第二十二讲
我们研究的一般性特色:研究的任务和困难——我们的错误
诸位!在科学地研究动物现象的场合,可以在若干水平之上做这种实验。可以考虑生命现象必有的物理化学的基础而利用物理的和化学的方法,以分析基本的生命现象。其次,重视着活物质进化的事实,我们可以努力把活物质复杂构造的机能归纳于活物质基本各形态的特性。最后地在全部现实的范围上掌握着复杂构造的活动,我们可以探求这类活动的严密的规律,或者是这样的,就是一个活动的过程,在该活动的一切瞬间和一切变动之中都是由条件而正确决定的,所以需要确定这些一切的条件。这最后的水平,显然也就是我们上述研究所采取的水平。在这水平上引起我们注意的不是如下的问题:究竟兴奋与制止两个过程在最深的根本上是什么东西?我们把这两个过程当做我们复杂构造物的两个事实性的资料看待,当做两个基本的特性看待,当做这复杂构造的活动的两个最主要的表现看待。我们并不是如神经纤维研究所确定地,把大脑两半球活动归纳于神经组织基本性特色的办法当做自己的任务,我们甚至于不详细谈及这神经活动两个根本现象——兴奋与制止——在组织构造成分上可能的分配情形,即是不谈及兴奋与制止这两个根本现象在神经细胞与结合点之间的分配情形或在神经组织间的道路上的分配情形,而是以一个暂时的假定为满足的,就是假定这两个过程都是神经细胞的机能。我们条件反射的研究也就是大脑两半球皮质细胞活动的研究,这当然是毫无疑问的。绝对地证明这个主张的是,大脑两半球从外界和内界所接受的无数个别刺激,与此相当地,是由数亿乃至数十亿的皮质细胞所构成的各孤立点而表现的,而大脑两半球一被摘除,则所有的刺激就消失。还有一个似乎很可能的事情,就是兴奋与制止两个过程,经常不断地密切地彼此互相综错交叉,始终不断地彼此互相交替,这就是神经细胞的机能,不过是物理化学过程的不同的时相而已,而皮质细胞内这些物理化学过程的发生,是由于外界和内界进入皮质内的、及由于全身体各部位乃至特殊地由大脑两半球其他各点的无数刺激影响而引起的。我们基本的任务是大脑皮质活动个别表现的记录和特征描写、这些表现发生时的正确条件的确定和这些表现的系统化,换句话说,就是要决定兴奋过程与制止过程数量动摇的条件和这两过程彼此间的关系。这样看来,从研究的特征而言,我们的研究是与谢灵顿及其学派有关脊髓的研究很相类似的。不能不认识,我们在研究大脑两半球时所获得的许多事实,在许多点上,是与脊髓生理学的事实相一致的,这就证明两方面在基本关系上是自然一致的。
然而我们即在自己所预定的界限中做大脑两半球机能的研究,这研究依然是困难无比的。因为大脑两半球皮质的非常的反应性和不断地向大脑皮质接触的无数刺激,有两个基本的特征成为大脑皮质的特性,一个是异于寻常的条件制约性(обусловленность),一个是与这条件制约性自然有关的、构成这皮质活动的、各现象的流动性(текучестьявлений)。几乎对于任何现象,我们都绝对不能确信自己有掌握该现象存在有关的一切条件的可能。