第十六章 大脑两半球中刺激的分化过程
(根据B.B.贝利亚可夫博士的实验,在俄罗斯医师学会宣读,发表在《俄罗斯医师学会会报》,79卷,12—26页,1911—1912年)
中枢神经系统的活动机理是基于暂时联系和分析器——分析器与分析——分化作用基于抑制作用——表明抑制解除作用和分化作用的实验——分化作用中抑制作用的所在部位——这些实验的生理和心理解释比较起来,后者是不适当的——分析器的生理;中枢的定位
动物高级神经活动的客观研究,条件反射的学说获得了关于中枢神经系统的两个主要机理的概念,即,暂时联系 的机理和分析器 [1] 的机理。这篇报告是关于分析器机理的生理功用和它的活动。
我提醒大家,关于分析器我们的意思是指一种神经器官,它包括下列部分:某一个周围末端(眼、耳等,通常称为“感觉器官”),与它相关的神经,及最后,此神经的大脑末端,就是,这个神经终止处的一组细胞。我们要讨论的是关于这个神经的最高末端,它位于大脑两半球内。这个器官被称做分析器是很恰当的,因为它的机能就是去分解复杂的外在世界成为它可能最小的要素。它的活动可以分做几部分来研究:一方面规定分析的界限,另一方面去研究分析的机理。今天我们讨论这个问题的第二部分,就是,讨论它的机理。
为了向诸位解释我们如何表明这分析器的机理,我将要举出一个详细的实验。我们拿作用于这个或那个分析器的一些外在世界的动因,声音、气味、皮肤的机械刺激,等等,试把它和某一种生理活动发生暂时联系。在我们的实验中是经常与唾液腺的活动相联系的。把某一种动因与这个器官的正常生理刺激相结合,我们便可以把此动因引到我们所愿望的联系中去。
经过一些重复之后,我们得到所希望的结果:从前对这个器官没有效用的动因,现在与它的活动发生关系,并且很快地就成为它的刺激了。每当它起作用时,它就引起该器官的活动,在我们的情形,就是引起唾液的分泌。假如于这种联系已形成时,我们试用其他刺激于身体同一感受表面,它们也会起作用,虽然在以前这些刺激绝未曾与这个器官的活动发生过联系。例如,假如我把一定的乐音与唾液腺的活动联系起来,然后再试用别种乐音或声音,它们也同样是有效的。但是这只是一个阶段,是一个一定的时期。我们注意到,如果把所选择的动因重复多次,我们的刺激,在起初虽似乎具有一种普遍的特性,渐渐就变成为特殊的了。以前,各种各样的乐音和噪音都有效力,后来这些中有许多变为无效,到最后,能起刺激作用的声音越来越少,就只剩下与我们选择的乐音有临近关系的那些乐音了。我们相信,这种从一个分散的兴奋转变到一种特殊的限度狭小的兴奋,这种分化作用的发生,是因为在神经系统中的某一点上发展了一种抑制性的过程。
我们这种确信的根据是什么呢?它是根据一些如下经常重见的事实:
例如,我选择一个1000振次的乐音;这个乐音已经成为唾液腺的一种刺激,经过许多次重复之后,我达到了可以使1000次的振动发生刺激,而与它相近的1012振次的乐音却不能发生刺激的情况。因此,刺激声音的范围缩小了,一个仅仅相差12振次的乐音,即是,与原来乐音仅差八分之一音的乐音,便不能发生刺激。如我所说过的,刺激的这种分化作用是通过一种抑制过程的发展而形成的,其证明如下:我用1000振动次数的乐音,它引起唾液流出;然后再应用1012振次的乐音,就不流出唾液;这两个乐音间的完全分化作用便发生了。如果在1012振次的乐音之后,我立刻应用原来的乐音是没有效力的,即使有也是很弱的,必须等候几分钟才会再发生。
