- 下篇
- 【1】 ">脓细胞的化学成分 【1】
- 【3】 ">对了解细胞及其生命现象的贡献 【3】
- 【5】 ">卵成熟和受精的研究 【5】
- 【7】 ">群体遗传与纯系 【7】
- 【11】 ">染色体遗传 【11】
- 【20】 ">昆虫染色体与性决定的关系 【20】
- 【24】 ">细胞核的化学成分 【24】
- 【26】 ">孟德尔遗传的随机分离与相引 【26】
- 【28】 ">果蝇的六个性连锁因子由其联合方式所表示的直线排列 【28】
- 【32】 ">性别对染色体和基因的关系 【32】
- 【35】 ">基因的人工蜕变 【35】
- 【37】 ">人类血液的个别差异 【37】
- 【40】 ">玉米细胞学和遗传学交换的关系 【40】
- 【43】 ">一种研究染色体重排和绘制染色体图的新方法 【43】
- 【45】 ">链孢霉生物化学反应的遗传控制 【45】
- 【50】 ">对引起肺炎球菌类型转化的物质化学特性的研究 【50】
- 【56】 【57】 ">在细菌生物化学突变型混合培养物中出现的新基因型 【56】 【57】
- 【59】 ">沙门氏菌的遗传交换 【59】
- 【61】 ">噬菌体生长过程中蛋白质和核酸各自的功能 【61】
- 【64】 ">核酸的分子结构 【64】
- 【66】 ">脱氧核糖核酸的结构 【66】
- 【69】 ">基因 【69】
- 【71】 ">细菌噬菌体遗传区的精细结构 【71】
- 【73】 ">和核酸相似的多核苷酸的酶促合成 【73】
- 【78】 ">病毒重建 【78】
- 【82】 ">脱氧核糖核酸的生物合成 【82】
- 【84】 ">蛋白质遗传密码的一般性质 【84】
- 【86】 ">大肠杆菌无细胞系统蛋白质合成对天然或合成多聚核糖核苷酸的依赖作用 【86】
- 【88】 ">蛋白质合成的遗传调节机制 【88】
- 【98】 ">核酸的分子构型 【98】
- 【100】 ">噬菌体、大肠杆菌素和大调节现象 【100】
- 【103】 ">核糖核酸指导的脱氧核糖核酸合成 【103】
下篇
自1900年孟德尔定律被欧洲三位科学家重新发现后,才诞生了现代意义上的遗传学。此后历经细胞遗传学、微生物遗传学、分子遗传学等不同的发展阶段,其发展极为迅猛,至今方兴未艾。

孟德尔所在的修道院
脓细胞的化学成分 【1】
约翰·米舍尔(F. Miescher) 【2】
(1871年)
直到目前为止,脓化学几乎完全是从病理学的角度来研究的。最近也利用脓来研究原生质特性。这个材料是不完整的,而且使用必须小心,但它是唯一容易得到的材料,为此,作为一个起点,它是合适的。
由图宾根外科医院供给研究材料。从包扎伤口的敷料上收集脓液,并立即使用。废弃那些从外形和气味说明已进一步解体的材料。我所处理的材料在数量上常有变动,但这种变动很少超过两盎司重,常常是略有变动。
显然,我没有用生理上新鲜的,也就是说活的脓细胞。因此,本人的试验结果需要进行专门的观察予以校正。
本研究首先迫切要求把细胞从血清中分离出来。通过过滤常得到稍微清洁的血清,但通常这只是赠品的一部分,盐溶液沉降用在血球上卓有成效,但此处未见成功,因为整个细胞团膨胀成发黏的东西,即使变换各种浓度也是这样。为此,我改用了其他的盐溶液。经过试用各种碱盐和碱面,最后确定了一种混合物:一份Glauber盐(硫酸钠)饱和溶液和九份水。用这种溶液冲洗吸有脓液的敷料,这样在清晰过滤的脓血浆中不产生一点混浊。用亚麻布过滤液体以除去棉花纤维。大多数细胞很快地从这个液体沉淀出来,即常常在一两个小时以后,能从浆状沉淀物中撇出一种混浊液。通常在一天内反复冲洗两三次。通过过滤把大多数冲洗液从脓细胞黏液中除掉。所得到的细胞在显微镜下的样子为球形,略膨胀鼓起,色暗淡,好的材料没有一点解体的迹象。
核和核素
像现在这样具有纯细胞的材料,特别需要解决细胞核的化学成分问题。我在别处提到,用很稀的苏打(碳酸钠)溶液提取细胞时,人们可以在其他东西中得到一种溶液加酸沉淀的物质,这种物质既不溶于过量的酸,也不溶于盐,但加上微量的苛性碱或碳酸碱,它就溶化了。同已知的组织化学事实相符,这一材料非常可能是细胞核。然而,我用稀释的酸想令人满意地把它从联结在一起的蛋白质里分离出来,却未获成功。仍有难以掌握的不能过滤的混浊度。所以我试图把细胞核本身分离出来。
为此,我首先使用了很稀的盐酸,延长其作用时期以溶解原生质,留下裸露核,但结果不完全,处理数天后,有些核几乎总是被分离出来,有时数目还不少;但多数情况下一些原生质顽固地同核粘在一起,即使更换液体6~10次也是如此,而且酸只吸收一点蛋白质。因此,没有溶解的原生质残余物沉淀是不完整的,且过滤冗长费时,醋酸的结果甚至更差。
用一种更机械的方法,以稀盐酸处理细胞数周(严冬条件下),得到了少量核。我用乙醚和水长时间有力地摇晃这些没有溶解的残余物。细胞团仍然含有在两个液体之间界面上所收集到的原生质残余物。然而,过一段时间在水层底部看到细粉末沉淀。可用一过滤器把它收集起来。它完全是纯核,外形光滑,含有物均匀一致,且明显地分出核仁。比它们原有体积略小。用新鲜水摇晃,可从剩下的细胞中重新取得核,但数量总是很少。核的比重高于原生质的比重,可依此作为这一分离方法的基础。
所得到的核在纯水中完全没有改变,但在非常稀的碱液中就变得膨胀,色浅。核仁颜色也变浅,看不清。添加酸后可恢复原来的结构样子。核在NaCl溶液中也有点膨胀,碘染色明显地呈黄色。上述稀苏打溶液从核中萃取出一种黄色溶液物质,当它被稀释的醋酸或盐酸处理后,产生出不溶于过量酸的簇毛状沉淀物。这些沉淀物在纯水中丝毫不膨胀,但如在煮沸时在仍然清洁的液体中加一点点苛性碱或碳酸碱以及普通的磷酸氢钠就溶解了,加NaCl则不溶解。加硝酸产生黄蛋白反应,尽管仔细漂洗也仍然如此;加氢氧化钠和硫酸铜产生蓝紫色溶液。它可溶于强盐酸,在稀释液中产生的沉淀即使加大量水也不溶解。从而证明该物质本身与蛋白质有关,但不是蛋白质的一个成员。除在中性水中完全不膨胀或溶解外,总的说来同艾希瓦尔德(Eichwald)的黏蛋白是一致的。
可以在滤器上得到这样一种物质,这种物质即使在浓苏打溶液中也不溶解。用酒精和乙醚干燥后,它像一棉胶似的薄片从滤器上顶出。在显微镜下它仍然清楚地呈现出核及其核仁的轮廓。该薄片可溶于浓HCl和苛性碱中,尽管不是顷刻发生,但另一方面,即使把它放在一个密闭的玻璃管中用冰醋酸加热到140℃达数小时,却仍然无变化(与角蛋白物质形成对照)。根据这些溶解特性,大体上有点像弹性物质。用上述方法得到了少量核,仅满足于上述少数反应,作元素分析是毫无问题的。
