二倍体雌性与单倍体雄性
轮虫首先经过许多世代的单性生殖,各代雌虫有两倍数目的染色体。卵子没有减数分裂,只放出一个极体。单性世代在一定营养条件下似乎可以无限期地绵延。不过正如Whitney所证明的,改变营养——例如用绿色鞭毛虫类饲养轮虫,便能结束单性生殖世代。在这种营养下,雌虫所产生的下一代雌虫(借单性生殖)有双重可能性。这一代的雌虫如果同雌虫(当时可能出现)受精,则在每个卵子成熟以前,只有一条精子进入卵内。卵子在卵巢内长大,卵外披一层厚壳(图137)。卵子放出两个极体,以后卵子的单倍型胞核同精核(单倍数)结合,从而恢复了染色体的全额。这个卵子称为休眠卵或冬卵,有二倍数目的染色体,不久,卵子发育成母系,重新产生一个新系的单性生殖的雌虫等等。
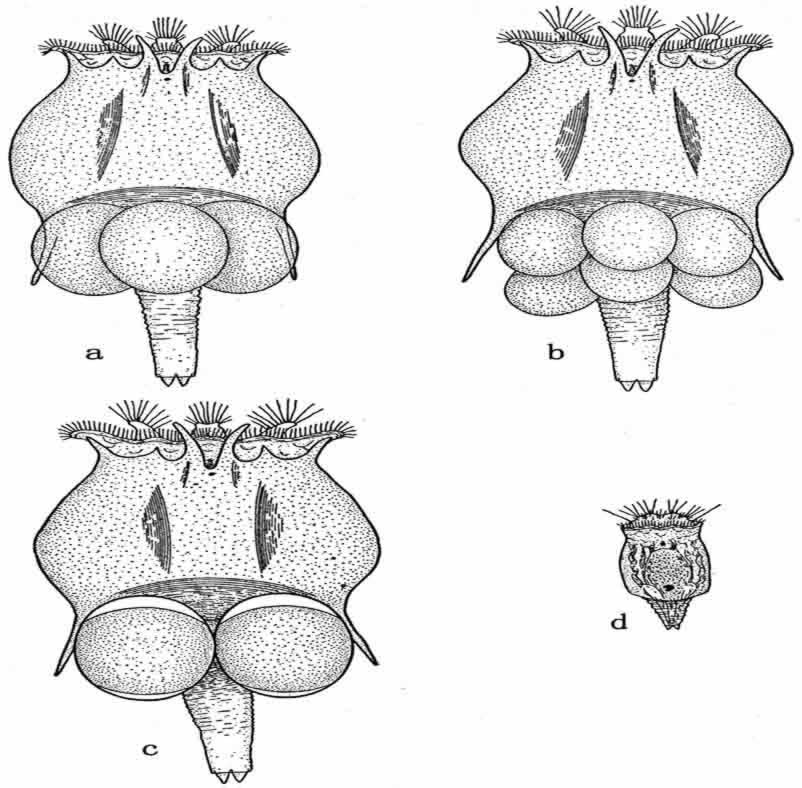
图137 a.臂尾轮虫Brachionus bakeri的雌虫,附有单性生殖的成雌卵子。b.雌虫,附有单性生殖的成雄卵子。c.雌虫,附有成雌和成雄的卵子。d.雄虫(仿Whitney)
另一方面,该雌虫如果没有受精,则其所产生的卵子比普通单性生殖的卵子小。卵内染色体两两接合,放出两个极体。卵内留下一组单倍染色体。卵子分裂,但染色体数目不加倍,终于发育成为雄虫。在单倍体雄虫的精子形成中,究竟发生了什么变化,还不明确。无论是Whitney(1918)的研究,或者是Tauson(1927)的研究,对于这种变化都没有作出确实可信的说明。
从表面上看,以上证据似乎意味着单倍数目的染色体产生雄性。二倍数目的染色体产生雌性。一点也看不出有性染色体的存在,所以不能假定特殊的性基因的存在。即使承认该类基因并不存在,为什么半数的染色体会产生雄性而二倍数目的染色体会产生雌性,依然无法解释,除非认为这里所涉及的分化因子,就是两种卵子内胞质多少和其染色体数目之间的关系。然而这种说法又和蜜蜂(说明如下)一例不合。在蜜蜂里,二倍型卵子发生成雌性,单倍型卵子发育成雄性,但两者却是一样大小。以上两例中突出的事实,是单倍数目的染色体和雄性有关,即令在轮虫内另有决定哪一种卵子成为单倍型的其他因素。
也许可能创立一种性染色体的解释,设想有两种不同的X染色体,又设想在减数分裂中一种进入雄卵的极体,另一种则由有性卵子排出(两种都留在单性生殖的卵子内),但是必须承认,目前还没有理由,也没有必要提出这样的臆想。
蜜蜂和亲缘关系相近的黄蜂、蚁类,其性别决定也同胞核的二倍和单倍状态有关。这是已经确定了的事实,不过还没有得到明确的说明。后蜂在后蜂室内、职蜂室内和雄蜂室内产卵。产前的卵子完全相似。职蜂室和后蜂室内的卵子在产卵时受精,雄蜂室的卵子不受精。所有卵子都放出两个极体。卵核内留下单倍数目的染色体。在受精卵内,精子带来一组单倍染色体,和卵核结合,得出二倍数目。这种卵子发育成雌性(后蜂和职蜂)。后蜂室的幼虫得到丰富的营养,发育完全,成为后蜂。职蜂室内幼虫的食物不同。像上面所说的,雄蜂是单倍体(注:已经知道,当没有受精的雄卵子进行分裂时,每一条染色体断裂为两个部分(形成种系的胞核可能例外)。这一过程似乎不是染色体纵裂成两条染色体,而是横裂成为两片。如果这项解释是正确的话,那么,基因数目实际上并没有增加,这个碎裂或分开过程(在一些线虫里也发现过)对于性别决定机制,也没有说明什么。)。
这里,不能假想性别是由成熟分裂以前的任何影响来决定的。没有证据来肯定卵子内的精核可以影响染色体的成熟分裂方式。也没有证据来肯定环境(雄蜂室或职蜂室)对于发育过程有任何的影响。事实上,这里也没有证据来肯定有任何一组的特殊染色体可以区别为性染色体的。在雌雄两种个体之间,我们所知道的唯一区别,只是染色体数目上的不同。目前我们只能依赖这点关系,把它看成是同性别决定有着某种还未知道的联系。现在这种关系还难于同其他昆虫中性别决定于染色体基因间的平衡一致,但是它仍然可能起源于染色体(基因)和胞质之间的平衡。
