第13章 种间杂交与染色体数目上的变化
由于杂种生殖细胞内染色体不能结合,所以就保留了双倍数目的染色体。这个双倍数目可以通过回交继续维持下去,但是由于杂种缺乏生殖力,所以在自然条件下,就很难从这种组合产生任何永久的类型。
配制饲养果蝇的培养基所需的部分原料(李凤霞提供)。
从不同染色体数目的物种相互杂交的结果中,揭发了一些有趣的关系。一个物种的染色体或者恰好是另一物种的二倍或三倍,在另一些例子里,数目较多的染色体群也许不是另一群的倍数。
1903—1904年,Rosenberg所进行的两种茅膏菜杂交实验,便是一个经典例子。
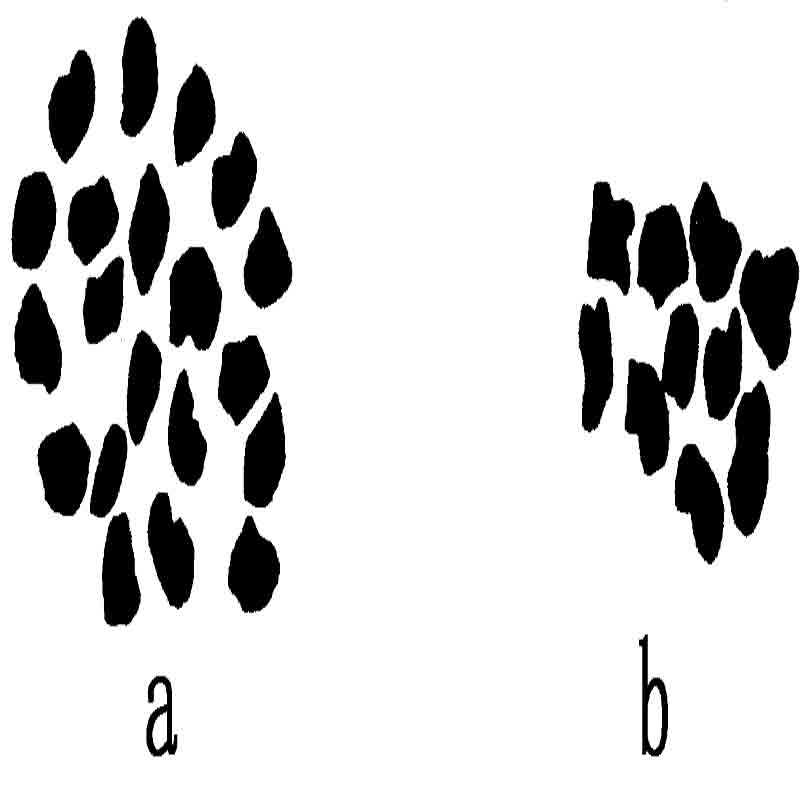
图108 圆叶茅膏菜的二倍染色体群和单倍染色体群(仿Rosenberg)
茅膏菜的长叶种(Drosera longifolia)有40条染色体(n=20)、圆叶种(D.rotundifolia)有20条(n=10)(图108)。杂种有30条(20+10)。杂种生殖细胞成熟时,共有10条接合染色体,即通常所谓的二价染色体,和10条单染色体(单价染色体)。据Rosenberg的解释,认为这意味着长叶种的10条染色体同圆叶种的10条染色体联合,长叶种的其他10条染色体则没有配偶。在生殖细胞第一次分裂时,接合染色体各分为两条,分别向相反的一极进行;10条单染色体没有分裂,不规则地分布在两个子细胞里面。可惜这样的杂种没有生殖力,不能供进一步遗传学研究之用。
Goodspeed和Clausen广泛地研究了两种烟草(Nicotiana Tabacum和N.sylvestris)之间的杂交。但两者的染色体数目直到最近才确定下来。N.Tabacum有24条(n=12),N.sylvestris有48条(n=24)。这种染色体数目上的差异,还没有同遗传学研究结果联系起来;染色体在成熟分裂中的行动,也没有见过报告。
两种烟草杂交后的杂种,完全同Tabacum亲型类似,甚至该亲型的基因对于Tabacum种的正常因子呈纯隐性作用时(即同Tabacum型的一些变种杂交)也是这样。Goodspeed和Clausen以为这是Tabacum的整群基因对于sylvestris基因呈显性作用的结果。他们这样表示说:Tabacum的“反应系”,在杂种的胚胎发育过程中,占优势,或者说:“两系的要素相互间一定是极其矛盾的”。