所以,我抓住了一种方法,这种方法早就使用于蛋白化学,即含有胃蛋白酶的液体,对蛋白质的溶解是有效的。把猪胃提取液每一升水加10毫升强盐酸过滤澄清。在40℃条件下,用这种液体直接处理刚漂洗过的脓细胞没有得到令人满意的结果。多数材料溶解了,但释放出一定量的油滴,这可能是卵磷脂解体的部分结果,这使得未溶解的残余物悬浮成难以过滤的混浊液。因此我延长用温酒精处理的消化时间,通常为3~4倍,然后处理残余物,现在已差不多没有卵磷脂了,在37~45℃条件下消化。在几小时内,从清净的黄色液体中分离出细小粉末状的灰色沉淀。这肯定是一个完全的反应。消化18~24小时,在此期间两次倒出和更换液体。第二次提取后,沉淀物在数量或显微特性方面不再有变化。沉淀物只有分离出来的核,没有一点点原生质残留物。有时候混杂几个细小、中等折射的颗粒,但多数颗粒穿过滤器被洗掉。如酒精提取没有用尽,会看到一些油滴。沉淀物加新鲜乙醚反复摇晃以除去脂肪。最后一次倒出乙醚后,很容易在滤器上收集到黏土般的灰色团的核,这时用水随意冲洗不会产生任何改变。冲洗持续到丹宁不再使滤液混浊为止。
一旦弄清楚必须注意的要点,我完全担保用这种方法能从脓细胞得到数量理想的核。得到的核完全是裸露的,但至少大多数不像单用盐酸分离出来的核那样光滑。虽然这些核在大小上与上述方法制备得到的核无显著差别,但它们常常有点皱缩,显然被折射了的样子,有的看起来好像它们的膜厚度不等,或出现球形模糊的样子。这是否因为内含物的改变,还是表面的皱缩和变粗糙?这一点尚未能肯定。有些核的轮廓光滑,其他一些核看来被腐蚀了。球形不明显的核,其核仁清晰可见。然后用温酒精把这种方法得到的经过漂洗的细胞团处理几次。酒精提取出少量在蒸发时呈油状和浅棕色的物质。它在乙醚中慢慢地溶解,留下有点发脆的残留物。根据这些特性,它很像卵磷脂;遗憾的是忘了用磷作分析。通过第三次提取,就不值得再去追踪该物质了。
经过纯化的细胞核团,除了其显微情况外,像是用稀HCl分离出来的核。用稀苏打溶液产生黄色液体,可以用醋酸或HCl从这个液体沉淀出一种不溶于过量酸的沉淀物。酸性滤液不论是中和还是添加亚铁氰化钾都不会变混浊。这物质的主要部分还是不溶解的,但它慢慢地溶于苛性碱。从单用HCl分离出来核的相当类似行为来看,不溶性并非是酒精或煮沸加热的问题。相反,我相信显微形状的改变可能是用酒精提取一种物质,即卵磷脂的结果。有些核混浊度不一;这可能因核的发育时期不同,而提取材料的量就有所不同。
溶于苏打溶液的材料,像上述用HCl分离核那样,发生类似粘蛋白的反应。但我不能得到更多的量,过滤器填满了膨胀的残留物,而如延长过滤进程,溶液中材料就会变质,形成的产物可能是被丹宁而不是被醋酸沉淀。我用少量材料作氮的测定。所以后期试验我只用整个核,而把更好的材料留下作组分分离,这里我暂时把它称为可溶性核素和不溶性核素。
纯化的核,尽管在浓HCl中不是顷刻间溶化,但它完全是可溶性的。如试剂作用时间不长,然后用水稀释,几乎全部材料都再一次下沉为毛丛状沉淀物,加大量的水也不溶解,然而,用亚铁氰化钾处理和中和,滤液产生一点点沉淀,丹宁使后一种滤液有点混浊。再延长作用时期,这种转化产物的数量就增多,到最后不论是稀释还是添加亚铁氰化钾都不产生沉淀。多数沉淀仍然是用丹宁处理形成的。而溶液常常是紫红色。
苛性碱的效应相似,它完全溶解核。首先,当用HCl或醋酸进行酸化,几乎什么都再次沉淀,但这种沉淀很容易在最稀的苏打溶液中溶解。我据此作出假设:可溶性和不溶性核素无本质差别,它可能只是一些变更,却易于从一种转换成另一种。当然,这一假设尚需进一步验证。同样,酸性滤液进行中和或添加亚铁氰化钾变得混浊。用相当稀的氢氧化钠处理数天后,这种中性溶液形成大量沉淀,此沉淀几乎完全溶解于1/1000HCl和稀醋酸中;但丹宁也会使这种中性滤液变混浊。这证明上述类似白蛋白的反应未必是夹杂有白蛋白的结果。看来这种类似白蛋白或类似酸肌球 的物质很可能是在核素转化中形成的中间产物,但最后形成胨那样经常成团的产物。我得到的是转化的哪一个阶段?对各别情况我不能确定,在显然相似的情况下,不同时间所得结果不一。显然,只有元素分析和对所形成的产物作精确的研究,才能对这两种反应是否可靠而作出肯定的结论。煮沸的冰醋酸既不溶解可溶性的核素,也不溶解不溶性核素。但看来也在慢慢产生种类相似的转化物。我没有用金属盐作任何反应,因为我知道只有核素碱溶液。另一方面,尽管材料很缺,只要许可,我就用它来确定元素成分的重要特性,我宁肯对少数极重要的成分作重复分析,而不是去进行一项单个的全元素分析,因为这一措施有助于更好地初步确定:所涉及的究竟是一个化学实体,还是一个混合物?一旦我发现有可能时,我将完成这些资料。此物质含有氮、硫,且特别富有磷。这样,蛋白质含磷的老习惯有一个真正的基础。
的物质很可能是在核素转化中形成的中间产物,但最后形成胨那样经常成团的产物。我得到的是转化的哪一个阶段?对各别情况我不能确定,在显然相似的情况下,不同时间所得结果不一。显然,只有元素分析和对所形成的产物作精确的研究,才能对这两种反应是否可靠而作出肯定的结论。煮沸的冰醋酸既不溶解可溶性的核素,也不溶解不溶性核素。但看来也在慢慢产生种类相似的转化物。我没有用金属盐作任何反应,因为我知道只有核素碱溶液。另一方面,尽管材料很缺,只要许可,我就用它来确定元素成分的重要特性,我宁肯对少数极重要的成分作重复分析,而不是去进行一项单个的全元素分析,因为这一措施有助于更好地初步确定:所涉及的究竟是一个化学实体,还是一个混合物?一旦我发现有可能时,我将完成这些资料。此物质含有氮、硫,且特别富有磷。这样,蛋白质含磷的老习惯有一个真正的基础。
(1)0.1915g可溶性核素产生1811Pt.=13.47氮。分离后用酒精没有提取出核。以后的测定是用热酒精提取的完整的核来进行的;
(2)0.2278g得0.2378Pt.=14.60氮少量氯化铂在蒸发时意外地被分解了;
(3)0.2545g得0.2518Pt.=13.99氮;
(4)0.1862g得0.1840Pt.=13.97%氮;
(5)0.3882g,用氢氧化钾和硝酸钾燃烧,得0.0494 BaSO4 =2.005%硫;
(6)0.4611g得0.0598 BaSO4 =1.78%硫;
(7)0.2453g得0.0318 BaSO4 =1.77%硫;
(8)0.3882g得0.0350 Mg2 P2 O7 =5.76%P2 O5 ;
(9)0.4611g得0.0430 Mg2 P2 O7 =5.96%P2 O5 。
上述的(5)和(8)以及(6)和(9)的分析是以同一样品完成的,但这两次取样来自不同制备物。氮测定用Will和Varrentrapp法,除(5)和(8)外,用苏打和硝酸钾进行燃烧。
我相信,尽管这一分析并不完全,但仍可通过它作出结论,我们打交道的不是一种意外碰到的混合物,而是一种化学实体,或是一种关系非常密切的物质的混合物,有少量杂质时除外。有利于这一结论的另一点是可溶性核素和完整核之间含氮量的密切一致,尽管它们在制备方面明显不同,而这必然会减少含氮量。在质量分析方面,随着晋升到卵黄磷蛋白或鱼卵磷蛋白,人们会想到它是一种含有蛋白质或蛋白质衍生物的卵磷脂化合物。在一种物质和相同物质中存在5.8%P2 O5 和14%氮而否定这假设。它更可能是一种特殊物质,这种物质不能同目前已知的任何一种物质(sui generis)相比较……