外在环境的与内界极小的、往往几乎不能捉摸的、或者完全不能想象的动摇,能使现象的进程(ходявлений)强烈地变化。显然,在这样情形之下,特别痛苦地表现的是两个思想上的通常弱点:一个是常同性(стереотипность),另一个是偏见(предвзятость)。可以说,思想不能赶上各式各样的关系。所以在做这类研究的时候,往往不能不犯错误。我确信,在已经说明的资料内也有不少的缺点,甚至有些很大的缺点。然而,既然想领会这样的复杂性,也不必耻于错误。这就是为什么我很久地迟疑着不曾对我们如此长时期的研究做一个系统性说明的缘故。因为这研究对象的上述特性,新问题不断地发生着,而后方还残存着许多未解决的问题。往往有许多意外的事情,使我们不能照预定的任务进行,并且从我们已接受的观点往往还有许多不能解释的事情,使我们迷惑。正是在这结论性的一篇里,我想叙述大脑两半球研究的一般性特色,并且根据在以前各讲里为地位所限制而不曾被记载过的事实材料。
我想,我们最近研究内的下方两个观察资料可以特别鲜明地昭示,我们研究范围内的各现象是复杂到什么程度,这些现象怎样精微地、多种多样地受着条件的限制。
在以前的各讲义内不止一次提及过的一只狗,受了非常的洪水的强烈影响。它在受了我们一定的处理手续以后显出了声音性分析器孤立性机能性障碍。在解决若干任务的时候,即是在利用音的高度形成分化相的时候,这只狗在整整一个月之中保持着完全正常的态度。在孤立的条件刺激时间不过是10秒钟的场合,唾液分泌性效果是5滴。强刺激物与弱刺激物间的条件反射的效果也有显然的区别。在条件刺激物以后被给予的食物,这只狗就即刻抓住它而贪食。在实验架台上,它泰然地站着。现在这实验有了似乎很小的变动。孤立的条件刺激时间更加长了5秒,于是所有的条件反射性活动即刻都强烈地有了障碍。
这实验的实在数字如下。

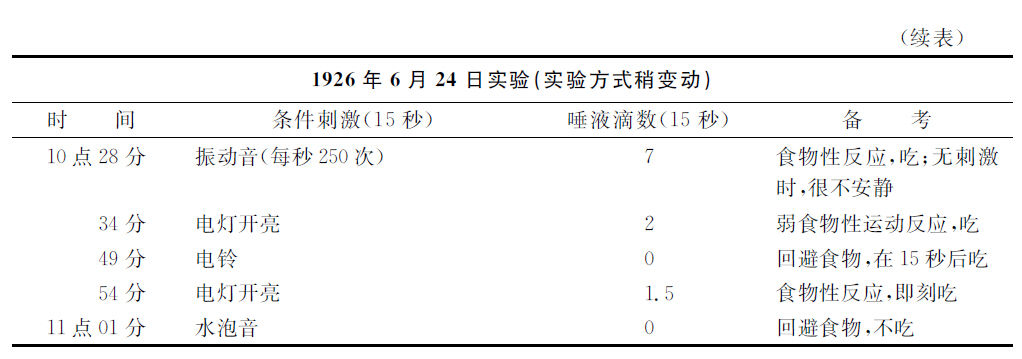
第二个实验里的第一个刺激物显出了比平常更强的效力,这是容易理解的,因为孤立的条件刺激时间加长了5秒。对于条件刺激物的食物反应是活泼的,食物即刻被吃完了。这是通常实验所应有的事情。并不曾显出这个实验的任何特别异常的征兆。然而在实验中的休息时间内,动物某些惶恐不安的状态已经引起注意。以后,显然的反常时相出现了。对于各强有力的刺激物,唾液分泌反应消失,并且这只狗对于第一次的强刺激,很迟缓地吃食物,对于第二次的强刺激却完全不吃食物。对于弱刺激物,分泌虽然减少而是存在的,狗即刻吃食物。在这实验的翌日,再应用寻常10秒钟的孤立条件刺激,情形就更恶化。对于全部的各刺激物,分泌效果都缺少,狗在强刺激物的场合不肯吃,而在弱刺激物的场合倒肯吃。在第三天,一切都恢复正常,不过当天第一次被应用的、时间加长的孤立条件刺激音比正常时有了减弱一半的分泌效力。此实验再重复了一次,结果与前相同。这是多么精确的反应,实验条件微不足道的变化有了多么显著的作用!