这个事实仅可以作如下的解释:当应用分化了的乐音时,在神经系统中就产生一种抑制过程,假如在这个时期我再应用原来有效的那个乐音,它的作用就被抑制过程所压抑。必须经过一些时间以后,这个抑制过程在神经系统中才会消逝,原来的乐音才会有效,这里有抑制的发生是毫无疑义的。
于是,分化的过程,分析刺激的过程必须这样来说明:假如把我们所选择的特殊动因最初与某种生理机能相联系,那么,这个动因所产生的刺激,当达到皮层中某一点的时候,就向着与它相关的感受中枢扩散或散布;这样,不但某个分析器脑末端的一点进入了一定的联系,就是整个的分析器,或是它的大部分或小部分,都会发生一定的联系。只有在后来,由于抑制过程的对抗,刺激影响的区域才变小,直到最后获得一种孤立的作用为止。这便是前面的实验所解释的重要事实。
显然,这仅仅是问题的开端,还有许多疑问发生。其中一些问题已被贝利亚可夫的实验所解决。我将要报告他的记录。第一个实验如下:如果分化作用基于抑制的说法是正确的话,那么我们应当可能用损毁抑制的方法去随时损毁分化。这是为什么呢?因为在复杂神经活动的研究中,我们经常碰到抑制解除 [2] 的过程。如果分化作用的确是基于先前有效的所有邻近刺激的抑制,那么,应当有可能从抑制下去释放它们,而使它们再度发挥效力。我们可以在附列的记录中表示出这一个事实。
第一表 狗“稻刚亚衣”,1911年5月9日 [3]
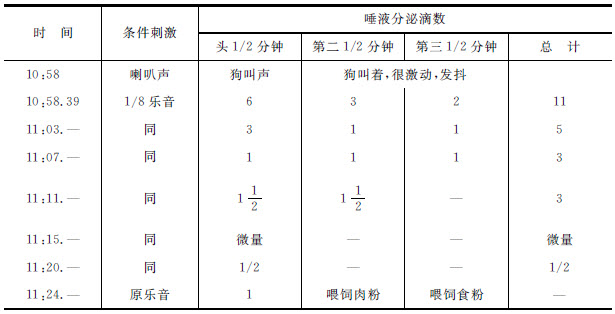
这是我们用一只名为“稻刚亚衣”的狗的实验的结果。经过9个月的训练,我们建立了并经常练习了一个与原乐音差别1/8音的乐音的准确分化。即是,分化了的乐音没有引起分泌的效力,而原来的乐音很迅速地可以刺激唾液的流出。现在我们用另外一种乐器的声音来刺激狗——一个包含许多陪音的尖锐喇叭声。它在动物身上产生了很显著的效力,狗开始吠叫,在架子上挣扎,并且发抖。当喇叭声停止,它安静下来的时候,我们试用分化了的(阴性的)1/8乐音,这时连分化作用的痕迹也没有了。起初,在30秒钟之内得到了6滴,与原来的乐音(1,000振动次数)所得到的完全一样,在以后相继的两个半分钟期间之内各得到了3滴和2滴,总计11滴。在5分钟之后我们又重复应用同一乐音,它发生了作用,在一分钟之内得到了4滴。而再过4分钟之后它的作用还没有完全停止。假如我们查看表中最后一栏,可以看到由于多次重复分化乐音所得的唾液分泌的总量是很大的。被分化了的乐音好像是起着通常刺激的作用;因为被重复时,它的效力就渐渐消灭了。这个抑制解除作用维持了10到15分钟,而没有分化作用的痕迹。我们有许多类似的实验。这里我们举出一个,它明显表示出被分化了的乐音就如同一个很熟练了的老的条件反射一样地被消灭。
再有,如果抑制是分化作用过程的基础,那么我们应当可以加强、累积并综合这个抑制。怎样做呢?连续重复被分化了的刺激多次。第二表描写这样的一个实验。
第二表 狗“克拉萨维茲”,1911年6月11日 [4]
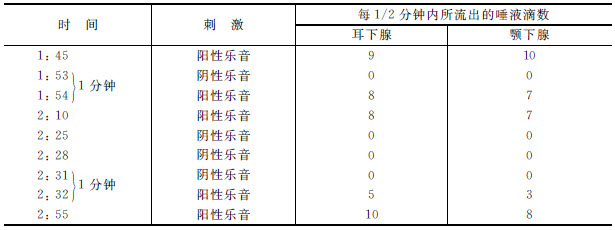