根据有待扼要报道的其他组织的试验,我看所有这些彼此略有差别的含磷物质将作为一组核素物质出现,这种物质或许值得同蛋白质作等同的考虑。
我只能设想,磷在生物体中起着最重要的生理作用。我特别记得人所共知的明显的事实,即在植物中,磷主要地或几乎独一无二地积聚在生长点;可以肯定核的出现局限在正在生长的部分,即局限在增殖过程中的细胞……
根据现有的材料,我谈得离题了。虽然除元素分析外,缺少一些更简单和更清楚的试验,从这些试验或可指望对核素物质同目前已知物质的关系有重要的发现。本人将尽早发表更多的消息。我想,所取得的结果尽管是零碎的,但吸收别人,特别是职业化学家们来从事这方面的研究,却是十分明显的。了解核物质、蛋白质及其新陈代谢中间产物的关系,将有助于逐渐打开至今还如此完全地把细胞生长的内部过程隐藏起来的屏幕。
对了解细胞及其生命现象的贡献 【3】
沃尔瑟·弗莱明(W. Flemming) 【4】
(1879年)
蝾螈幼虫尾鳍无色素区域是观察表皮活细胞分裂最好的物体。透明度极好的鳃丝没有显示出活的表皮细胞,而且尽管它们出现分裂核,但太暗淡,看不清楚。
尽管如此,还是用鳃丝作细胞分裂的固定和染色制品,因为它不要作切片或制备。我用过若干种试剂作固定剂,但总是归结到相同的3个结果最好的固定剂:苦味酸、铬酸和效果较差的氯化金。
苦味酸处理的主要优点是,当接下来再用苏木精或洋红(稍差些)染色时,会得到非常漂亮的核。染色前必须把酸充分洗净,苏木精溶液最好大大稀释。
蝾螈细胞分裂(活体后)比较其染色制备的说明
我的说明主要为鳍的表皮细胞和鳃丝。
人们很容易在充分喂饱的幼虫尾鳍的细胞表层和深层的静止核之间找到各分裂期。在活组织中能识别的最早各期如下:
第一期:细丝紧密缠绕成小篮子编织物状
与暗淡但可以明显区分的静止核不同,表皮细胞的中央有一个暗淡、且区分不明显的物体,它常常比静止核稍大些或大得多,这种核在活动状态下看起来像稠密、精细的颗粒,但这种颗粒只是外观上的:这一期的染色制片十分清楚地说明,它是一种由精细的螺旋细丝聚合在一起的稠密而有规则的结构,它在活动状态下太暗淡而不能全部看清,所以在光学透过和其螺旋体的斜切面,使人看起来像是颗粒。染色浅的切片中肯定已不再存在核仁。同时,仍然可以从染色物体上看得见的原生质,明显地区分出核的形状来,其外形细小,但明显。
同静止核的结构相比,活动核的盘旋结构密集得多,分布更有规律,更明显地成团;活动核的细丝完全或差不多一样细,静止核却不是那样。
碎片物质在其静止状态,对像网状似结构和核仁那样的可以染色,但染色程度稍差。另一方面,在特别清楚的卷绕时期(前期),染色的碎片物质不再出现。现在我们认为这种物质被吸收形成核部分,形成网状物,准备分裂(与此有关,核仁在此时丧失其形状并消失,大概在分裂)。这时染色物质尚未完全变成网状物。碎片物质染色暗淡,可能存在它的没有变化的残余物。恰好在静止核中,试剂使它们显现成颗粒,这两种残余物以后也消灭,并不再在核中出现任何能凝结成颗粒的物质;每一个染色的物质都被吸收到结构成分中。此后结构成分体积增大,且通过核同时分裂成几乎相同的螺旋系。所有这些首先发生在核的周围。
这种物质必须在核中发生实际的转化,这一点是很清楚的,必须记住在静止期网状物的碎片物能染色,而在分裂时失去染色能力。据此判断,发育核的盘绕网细丝,仅仅是包括核仁在内的静止网状物的一种形态上的重新调整,这一点是不可能的。实际情况是,盘绕细丝团显然比静止核的结构大,同时,如果可以这样说的话,可以估测篮子状结构所积聚的染色量,与包括碎片物在内的整个核在静止状态下所吸收的量相等……
第二期:母核的松散盘绕成篮子形状
倘若对中期形态核观察一段时间,可见的颗粒逐渐变粗和相互分开,不久便清楚地看出盘绕细丝联结在一起,但由于它们暗淡因而看不清楚。该物体的染色清楚地说明,确实存在着这种联结。核的形状是一个非常精细的由粗细均匀的盘绕细丝相互广泛联结在一起的篮子状结构,它能清楚地染色。同时碎片物质不再有一点点染色,并且核中不再出现任何精细的颗粒团(凝结)。核仁在此以前就消失掉。细丝越来越松散,它们的方向大部分同核的直轴成直角,或差不多成直角,这是一种在分裂后形成核时相当典型发生(甚至更明显地)的状态。
如果把这团篮子状结构同早先时期的染色物质团作一比较估测,这两者似乎是相等的。但如果比较细丝的粗度,第二期比第一期显著增粗,同时盘绕的密集程度减少了。关于这种细丝变粗可能由相邻的细丝合并在一起的想法已被排除。首先,根本没有找到这样一种情景:细丝中有一部分的粗度与第一期相同,另一部分的粗度与第一期比,则增加近一倍;其次,发现从细丝紧密地缠绕盘旋到粗丝松散地缠绕每一个变迁时期。据此出现下述关于发生转变方式最能被接受的意见:细丝密集篮子的丝变短了,同时粗度增加了,有点像肌肉纤维收缩成一根橡皮带,从其伸展的状态收短的方式,但要慢得多,以及同时慢慢地换成这样一种方式:它们总是同最大的可能范围保持一个等距离。
第三期:母核的星形状
分裂活动变迁到这一时期,网状丝似乎变得更加松散,而且从周围把环伸到明显的地方。中央仍然不清楚。变迁到这一状态的染色制片看到丝的层叠作用,其次序常常难以看清,但很快集聚一起,就看到细丝典型的层叠,尽管它并不是在所有情况下都是同样清楚的。这就是说,细丝中央和周围发生弯曲,我把它们简称为环。
到目前为止,我已看到过另一个非常突出的现象:细丝本身纵裂为二。这个过程在盘绕期终了时就已经发生,或发生在现在谈到的第三期的过程中。因此,在所有这些情况下,单丝或双丝这两者都会碰到。在转化到星体的时期,这些丝可能还是单根。但无疑在蝾螈方面有大量图片证明丝的纵裂是一典型过程。
表皮细胞核和红细胞细胞核,其丝的各半几乎完全是平行的,在内皮细胞则有点分开,以及在结缔组织细胞常常在同一方向上折回一个短的距离。
以后这些丝沿着它们的整个长度彼此分开,在这种情况下出现一很好的丝星(threaded star),它的射线同过去的相比,数目上多一倍,粗度上少一半。
下一期(赤道板)的丝粗度也经常是单丝星(single-threaded star)粗度的一半,这一简单的事实说明,这种丝的纵裂(至少在蝾螈中是这样)是一个重要而稳定的时期。
目前还没有核分裂的研究人员作过这种丝分裂的报道,因而我马上向自己提出问题,这是否是试剂的作用?由于用苦味酸和铬酸制片都经常出现相同的现象,却又未必是试剂的反应。鉴于我幸运地在几种情况下都看到活动状态下的双丝,我就能排除上述想法。
第四期:赤道板
这一期发生得快,通过得也快,因此用试剂不太容易使之固定;但是,在研究每一个活表皮细胞分裂时,总看到重现这种典型的时期,同时由于它特有的形状,值得把它作为一个单独的时期。
