第十五讲
内制止与睡眠在物理化学基础上是同一的过程
诸位!在前一讲里,我们达成了一个极为重要的假定,就是在条件刺激影响之下,皮质细胞迟早必定会移行于制止状态,而且在频繁地反复给予刺激的场合,皮质细胞会很迅速地移行于制止状态。最合理的解释一定是这样的,就是可以说,这种皮质细胞是生物个体的警卫点,具有高度的反应性,所以也具有高速的机能损坏性(функциональнаяразрушаемость),极迅速的疲劳性。所以制止过程的发生,并不是皮质细胞本身的疲劳,而是具有皮质细胞保护者的作用,以预防这类特殊细胞继续的、过度危险的消耗。在制止过程的期间,皮质细胞不做任何工作,因此它就恢复正常的成分。全部皮质细胞都是这样的,所以在大多数皮质细胞已经工作的条件下,全部皮质都必须移行于制止状态,这正与条件刺激物对皮质个别细胞发挥作用因而引起个别细胞制止状态是相同的。这就是每天发生的事实,也就是人类和动物的睡眠。与大脑两半球有关的我们25年间的全部研究,继续地、不断地证明了这个结论。现在在大脑两半球生理学的方面,如我们用我们方法所研究的结果,这是一个最确实可靠的命题(положение)。从我们开始研究起,动物的瞌睡和睡眠都是与研究相伴随的,并且直到现在,我们都是不断地与这现象有关系的。这样,在我们研究的各个不同的期间,我们汇集了巨大的事实材料,使我们有理由地做成许多彼此多少不同的若干假定。可是在几年以前,全部的这些假定都互相融合而成为一个与我们全部材料确实地相调和的最后的假定,这是,睡眠与我们所谓内制止过程正是一个同样的过程。
睡眠与这制止过程两者的出现和发展的基本条件是完全相同的。这就是多少时间较长的孤立的条件刺激,或者是多次反复作用的孤立的条件刺激,也就是说,这就是皮质细胞的刺激。在第四、第五、第六和第七各讲里,我们知悉了各种制止过程的一切事例,我们不断地遭遇着动物瞌睡与睡眠的现象。如果我们引起反射的消去,那么,甚至在第一次实验的场合,有些动物,除条件唾液分泌及与此相关的运动反射停止以外,与消去过程发生以前的状态相比较,就显出更高度的疲倦。在几天内重复地进行反射消去实验的场合,即使交替地应用着受过强化手续的刺激物,最后几乎总是动物在实验架台上发生明显的瞌睡和睡眠,而这是该动物以前从来不会有过的现象。在分化相形成的场合,同样的情形更强烈地发生。譬如一只动物具有许多条件刺激物,并且同时其中的一个条件刺激物是一定的音。动物在实验架台上总是保持活泼状态。其次,我们用与该音相近的一个音形成分化相。于是与分化性制止的开始发展同时,瞌睡会出现,并且瞌睡越过越深,最后就陷于深深的睡眠,骨骼肌肉完全弛缓,发出鼾声,结果就是在应用其次各阳性条件刺激物和给予食物的场合,不能不推动该动物,或者甚至于不能不把食物放进于动物的口内,使它开始摄取食物的动作。同样的情形在形成很强的延缓性条件反射的场合(延缓3分钟)也会发生。所以当我们以前关于有些动物发生这样睡眠还很少了解的时候,我们不能够获得我们所需要的条件反射。末了,在形成条件性制止的场合,同样的情形也会发生。需要注意,在内制止各种不同的场合,睡眠的障碍程度是多少不同的。在条件性制止的场合,睡眠显现的程度是最弱的。
这就是一切可能的场合,其时由于条件刺激物不并用无条件刺激物的结果,内制止就会迅速地发展。而且同样的事情也会发生在条件刺激物受着强化手续并多次重复地被应用着,以致具有慢慢地逐渐增强的制止过程的场合。这是在前一讲里特别说明过的一种制止现象。在几个月或几年实验的经过之中,这种情形最后的结果就产生各种不同移行状态,如觉醒状态与睡眠状态间(次篇讨论这个问题)及与完全睡眠间等等的移行状态,这是因动物而有差异的。动物在这一关系上是彼此互不相同的,正如制止过程发生的速度的各有差异。
