雄蟾的Bidder氏器官向卵巢的转化
雄蟾精巢的前部由像幼小卵细胞似的圆形细胞组成(图147)。甚至在精巢后部或精巢本身的生殖细胞还没有分化以前,幼蟾的精巢前部便已经够显明了。前部称为Bidder器,许多年来引起了动物学者的注意,他们对于Bidder器可能的用途提出了许多看法。最普遍的解释,是把Bidder器当为卵巢;Bidder器的细胞同卵子类似这一点,便是这项解释的有力证据。不过雌性幼蟾在其真正卵巢的前端也有Bidder器,这就不容易符合前一解释,否则雌蟾便会前端有退化卵巢或祖型退化卵巢,而后端又有作用的卵巢了。
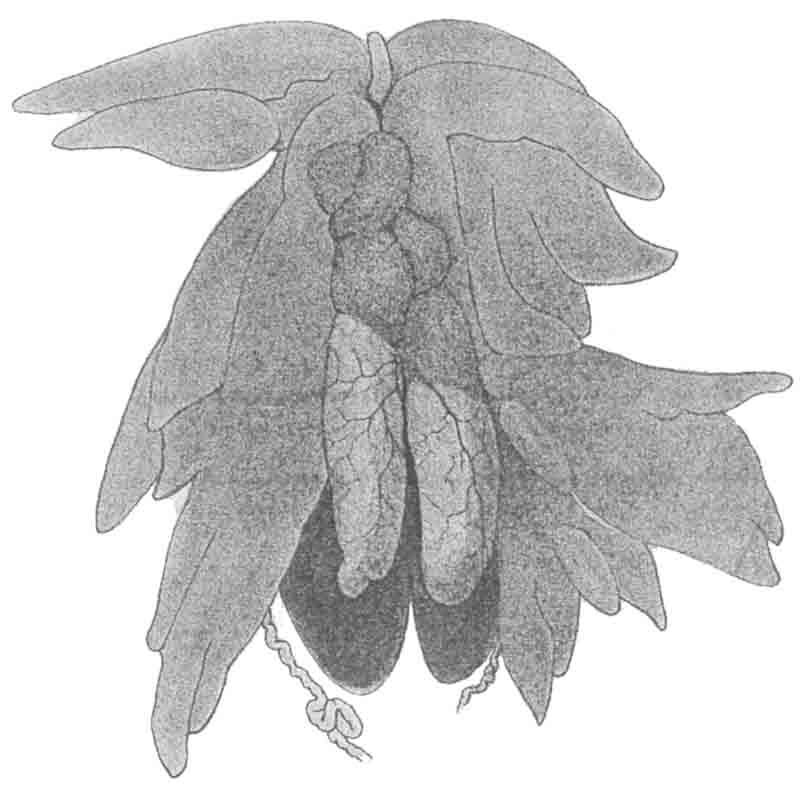
图147 半成年的加利福尼亚种雄蟾,精巢的前端有Bidder器,两侧有脂肪体的各叶,下面有肾脏。壁上有分支血管的,为精巢
Guyénot和Ponse(1923)以及Harms(1923,1926)的实验先后证明:在幼蟾的精巢被完全摘除以后的二三年内,Bidder器发育成为卵巢,并产生卵子(图148)。卵子由母体排出,经过受精以后的发育,也被观察到了。这里,毫无疑问,在摘除精巢后产生了雌性;至于接受手术的个体,究竟应该称为雄性或者称为雌雄同体,也许是一个定义的问题。我个人会把它称为雄性,并且认为上述结果意味着:由于摘除精巢,雄性才转化成了雌性。我认为雄蟾器官里的细胞可能发育成卵子这一点,还是次要的问题,因为一般说来,即使性别决定于染色体机制,这也并不意味着,预定生殖腺所在地区的未分化的细胞,由于具备了在另一种情况下产生雄性的那种染色体群。便不能在不同的环境下变成卵细胞。从基因方面看,这就是说,蟾蜍有这样的一种基因平衡,在正常发育条件下,一部分生殖腺(前端)开始发育成卵巢,而另一部分(后端)开始发育成精巢;随着发育的进展,卵巢的发育赶上了精巢的发育,并且制约着精巢进一步的发育。