上期特点是平展的星体以两个圆锥体的形式朝两极伸展,这一期则不同,丝的集聚方式是所有的丝最初有点盘旋,以后展开成越来越同分裂轴平行,占据了一个厚板的位置,约为整个细胞长度的1/5~1/4,有时为1/3。这个板经常在赤道位置,并坐落在同分裂轴呈直角之处。
人们可以就这一时期提出以下两点可能性,而无须对其中的每一点提出证据。
(1)两个未来的核物质早已准备就绪,并在星体时期聚集在一起。在这种情况下,赤道板时期只是把这些成分重新整理而已。
(2)两个核各半的分离可以发生得比赤道板时期早,这也是可能的。在这种情况下,一个特别值得注意的现象是丝的纵裂。它总的来说预示着什么?当发现这一现象时我立刻想到,尽管方式很不相同,或许它代表着分裂为二的同源性,核板成分将按照Strasburger、Bütschli和O. Hertwig等人的发现进行分裂:每根丝纵裂一半的丝移入核的一半,另一半丝移入核的另一半,换句话说,各自进入未来的子核中去。
如果我们假定一个双股丝的纵裂一半预定给一个新核,而另一个纵裂一半给另一个新核,和所有的双股丝一样都是这样分配,我们还可以进一步假设(确实不能证明),在进入第四期前,所有中央的环分开,这样,每一个双股丝分为4个1/4丝,而不必要再分裂,但只要把4个1/4丝的每两个分别进入核的一半中去,这可以发生在赤道板时期。从核图像的极一边伸到赤道板水平的每一根丝代表一个1/4股丝。
可以认为,在这期看到两边的丝在赤道板上的联结,必须是无特别重要的丝端的第二次接触或暂时的融合。
我之所以在这里提出此假设,是因为据我看来对丝的纵裂还不值得去作一些解释,那就太奇怪了。同时,我提出这种假设纯粹是一种可能性,决非坚持己见。
第五期:核图像的分开
这期的表现只说明核两半的分开移动,因为核两半的实际分开早就开始了。
每一半核图像有点像一只阔的盛鱼的篮子,但带有向外稍凸起的绳。在极的视阈看这时期,它的样子像一颗星。但在表皮细胞观察不是很清楚,上面已经提到,因为细胞早已在水平面上分裂。
第六期:子核呈星状
对目前末端自由的两个各半核的丝各朝着相反的另一边移动,而越来越离开,这样,有些位于周边的丝常取向于细胞的极。在这种情况下,图像为一个伸长的星体样子,有时非常有规则,有时不太有规则。
此时细胞体的一边常常出现缢痕沟。
第七期:子核呈环圈和盘旋状
活细胞中的每一个子核有点像早些时候第二期的母核。其特点是不断地深入到极一边,因此这两个形状为外凸内凹板的篮子,彼此把它们外凸的一面对着另一个篮子。在这期的稍后过程中,它们盘绕在一起如此密集,以致活动的幼核给人的印象是一丛粗糙的内部均匀一致的物质,染色非常清楚,但这是一种错误的印象,而一个完全均匀的时期根本不会在这里产生。只要在这个明显均匀一致的类型中加上醋酸,就立刻看到不规则的棒结构的清晰图片。
细胞在这期分裂。早在以前的时期就出现分裂的第一个标记。缢痕沟也逐渐影响到另一边,赤道渐渐变窄,细胞体收缩成两个;在表皮细胞中,这一过程在无干扰、无停歇的情况下相当缓慢地发生(我没有在其他细胞直接观察到这种过程)。细胞内部的赤道板无明显区别。
第八期(如有人希望这样鉴别):网状子核,回到静止状态
从第七期变迁到静止状态全部时期到处大量发现成对的幼核;因而这种变迁继续相当长时间。很清楚,细胞分裂后首先是丝盘旋,然后大多数丝排列成横向伸展到核的纵轴。由于这样的结果,这些成对的横断幼核初看起来,除它们小些外,像静止时期。细丝结构从这种状态进入到均匀一致的网状结构的状态;但丝不再盘旋。网状结构逐渐变得更浓密,却始终呈暗淡色,而核慢慢增大。与此同时,核在细胞体内轮廓勾画清楚,而丝之间的间隙物质现在能染上色。但是在核那里还看不到确切的有形的膜。在下一个呈黄色、更暗淡的时期,出现的轮廓更加清楚,于是它的形状恢复到静止时期的核。这些网状幼核,尽管呈色暗淡,但比活动状态和保存状态下的老核更清楚,更有规则。我的印象是核膜并不长成一连续层,它变硬,或模糊不清,而实际上它是网状物的边缘部分在细胞质的周围联结成一个薄层。
我没有直接观察到核仁的样子。在幼核和在暗淡、较大的核中,还是看不到核仁。因此,这一非常重要的问题仍有待于确定。
所有这一切都明显地看出,子核的形状最初是一个伸长的星体。它转变成一个星体或带有盘旋细丝的环,它们又变成两个环,一个在外和一个在中央,从这些环长出一束旋卷丝。从这一束旋卷丝形成带有间隙物质的网状物。同样明显的是,除双星外,这整个过程是母核所经历过的一系列恢复变化。
卵成熟和受精的研究 【5】
埃德瓦·贝内登(E. V. Beneden) 【6】
(1883年)
如果我可以这样表达的话,我希望提请注意,卵在其成熟史中,各连续时刻的染色质成分的数字组成问题。
在全部发育时期,我发现卵核染色质团分成两个部分,每个部分由4个成分组成。这4个成分常常成对聚集在一起。
萌发粒中有两个核板,各由4个染色质体组成。
在第一个极体中,我们发现两个染色质体,各由2个或4个或多或少明显分开的部分组成。
在未成熟卵核(次生卵母细胞)中,我们发现两种染色质残留物,它们分开成两组成分。
第二个极体有两个染色质体,各由两部分组成,每一个部分又由两个黏着的成分组成。
雌性原核有两个染色质聚合物。每个染色质聚合物由两个小型染色团组成,每个染色团有两个染色深的棒……
至于雄原核的成熟现象,它们同我关于雌原核的描述相同:当它们到达其完全发育时,同样构成两个成分……然后两个原核朝着卵的中央相互靠拢。通常在卵的上部附近长出雌原核,它为了配合比雄原核移动的距离要大得多;后者移动的距离不多。
卵通过间接方法(有丝分裂)发生分裂,两个没有融合的原核,都参与形成一单个的具双着丝的图像。尽管这两个原核仍然完全不同,但它们同时进行,在有丝分裂过程中,正常细胞的核发生相同变化。每个原核给赤道面的染色质星体提供两个环;星体由两个雄环和两个雌环组成。每个原核纵裂为二,各一半。一份进入到其中一个子核中去,而另一份进入到第二个子核中去。开始时,雄的和雌的染色质成分之间不发生任何融合。如果确实发生这种融合,它只能在头两个胚细胞核中发生。赤道板的染色质星体一部分由雌原核,一部分由雄原核提供,这一点同分裂球的分裂形式是相同的。两边的星体由4个染色质环构成。如果在头两个分裂球分裂时,看到的染色质星体代表分裂过程中的核,那么由原核发育的第一个星体就等于一个单细胞核。
但是,既然这种表达方式也十分适用于较早时期,这就使我们考虑到,尽管这两个原核彼此相距一定距离并完全分开,却共同代表一个细胞的单核:每一个原核等于核的一半。但这一结论不能使我们忘记,不论在有丝分裂之前或有丝分裂期间都没有发生任何融合,至少就原核的染色质成分来说是这样。
两个原核的非染色质物质都不会掺合成一团;简而言之,尽管两个原核表现为好像它们一起构成一个单核,但在分裂的头几个时期,它们是可以区别的。没有发生两个原核结合成一个形态上单独的胚核。在叙述卵分裂成头两个分裂球之前,我们必须向自己提出问题,在哪一个时刻应当认为卵是受精了以及受精主要包括些什么?