我们有关条件反射的研究充满着极多的由制止过程移行于睡眠的事例,所以不需要举出多余的个别的实例。这样看来,实际上制止过程是与睡眠有密切关系的。如果不应用适当的办法,制止过程就移行于睡眠。
在一定条件之下,差不多没有一种刺激物不受内制止和睡眠的影响,这是富于兴趣的。在叶洛菲耶娃实验里,应用了强有力的电流于皮肤当做条件性食物刺激物。虽然对这条件反射施行了强化手续,但在实验经过几个月之后,终于显现了逐渐增强的制止过程,而在彼特洛娃所做的同一实验里,同样的电流刺激却显然地引起了睡眠。从另一面说,当做条件刺激物的各种不同的外来动因,在内制止发展速度的关系上与在睡眠发展速度上次序完全相同。在前章讲义内已经说明过,制止过程在温度刺激的场合最容易发展,在声音性刺激的场合最困难。严格地与此相并行地,睡眠在温度性条件反射的场合发生最快,而在声音性条件反射的场合却是又慢又较少。在温度性条件刺激的场合,睡眠所引起的障碍非常顽固,妨碍了我们的研究,所以在我们研究的初期,很难获得应用这些温度条件刺激物做实验的工作同人的赞成。
末了,如以前指出过的,条件刺激时间的长短对于制止过程和睡眠的发生,具有决定性的意义。在条件刺激物延缓时间是10~15秒的场合,在长期的研究期间,有些实验狗保持完全觉醒的状态。只要将狗的条件刺激延缓到30秒以上,瞌睡和睡眠即刻就会出现。这样方式的实验往往是实在令人惊异的。在实验条件不过具有似乎意义很小的这类变动的场合,实验动物由完全觉醒状态这样迅速地移行于真正的睡眠,这是完全出乎意料的。在我们的研究里有多数这样的事例,不过由于条件刺激物延缓时间的不同,表现的程度也各有不同。
在长时期内条件反射被应用着并不断地受强化处理的场合,会有逐渐增强的制止过程的发生。在前讲里引用了许多处理方法,既可以阻止这类制止过程的发生,也可以把制止过程除去。没有例外地,这些一切的方法同样地也能克服睡眠。
在刚才所引用的一切材料之后,会发生一个极当然的问题。既然睡眠的出现与消失都与内制止互相一致,那么,内制止怎样才可能是觉醒状态最重要的因素呢,怎样地才可能是生物个体保持与环境的平衡的基础呢?初看起来,这些问题的答复是似乎困难的。据我的意见,以前在各篇所说明的事实必定能够完全排除在答复方面的这个困难。在觉醒状态的内制止过程是一种微细地碎分的睡眠(раздробленныйсон),是皮质细胞个别群的睡眠,而睡眠却是一种扩展性的内制止,散布于全部的脑实质与大脑下位的各部分。所以事情是这样的,就是制止过程在空间上受着限制的,是封闭于一定的范围里的。而引起这样情形的,当然是互相对立的神经过程,正如我们在有关大脑皮质镶嵌细工式及有关皮质分析性机能的讲义内所看见的事实一般。
在反射消去的实验场合,只是在反射消去过程成立以后,我们系统的应用依旧受强化处理的各条件刺激物,并且重复应用消去性反射的回数不太多,那么,睡眠才不会发生。在分化相的场合,已展开的制止过程,起初并有睡眠,可是只在如下的情形下才能保持所需要的制止过程而不致有不适当的睡眠,就是与使用未受强化处理的动因同时,需要交替地、并且比通常更多次地应用条件刺激物,即是应用具有阳性作用的动因,这就是说,兴奋过程不断地抵抗制止过程的扩展。在条件性制止和延缓性反射的场合,情形也是相同。在一切场合,如果实验的执行是合理的,瞌睡与睡眠只是一种时相性现象(фазовоеявление),其时在兴奋过程与制止过程两区域之间还不曾有精确界限的成立。