然而事情不仅是限于反感性的。在以前提及过的另一只狗的场合,即在一只非常制止型的叫做“聪明的女人”的那只狗的场合,实验条件的这样变动,引起了相反的结果。因为在安静的环境里,这只狗在实验架台上是容易瞌睡的,并且对于各条件刺激物以及对于食物的反射都是完全缺乏的,所以应用了已经考验过的一个引起兴奋的处理方法,就是在条件刺激物作用开始以后极短的时间(0.5~1秒)内把食物给它吃。应用这样处理方法3周以后,狗的瞌睡状态消散了,并且它每次都接受食物而贪食。当以后把孤立的条件刺激加长到5秒钟的时候,条件性分泌效果也就出现了。可是这效力只维持了几天,以后就消失了,这只狗又容易瞌睡。然而只要把各条件刺激的时间延长到10秒钟,这只狗又变成更活泼,并且分泌性效力又出现,而在本质上值得注意的是,这分泌效力在条件刺激的起初5秒内已经出现了。以后的情形又是老一套故事的重复。在几天以后,这只狗又陷于以前的瞌睡状态,已经没有条件分泌的反射。条件刺激时间延长到15秒钟,这也与第一次延长到10秒钟的作用相同,就是这只狗又变成活泼,条件唾液分泌性的效力很显著,并且在刺激的最初5秒内已经开始。在条件刺激物延长到20秒乃至25秒的场合,情形也是如此。
我举出实验记录如下。

在下次的实验,各条件刺激的时间延长到20秒。

这样,孤立的条件刺激时间每一回不很大的延长,都引起狗的一时性兴奋:瞌睡情形消散,并且唾液条件效果的出现时间与条件刺激开始时间很相接近,这是在条件刺激时延长以前所未有的。所以这是大脑皮质兴奋的结果,而不仅是由于以发现这效果的目的而把时间延长的结果。
在实验内的同样的一种完全微细的变动,对于两只狗却决定了完全对立的效力。对于一只狗,这类变动引起了兴奋;相反地,对于另一只狗,排除了兴奋。不能不这样想,效力相反的根据是在于第一只狗原有的活泼(兴奋的)状态和第二只狗原有的睡眠(被制止的)状态。
我再引用一个事例。显然,在此例中,现象根本关系的乖离是由于目前还不明了的次要性的条件而起的。如在以前各讲内多次指明过的,属于各个不同分析器的外来动因所构成的条件刺激物,在其他各条件都相同的场合,会产生各不相同的效力数值。我们很长久地只受了这样一个先入之见的影响,以为光线、声响等等感觉上的差异必定是基于各有关的皮质细胞物质基础上的区别,因为在现代生理学上,一切神经内的神经过程都是认为相同的,所以我们就抱了一个倾向,以为应该把条件刺激数值的差异归纳于各种不同分析器的细胞个性的缘故。但在这些讲义的进行期间,若干事实已经要求我们特别地做与此点有关的研究。并且同时明了了,正如已经说明过的,在各种不同分析器的动因构成条件刺激物的场合,各条件效果量的差异在本质上是与每个动因送进大脑皮质内的能量(количествоэнергни)大小有关的。像你们可以想到的,这个结果是这样获得的:我们老早知道,我们通常从两个不同分析器各采取一个动因而做成一个条件性复合刺激物,于是这复合条件性刺激物的作用,主要是,或者几乎完全是一个成分的作用,这是由于对两个成分的条件性作用分别地加以检查而能证明的。譬如我们通常的复合刺激物中的声音性动因总是掩蔽光性动因的、皮肤机械性的和皮肤温度性各动因的效力。可是,如果故意地把声音性动因非常弱化而把光性动因强化,我们就获得相反的关系,因此也可以证明,能量具有决定性的意义。然而条件反射量与能量有关的重要事实,在若干实验动物个别实验的场合,并不能证明,这虽然不是很常有的、但是有时有的事情。我们对于这一点虽然非常注意,可是直到现在,我们并不能确定地指明与这些乖离有关的特殊条件。个别的条件虽然或多或少地被确定了,可是这些条件的全体依然是不能捉摸的。这种乖离是显然与神经系统的一般特性相关的,就是与神经系统的兴奋型和制止型有关的。在制止型狗的场合,效果量与刺激强度的通常比例显现得特别明了,并且除了显著的病态以外,这是几乎无例外的。其次,孤立的条件刺激时间也具有一定的意义。如果这刺激时间不很大,差异就可能不发生。假定强刺激与弱刺激起初的效力相同,但在弱刺激的场合,效力在刺激继续的时期以内并不十分强烈地逐渐增强,而有时或者完全并不增强,相反地,强刺激的效力却几乎总是逐渐增强的。相反地,在非常兴奋型的、贪食的狗的场合,孤立的条件刺激时间的缩短,会促进条件效力数值与刺激强度间的通常关系的出现,而在比较长时间的条件刺激的场合,这样关系是不显著的。