从我刚才小结的事实明显地产生一种想法,由两个原核所提供的卵,其行为像一个单细胞,而卵含有的两个核成分的总和等于一个正常的核。在两个原核形成的那一时刻,就完全构成了胚的第一个细胞;因而受精是与这些原核的发生相吻合的。
上面我已经提出,只要第二个极体没有消失,那些预定产生雄原核的精子成分不会发生任何变化。但是,从卵本身摆脱掉它的第二极体和它的第二卵周层的那一时刻起,精子产生原核,与此同时,形成了雌原核。因此,看来当卵清除掉本身的产物后,卵只对精子有影响,只决定精子在形成半个细胞核(雄原核)时所起的作用。只有在此之后,卵本身才显示出性的特性;初级卵细胞只留下一个带有半个细胞核的退化的卵黄。
雄原核继续完成这个我称之为雌性原细胞的退化细胞,并使之成为一个新的完全的细胞;受精显然主要包括第一个胚细胞的重新构成,这个重新构成恢复并提供胚细胞本身转化所需要的能量,以通过一系列越来越复杂的阶段成为一个同亲本相似的个体。看起来细胞起源于两个不同成分的奇妙的现象,是同卵细胞在一个方面和精母细胞在另一方面排出某些物质紧密地联系;卵排出精体和卵周层;精母细胞排出被寄生细胞部分……在这一点上,卵细胞和精母细胞同所有其他的细胞是有区别的。当卵为其雌原核补充时,它就不再是我称之为雌性原细胞的一部分了,精子同样是一个退化的细胞,是一个没有寄生细胞部分的精母细胞,我称之为雄性原细胞。据我所见,不把这些退化或消失的现象同再建造或受精的现象联系在一起,不把后一种现象看做是一种更换或替代是困难的……
两个原核的非染色质外形变得越来越不清楚;原核体相互压挤在一起形成内有4个染色质环的明显的一团东西;这些染色质环都取向同一个平面,即位于分裂过程中的细胞的赤道面上。环的数目是稳定的:人们总是有规律地发现它们是4个;长度几乎相同。从最近研究发育早期的证据看出,4个环中的两个来自雄原核,另外两个来自雌原核。
现在我必须提到很多卵在进入被研究的那个阶段所碰到的另一个现象,这种现象有时在前一个阶段就已经出现,我所指的现象是染色质环的加倍。染色质丝纵裂这一事实是由弗莱明(Flemming)在蝾螈组织的分裂细胞中发现而后为雷济厄斯(Retzius)和普费兹纳(Pfitzner)所肯定。按我的意见,这是核分裂最重要的事实之一。如果记得构成赤道面的4个环,两个由雄原核供给,另两个来自雌原核,我们就得出结论:每一个子核从精子接受一半染色物质,从卵接受另一半染色物质。如果原核有性的特性,如果一个是雄性,另一个是雌性,那么显然卵裂球的头几个核是两性的。
对于在核分裂时染色质丝加倍的原因,弗莱明表示过怀疑,他怀疑每一个初级环是否没有给每一个子核提供一次级环。可能他是以这个假设来理解为什么有这种加倍,他不能用任何实际观察支持这种假设;蝾螈核中环的数目很多,以致不能追踪每一个环以观察其变化情况。马蛔虫的卵或分裂球的典型图像则没有碰到这种困难。正是这种相对的单纯性,使我得以明确地解决对说明细胞间接分裂如此重要的这一问题,并确定细胞核的染色质物质,是从雄性和雌性性原细胞,即一方为精子,另一方为成熟卵那里遗传来的。
群体遗传与纯系 【7】
为解决选择中的突出问题而作
威廉·约翰森(W. Johannsen) 【8】
(1903年)
摘要和结论
本文即将讨论的全部问题将同时对著名的伽尔顿(Galton)回归定律进行一次全面的肯定和总的阐述,它论证了亲本与后代之间的关系。本文不涉及其他的回归关系。
根据我的研究材料看来,非常符合Galton定律。这一定律说明了不同于群体平均性状的各个体所产生的后代,与亲本的平均数差别一般较小,但方向相同。群体内部的选择使平均性状的选择方向发生或多或少的变迁,有关个体就是在这种平均性状的周围上下起伏。
而结果是尽管我用的材料能够分解成纯系,但我还是未能继续把群体看成是完全一致的。现已表明,在所有的情况下,纯系内部的上述退化已经实现了纯系内部的选择没有发生新的基因型变迁。
因此,群体内的选择经常会发生的平均性状变迁是一种标志,它说明总的群体由基因型能多少加以区分的不同的系所组成,至少我所用的材料是如此。在通常的选择过程中,一个群体有可能变得不纯,这是这些系没有完全分离出来的结果,其基因型就使群体平均性状在方向上发生偏离。
典型的、众所周知的选择结果,是每个世代都朝着选择方向有阶段性的进展,因而阶段性的进展又取决于被选择的各系每个世代的阶段性进展级数。现在很容易理解,选择的作用不能超越已知的限度。或具体说,当纯化时偏离最强烈的纯系已经被分离出来时,选择的作用必定终止。在这方面,必须指出仅仅根据一个样本所表示的变异表或曲线同二项方程式数字比例之间的一致性,人们决不能确切地肯定在这个样本中只存在一个单独的基因型。在正常的含义上,代表一个种性纯的群体的个体变异曲线,常常或的确在多数情况下能证明它是代表群体里面不同纯系的许多基因型的结果。这样,平均值并不总是能代表一个真正的基因型。在这一点上,一种纯统计手段就有很大的缺陷。
为此,我试图在本文的自始至终把平均数的概念(平均性状、平均值等等)同基因型的概念予以严格区别。这两种截然不同概念之混淆,只能如此频繁地造成误解和错误的推理,或者不仅在遗传学领域中如此。必须承认,不作详尽分析而想区别这两种概念,常常十分困难。在纯系中这两种概念常常是同一回事。一个基因型的数位表示经常是一个平均值,但决非总是如此。
就形态学性状来说(至少形态学性状的整个体系,在分类学研究中的价值已获得一致公认),不同基因型之间的区别,是对单独的个体通常能识别出来,尽管有变异,但这种变异属于这一种或另一种最狭窄的分类范畴(例如Jordan的亚种)。
通常能把这些形态类型编成一套精确的变异组,但鉴于以下原因却有很大的困难:因为一个不同基因型个体的混合群体,可以同一批属于同一个独特基因型的个体结合在一起。德弗里斯(Hugo de Vries)的一个混合体如月见草类型或拉恩基尔(Raunkiaer)的蒲公英“Geschlechter”,其主要形态特性同单独一个类型的纯种栽培的情况是不同的。
关于各种性状中一种生理性更明显的性状,即Hj. Nilsson的非植物学性状,例如最高度和其他比例、生化性质、可靠的数学关系等,我们碰到的是另一种情况。通过隔离栽培很容易证实确实存在这种明显的基因型,它们只有数量差异,所以不同基因型的变异曲线会重叠,而有人则得到德弗里斯的越亲曲线。如果一个混合群体的个体属于在这类特性中有一个特性明显不同的基因型(比较蚕豆的大小和宽窄),这个混合群体很容易产生一系列连续变异,却不可能在基因型之间直接识别这种特性的差别,而平均值将被错误地当做是一个单一种基因型的平均数。在这种情况下,不可能辨认出一个单独个体所属的那种基因型。表1对这一点作了很好的说明。
表1①
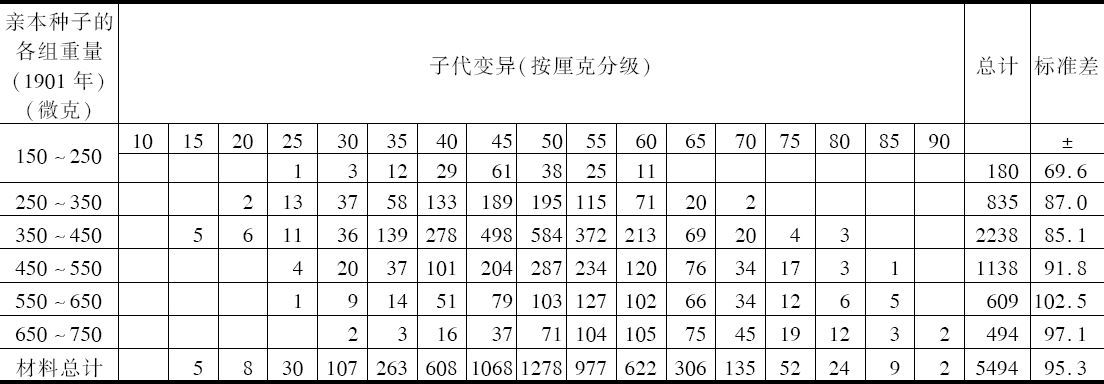
①子代种子重同亲本种子重的关系。1901年得到的种子按重量分级,然后种植。
每一重量级成员所产生的种子进行称重并分级,其结果见“子代变异”下各栏所示。