如果实验条件使制止占优势,睡眠就发生。鲜明的例子如下。在关于镶嵌式的讲义内我提及了一只狗的实验。对于这只狗,用一定音形成了阳性条件反射,并且与该音上方及下方相近的20个音是分化的,就是说,这20个音是阴性条件刺激物。在阳性和阴性刺激物的应用上有一定关系的场合,一般地说,这只狗并不倾向于睡眠,并且总是对于一切阳性刺激物都显出完全的效力。但是只要接连几次地发动分化音的作用,这只狗就陷于很深的睡眠,虽然应用其他强有力的新异动因,也不能使它觉醒。相反地,如果把阴性条件刺激交替地与阳性条件刺激共用,那么,即使频繁地应用阴性条件刺激,睡眠也并不发生。在这一关系上,前述有关镶嵌式的实验,尤其有关条件皮肤机械性镶嵌式的实验是值得重视的。虽然条件性皮肤机械性刺激很有引起睡眠和瞌睡的倾向,可是库帕洛夫对于一只实验狗,2年以上,只应用了皮肤机械性刺激,这只狗丝毫不会显出睡眠的倾向。这明明是因为制止过程对于兴奋过程不断地保持一定的界限,因而制止过程由兴奋过程而限制于狭隘范围以内的缘故。对于广泛地扩展的制止过程,我们还有对抗的方法,就是把阳性条件刺激物增多,以限制制止过程从发端点向近旁部的广泛扩展。从彼特洛娃的研究,举出一个更复杂的一部分与此有关的例子吧。对于一只狗,即刻着手于很强延缓的(延缓3分钟)拍节机食物反射的形成。这只狗很快地开始陷入于瞌睡状态,最后就完全睡眠。很显然,在拍节机作用的第一期,因为拍节机刺激与无条件刺激物相结合的瞬间相隔过远,其时必然发展的制止过程占了优势,这是因为正在无条件刺激物作用直接以前的时期,即在拍节机作用的第二期,并不曾有兴奋过程的适当的发展,所以发展中的制止过程不受任何的抵抗作用而占着优势。于是再应用了五个新的动因而形成了条件刺激物,并且在给予这些刺激物以后都经过5秒钟就给予食物。瞌睡状态就很快地消失了,并且5个反射全部都是很容易形成的。在这以后,又渐渐地把给予无条件刺激物与条件刺激物的间隔时程加长,每日延缓5秒。与此相当地,各条件反射的潜在时间也逐渐加长,并且在最后,没有任何睡眠的妨碍,形成了6个条件反射,其潜在时间是1~1.5分钟,这就是一种初期的制止过程的表现。这样地,兴奋过程先在大脑皮质6个点内发生,只渐渐地把地位让给制止过程,使制止过程在时间上与空间上都受着限制,睡眠就不能发生。
在实验架台上限制动物的运动,这对于若干的狗,尤其对于具有瞌睡倾向和制止过程倾向的少数狗,必有影响。在这样场合,有时把狗放在地面上,至少暂时地使它获得自由以进行实验,这是很适当的。我们需要这样想,就是在运动的时候,由运动器和皮肤两方面出发的刺激,必定会在大脑两半球皮质内陆续地形成兴奋的中心,这也对于制止过程的扩展是相当对抗的。不过此时还有另一个因素,似乎具有巨大的意义,关于此点以后当有说明。
我们已经看见,在重复地应用条件刺激的场合,在大脑皮质细胞内就会有制止过程的发生。与此相同地,如果重复地应用一切并无特殊条件性生理学意义的各动因,以刺激脑皮质细胞,制止过程也会发生。像以前曾经提及过的,在各种反射之中,有一种探索反射。这个反射的作用点是在大脑两半球和下位脑部的细胞之内都有的。在大脑两半球现存的场合,探索反射显然是由大脑两半球细胞的参与而成立的。无疑地,这是由于本反射的精确性而能证明的,因为在环境有任何极微小摇动的场合,探索反射也会发生,这只是在大脑两半球最高的分析机能存在的场合才是可能的,而绝不是脑下位部的关系。像我们都知道的,在重复地被应用的场合,探索反射必定会减弱,并且纵然唤起探索反射的动因仍然继续存在着,最后探索反射会完全消失。在我们研究室里柏柏夫(Н.А.Попов)的特殊实验证明了,探索反射的消失是由于制止过程的发展,并且在详细的经过上,这与条件反射的消去完全相同。