构成大脑两半球活动的各现象是具有非常的条件制约性的。由于这异常的条件制约性,并在许多简单的、似乎已经经过相当研究的事实的场合,我们还是不能不感觉,现在生理学的分析是不能使人满意的。我们试举一个不久以前的事例[柏德可琶叶夫(Н.А.Подкопаев)和魏尔及可夫斯基(С.Н.Выржиковский)两人的实验]。这只狗在如下变式的各条件之下,用各种不同的动因形成了条件反射。第一个无关性动因的应用,一次是与无条件刺激物(食物)并用的,其次的一次却是单独地作用的——这样地重复应用若干次。条件反射相当迅速地形成了(在第20次)。其次动因的应用,是每3次之中,只有一次并用食物。反射也形成了,并且形成是更快的(在第七次),但是其时这只狗变成非常兴奋。末了,一个动因的重复应用是每4次之内才并用食物一次,于是这个动因不曾成为条件刺激物,并且这只狗多少瞌睡了。这个动因一共被应用240次,其中只有60次并用了食物。我们试用其他一些基本的事实理解这事实的机制:为什么在这场合条件反射不曾形成呢,或者至少假定,在应用回数更多的场合这条件反射是可能形成的,那么,为什么条件反射的形成如此缓慢呢?条件反射形成的基本的机制是两个刺激的遭遇,就是大脑两半球皮质某一定点的兴奋与另一个点的(即似乎与大脑皮质另一个点的)更强的兴奋,在时间上的一致发生,因此在这两点之间就或快或慢地拓通一条比较轻便的道路,形成了联合的关系。相反地,如果长久地两个刺激不再相遭遇,拓通性(проторенность)就会消失,联系就会破裂。然而当拓通性达到最大限度的时候,它就不需要继续的练习而会保存几个月乃至几年之久。在适当的条件之下,就是说,在刺激互相遭遇的时候,拓通性必定在第一次的遭遇时就会形成,并且在以后各次遭遇的时候,拓通性会不断地增强,会有累积的效力。于是发生一个合理的问题:为什么在上述最后方式实验的时候,条件反射不曾成立呢?确实,在这个实验里,两种刺激物的遭遇回数有60次之多,条件反射并不曾成立,而在通常条件反射形成最顺利的场合,不过需3~5次的遭遇而已。但在上述第一个变式实验的场合,在两刺激物互相遭遇20次以后,条件反射才形成,这也是通常在我们实验环境里一只狗的条件反射的最初形成的时候所需要的刺激回数。当然,起先我们会想及两刺激物互相遭遇各次间间隔时程长短的影响。但在这个事例,这是没有意义的。如果各次遭遇的间隔时程是相等的,而所应用的无关性刺激物没有一次不并用食物,那么,条件反射无疑地也许会形成了。就是说,障碍不是在此。我们不能不假定,正是多次重复地只应用无关性动因而不并用无条件刺激物,这是对于条件形成发生了障碍。我们应该怎样地解释这个障碍呢?像我们已经知道的,任何一个新的刺激物起先引起探索反射,可是如果以后该刺激物不再对动物发生作用,就不再引起探索反射。这刺激物作用的停止,是由于在刺激所趋向的皮质细胞里有了制止过程展开的缘故。所以在上述第三变式的实验里一个动因反复应用3次而不并用食物的场合,有关的皮质细胞的制止过程是可能发生的,于是对于以后所使用的食物刺激,并不曾有兴奋过程的产生。这个假定似乎有充足的根据,可是在本事例,这并不能认为是圆满的。当第三方式实验的新动因被重复应用到240次的时候,我们检查了它的特性。在此动因作用以后很短的时间以内(30秒)应用了这只狗原有的条件反射中的任何一个,该条件反射决不曾显出被制止的状态。所以该动因不是制止性的,并且不曾引起后继性制止。可是因为长久应用的结果,它的制止性作用必定是非常集中的。但是这个假定也不曾被证实。当在240次应用以后开始把这动因与食物接连地并用的时候,该动因在第三次应用以后就显现了著明的条件性分泌的效力。需要再补充几句话,就是在此动因第一天与食物并用两次的时候,早期的各条件反射的效力很微弱,就是说,这只狗的兴奋性是不大的。这样看来,新条件反射形成的速度是极大的,于是以为皮质细胞由这无关性动因而具有制止状态的想法,也是没有根据的。我们把从我们一些既知事实所能做的假定都试验过了,也不曾发现这个现象的原因。我们将继续检查这个原因,并且不能不相信,我们是会发现它的。然而直到现在,我们所做的试验却昭示着我们关于决定条件反射形成机制的一切条件还不曾有完全的知识。在第二讲内所引用的实验条件,虽然对于我们直到现在所能掌握的一切条件反射的形成都是充足的,然而还是不完全的,所以即使我们对这些条件加以考虑,也还不能即刻理解本分析例的结果。必定还有被我们所忽视的某个条件的存在。就是说,这又是这一类现象的非常的条件制约性。