但是如果摘除精巢,这种制约也就消失了,于是Bidder器的细胞便重新发育,成为有作用的卵子。Ponse从转化了的雄蟾的卵子,得出9雄和3雌。Harms从同一只雄蟾育成了子代的104个雄性和57个雌性。假定雄蟾为XY,预料转化后的雄蟾会有X和Y两类卵子,各占半数。这些卵子如果同正常雄蟾的精子受精,预计子代中,将为1XX+2XY+1YY。YY型个体多数不能发育,从而造成了2雄和1雌的比例,同实际结果密切符合。
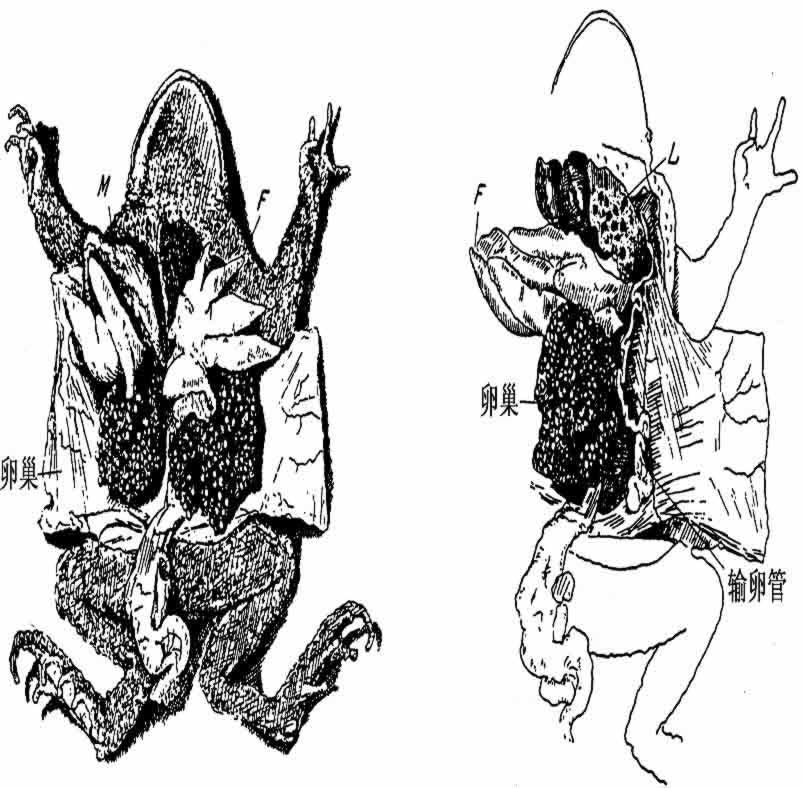
图148 摘除精巢三年后的蟾蜍,Bidder器发育成了卵巢。在右边图里,为了显出胀大的输卵管,于是把卵巢翻到一侧去了(仿Harms)
Champy描述过蝾螈Triton alpestris中“完全性转化”的一个例子。一只雄螈原呈雄性作用,有生殖力,以后任其断食。在这些情形下,精子不能进行正常的更新,不过蝾螈依然保持某种“中性状态”,它的特点,是精巢里面有了原生殖细胞。在这种状态下渡过了整个冬天。再取两只断食的雄螈,加强营养,以后便发生了从雄性色彩到雌性色彩的变化。几个月以后,检查了其中的一只,Champy认为检查结果可以证明性的转化。鉴于这个例子近来会被用为性转化的充分的证据,所以值得比较详细地谈一谈Champy的实在记载。他看到的,不是卵巢,而是一个大致和幼小卵巢相似的长形器官。在它的切片上看到了器官内有幼小的卵形细胞(“卵母细胞”)同变态期内幼螈的细胞相似。又有一条输卵管,白色而曲折,所以显然可辨。Champy由此断言:这里是一个具备幼小雌螈卵巢的成年动物。这项证据似乎指明,断食处理导致了精母细胞和精子的吸收;这却不能明确地指明以后发生的新细胞,究竟是胀大的精原细胞,还是原生殖细胞,或者是幼小的卵子。根据两栖类的其他证据(Witschi、Harms、Ponse),下面的结论也并不是不可能的:就是,这些细胞实在是幼小的卵细胞,并且也发生了不完全的性转化。