如果在一个实验的期间以内,以一定短的间隔时程反复应用某一个动因,起初会引起探索反射,以后却不能再引起与此相当的运动反应,那么,在同一个实验内安排较大的间隔时程以后,该运动反应又会恢复,这正是与消去性条件反射的场合相同的。在一个实验之中,因为重复应用的结果,某个一定的探索反射会消失,但是如果在它消失以后,即刻应用另一个新异刺激物,探索反射就暂时会出现,这就是说,另一个探索反射被引起了。所以,探索反射与被抑制的条件反射相同地,也是会解除制止化的。如果在几天以内重复地应用某一定动因的探索反射,那么,这探索反射就会慢慢地消失,这是与系统地不受强化手续处理的条件反射相同的。末了,已经消失的探索反射在兴奋剂影响之下(咖啡因),会暂时又再恢复,这与在条件性分化性动因的场合所观察的情形相同。探索反射的这个制止过程必定地,并且比条件反射的制止过程甚至更易引起瞌睡与睡眠。现在从契求林(С.И.Чечулин)的实验举出例子,说明在探索反射的场合,制止过程和睡眠的发生是与条件刺激物的应用相结合的。
对于一只狗,用哨笛音形成了食物性条件反射。当做探索反射的刺激物而开始被使用的动因是:嘘音,水泡音,皮肤机械性刺激及其他等等。

以后,从第21分钟开始,应用水泡音30秒钟,每隔2分钟应用一次。在起初三次应用的时候,动物向着发声的方向运动,不过运动是渐渐减弱的。从第四次应用的时候起,瞌睡出现。直到第八次应用的期间以前,睡眠因为各种不同的刺激因素而时时中断。在第八次和第九次应用的时候,狗的睡眠是连续的,不因为刺激物而有任何运动。在43分钟的时候,起初只应用水泡音10秒钟,其次再应用哨笛音30秒钟。没有任何反应——运动的和分泌的反应都没有,睡眠继续着。食物的给与会使狗觉醒,狗开始吃,可是吃了以后依然瞌睡。用条件刺激物所做的实验继续进行如下。
在这实验以前,狗在实验架台上从来不曾睡眠过。在以下的实验里,引起探索反射的一些新的动因重复地被应用着,直到睡眠出现的时候为止,或者直到运动消失而瞌睡尚未出现的以前的时候为止。在上述实验以后过21天,用皮肤机械性刺激做实验。实验的进行如下。

从第25分钟开始皮肤机械性刺激,每次继续30秒钟,每隔1分钟,重复地应用一次。在起初三次应用的时候,狗头运动是倾向于受刺激部位的一侧的。在第四次和第五次应用的时候,不再有运动。在第32.5分钟的时候,只应用皮肤机械性刺激10秒,其次,哨笛音被结合而应用10秒,在哨笛音开始以后过15秒,唾液分泌开始,并且在哨笛音的30秒内有唾液2滴的分泌。以后实验的进行如下。

在实验开始的时候,条件刺激物或者在5秒钟以后被强化,或者在30秒以后被强化,目的是要保持条件刺激物的正常值,一直到实验终结的时候。
我们看见,如果把引起探索反射的动因重复地应用下去,它的运动效果就总会慢慢地减少下去,如果再继续重复地应用下去,那么,有时睡眠与瞌睡会即刻发生,越过越加深,或者在睡眠或瞌睡的出现以前,动因暂时似乎依然没有任何作用。然而与该动因相结合的条件刺激物(如刚才所引用的第二个实验)证明着,这其时似乎没有任何作用的该动因却是引起制止作用的。对于条件刺激物的这种制止作用并非由于我们所谓外制止作用的机制而成立的。这是可以这样证明的,就是(像在第六讲研究有关探索反射对延缓性条件反射所发生的作用的场合)探索反射如果是微弱的,它就只能使条件反射解除制止化,而不能制止条件反射。所以与现在所说的事例有关的是制止过程的作用,而这制止过程的发展是探索反射重复地被应用的结果,并且以后这制止过程会移行于睡眠,其时条件反射也会完全消失(刚才所引用的实验中的第一个)。