还有一件有关蜜蜂中性别决定的事实。在雄蜂生殖细胞成熟分裂中,第一次分裂失败,分出了一个没有染色体的极体(图86)。第二次分裂时染色体分裂,一半进入一个很小的细胞,这个细胞以后退化;另一半则留在大细胞内,这个细胞变成有作用的精子,含单倍数目的染色体。像上面谈过了的,精子带着单倍数目进入卵子,随后卵子发育成雌性。
有几个例子,谈到两族蜜蜂杂交,杂种的后代被记录下来(Newell)。据说孙代雄蜂只表现了原来一个族的性状。如果两族的区别只是同一对染色体上两组基因间的差异,则孙代雄蜂的特点是可以预料得到的,因为这两组基因在减数分裂时,会彼此分离,这一组或那一组会留在卵子里面,以后卵子发育成雄性。但是,如果族间的区别决定于异对染色体上的基因,那么,孙代雄蜂当然没有区分为鲜明的两族的希望了。
职蜂(和职蚁)有时产卵。这种卵子一般发育成雄性,这是意料得到的,因为职蜂不能从雄蜂受精。有人报道在职蚁的卵子中偶尔出现有性类型的雌蚁。可以设想,这是由于卵子保留了双组染色体。据说,在Cape蜜蜂中,经常有职蜂的卵子发育成雌性(后蜂)。我们不妨暂时采用上面所说的同一解释,来说明职蚁中的雌性偶尔产卵,其中有些卵子在特定条件下发育成雌蚁。
研究寄生蜂Habrobracon,已经比较充分地证实了母虫性状直接传给子代单倍体雄虫。普通型有黑眼。在培养中出现了一只橙色眼的突变型雄蜂。和黑眼雌蜂杂交,经过单性生殖得到了415只黑眼雄蜂,又从受精卵得到了383只黑眼雌蜂。
子代的四只雌蜂被隔离后,经过单性生殖,产生了268只黑眼雄蜂和326只橙色眼雄蜂,没有产生雌蜂。
另外有八只F1 雌蜂(从第一只橙色雄蜂受精而来)同子代雄蜂自交。孙代有257只黑眼雄蜂,239只橙色眼雄蜂和425只黑眼雌蜂。
第一只橙色眼突变型雄蜂同它的F1 雌蜂交配,得到221只黑眼雄蜂,243只橙色眼雄蜂,44只黑眼雌蜂,和59只橙色眼雌蜂。
在雄蜂为单倍体,又由未受精的卵子发育而成的假设下,这些结果都是可以预料到的。当杂种母蜂的生殖细胞成熟时,橙色眼基因同黑眼基因分离,半数的配子得黑眼基因,另一半配子得橙色眼基因。任一对染色体上的任一对基因,都将产生同样的结果。
橙色眼雌蜂同黑眼雄蜂交配。从11对杂交中,共得预料到的181只黑眼雌蜂和445只橙色眼雄蜂。另有22对杂交,除了产生816只黑眼雌蜂和889只橙色雄蜂以外,还产生了57只黑眼雄蜂。这一类雄蜂的存在要求另一种不同的解释。它们显然是由同黑眼精子受精的卵子发育出来的。一种可能的解释似乎是:单倍型精核在卵子内发育,而且产生了至少是形成双眼的那些部分。卵子内其他部分的胞核也可能从单组型卵核得来。事实上,有些证据肯定了这是一个正确的解释,因为Whiting已经证明,在这种特殊的黑眼雄蜂里,有些可以繁殖,好像它们的全部染色体只载有母方的橙色基因似的。但是其他事实指明,这里的解释也并不这么简单,因为这些雄蜂大多数都没有生殖力,从有生殖力的雄蜂(嵌合型雄蜂)又产生出了几只雌蜂(注:根据Anna R.Whiting(1925)报道:“黑眼偏父遗传的雄蜂,在形态的畸变上,比正常产生出来的雄蜂和雌蜂,有着更高的百分率。大多数偏父遗传的雄蜂已经被证明是没有生殖力的,有一些作为黑眼来繁殖的有部分的生殖力,此外还有少数嵌合体产生了橙色雌性后代,后代有完全的生殖力。偏父遗传的雄蜂的下一代橙色雌蜂,在形态和孕育性上,都是正常的。偏父遗传的雄蜂的下一代黑眼雌蜂数目少,畸形百分率高,并且几乎完全无生殖力”。Habrobracon的特种雄蜂可以说明蜜蜂里出现的一些不规则的情况。)。不管这些特殊情况最后如何解决,这些杂交的主要结果,证实了雄性是单倍体这个理论。