我们现在还谈不到,从生理化学的详细事项可以理解大脑两半球的一切活动,这是很显然的;我们从神经组织的基本性特色而理解这种活动的可靠机会很像是没有的;甚至于有关这种机能的根本现象的本身,我们也还不曾有完全的知识。既然如此,剩下的需要我们研究的是什么,我们的研究的要点是什么?显然,把无数的各种不同的个别现象都归纳于比较少数的共通的基本现象,这必定就是构成大脑两半球生理学研究现在阶段的最近而最实在的任务。我们也正在从事于这类工作,时而在个别的场合多少接近着这个目的,时而站在尚未解决的、正在研究中的问题的前面,时而犯着错误。
在我们这研究的初期,根据强烈的、外在的特征,我们区别了三种制止过程:外制止、内制止及睡眠性制止。如你们已经知道的,我们所汇集的事实材料允许我们把后方两类合并为一类,就是把这两者表面的区别当做次要的细情看待。两者是同一的内制止过程,不过一个是断片的、限于一个部位的,而另一个是融合的、广泛地扩展的。当我们继续研究条件反射的时候,我们认识了互相诱导的现象。当然,负性诱导的事实与外制止过程的事实,两者在基本的过程上彼此相似的情形,引起了我们的注意。现在,我们旧分类的各种制止过程,其根本类似性似乎是已经很可能了,所以我们也在这个研究方向上,着手于资料的汇集。这些资料的一部分,以前已经说明过,一部分是我将要引证的。在诱导相有关的一章讲义内记载了一个如下的实验,就是皮肤某一定部位的机械性刺激的条件性防御反射,在它的后作用的期间,制止了其他各皮肤部位的机械刺激性条件性食物反射。其时一个细节证明了,这个制止过程正是发生于大脑皮质内两点之间的(兴奋点与制止点)。在向完全的内制止过程进行的各移行时相有关的一章讲义内,在若干事例的场合曾经指明,即在外制止的时期以内,也可能观察各移行的时相。现在我能再补充一些新的见解和新事实,这是有利于内制止与外制止的相同性的结论的。在上述的复合条件刺激物中弱刺激物为强刺激物所掩蔽抑压的事实上,不能不发现外制止过程的显现。强刺激物的皮质细胞制止了弱刺激物的皮质细胞,所以后者的皮质细胞只能与无条件刺激物的皮质细胞成立薄弱的联系,因为我们有根据地可以认为,结合道路的拓通性是与条件刺激物的强度相当的。但是在阳性条件刺激物皮质点强化了或者甚至于恢复了条件制止性刺激物皮质点的制止状态的时候,负性诱导在该场合与外制止过程可能有什么区别呢?最后,也可以证明我们所树立的论题,是如下的事实。老早以前,我们的许多工作同人[密仕托夫特(Миштовт)、克尔瑞序可夫斯基(Кржышковский)、来柏尔斯基(Лепорский)诸人]已经指明过,在条件制止物形成的场合,在迅速地完全地实现制止过程的关系上具有重要意义的是,在物理学的强度上构成条件制止物的一个动因不可比复合制止物中另一个成分的动因即阳性条件刺激物更薄弱。不久以前的一些特殊实验(弗尔西柯夫)完全证实了这些说明,并且补充了一个事实,就是新动因的作用往往以外制止过程而开始,引起探索反射,其次就不触目地移行于复合刺激物的制止作用,这就与内制止过程相当。然而还剩下一个问题:这就充足地成为内制止与外制止相同的理由吗?
在第十九讲内举出的一个问题,这是与三种直接引起大脑两半球皮质制止状态的刺激物有关的。很弱的刺激物、很强的刺激物及不寻常的刺激物,同时假定地指明过这个事实在生物学方面的意义。有关这些各不相同的刺激物作用的生理学机制的问题,被搁置到本讲。然而即在此地,现在我也认为,从制止过程的全范围提出问题而加以讨论,这是不可能的。不管我们的实验材料如何多,但如果关于制止及其与兴奋的关系而要作成一个共通的、一定的观念,这些材料就显然是不充足的。对于一组事实很适当的说明,并不能包括一切的事实。许多的事实顽强地不能由理论的分析而加以说明,并且在我们研究进行之中,有关这些事实的概念并不是能满意的,所以我们不能不几次地更改了我们的概念。所以在这个问题上,正如在我们一般工作上,目前只能进行事实材料分类的工作。