对于“克拉萨维茲”,某一个乐音已经被用作一个条件刺激。第二表中的第一行的数目字表示条件反射的平常数量——耳下腺得9滴,颚下腺得10滴。现在我们试用大约低1/2音的被分化了的乐音。它没有作用。我们应用它一次,一分钟之后再重复原来的(阳性 )乐音。我们看到,这里若是有抑制的话,也是很小的——我们得不到9滴而是8滴和7滴。现在我们把这个同一被分化了(阴性 ) [5] 的乐音连续重复三次,就是说我们把抑制作用累积起来,我们看到,在试用被分化了的(阴性)乐音的同样间隔之后,再应用平常的(阳性)乐音(像从前一样),这个平常的乐音的效果就锐减,不给出8滴和7滴,而给出5滴和3滴。如果我们给予一段时间使这个抑制分散开,然后再试用阳性的乐音,我们便看到它会恢复它的通常效力——10滴和8滴。由此我们可以说,抑制是分化的基础,它可以由重复分化了的(阴性)刺激而被综合起来。
以下是另一个事实。假若抑制是分化的基础,那么,分化作用的任务越艰巨,分化作用越精致,抑制作用也就越大。很明显,分别两个相差仅1/8音的乐音,是要比分别两个相差一全音的乐音难些。我们可以设想抑制作用的强度也是会不同的。分化作用越是精细,抑制作用也就越强,反过来也是如此。下面是一个实验。
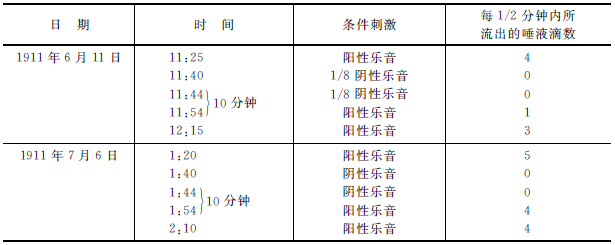
在常态条件之下,我们从狗“稻刚亚衣”用阳性乐音(1,000振动次数)得到4滴唾液。然后我们试用阴性乐音(1,012),它不产生唾液。连续试验了两次,它都给出0滴。10分钟以后我们再试用阳性乐音,于是它被抑制住了。被分化了的乐音的抑制作用持续了一些时候,表现在阳性乐音的效力的减低。让我们把这个实验与第二个实验(见第三表下半,1911年7月6日的实验)相比较。我们在第一行中看到条件反射的正常数量——5滴。然后我们试用一个分化了的乐音,但是这个乐音是很容易区别的,就是说,是一个比阳性乐音相差两个全音符的乐音。这个阴性乐音被重复两次。10分钟以后,我们试用阳性乐音。它丝毫没有变动;它给4滴到5滴。如此我们看到一个精细的分化作用(区别出相差1/8音的阴性乐音),造成一个强烈的抑制作用,然而一个粗糙的分化作用(区别相差两个音符的阴性乐音),并没有引起任何显著的抑制效力。
第三表 狗“稻刚亚衣” [6]
这里发生了一个有趣的问题:这个作为分化作用基础的抑制是在什么地方发生的呢?很自然地我们会想到它是在相应的分析器中发展出来的,就是说,在刺激作用被分析的那个地方。但是这自然需要证明。现在我将要举出一个实验,可以引致抑制作用正是在抑制性刺激所隶属的那个分析器中发生的这个结论。我们试用了来自不同分析器的各种刺激,去试行把这个分化作用解除抑制,得到在第四表所表示的结果。
第四表 狗“克拉萨维兹” [7]
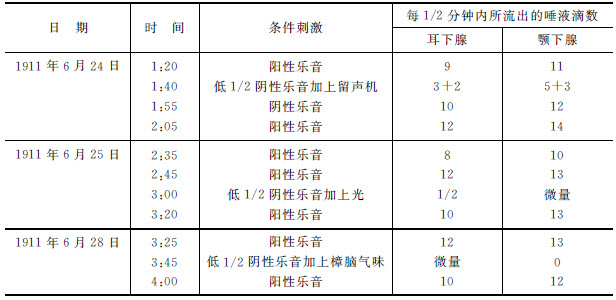
表中第一横行表示对于阳性乐音的平时正常分泌量,9滴和11滴。然后,与阴性乐音的同时,再应用可以产生动物方向反应的一个新刺激,即留声机的音乐声。它产生了很大的抑制解除。被分化了的乐音和留声机声共同产生的不是0,而从耳下腺给出3滴和2滴(每15秒钟),从颚下腺给出5滴和3滴。因此,留声机声解除阴性乐音的抑制。在下一个实验中(第四表中部,1911年6月25日),我们应用一个光的刺激作为解除抑制者;它几乎没有效果。分化作用仍旧保留着。光的刺激并没有发生解除抑制的效用——它并没有破坏分化作用。最后,在第三个实验中(第四表下部,6月28日),我们施用了樟脑的气味作为解除抑制者。这也没有效力。因此,我们用三种不同的刺激:用光亮、留声机和樟脑分别刺激眼、耳和鼻分析器。我们的分化了的(阴性)乐音影响耳分析器,而留声机也影响同一分析器,于是呈现一个强烈的抑制解除;但是落在眼和鼻分析器上的刺激是没有效力的。虽说光亮只是一个微弱的刺激,但是正如大家所看到的,气味并没有释放我们的阴性乐音的抑制。