正因为这些原因,它或多或少使人们已清楚地认识到或刚刚感觉到,以上我称之为真正的形态学性状的研究为分类学提出了一个中心问题。只是近年来在更低等的类型方面,生理性更强的性状才使分类学家感兴趣,这些理由也说明为什么突变研究者在真正的形态性状中找到了他们研究的主要依据;另一方面,这些用来主要确定植物全部习性的特性或者不能够、或者只能部分地用数字测量表示,而这种数字测量几乎总是把它们的值转换到精确的测量和计算方法的范围之内。
这样,探究变异和遗传法则的生物统计研究,首先包括生理性更强的性状,或一般来说即Bateson所谓的比例性状,这些性状能用数字清楚地表示,如体积大小或重量。个体之间的比较使人们无法根据彷徨变异性的表现来区分基因型的差异,这就是以下Galton-Pearson概念的主要依据。当人们不考虑纯系时,必定得到结论,选择强烈变异的个体(或正的变量或负的变量)能在被研究的基因型中引起真正的变异。虽然,这种截至目前已一致公认并得到生物统计学家韦尔顿(Weldon)和皮尔逊(Pearson)支持的概念,必定反对把突变看做是另一种同彷徨变异同样重要的变异。根据所积累的群体遗传的全部统计学知识,或者感到生物学并不需要承认突变理论,我之所以在这里说“或者”,是在某种程度上碰到了生物统计学家的反对意见,就我自己来说,德弗里斯规模宏大的实验,业已证明突变的存在是毫无疑问的。
从研究结果清楚地看到,我在这里提出的有关亲本和子代间关系的Galton-Pearson法则,其根据同至今已予承认的情况有所不同。就我们的研究而言,父母亲、祖父母、或任一个其他祖先的各别特性对子代的平均特性不产生影响。一个系的基因型是在同特定时间、特定场所的外界环境发生紧密相关的情况下工作的,而这种外界环境又决定着一个个体的平均特性。因此,正如德弗里斯根据相似的情景已如此清楚地指出,这个系是完全稳定和高度变化的,尽管这显然是一种相互矛盾的方式 【9】 。
与此同时,这决不意味着纯系是绝对稳定的。
首先,经过无数世代对彷徨变异体的选择,有可能最终使一个系的基因型发生变迁。这就是经常指出的把生物统计学家的见解,应用于那些现在不是纯系,但可以分出纯系的群体中,但这一点尚未得到肯定的论证。证明这种可能性的着重点是人们希望确证这种选择的有效性。
其次,我们必须考虑杂交育种,在发生杂交育种的情况下,纯系必定丧失掉它们纯的状态!然而,整个杂交种问题不是我们所要讨论的部分。
第三点是突变,即基因型不规律变异的可能性。要在最大程度上规定突变还不成熟;首先必须证实在更多的有机体中存在突变。据我所见,突变的确存在,这一点毋庸置疑;我希望在以后发表的刊物中提出特定的明确的证据。本文除涉及根据子代各个体方向不规律的偏离,而不能特别地严格地鉴定它们为定向突变这一问题外,其余的就不准备多谈了。
我必须提到怎样才能解释德弗里斯的意见,这一难以处理的问题,这就是人们经常观察到负的变异,这在新发现的基因型中尤其突出,这一事件有理由引起生物统计学家对此提出疑问。希望本文提出的研究将对这一问题有所启示,它或者至少看来是把彷徨变异和突变之间的界限拆掉了。
德弗里斯在他的《突变理论》 【10】 一书中有一章关于“营养和自然选择”,其中讨论了母本植株营养丰富或缺少的结果。我毫不怀疑营养方面实际的或想象的差异,是与出现选择或不出现选择同时发生,这一点将说明德弗里斯的例子。还有,个体发育期,即德弗里斯所命名的敏感时期特别有意思。我能在我的研究材料中引证相似的现象,但存在着极大的困难。必须知道,我的意图不是用纯系原理来解释这一问题,因而除了所选择的性状方面的差异同截然不同的或实验设计的产地有联系外,不可能得到别的什么东西。在这一点上,新拉马克主义者特别感兴趣,但还要作大量的研究工作,并肯定要使用真正的纯系作为研究材料。
我的主要目的是对祖先和后代之间的伽尔顿回归有所阐明,而我相信我的材料同伽尔顿的材料一样明显地有其自然特性,这种材料作为把伽尔顿定律应用在群体方面进行分析的基础是有价值的。我的见解同伽尔顿或德弗里斯的见解皆不矛盾。
如果我的研究得到广泛传播,并抓住了它们的意义远远超过本文讨论的特殊例子,这项工作总的结果将会对贝特森(Bateson)和德弗里斯关于非连续性变异或突变在进化理论方面的巨大意义赋予重要的支持。就本人所进行的选择而言,仅在选出一个早就存在的基因型的代表者方面是有效的。通过保留那些同所需要的方向不一致的个体,就不能连续地得到这些基因型;它们将很少被发现和分离出来。
从纯系研究得到的知识,与杂交方面的知识相结合,可作为群体遗传研究的一个起点,此群体由于需要不断的异花受精或杂交,致使纯系不能得到完全的分离。早些时候曾经指出,这种见解同德弗里斯伟大著作中的基本意见完全相符,并已看出我是通过同德弗里斯略为不同的途径得到这一概念,注意到这一概念是根据不同资料得出的,这一点也很重要。
此外,相关变异的重要问题是一个轻微变化的性状取决于究竟所研究的是一个纯系还是一个群体。在后一种情况下,我早些时候就打算证明,某一个相关比例(Pearson的术语)不一定能强有力地代表一种合理的关系,在一个纯系内所表示的一种相关的关系,其意义要重大得多。我的综合表很好地说明了这个概念,即纯系内的选择不可能使蚕豆的长度和宽度间的相关性发生变异,而要从原始群体中分离出真正不同的基因型,例如看来是完全同质的、窄的和宽的蚕豆则不难。
还有,我们必须估计到突变的可能性,这样即使最强的相关关系也会被破坏。目前我不想涉及这一问题,在以后发表的一篇文章中,我希望用作为研究基础的纯系原理对此作更多的阐明。
如果读者经了解本项工作而对伽尔顿、皮尔逊和其他生物统计研究工作者的重要工作价值置于怀疑的地位,我必将真正地感到遗憾。我不能冒昧地批评皮尔逊的论述,特别是他对一个特殊群体内有关祖先影响问题所作的论述。但我的确这样想,如果纯系原理为皮尔逊这样一种人所掌握,它就能把生物统计的研究较之其群体研究大大地推向前进。很明显,皮尔逊所研究的这种关系具有伟大的科学意义和重大的实践价值,但它们不适于完全地说明遗传学的基本定律。
而什么东西特别地影响伽尔顿的研究?据我估计,是本文提出的结果以一种漂亮的方式支持伽尔顿早在1876年已经提出的“血统论”的基本意见。这个定律包括最近关于“种质连续性”的魏斯曼理论中几乎全部有实际价值的内容。魏斯曼的这个推测使提得更简单的但并非没有独创精神的且相当原始的伽尔顿意见相形见绌,这或许在某种程度上应归结于伽尔顿本身,因为在他的最近著作中,他没有看出借助于研究进展坚持他的血统论是合适的。血统论同伽尔顿的回归定律并不完全相符,这一点是对的,但再没有比我在下面读到的结果能更好地支持和说明这种论点了:据我看来,对一个纯系基因型常见的完全回归,是对血统概念稍作修改的最漂亮的证据。伽尔顿的血统概念不能不加改变地维持下去,这一点是正确的。尽管魏斯曼最近把伽尔顿当做通过决定了实现细胞限制的“声音”,但人们可以命名这些理论上的遗传颗粒,而德弗里斯在识别出遗传颗粒的单位性质方面是值得倍加赞扬的,他把这种遗传颗粒称之为泛子,于1889年首次公布了这个概念,并在《突变理论》一书中得到了进一步的发展。我认为伽尔顿-德弗里斯理论是唯一正确有用的遗传理论。