我们在用幼犬做实验的场合,同一的情形会特别显著地发生[洛仁他里(И.С.Розенталь)实验]。在任何刺激物单调重复地被应用的场合,在周围环境一般地没有任何动摇的场合,幼犬们往往非常正确地迅速地陷于睡眠。从另一面说,一切的人,尤其不具有坚强的内在生活的人,在受单调性刺激的场合,就不管什么地点和什么时候,不可克服地会陷于瞌睡和睡眠。这虽然直到现在不曾有过科学性的说明,但是谁都知道的一个事实。这就意味着,对于长时间继续作用的外方动因而发生反应的一定的、一些大脑皮质细胞,不断地消耗着,就移行于制止状态,如果其时从皮质其他各活动点不发生与这制止过程相反的作用,制止过程就更加扩展而成为引起睡眠的条件。大脑皮质细胞的非常迅速的消耗性(истошаемость),容易移行于制止状态,而与此强烈地相对照的是,大脑下位部的细胞在同一条件之下却具有很大的忍耐性。我们研究所的泽廖尼(Зеленый)的实验证明过,在正常狗的场合,对于一定声音的探索反射会迅速地消失,但是在狗的大脑两半球被除去的场合,在同一环境里用同一的声音做实验,这个探索反射会常同型地(стереотипно)发生极多次。
我们再回到条件反射的问题上去。
大脑皮质细胞的消耗和一般地大脑皮质细胞的疲弱,会成为制止过程发展的基础,并且会成为睡眠在制止过程以后出现的基础,这是由于如下的常见的事实而可以证明的。如果我们对大脑两半球施行外科的手术而损伤了某一个分析器,那么,与该分析器有关的各阳性条件刺激物的单独应用,或者甚至于在短的时间内也是不可能的,这些阳性刺激物会迅速地变成阴性刺激物——或者各阳性刺激物会完全失去阳性作用,而成为阴性制止性的刺激物。在皮肤分析器受伤的场合,这是特别容易常常观察的事情。
当一只狗的大脑皮质的冠状回转(guri coronarius)和外雪儿维氏回转(ectosylvius)摘除以后,在相当的时期以内(往往好几周),狗的四肢、肩、腰等部位的阳性条件性皮肤机械性反射都会消失而为阴性制止性反射所代替。这情形是可以这样证明的,就是在应用皮肤机械性刺激以前,其他分析器有关的一些阳性条件刺激物,具有显著的阳性作用,但在皮肤机械性刺激以后却丧失阳性作用。同时,这些皮肤机械性刺激非常容易地迅速地引起这些狗的睡眠,而这些狗在受上述手术以前,从来不曾在实验的时间以内睡眠过。事实是往往采取如下可惊异的形式的。如果对相当于脑部手术损伤的皮肤表面某一个部位应用条件性皮肤机械性刺激物,就会引起制止过程和瞌睡,而同一刺激物如果被应用于不因脑手术而受损伤的皮肤部位,就会显出阳性效力,并且使动物依然保持活泼觉醒的状态[克拉斯诺高尔斯基(Н.И.Красногорский)、拉仁可夫(И.П.Разенков)、阿尔汉格里斯基(В.М.Архангельский)等三人的实验]。
可以很正当地在此地引用的一个事实,是在几年前的我们饥馑时代实验室内所观察的事实。对于衰弱疲惫的动物,我们不曾能够做条件反射的研究,因为一切的各阳性条件刺激物对于动物都非常迅速地变成阴性刺激物,并且同时一应用条件刺激物,动物就开始睡眠。很显然,动物全身的疲惫对于大脑皮质细胞特别发生了影响(弗洛洛夫、洛仁他里等人的实验)。