我们还有其他的实验可以直接证明抑制是在分化了的刺激的分析器中发生的。第五表中列举出这样的一个实验。
第五表 狗“稻刚亚衣” [8]
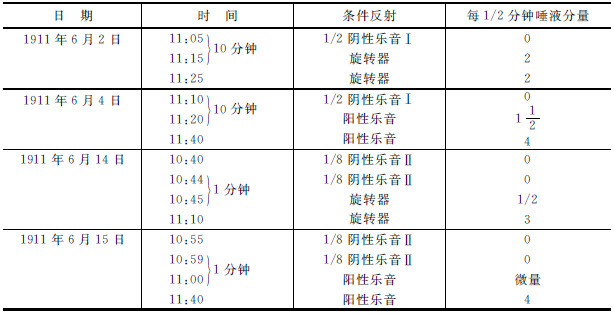
在这里,我们比较两个条件反射,一个是对于一个乐音,另一个是对于一个旋转物体(旋转器)所起的条件反射。应用阴性乐音之后,在中枢神经系统中就保留有抑制性痕迹,这个抑制性痕迹的作用,可以用测量对于一个阳性刺激,一个乐音(是与阴性刺激同一个分析器,即是耳分析器)的反射的大小,与测量对于另外一个分析器(眼分析器)的阳性刺激(旋转器)的反射的大小来比较。在起初,试了一种粗糙的分化作用(半音)。“稻刚亚衣”对于这个分化作用产生了微弱的抑制作用;因为在它的身上,我们已经建立了一种更精细的分化作用,它能分辨出相差1/8音的乐音。我们用这种分化作用所产生的抑制性痕迹的效力去试验眼分析器的反射,即对于旋转器的反射。这个反射并不因应用阴性乐音而被抑制,它给出与当天前一次试验中所得的同样数量的滴数,就是2滴。所以,在当时的(时间的)条件之下,耳分析器中的微弱分化作用(就是,一个不重要的抑制过程)对于另一个分析器(眼)的兴奋过程是没有效用的。同样的分化作用(1/2音),就是说,有同样强度并在同样条件之下的抑制性过程,对于同一分析器(耳)的一个条件反射,给以显著的抑制性效果。在当天,对于阳性乐音所起的唾液分泌是4滴。试验粗糙的分化作用(1/2音)所产生的效果是0。10分钟以后,应用阳性乐音,不得4滴,而产生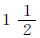 滴的唾液分泌。因此,可以证明出,同一分化作用,同一分析器的抑制性过程,对于同一分析器的反射具有一种抑制性效果;但是对于其他分析器的反射,便不具有抑制的作用。所以,由于分化作用所引起的抑制,其位置是在被分化了的(阴性)刺激所作用的同一分析器中 [9] 。
滴的唾液分泌。因此,可以证明出,同一分化作用,同一分析器的抑制性过程,对于同一分析器的反射具有一种抑制性效果;但是对于其他分析器的反射,便不具有抑制的作用。所以,由于分化作用所引起的抑制,其位置是在被分化了的(阴性)刺激所作用的同一分析器中 [9] 。
由已往的报告大家还能记得,中枢神经系统的最高部分的神经过程是经常在流动着,扩散着和集中着。这个理由使我们相信,来自某一分析器的抑制过程可以散布到整个两半球中。为了证明这一点,我们不能再用简单的分化作用(它只要求一种微弱的抑制过程),而必须应用一个较高级的分化作用,或累积分化了的抑制作用;因为抑制性波浪不仅限于所涉及的分析器,而且包括了邻近的和遥远的分析器。