本文把纯系原理作为一个绝对需要的原理,而带入遗传学研究的真正深入的探索中必将取得成功。然后将达到它的最高目标。以后发表的文章将用以说明各纯系方式多种多样的作用。本文中我只研究了一种方式的变异,以便以其最简单的例子提出我的概念。
可用下面伽尔顿常引用的一句话简练而最清楚地表示从事这项研究的思路。
“在你能掌握它以前,你首先必须把它拆开,然而再把它组合起来。”
Vilmorin强调各部分的区分,伽尔顿证明重组的合理基础;我在此则尝试着把这两位在他们各自的观点方向都值得尊重的、具有独创精神的研究者的观点结合起来了。
染色体遗传 【11】
沃尔特·瑟顿(W. S. Sutton) 【12】
(1903年)
作者在最近发表的Brachystola 【13】 不同细胞世代染色体精确研究的一些结果中,扼要地指出,染色体研究中所叙述的一些现象,同孟德尔在1865年根据植物杂交种方面的观察首先作出的,最近又被一些有才能的研究工作者予以肯定的某些结论之间,存在着肯定的关系。威尔逊(E. B.Wilson)教授在其简讯中又进一步提到其理论问题。本文针对这些问题作更详尽的讨论,其推理性可通过指明工作的某些线索予以证实,以便测定所下结论的可靠性。此处提出的一般概念是作者在知道孟德尔定律之前纯粹从细胞学资料推导出来的,而现在却是细胞学家不容推辞地对完全熟悉遗传学试验研究结果的贡献。正如以后会看到的,这些一般性的概念完全满足典型的孟德尔情况的条件,而且看来许多同孟德尔类型已知的偏离可以用正常染色体过程中容易想象到的变异来解释。
我们早就认为,必须注意生殖细胞机体,以便最终确定遗传现象。孟德尔充分重视这一事实,并甚至安排专门的实验以确定生殖细胞机体的本性。他根据这些实验作出了伟大的结论:在有机体中,当母本和父本的潜力表现在每个性状的范围时,与每个性状有关的生殖细胞是纯的。然而,由于对细胞分裂的性质一无所知,孟德尔没有在这方面进行过比较,但是,对于近年来重新恢复并扩大孟德尔实验结果的人们来说,反复地看到在细胞机体和细胞分裂之间可能有关系。对此,Bateson用下面一段话清楚地谈到他的印象:“不可能有这样的事实:在孟德尔情况下,杂交种会产生平均来说每种配子的数目相等,这就是说,一种对称的结果,无疑是这个事实必须同细胞分裂所产生的配子的一些对称的分配情景相一致”。
作者在将近一年之前根据在Brachystola中看到的情况,弄清楚生殖细胞染色体组的高度组织性在遗传方面肯定有意义。在文章中早已指出以下几点:
(1)前联会生殖细胞的染色体组是由两个相等的染色体系统组成,有充分的根据作出结论,其中之一是父本染色体系统,而另一个为母本染色体系统。
(2)联会的过程(拟减数)包括两个染色体系统的同源成员(即大小相同的成员)成对地联合。
(3)第一次后联会或成熟有丝分裂是相等的,其结果无染色体区别 【14】 。
(4)第二次后联会分裂是减数分裂,它使得在联会时成对结合的染色体分开,并把它们送往不同的生殖细胞中去。
(5)贯穿各细胞分裂的全过程,染色体都保持其形态特性。
众所周知,许多种卵,其母本和父本染色体组对大量有丝分裂来说,彼此间明显地保持其独立性,由于牢记这一事实,作者首先倾向于认为,在减数分裂中,所有母本染色体必须移向一个极,而所有父本染色体则移向另一极,生殖细胞就这样分裂成两种,把它们分别称之为母本和父本。这同最近Cannon进一步提出的概念是一致的,但不久就发现这个概念同许多已知的育种实践有矛盾,因此:
(1)如果杂种的生殖细胞血统是纯的,则杂交育种的效果不会超过第一次杂交的情况。
(2)如果任何一个动物或植物只有两种生殖细胞,那么一个单一配对的后代,只能有4种不同的组合。
(3)如果从每一个成熟的生殖细胞整个地排除掉母本或父本染色体,一个个体就只能从每一个后代亲本系的每一代接受一个祖先的染色体(特性),也即不能从祖父和外祖父,或祖母和外祖母都把染色体(特性)遗传过来。
在这些考虑的推动下,更仔细地研究了整个分裂过程,它包括分裂前染色体在核中的位置,纺锤体的起源和形成,染色体和发散中心体的相对位置以及纺锤丝同染色体的接触点。研究结果没有证明整个配子染色质的亲本纯洁性。相反,发现有许多地方有力地表明 【15】 ,减数分裂赤道板上的二价染色体的位置纯粹是一种机会而已,这就是任何一对染色体可以同走向哪一个极都无所谓的母本或父本的染色单体靠在一起,而不管其他对的位置如何。因此,一个个体的成熟生殖产物可能有大量的母本和父本染色体的不同组合。为说明这一点,我们可以考虑一种形式,在体细胞和前联会生殖细胞有8个染色体,最后在成熟的生殖产物为4个染色体。该物种生殖系统,总的说来,可用字母A、B、C、D来表示。而任何一个分裂核可以认为它含有A、B、C、D的染色体来自父亲,a、b、c、d则来自母亲。作为联合同源染色体的联会,其结果形成了二价染色体Aa、Bb、Cc、Dd,它通过减数分裂再分解其成分。由减数分裂得到的每一个成熟生殖细胞必定接受每一个联会对中的一个染色体成员,但母本和父本染色体有16种可能的组合,形成一完全的系统,即a、B、C、D;A、b、C、D;A、B、c、D;A、B、C、d;a、b、C、D;a、B、c、D;a、B、C、d;a、b、c、d;和它们的配对:A、b、c、d;a、B、c、d;a、b、C、d;a、b、c、D;A、B、c、d;A、b、C、d;A、b、c、D;A、B、C、D。这里,一个在其减数系统中具有4个染色体的有机体,不是两种配子,而是可以产生16种不同配子;以及两个无亲缘关系的后代可以出现16×16或256种组合,而不是亲本配子纯合性假设所限定的4种。此外,只有少数有机体,其染色体少到8个,还有,由于每增加一对染色体,使生殖产物 【16】 可能的组合数增加一倍,而接合子的可能组合数则增加4倍,显然,在具有24~36个染色体的正常形式中,其可能组合的概率就十分大了。下表表示在前联会细胞中具有从2~36个染色体的可能组合的形式。
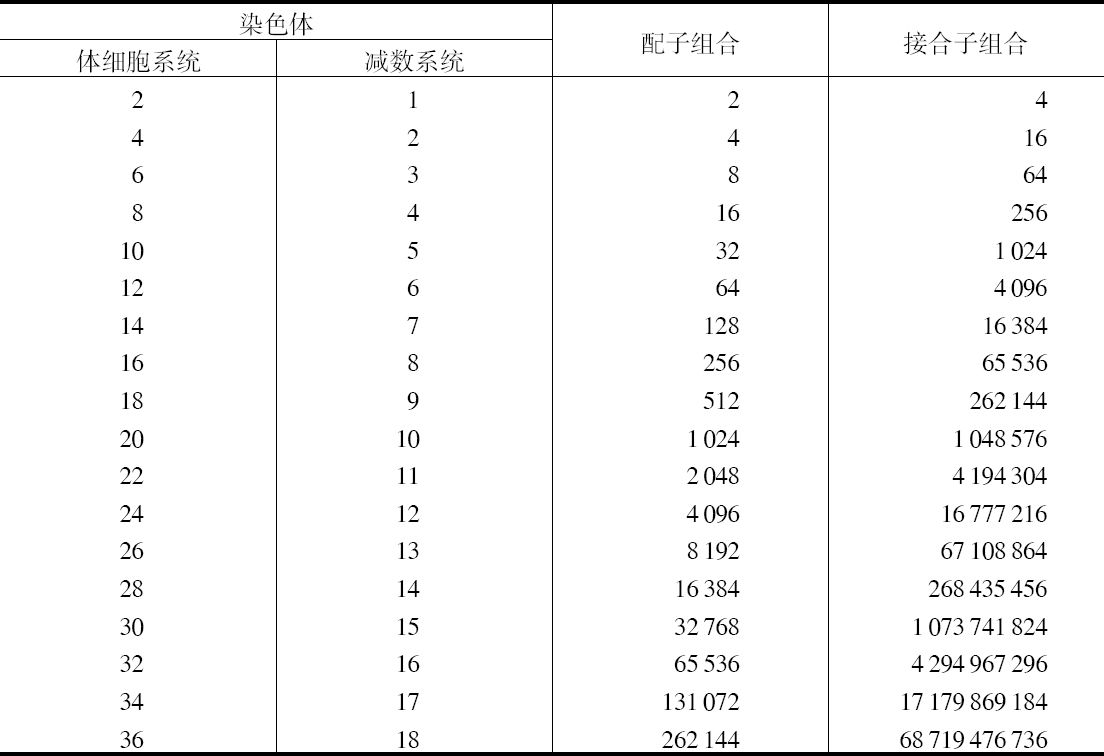
如果Bardeleben估测人有16个染色体(所作的最低的估测)是正确的话,每个个体参照某染色体组合,能够产生256种不同的生殖产物,以及一简单配对后代的可能组合为256×256,或65536;而且有36个染色体的Toxopneustes其单个个体配子和配对合子的不同组合的可能性分别为262144和68719476736。就因为配子中母本和父本染色体有数目如此之大的组合的可能性,它把染色体理论同遗传学方面已知的事实带来了最终的联系;孟德尔本人找到两个或3个不同性状确切的组合,和发现它们彼此间独立地遗传,并在第二代中出现大量各种各样的组合。
在Brachystola染色体中观察到其恒定体积上的差别早使我产生怀疑,单研究精子发生不能证实减数系统的各别染色体在发育中所起的不同作用。看来以后Boveri在一项有关正常系统中确实缺乏某些染色体的幼虫研究,所得结果证实了这一疑虑,其唯一的结论是染色体在质量上是不同的,并各自代表不同的潜力。采纳这一结论,我们必须能在任何一个染色体的遗传行为,同有机体中与该染色体有联系的性状的遗传行为之间找到确实的关系。