杂种有高度的不孕性,但也形成了少数有作用的胚珠。像繁育结果所证明的,这些胚珠或者完全(或大多数)属于纯粹sylvestris型,或者完全(或大多数)属于纯粹Tabacum型。看来,杂种的胚珠中只有具备任一型的整组染色体(或几乎是整组染色体)的,才能(或大多数能够)有作用。下述实验是这种见解的根据。
杂种同sylvestris花粉受精,产生了各种各样的类型。其中,很多植株的性状完全为纯粹sylvestris型。这些植株都是可孕的,产生纯sylvestris型的后代,因此必须假定它们是由含sylvestris染色体群的胚珠同sylvestris花粉受精而成。也有和sylvestris相类似的植株,却含有大致由Tabacum染色体群得来的其他要素。它们都没有生殖能力。
杂种回交Tabacum,没有成功,但从田间自由传粉中出现了少数杂种,同Tabacum相似,一定是和Tabacum花粉受精而成。其中有些是可孕的。它们的后代绝不表现sylvestris性状。不论它们有什么Tabacum基因,这些基因都表现(孟德尔式的)分离现象。这里也有不孕性的植株,同Tabacum和sylvestris杂交所生的杂种相似。
这些不平常的结果,还有另一方面的重要性。子代杂种可以用两个方法产生出来,即每一个物种都可作为胚珠的母株。由此得出结论:即使是在sylvestris的胞质里面,Tabacum的一群基因也完全决定了个体的性状。鉴于这项结果是由大相悬殊的两个物种的胞质得来的,所以这就成为基因在决定个体性状上有影响的一个有力证据。
Goodspeed和Clausen所提出的反应系观念虽然新颖,原则上却与基因的一般解释毫无矛盾。它仅仅意味着:当sylvestris的单组基因同Tabacum的单组基因对立时,sylvestris的基因完全潜伏,没有作用。但sylvestris染色体依然保持原状。它们未被扔掉,也没有受到亏损,因为从杂种同sylvestris亲株的回交中,可以重新得到一组有作用的sylvestris染色体。
Babcock和Collins用各种黄鹌菜(Crepis)进行了广泛的杂交。Mann女士(1925)也研究过这些杂种的染色体。
Crepis setosa有8条染色体(n=14),Crepis capillaris有6条染色体(n=3)。Collins和Mann用两种杂交,杂种有7条染色体。在成熟时,有些染色体结合成对,另一些染色体不经过分裂便分散在花粉母细胞内,形成胞核,各有2~6条染色体。第二次分裂时,所有的染色体,至少在为数较多的一群里的染色体,都各自分裂,子染色体分别进入相反的两极。胞质往往分裂成四个细胞;但有时也分裂成2粒、3粒、4粒或6粒小孢子。