在同一只狗“稻刚亚衣”的身上,我们现在试用一种较高的分化作用,用相差1/8的乐音并重复它。你们可以清楚地看到,它的作用并不只限于同一分析器,而散布到另外的分析器。第五表(6月14日)有一个实验表示耳分析器中所引起的分化了的抑制作用对于眼分析器反射的影响——对于旋转器所发生的反射的影响。在施用阴性乐音之后旋转器只产生了半滴;但是若等候25分钟以后,使抑制作用的波浪能有充分时间去消失,那么它就显出十足的效果——不是半滴而是3滴。显然,当两种反射是在同一分析器中时,即是,在耳分析器中时,也发生同样的情形;因此,假若反复施用(阴性的)1/8的乐音之后,再试用阳性乐音,后者便没有效果(见第五表,1911年6月15日)。假若抑制过程在遥远的分析器中达到某种力量,很明显,它应当在原先产生它的分析器中有更大的效力。
这就是我们与贝利亚可夫博士所建立的一些事实。显然我们由这些事实去深入,就是说,我们可以提出关于这个机理的很深入而奥妙的问题,并且可能得到确定的答案。我们不仅能够建立分化了的抑制作用的事实,而且实际上我们能够用实验方法把它引导到某些路线中去,加强或减弱它,并且找出它是从什么地方发源的,等等。
我们在回顾这些结果中,很有兴趣地对于我们的客观观点提出一个比较性的判断,这个客观观点是不难于维持的。你们看到我并不只是在想象着,而是经常站在事实的基础上。我用实验来检验我所有的命题,因此,我的观念总是靠事实来决定的。为了要正视这个客观的生理学观点的力量,我希望诸位先生们试从心理学观点来理解并解释上述的事实。你们会看出很显著的区别。让我们看一两个例子。我把一定的乐音做成一个条件刺激。我们可以想象并且说这狗很熟记这个声音就是进食的信号,在它之后必定有食品吃,为了这个期望,狗便分泌唾液。现在在这个乐音之后,我试用一个仅相差1/8音的乐音,狗不能立即分辨它们,而将它们弄混,结果就分泌唾液。它记忆得不好。此后我把平常(阳性)的乐音和非平常(阴性)的乐音重复多次,使狗熟记阳性乐音是伴随进食,而阴性乐音是不伴随进食的。当我应用阳性乐音的时候,狗分泌唾液,并且预备吃食,但是当应用阴性乐音的时候,它安静而不期待食物。现在在阴性乐音之后立刻施用阳性乐音,它没有效果。这是为什么呢?狗是很熟知这些乐音的;它能记得哪一个乐音是进食的信号,哪一个不是进食的信号。为什么它现在听见阳性的乐音反而不分泌唾液呢?这将如何解释呢?还有一层,我第二次又重复阴性乐音,仍无唾液。这是说狗记住了,这个乐音是不会随以进食的。我第三次重复阴性乐音,仍旧得到同样的结果,这证明狗已经记得很准确。但是为什么它单单忘记了阳性的乐音呢?从心理学的观点看来这是不可能被了解的。如实验中所示,在阴性乐音15分钟以后,它又能回忆起阳性乐音,这更是不可能理解的。但是若从我们的生理学观点看来,问题是很简单的。如果分化作用是一种抑制作用,又假定分化作用的重复是一种综合,是抑制作用的累积,那么,我们必须等候一段时间让抑制作用消失,然后常态关系才可以恢复。
去检查所有的心理学概念,并且去与我们的客观材料相比较,指出它们是粗糙的、经验的和空想的,并且在分析高级神经活动的最微细现象中,这些心理概念的特质是不可克服的阻碍——这种高级神经活动的分析就是摆在我们面前的任务。
现在我再回到分析器的问题。我们已经收集了关于分析器的活动的事实并将它们加以系统化。再者,我们还有材料说明在某些条件之下它们的活动如何变化。根据上述事实看来,如果我们破坏大脑两半球代表分析器组合的某些部分,这个破坏所表现的方式应当是我们所预料到的。如果我们把一个分析器的一部分损毁,这便立刻在它的机能上反映出来。机能损伤的程度是被大脑损坏的大小及从施手术到观察时所经过时间的长短所决定的。正如我们所知道的,这些扰乱在这种程度上是可以逐渐地被补偿,但是永不会完全平复的。