我们正面临着无数未解决的问题:就是有关解除制止的问题,制止性刺激物阳性作用的问题,在若干事例大脑两半球有障碍的场合阳性条件刺激物的直接制止性作用的问题,以及有关弱阳性刺激物及强阳性刺激物的制止性作用占优势的问题及其他等等的问题。在许多场合,我们不能说:许多事实之中,那些是彼此相近、彼此相同的,或者那些是彼此完全各别的、彻底互不相同的。为了说明这些场合的困难,我要仔细研究下方一个例子。为什么新的现象,或若干现象的新结合会对于动物发挥制止性的作用呢?这个抑制过程的生理学机制是什么?我们对于一只实验狗,本来是在条件刺激物应用以后,把装盛食物的小碗从影幕后向前方推出来的,但现在改用另一个方法,就是在狗的面前放一个小碗,从位于上方的食物储存器内经过一个小管子,把食物投入于小碗之内,这样做,实验狗中的多只就会顽固地不吃这食物,条件反射也都消失。当然,这是制止过程发生了。这个制止过程应该归纳于什么原因呢,应该与什么已知的其他事实可以相并列呢?也许这可能与第十三讲内所记载的一个事实相并列罢,就是在应用各种不同条件刺激物的时候,如果非常更动了各刺激物原有的次序,全部各条件反射性活动的制止过程就会或多或少地显著地发生,特别在若干狗的场合是这样的。并且纵然恢复各条件刺激物原有的排列次序,这制止过程还会继续存在几天。关于狗的周围现实环境的一切,我们也可以进行同样的想象。在动物的面前,外在的、以一定的连续关系而反复发生的各现象,可以说,在大脑两半球里构成一种常同性的活动。任何一个新的现象,或者一些旧有的现象所形成的一个新结合,都会损害这个常同性,因而就引起制止过程的发生,这正与我们若干实验里各条件刺激物排列次序的变动所引起的制止过程是相同的。于是就会发生如下的一个问题:这最后场合的制止过程是怎样地,以什么生理学的过程而发生呢?或者更正确地说,这个场合的制止过程是与什么其他的制止过程的场合相同呢?在外方环境有新动摇的场合,会有探索反射的发生,而上述制止过程不就是这类探索反射制止性作用的结果吗?或者这是些完全个别的、独立的现象吗?在非常制止型狗的场合,敏感的方位判定反射与强烈的后继性制止过程的联系,表现得非常显著(在前述狗“聪明的女人”的实验的场合,这个事实很明了地发生了),这对于第一个解释也许是有利的,所以在这样实验狗的场合,这样假定的机制是很像可能的。然而在一些其他的场合,并没有任何方位判定反射的征兆,而制止过程却是直接地发生的。
这个例子昭示着,我们还是与个别事实满意的分类法相距很远的。
这就是为什么,关于各制止过程的现存理论,我还不肯主张哪一个是正确的,并且我不肯提出新的理论。为了我们事实材料系统化和新实验设计的目的,目前我们利用着一些暂时的假定。
最后我转而说明我们的错误。在过去,我们犯了不少的错误,可是我们渐渐改正了,现在也改正着。现在先说明一个错误。这个错误是在这些讲义进行中各旧实验反复进行的时候所发现的,并且是已经改正的,我还说明另一个很像可能的、正在研究中的错误。
在第四讲内,我们关于消去性条件反射的恢复有过说明,这类恢复有3个相异的场合:一个即是在一定时间经过以后的自动的恢复,这是稳定的、但或快或慢地逐渐增强的恢复;而其次的两个场合却是加速的恢复,或者是利用食物强化的手续,就是说,应用特殊的无条件反射,或者是应用某种新异的反射。这最后的两种场合在第四讲内是当做本质不同的东西而记载的。在前者的场合,恢复不仅是迅速的,而且是稳定的。在后者利用新反射的场合,我们已经确定了,恢复虽然很快,但不过是一时性的,就是在新异反射停止及其后作用消失以后,恢复的效力就会很快地消失而又为制止过程所代替,于是继续到消去性反射通常所需要的时期为止,好像并不曾有任何新异反射被应用过。所以这个恢复获得一个特殊的名称——“解除制止”。并且在第四讲内指明过,既然消去性过程并不是条件反射的根本破坏,所以很难了解这个区别。最近我们关于条件刺激物与无条件刺激物的相互作用已经获得了新的资料,所以我们认为有把旧实验尽可能精确地再做的必要,并且果然确证了旧实验的错误(柏德可琶叶夫的新实验)。在应用特殊的无条件反射或任何其他新异反射的场合,消去性反射的恢复都不过是一时性的,就是说,在应用这两种反射以后的最近时间以内,消去的条件刺激物就具有阳性作用,但经过若干时间以后又会丧失这阳性作用,直到消去性反射的自然恢复为止。