在直到现在已列举的很多数的场合,我们不断地看见,制止过程会移行于睡眠,可是也可以观察相反的情形——就是睡眠会移行于制止过程。我们形成过延缓3分钟的条件反射。个别实验有时是进行如下的。我们把动物放在实验架台上。动物是很活泼的。但是条件刺激物一被应用,动物即刻就瞌睡,并且在3分钟里,唾液分泌效力完全缺乏着;而且在给与食物的场合,动物并不即刻就吃,而是懒慢地吃。我们在这个实验里,以通常的间隔时程再应用这条件刺激物若干次。与每一次的应用并行地,动物在受刺激的场合越加活泼,在3分钟刺激的末了,唾液分泌就出现了。与刺激物重复应用相并行地,唾液分泌越加增多。于是大约可以把3分钟的刺激分为两个时期:在前半期,动物虽然完全是活泼的,但无唾液分泌,而在后半期,唾液分泌极多,并且动物迅速地很贪馋地吃食物。这就是因为起初在条件刺激物作用前半期,制止过程占着优势,发生了扩展性的制止过程,就是说,睡眠发生了,而这扩展性的制止过程会渐渐地移行于局限性的集中性的制止过程,这是与该同一条件刺激物作用后半期的兴奋过程逐渐增强的压力有关的。在与此类似的一些场合,也不能不看见制止过程为纯粹的睡眠所代替的情形。既在较长的延缓3分钟的场合,也在刺激较短的30秒钟的场合,都有时可以观察如下的情形。在实验的时候,实验台上的动物虽然总是觉醒的,但在正确地重复地被应用的条件刺激物每次开始作用的时候,动物就睡着:眼睛闭着,头部下垂,全身弛缓地悬在粗绳上,有时甚至于可以听见鼾声。可是经过一定时间以后,就是在刺激延缓较长的场合过1.5~2分钟,在刺激延缓较短的场合过25秒钟以后,动物迅速地醒来,唾液分泌也出现。并且显出强烈的食物性运动反应。很显然,在这场合,集中化的制止过程是慢慢地为弥漫性睡眠所代替的。最后,有时可以看见,两个制止性刺激的同时作用会引起瞌睡。
一只狗(弗尔西柯夫实验),具有良好形成的条件反射,孤立的刺激时间3分钟。在最初的2分钟,没有唾液的分泌,2分钟后,唾液分泌开始,在第三分钟的终末达到最大数值。在这实验里,与条件刺激物同时应用了另一个新异动因——微弱的嘘音(шипение)。这嘘音能消除条件反射前半期的制止过程,并且在应用嘘音的时候可以看出微弱的运动反应——探索反射。条件反射被强化了。在重复地应用这复合刺激物的场合,对于嘘音不再有方位判定反应的出现,而条件反射完全消失,动物非常瞌睡。反射再受强化处置。显然地,结果是应该解释以下的。对于嘘音的探索反射,在新刺激物嘘音第二次被应用时候已经消失了,就是说,现在对于嘘音发展着制止过程,这是在本讲的前部已经提及过的。这制止过程与延缓性反射第一时相的制止过程互相连接,就把制止过程更加增强,于是反射的活动性时相完全缺乏而让瞌睡占着优势。我们继续做这个实验,就证明与这实验有关的这解释是正确的。在条件刺激继续地反复地被应用着而不并用嘘音的时候,正确延缓性反射就显现出来,并且有两个时相。如果再用嘘音与拍节机配合起来做一次实验,就再引起瞌睡,条件反射会消失。
此实验的具体数据如下。
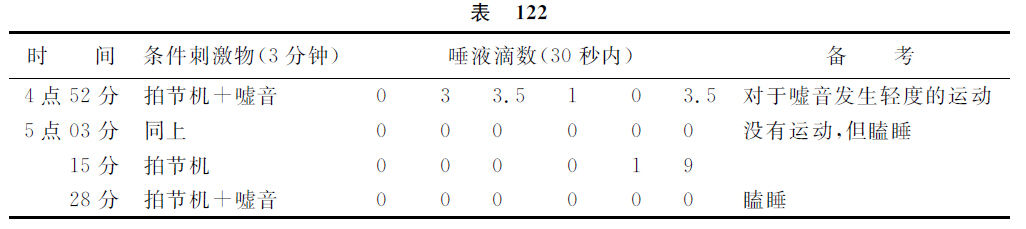
在这一场合,为了明了起见,请诸位注意于如下的情形,我认为这不是徒然的。显然,这个实验和上述契求林的实验,在新异动因对于条件反射所发生作用的关系上,增加了一个多余的新的时相。如果应用强有力的外来动因,那么,它的作用如何,请你们想起第六讲内延缓性反射有关的说明吧。就是强力的外来动因由于所引起的探索反射,起先抑制延缓性反射的全部,其次在重复地给予强力新动因而探索反射显著地减弱的时候,该新动因就仅能解除延缓性反射第一时相的制止过程,但是到了最后,就如诸位现在所看见的,该强有力的新动因又制止这延缓性反射,而此时的制止作用是另有一个根据的,就是这强有力的刺激本身此时在大脑皮质内成为制止过程的原发性刺激物了。而弱的外来动因,正如在弗尔西柯夫的实验里所表现的,从最初起就引起微弱的短时间继续的探索反射,在第一次被应用的场合就对于延缓性反射具有解除制止的作用,最后就以第二次的制止过程而完结。