还有,在分析器机能的扰乱中,必须解释哪些扰乱是由于损伤,及哪些扰乱是由于其部分的割除。的确这是一个远大的问题,而且我不知道到何时才可以解决,但是我应当说明在我们已经做过的实验中,能有答案的某些线索。例如,我们有证据说明分化作用中的扰乱是依赖于抑制过程的正常进程的某种歪曲和中断。
所以诸位可以看到,神经系统的最高级活动,大脑两半球的活动的分析机能,可以加以严格的生理学的探讨,而绝对不需要心理学概念的帮助。而且这个分析的机能就是大脑两半球的主要任务。
关于事实和一些片断知识的这个报告,虽然很贫乏,我想可能给予我们一些指示,以解决关于分析器的生理活动的深奥秘密。在我们面前使我们非常迷乱的现象之一,便是大脑两半球相当大部分被割除后,过了一些时候,在神经系统的活动中不一定常能发现任何缺陷。看来好像你是在研讨一个珍贵而又特别重要的机理,但是在另一方面,你破坏并损毁它的一大部分,所以你看不出有任何后果。我愿意着重地指出,大脑惊人的补偿能力是很值得注意的。因此你看到,差不多100年以前关于大脑两半球整体的说法,在后来虽然曾遭到否定,但是现在从它的个别部分的关系看来,这个说法是一个活现的事实。大脑生理学是从法国学派的观察开始的,此学派曾有条理地指明,在两半球中并没有定位,并且两半球受损坏时,如果只有一部分还保留着,状况便会复原于常态。
到1870年,当福利契和赫齐葛做了他们有名的实验,这些看法便被完全放弃了。他们的实验奠定了中枢定位学说的基础。在当时早先的看法好像是一个极大错误;然而现在当我们把分析器进行了详细的研究后,这个被拋弃了的观念又重获光明了。当割除了大脑两半球的大部分时,起初看来好像分析器是被消灭了;它的作用几乎毫无表现。但是经过几个星期或几个月之后,这些损伤已经被补偿得使我们很难看出这个动物究竟在哪一方面是不正常。
大脑半球的大部分存在有定位是毫无疑义的事实。但是脑的个别区域中的定位是怎样的——这是生理学所面临的困难而艰巨的工作。一个结构可以被破坏和毁灭,而我们看不出这个损伤的结果,这怎样解释呢?显然,在个别分析器中我们必须承认一个无疑的事实,就是机能中存在有某种代替作用和补偿作用。这将如何被了解,我们采取什么推测呢?我们的概念当然一定是机械论的。
关于这个问题,显然已经有了一些希望和一些进展。我今天在开始时所叙述的话很可能是具有重要意义的。我们指出的事实是当条件反射刚刚被形成,它便转为普遍化了。由此看来,很显明分析器的脑末端代表着一种共同的物质基础,其中的各部分都紧密地联系着,并且其一部分可以代替另一部分。我们可以假设,在分析器的周围这一端,对于刺激因素有一种严格的分化作用,每一部分都是个别的并且与其他部分彼此分别开,同时在分析器的脑末端,有联系可以通达大脑全部,以至于所有的周围刺激作用可以传达到脑髓的每一点。因此,有可能性以分析器的小部分来代替分析器的大部分。
我所说的话不够作为一个建议,而是这些极端复杂而重要的问题,怎样才可以得到解决的一种预感。最后,我愿意表示出我的想法,我们距离大脑两半球机理的任何真正概念还差得很远呢!
