回原突变(返祖性)在解释突变过程中的重要性
如果隐性基因起源于基因的损失,则隐性纯种中势必没有再度产生原有基因的希望,否则高度特化的某物竟能无中生有,这是说不通的。另一方面,如果突变起源于基因结构内的一种变化,则突变基因有时恢复原态,似乎是不难想象的。也许我们对于基因了解得太少,以致对这样一个论证,还不能给以很高的估价;但是后一种见解,对于返祖突变体的发生,似乎解释得比较合理。关于这方面的证据,不幸还不完全满意。的确,有几个果蝇的例子,在其突变型隐性原种里,出现了具有原来性状或野生性状的个体。但是除非在控制情况下,否则这类事件还不能被认为是证据充足,因为隐性原种中沾染一个野生型个体的机会,是不可以忽视的。只有在一个突变原种具有几种突变性状作为标记,其中仅有一种性状复原,而且当时附近又没有这些突变体的其他组合的情形下,这种回原变化才能提供注意的证据。在我们培养的原种里,有少数记录的例子,满足了上述条件,就证据所能涉及的范围来说,也表明了回原现象可以发生。也须提防另一种可能性。有一些突变原种,经过若干时间以后,似乎或多或少的失去了该原种的特征,但在杂交以后,却又完全恢复了那个突变性状。例如第四染色体上的弯翅性状(图30),原来变化不定,并且容易接受外界影响,如果不加选择,外貌上即有重返野生型的趋势。这种外貌上返祖的果蝇如果同野生型杂交,所得子代再进行自交,则在所预期的弯翅一类的孙代中,会有许多个体显出弯翅性状。在另一种称为楯片的突变原种里,也发现了同样的结果。楯片原种的特征是:胸部缺少某些刚毛。在某些楯片原种里,出现了一些具有这种“失去”了的刚毛的个体。表面上,这种突变体似乎回到野生型,但如果让这种果蝇同野生型原种交配,却证明并没有这回事。在杂交二代中,楯片果蝇重复出现。研究这个例子,证明楯片之所以还原为正常性状,是由于出现了一个隐性基因,该隐性基因在楯片原种的纯合状态下,促使所失去的刚毛再度发育。除了这个结果对正在讨论中的问题有关系以外,一个新隐性突变使原有突变性状回到原型,这件事本身也是一个饶有兴趣的重大事件。
最后,还有细眼还原为正常眼的奇异现象。细眼[图49a、b]为显性或半显性性状。若干年来,根据May和Zeleny二人的观察,已知细眼可以还原为正常眼,并且有人引用它作为回原突变的证据。回原突变的频率随不同的原种而异,估计每1600次中约发生1次。以后,Sturtevant和摩尔根发现,当细眼回原时,细眼基因的附近便发生交换。Sturtevant在判明所发生的变化的性质方面已经取得了决定性的证据。
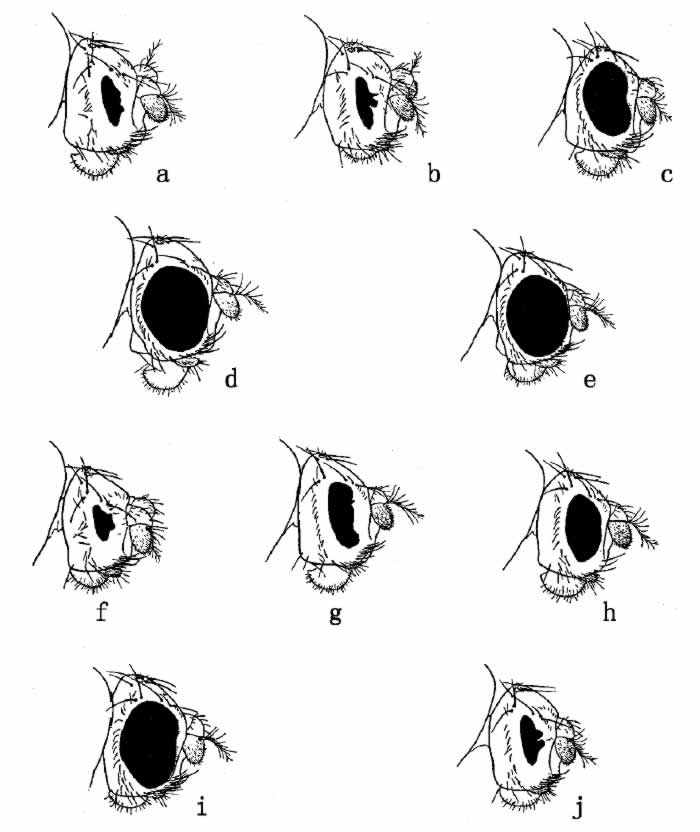
图49 果蝇细眼的各种类型:a.纯合的细眼雌蝇;b.细眼雄蝇;c.细眼对圆眼的雌蝇;
d.由回原作用得来的纯合圆眼雌蝇;e.由回原作用得来的雄蝇,含有一个圆眼基因;
f.双细眼雄蝇;g.纯合的次细雌蝇;h.次细眼的雄蝇;i.次细眼对圆眼的雌蝇;j.双次细眼的雌蝇
证实每次回原都有交换作用的方法如下:有一个称为叉毛的基因,紧贴在细眼基因的左侧(1/5单位);另一个称为合脉的基因,位于细眼右侧的附近(2.5单位)。一只雌蝇有下列组合:一条X染色体上含上述三个基因,细眼位于叉毛与合脉之间,另一条X染色体上除含细眼外还有叉毛、合脉两者的野生型等位基因(图50)。这只雌蝇同叉毛、细眼、合脉的雄蝇交配,其子代的普通雄蝇从母方接受一条含叉毛、细眼、合脉的X染色体,或者接受另一条含非叉毛、细眼、非合脉的X染色体,所以表现出的性状或者是叉毛、细眼、合脉三者,或者只是细眼。当回原突变发生时(很少发生),即当一只圆眼雌蝇出现时,可以看到在叉毛和合脉之间发生了交换。例如,回原的雄蝇或者具有合脉,或者表现叉毛,但从来没有兼具合脉、叉毛或者兼具非叉毛、非合脉的。所以在母体染色体上,叉毛和合脉之间一定发生过交换。总计叉毛与合脉间的交换,不到百分之三,但已经包括所有回原突变在内。