这些含7条染色体的杂种,不产生有作用的花粉,不过有一些胚珠还是有作用的。杂种胚珠同某一亲株的花粉受精,产生了五个植株,各有7条到8条染色体。检查一株含8条染色体的成熟分裂,发现有4条二价染色体,其分裂情况正常。这一株的性状同C.setosa相似,并且有同样类型的染色体。这样便恢复了一种亲型。
另用Crepis biennis同C.setosa杂交,前者有40条染色体(n=20),后者有8条(n=4)。杂种有24条(20+4)(图109)。当杂种生殖细胞成熟分裂时,至少有10条二价染色体和少数单价染色体。
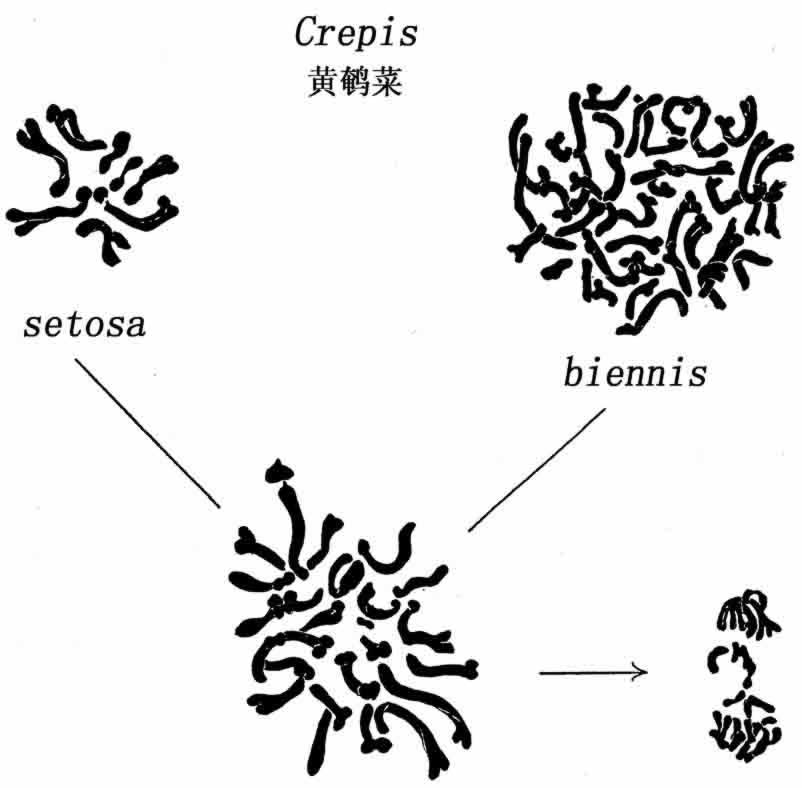
图109 Crepis setosa和C.biennis染色体群(仿Collins和Mann)
由此可知,既然setosa只提供了4条染色体,那么,biennis染色体中一定有一些互相结合。以后细胞分裂,有2条到4条的染色体比其他染色体落后,但最后大都进入一个胞核。
子代杂种是可孕的。孙代(F2 )植株有24条或25条染色体。从这里似乎有产生一个稳定的新型的希望;这种新型有新的染色体数目,其中,有一对或多对染色体从染色体数目较小的物种方面得来。杂种中有10条二价染色体,这一事实便意味着Crepis biennis是一个多倍体,可能是一个八倍体。子代杂种的同类染色体联合成对。这个含有半数biennis染色体的杂种(F1 )是一年生植物,与二年生的biennis不同。染色体数目上的减半,引起了生活习性上的变化。植株成熟时间,只有二年生型的一半。
Longley描写过两型墨西哥大刍草teosinte:一型名为墨西哥型mexicana,为20条染色体(n=10)的一年生植物,另一型名为多年型perennis,是多年生植物,有40条染色体(n=20)。两型都有正常的减数分裂。用二倍体teosinte(n=10)同玉蜀黍(n=10)杂交,杂种有20条染色体。在成熟时,杂种生殖细胞各有10条二价染色体。普通会把这种情况解释为10条teosinte染色体同10条玉蜀黍染色体之间的联合。
多年生teosinte(n=20)同玉蜀黍(n=10)杂交,其杂种有30条染色体。在杂种的花粉母细胞第一次成熟分裂中,看到了一些三价染色体群,疏松地结合起来,有一些二价染色体,也有一些单染色体:三者之间的比例为4∶6∶6或1∶9∶9或2∶10∶4(参考图110b)。在第一次成熟分裂中,二价染色体各自分裂,两染色体分别进入相反的两极;三价染色体也进行分裂,两条同入一极,另一条则进入另一极;单染色体行动迟缓,零乱地分布(没有分裂)到两极上去(图110c)。结果,分布极不平衡。