[1] “分析器”的定义(巴甫洛夫用“分析器”这个名词来代表一个机能单位,包括从外在世界接受刺激作用的身体表皮(感官),传送冲动到中枢神经系统的神经,以及这个过程所流到的中枢神经系统中的细胞。虽然对外在世界的现象作物理分析的最重要的部分是属于分析器的周围结构,但是具有最大生理兴趣的,还是在于分析器中央部分的细胞,特别是脑的最高级部分。这部分是被假设为与此过程有密切关系的;在这里条件反射所依赖的联系,暂时造成而又破坏。关于分析器作为一个生理单位的描述,将在下一章第二部分及第二十一章第一部分中提及。比较新近的研究指出分析器这个词的定义,并不包括这个生理器官的所有机能;因为它不但把外在世界分解成它的单纯而细致的现象,并且它也具有把许多单纯现象联合成为一个复合刺激的能力,即是,它做分析,也做综合。———英译者)。——英译者
[2] 读者会记得在在这个研究阶段,巴甫洛夫已经感到,在这种关于两半球的精细工作上,我们必须考虑到每个不同神经系统的个别特性。在第三十五与三十八章中,我们会找到关于这一点的更多事实。这个问题已在巴甫洛夫的《大脑两半球机能讲义》第十七章中系统地讨论到。在特殊实验中,可以证明属实,某种刺激若是与两半球某点的抑制状态,常常同时发生,那么,这种刺激再次应用时,就能够产生这种抑制作用状态。条件反射的消灭,被认为是由一种抑制过程而产生的。这种抑制过程的产生,是由于附加于消灭了的条件反射上的每一个非常的或新的动因的阻碍;所以,由于缺乏抑制,条件反射又成为有效的了。这种过程被视如停车器(抑制作用)的拉起,称为“抑制解除”。在描写第一种大脑机理的第十章中和在第十章中有抑制解除者的实例。———英译者和第十一章中关于抑制解除 所谈到的。所有的内抑制的过程可以用额外刺激来释放抑制,分化作用的过程既然是基于内抑制,则分化了的(阴性)刺激也能被释放抑制,就是说,遇到额外刺激时它们就会表现出一种兴奋的作用,而不是那种平时的抑制的作用。——英译者
[3] 喇叭是当做一种非平常而强烈的刺激(额外刺激)应用的;1/8乐音的意思是比用做条件刺激的原来的乐音高1/8音。1/8乐音是阴性的或分化了的乐音。
[4] 阳性乐音=伴以喂饲的乐音。阴性乐音=未伴以喂饲的乐音。
[5] 把这两个乐音叫做阳性和阴性似乎是很便利的,阳性的意思是伴以喂饲而又可使唾液流出的刺激,阴性的意思是较后试用的刺激,并不伴以喂饲,而且通常也不使唾液流出(被分化了的乐音)。——英译者
[6] 阴性乐音与阳性乐音相差两度。
[7] 阴性乐音比阳性乐音低1/2音。
[8] 阴性乐音=不伴以喂饲的乐音。阴性乐音Ⅰ=与阳性乐音相差1/2音。阴性乐音Ⅱ=与阳性乐音相差1/8音。
[9] 就是说,假若对声音的分化作用已经被建立了,那么阴性声音所形成的抑制作用并不影响所有的阳性条件刺激,而主要地影响那些声音分析器的条件刺激;假若来自跟分析器的刺激已经建立了分化作用,那么,阴性光觉刺激所形成的抑制作用主要地影响跟分析器的阳性条件刺激,而不影响,例如,声音刺激。