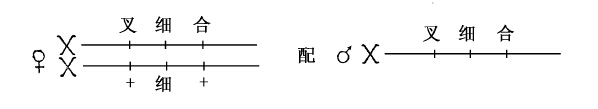
图50 一只细眼雌蝇在叉毛和合脉上为杂合子。该雌蝇与一只叉毛、细眼、合脉的雄蝇杂交[“+”代表野生型基因——译者注]
为了简化情节起见,上文仅提出回原型雄蝇。当然,回原型染色体也可以窜入一个卵子内,发育成为雌蝇。我们可以设计这样的一项实验,以便在回原型雌蝇体内探求交换作用的证据。子代普通雌蝇尽是纯合的细眼[图49a]。子代回原型雌蝇则为杂合的细眼,或者加上叉毛,或者加上合脉,没有一只是兼有叉毛和合脉的,也没有一只是兼有非叉毛和非合脉的。
引起还原为圆眼的这种交换作用,一定不只是使一条X染色体失去一个细眼基因,而且也必然会把这个基因移放在另一条细眼染色体之上[图51(a)]。含两个细眼基因(双细眼)的雄蝇,同含一个细眼基因的雄蝇,外形上相似,不过前者的眼比较小,小眼的数目也比较少。这型称为双细眼[图51(b)]。同一直线序列上有两个等位基因,这是在任何其他突变中从未见过的非常事件。我们可以这样去描画:即只有设想在交换前原来是对立的两个细眼基因,在进行交换时却稍微移动了一下位置,结果,双细染色体上至少延长了一个细眼基因,反之,另一条染色体上却随着一个细眼基因的缺失而相应缩短了。
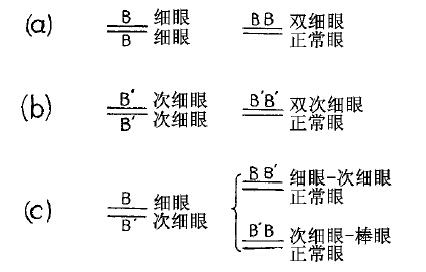
图51 示细眼、次细眼与细眼次细眼三者的突变[B代表细眼基因,B′代表次细眼基因——译者注]
Sturtevant对于回原理论进行过一些决定性测验。细眼的一个等位基因(由细眼突变而出),名为次细眼[图49g、h],在两眼的大小和小眼的数目上都同细眼型略有差异。次细眼原种内也发生回原现象[图51(b)],产生同野生型极相类似的完全圆眼型以及一个称为双次细眼的新型[图49j]。
一只雌蝇在一条染色体上有细眼基因,在另一条染色体上有次细基因[图51(c)]。当回原变化发生时,该雌蝇将产生完全圆眼型以及细眼次细眼型或次细眼细眼型[图51(c)]。
Sturtevant也利用了细眼次细眼型和次细眼细眼型,来证实下列事实:如图所示,如果突变基因都位于同一染色体之上,则当细眼次细眼与正常型之间发生交换时[图52(a)]结果会产生叉毛细眼型或次细眼合脉型;如果交换发生在次细眼细眼型与正常型之间[图52(b)]则结果或为叉毛次细眼,或为细眼合脉。

图52 a.示叉毛-细眼与次细眼-合脉之间的突变;b.示叉毛-次细眼与细眼-合脉之间的突变[f代表叉毛基因,fu代表合脉基因,B代表细眼基因,B′代表次细眼基因。——译者注]
由此可见,两型中的各个基因不仅保存各该基因的特性,而且也维持基因互相间的顺序。从fBB′fu和fB′Bfu的构成方式,可以知道这些基因之间的顺序。事实上,在所有的例子里,B与B′之间的断裂,都同原先决定了的顺序相符合。
这些结果,对于细眼由于交换而回原一说提供了有利的决定性证据。目前,这是独一无二的例子。似乎在X染色体上的细眼基因点上有某种特殊情况,能使等位基因之间发生交换。Sturtevant把这种交换称为不等交换(注:这些关系,涉及与细眼基因点有关的几个奇怪的问题。例如,细眼交换时,在细眼基因点上究竟留下了什么?是细眼基因缺失了吗?原来的细眼基因是由于野生型的基因突变而生,或者有另一个新基因的生成呢?这些问题仍在研究中。)。
上述结果提出另一问题:即一切突变是否都由交换所致?在果蝇里,显然证明了交换作用不能普遍解释一切突变的由来,因为我们熟悉,雌雄果蝇都可以发生突变,而雄蝇却没有交换作用。