图110 减数以后的染色体群:a.多年生teosinte;b.同玉蜀黍杂交后的杂种;c.玉蜀黍的减数分裂(仿Longley)
最近有一个例子,谈到染色体数目很不相同的两个物种互相杂交,产生了有生殖能力的、稳定的新型杂种。Ljungdahl(1924)用有14条染色体(n=7)的罂粟Papaver nudicaule和有70条染色体(n=35)的P.striatocarpum进行杂交(图111)。杂种有42条染色体。杂种生殖细胞成熟时,出现了21条二价染色体(图111b~e)。二价染色体分裂,每极各得21条染色体。这里没有一条单染色体,也没有一条逗留在纺锤体上。这些结果只能作下述解释:nudicaule的7条染色体,同striatocarpum的7条染色体结合,其余24条striatocarpum染色体则两两结合,形成14条二价染色体。两者共得21条二价染色体,正是观察所得到的数字。由此自然得出下列假设:有70条染色体的(n=35)striatocarpum型多份是一种十倍体,也就是说,每一类的染色体都有10条。
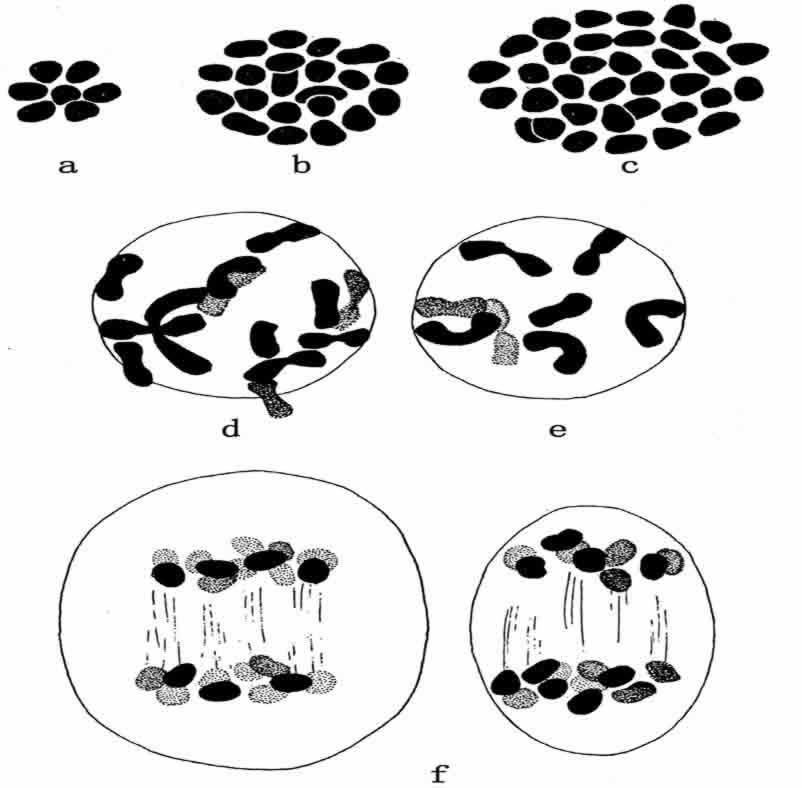
图111 两种罂粟之间的杂交:a.Papaver nudicaule有14条染色体(n=7);
b.杂种,有42条染色体(n=21);c.P.striatocarpum有70条染色体(n=35);d~e.杂种的胚胎母细胞;f.杂种第一次成熟分裂的后期(仿Ljungdahl)
子代新型产生含21条染色体的生殖细胞。新型平衡、稳定。它也是可孕的,预期可以产生一个稳定的新型。由此又产生其他稳定的类型,在理论上也是可能的。如果子代新型回交nudicaule,应该产生四倍型(21+7=28);回交striatocarpum,应该产生八倍型(21+35=56)。这里,通过二倍体同十倍体的杂交,可以产生以后世代中的四倍体、六倍体和八倍体,这些类型都是稳定的。
Federley关于Pygaera属各蛾种的实验(参考第9章),说明了一项极不相同的关系。由于杂种生殖细胞内染色体不能结合,所以就保留了双倍数目的染色体。这个双倍数目可以通过回交继续维持下去,但是由于杂种缺乏生殖力,所以在自然条件下,就很难从这种组合产生任何永久的类型。