当条件反射是食物性反射而新异反射是酸反射的时候,就是说,当两个都是化学性反射的时候,一时性的反射恢复,即解除制止现象的发生,都是在时间经过的关系上完全一样的。对于两只狗做了这样实验,所得的结果完全相同。我现在详细地说明一只狗的实验。在拍节机条件性食物反射消去而达到零的场合,在条件刺激物最后一次不受强化处理以后,这零相继续了20分钟。以后该反射开始慢慢地自动地恢复,在30分钟的时候,达到消去过程以前原有初期条件效果量的40%。但是在该消去性反射第一次零相出现的场合,条件刺激物即刻就受强化的手续,并在零相后过20分钟,再检查该条件刺激物,那么,条件效果依然是零的。在另一个实验里,在与上述相同的强化处理以后,只与消去性反射零相相隔10分钟即刻应用条件刺激物,效果却是阳性的。如果条件反射消去以后在同一时期内用酸注入狗口中以代替食物,并在同一时期内检查条件刺激物,实验的进行,也就与上述应用食物的实验完全相同。
现在当做例子,举出若干实验的实在数字。
在实验这一天第一次被应用的条件刺激物在20秒间给予了唾液6滴;在条件反射消去而成零相以后,该条件刺激物即刻受强化手续,从消去的最后起算,过10分钟再受检查,效力是3滴。
在另一天,条件刺激物被应用的最初20秒间的效力是7滴;在反射消去以后,即刻就应用强化手续,而在消去的零相以后经过20分钟,该条件刺激物再受检查,效力是零。
在实验这一天,条件刺激物第一次被应用的20秒间,效力是5滴;在条件反射消去而成零以后即刻用酸液注入于狗的口内,并且在零相最初的发现以后经过10分钟,就检查条件刺激物,效力是2滴。
在另一天,条件刺激物最初次被应用的20秒间,效力是5滴;在条件反射消去而成零以后,即刻用酸液注入于狗的口内,并在零相发现以后经过20分钟,条件刺激物再受检查,效力依然是零。
解除制止的最大值是在消去反射的零相发现后10分钟以前就出现。
很显然,以前的错误一部分是由于不精确的实验而起的。当时在应用特殊的无条件反射和其他新异反射的场合,最常用的新异反射是声音性的、光性的或其他的反射,而这些反射都只有短时间的后作用,而不曾应用过具有后作用时间较长的化学性反射,于是比较了消去性反射恢复时期的长短。以前错误另一部分的理由是,由于我们想法倾向于一个旧的结论,以为在条件刺激物消去以后而恢复的场合,与条件刺激物相结合的无条件刺激物必须对于条件刺激物具有特殊的关系。现在实验的结果对于如下的假定提供了一个充分的根据,就是制止过程是发生于神经细胞之内的,而不是发生于结合点内的,也不是发生于条件刺激物皮质细胞和特殊的无条件刺激物皮质细胞两者间的结合道路之内的。否则就难于想象,这两个刺激物的作用怎样会能够是完全相同的。
另一个错误现在正在暴露之中。虽然这对象的再研究还是正在开始,我认为现在可以谈及它:从一方面说,因为与这错误有关的一点具有特殊的重大性;从另一方面说,因为我们的研究范围具有一个共通的特色,就是在确定正确的事实关系的场合,充满了无比的困难。
在第二讲内,似乎形成条件反射的一切条件都被举出了,同时妨碍条件反射形成的一切事项也被举出了。第一个条件是,无关性动因与无条件刺激物在时间上是同时的。在这第一条件后,其次就是未来的条件刺激物必须以极短的时间先行于无条件刺激物之前。对于许多的狗,在无关性动因与无条件刺激物复合地重复应用很多次以后(甚至在300~400次以后),如果每次都先应用无条件刺激物而只在5~10秒钟以后才应用无关性动因,那么,条件反射就不会成立,但是对于相同的这些狗,如果按照普通的方法利用其他的一些动因,与无条件刺激物相结合,那么,在反复应用7~20次以后,条件反射就成立了。当然我们会这样想,强有力的无条件刺激物,被集中于大脑两半球的一定部位,由于外制止机制的关系,这就成为大脑两半球其他各部分制止过程发生的条件,于是向这些部分进行的刺激都成为无效。这是完全与我们自己知道的一种情形相同,就是当我们为某一件事情而非常忙的时候,我们不听见也不看见在我们周围所发生的事情。这样,从我们的观点和从日常生活的观点而言,这个事实是显然自明的。直到最近,这个见解使我们对于它不抱任何一点的怀疑。现在情形完全变了。当我们受了如下一个质问的时候:究竟怎样地条件刺激的早期强化手续,就是说,条件刺激物孤立作用的尽量地缩短,会妨碍大脑皮质内该刺激作用点制止状态的发生呢?