制止过程与睡眠两者共有的特性,也说明这两种过程的相同性。在前述的讲义内,我们看见了极多的事实,毫无疑问地证明制止过程向大脑两半球实质的进行,并且此过程进行得很缓慢,计算起来有几分钟之久或者甚至于是较多的几分钟,而且此外,由于各只动物的不同和实验条件的不同,这过程进行的速度也很有差异。很显然,睡眠也是一种运动性的过程。我们都知道,瞌睡与睡眠对我们的侵袭是逐渐的,并且是慢慢地相当困难地才会消失散去的。我们也有科学的参考资料,说明在睡眠的场合,各种感觉器机能和其他更复杂的精神机能的消失都是逐渐进行的。从另一方面说,各人的睡眠与觉醒的速度具有很大的区别,而在不同的条件之下,这个速度也各有区别,这都是周知的事实。在实验狗的场合,我们也观察了同样的情形。
此外,在我们的研究方面,我们不断地看见,制止过程的发展是起初很困难的,但是由于实习,即由于制止过程各种形成方法多次的应用,就会逐渐成为很容易再演成的一种过程。完全与此相同地,各条件刺激物在相当的条件之下会引起睡眠,并且无关系的刺激物,如果重复地被应用的回数越多,就会越迅速地引起睡眠。
如下的事实是特别有趣的。像以前指明过的,制止过程能诱导兴奋过程。像在稍稍以前说明过的,在若干狗延缓性反射的场合,条件刺激物在刺激的初期不引起制止过程而引起睡眠,有时在睡眠以前,动物先有极短时间的若干一般性的兴奋。在无关性刺激物长时间被使用的影响下或重复多次地被应用的影响下动物睡眠的场合,上述现象是更鲜明而恒常的。在上述的洛仁他里的实验里,这是常常被观察过的。当无关性刺激物显然开始刺激而幼犬发生瞌睡状态的时候,在幼犬最后入睡之前,先有极短时间的兴奋状态,它开始不安地运动着,用爪挠动,无故地向空中吠叫。同样的情形,我们在小孩睡眠的时候,也常常可以看见。这是一种特异的出乎意料的情形。可以有理由地把这种情形当做一种诱导现象看待。可以同样解释的是,这是在麻醉药作用开始以前的周知的兴奋期。
我以为,上文所引证的各种事实的全体可以充足地完全地证明我们一个假定的正确性,就是睡眠与内制止过程是一个相同的过程。现在我本人不知道,也不能看出,我们的研究有任何一个事实是会与这个结论严重地相矛盾的。不能不以为遗憾的是,直到现在,我们还没有表现睡眠的好的描写方法。不过间或地我们为了这个目的利用了动物头部位置的记载方法。当然,我们有关睡眠各实验的报告,如果并用睡眠的某种表现,也许就会更增加我们理论根据的确实性。
显然,我们通常睡眠的一切详细的情形也与我们的这个结论完全调和。我们有些人的每日工作是非常单调的,有些人的工作,相反地,是极复杂的,可是最后都同样地成为睡眠发生的条件。皮质的同一部分的各点,如果受了长时期的刺激,就会在这些点部引起深深的制止过程,而这制止过程当然强烈地扩展而侵袭大脑两半球,下降到大脑下位的各部分。从另一侧说,在复杂活动的场合,虽然皮质的个别各点并不曾达到显著程度的制止状态,可是这些刺激点数量极多,即使没有大量的扩展过程,也会构成广泛的制止状态,也下降到脑下位的各部分。当然,非常大量的、迅速地轮流交替的许多刺激,往往可能长时间地反抗制止过程对大脑两半球的侵袭,因而使睡眠出现很迟。并且与此相反地,如果在觉醒与睡眠两过程之间有严格地轮流交替的生活顺序,有一个确定的节奏,就可以增大睡眠的坚强性,而并不必有大脑皮质细胞过度的疲劳。在我们的实验里,有足够的例子可以说明,兴奋过程与制止过程彼此间的关系也与上述情形相同。