于是我们做了变式的新实验,获得了出于我们意外的事实。结果是,无条件刺激物也制止已形成的条件刺激物。然而其时,各事实如下的对照当然引起我们的注意。或者已形成的条件刺激物是存在着的,或者这还是一个无关性的动因,而条件刺激物会由该动因而形成,可是无条件刺激物对于该动因的影响是相同的。同时,这影响的表现是这样的,就是在外方条件和时间条件有极小变动的场合,条件反射形成的情形就会成立,就是说,一个无关性动因会变成一个一定的条件刺激物。不能不承认,条件反射形成的机制与外制止的机制仿佛是互相接近的东西,或者甚至可能是相同的东西,换句话说,外制止过程产生一个基础,以形成细胞间的联系。这就使我们想起另一个事实,就是第二级条件反射形成及条件性制止过程发展的事实,在这个场合,也是在共通的外方条件之下,不过由于刺激时间的若干微小的区别,就或者发生兴奋过程,或者发生制止过程。如果这个见解是可靠的,那么,不能不期望着,即在我们所谓遮蔽(покрытие)现象的场合,即从一个无关性动因为无条件刺激物所遮蔽的最初时候起,在大脑皮质内这两点之间,起初就可能产生了联系,可能条件反射会成立,不过在以后这两个刺激物再三地被复合应用的场合,条件反射却被制止住了。与我们最初的方位判定有关的一些实验也昭示着这一可能性。已形成的各条件刺激物,如果完全地为无条件刺激物所遮蔽,不过慢慢地引起条件效力的减弱,而在弱条件反射的场合,该条件反射的效力比较快地减弱,甚至有时减弱至零,但在强条件反射的场合,不过使该条件反射效力慢慢地减弱,并且往往减弱得不很多。根据这一事实,我们在一个无条件刺激物与一个无关性动因作成新的结合的时候,即在无条件刺激物先被给予的时候,我们只应用这个结合不过数次,以防制止过程的发展。在许多实验的场合,我们实在获得了我们所期待的结果。在这样的复合刺激物的实验以后,单独地被检查的该无关性动因却发挥了与条件刺激物相同的作用(巴夫洛娃、克列勃斯、柏德可琶叶夫、库帕洛夫诸人实验)。当我们用这种眼光及这一动向的实验审视我们这一类的各个做过的实验的时候(克列斯托夫尼可夫实验),我们看见了如下的事实:第一,各旧实验中无关性动因条件作用的检查几乎都是在复合物很多次应用以后,而且是在无条件刺激物作用开始以后,无关性动因才与无条件刺激物相结合;第二,即在这样的场合,强有力的无关性动因,如果单独地受检查,也往往很长时间地显出唾液分泌的效力。可是与这阳性效力有关的解释总是把它当做偶然的新奇的,而不当做条件性效力看待,这因为我们有了一个偏见,以为如果这效力真正是条件性的,那么,它也许因重复应用回数的加多而会更强,不致像这些旧实验的情形,反而会减弱或消失。可能的情形是,在当时检查的场合,另一个现象,即明显的条件运动反应的缺乏,也由于这先入之见,不曾完全客观地加以完全的研究。在当时有一只狗的实验,必定应该承认有真正条件反射的成立,可是其时也把这个情形认为是由于一个新异条件的缘故,当然,当时这是有若干根据的,可是根据并不充足。这个主题的研究应该是多方面的、严格的,同时要利用有关本问题很多的准备知识,并且需要在这新研究范围上,我们的思想有更多的特殊的训练。在我们的解释被完全认为正当的场合,并且在我们现在指南性实验以后继续被证实的场合,大脑两半球生理学也许可以掌握一个适用于人类的很重要的论题,就是大脑皮质内新联系的形成,不仅发生于大脑内最优良的兴奋性的部位,而也发生于大脑内多少被制止的部位。
然而我在这最后的、也是事实材料的一章讲义内,我想对我们研究范围的工作,尽可能地报告最基本的印象。我以为,在以前全部讲义内所说明的多数事实充足地证明着,在这个研究范围内,严肃的自然科学性的研究、精确的事实资料的汇集是可能的。所以不会因为本讲内所引用的事实而担心损毁我们这个科学性研究事业的信誉。当然,承认危险的存在,总比把危险置之度外的态度更好些。此外,我想提醒将来从事这一领域的研究工作者,他们在该范围内是会遭遇无比的困难的。
一般地说,生理学这门新的学科,是真正富于魅力的,可以满足人类精神两个永远并行的倾向——一个是不断地向新而又新的真理的掌握的努力,一个是对于仿佛知识已经完美的一种僭越主张的反抗。显而易见,在今后的长久时期内,未知事实的大山仍然要无可比拟地高于既知事实的断片。