就性状而言,孟德尔发现,由两个在某一特定性状上不同的个体进行杂交,产生的杂交种作自花受精,在大多数情况下,其后代在不同性状方面是顺应一完全明确的规律,用字母A表示原始亲本之一所看见的性状,字母a为另一个原始亲本的性状,那么该杂交种经自花受精所产生的全部后代,以该性状来说,可用公式AA∶2Aa∶aa表示。这就是说,接受原始的纯种亲本之一的性状只有四分之一,接受另一纯种亲本的性状也只有四分之一,而后代的一半接受两个原始亲本的性状,由此提出了杂种产生这些后代的条件。
至今我们还没有用图解公式来表示在相似育种实践中的染色体组合,但显然,根据早已得到的资料,现在可以作出这类公式。Bracbystola减数染色体系统由11个成员组成。它们中间没有两个染色体的大小是完全相同的。在我早些时候的文章中,用字母A、B、C……K来区别这些染色体。在非减数系统中,有22个成员 【17】 ,能看出它们构成像成熟生殖细胞那样的两个系统,而可以用A、B、C……K+A、B、C……K表示。联会使同源染色体联合和产生一加倍成员的单独系统,即:AA、BB、CC……KK,而减数分裂使这些成对的染色体分开,因而每一对的一个成员进入到每一个所产生的生殖产物中去。
有理由相信,Brachystola某一染色体的分裂产物在其各自的系统中,像它们亲本成员一样,保持着同样的体积关系;这一点加上不同染色体代表不同潜力的证据,使这种体积关系是某一组性状的生理基础的特点成为可能。但是物种中任何一个减数系统的每一个染色体,在其他任何系统中都有一个同源染色体,根据上述考虑,其结果是这些同源染色体在发育中占有同一地位。如果是这种情况,那么,在后代的前联会细胞中,来自父方的染色体A及其同源染色体,来自母方的染色体a,可以看做是父方A和母方a相对立的单位性状的生理基础。在联会中,同源染色体的联合产生减数分裂的二价染色体Aa,上面提到在减数分裂中,此二价染色体Aa分离成为A和a成员。在所有情况下,它们都是这样分到不同的生殖产物,因此在一个雌雄同体的有机体,我们就有以下4种配子:

它们将产生4种组合:
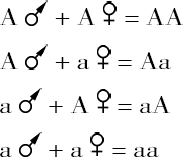
由于第二种第三种组合是相似的,其结果可用公式AA∶2Aa∶aa来表示,它同孟德尔情况中任何一个性状所得的结果是相同的。因此生殖细胞分裂的现象和遗传现象似乎有相同的本质上的特点,即单位(染色体、性状)的纯洁性和相同单位的独立传递;而结果是在每一种情况下,所产生的一半配子确实含有两个对立单位(染色体、性状)中的一个单位。
在以往的考究中我们有理由相信,染色体和等位基因 【18】 或单位性状之间存在着肯定的关系,但过去我们没有提问,究竟是把整个染色体还是把染色体的一部分看做单个等位基因的基础?毫无疑问,答案必然赞同后一种可能性,不然的话,一个个体所持有的各别性状的数字,就不能超过生殖产物的染色体数;这无疑同实际情况相违反。因此,我们必须想到,至少有些染色体同若干个不同等位基因有关系。然后,如果染色体永恒地保持它们的个性,那么无论哪一个染色体所代表的全部等位基因必定在一起遗传。另一方面,不要设想,所有的等位基因都必定在有机体中显示出来,因为这里有一个显性问题,且不知道显性是否是一整个染色体的功能。可以想象染色体能分成更小的单位(有点像Weismann的意见)它们代表等位基因,它既可以是显性,也可以是隐性。在这种情况下,相同的染色体可以同时代表显性和隐性等位基因。
这种设想大大增加在个体中确实看到的性状的可能组合的数目,而同时却不幸地增加了去确定哪些性状是一起遗传的困难性,因为隐性染色质单位(等位基因?)总是同显性染色质单位联结在同一个染色体上,以致经过好多世代都不能把它探查出来,直到在一种非常混乱的方式下作为一种返祖现象而可能显示出来。
Bateson和Saunders在紫罗兰属(Matthiola)实验中提到,有两种相互关联的性状可用其物质基础处于同一染色体上加以解释。“在某些组合中,①绿色种子和灰白色紧密相关;②棕色种子和grabrousness紧密相关,在其他组合中则没有这种相关”。此种结果可能由于此二性状的物质基础处于同一染色体上的缘故。当人们观察到密切相关时,可能二者都是显性;如缺乏相关性,则可能一为显性,另一为隐性。他们在另一段引言中又说:“紫花或深红色花的植株,系由绿色种子生长而成,这一规律具有普遍性。”这种情况可能是两种共同存在染色质单位恒定地表现出显性的缘故。
显性问题并非纯属细胞学概念。细胞学仅说明在一个细胞内存在着两种染色体,而这两种染色体都能表现某一特定性状,同时,有待通过实验予以说明当它们同时存在时的效应如何。实验表明,下述三种理论上的可能性都有可能实现,即①其中一方成为显性,而使其同源的另一方成为隐性;②可能产生一种折中的结果,这时可以表现出每一种染色体的作用;③两种染色体的共同作用可能产生一种完全新型的性状。孟德尔在遇到第一类的情况时,把看得出的性状(等位基因,染色质单位)称之为显性的,把另一个性状称为隐性的。Bateson和Saunders等人的实验,以及孟德尔的实验都表明,在许多情况下,只要环境条件没有起质的变化,显性性状往往在以后的世代中仍然保持显性。然而,Bateson所引证的某些实验说明显性可能起变化或有缺陷。此外,在多数(如果不是全部)情况下,每种性状都有许多不同的表现(如Bateson所说的关于人体高矮的许多不同的等位基因),它们在不同组合中表现相对的显性,这一点不仅可以想象,而且十分可能。豌豆的实验表明某些等位基因几乎永远是显性,诸如种子圆形对皱形、子叶黄色对绿色,但值得注意的是,如同多数孟德尔实验一样,这里只使用了两个相对性状。对一般相似、但某些特性表现不同的品种进行研究,肯定会得出有意义的结果。Bateson对单冠鸡、玫瑰冠鸡及孔雀之间杂交的观察,认为就是这样一种简单的方式。它将大大充实我们对显性本质的了解。
Bateson除提出许多实例说明孟德尔原理以外,还提出非孟德尔式的三种情况:①连续变异的普通融合遗传;②首次杂交后就不再分离的遗传形式;③Millardet的“假杂种”。
1.融合遗传
在这方面,Bateson明确认为这种情况可能“与孟德尔原理完全无关”,但表明它很可能与真正的孟德尔遗传有联系,他又说道:“比方说,应该承认根据某种假设,开化人种的身材这一典型持久性的性状方面,肯定存在一对以上的等位基因。可能存在有许多成对的等位基因。但我们不能肯定这种基因对和由之产生的不同种的配子的数量(甚至在身材方面也如此)是否都无限量。即使等位基因对数很少,比方说只有4对或5对,那么纯合与杂合的各种组合可能会依次排列,形成接近于连续性的曲线,其组成成分的纯度无须怀疑,实际上也无法查核。”这一假设说明,它在染色体理论方面是有根据的,在目前我们所了解的情况下,它足以将近乎连续性变异的多种情况与严谨的孟德尔遗传确切地联系起来;但另一方面,正如已经指出的,现在许多性状的个别变异,看来很可能被看做为严谨的孟德尔式变异,可以证明是由于物种本身存在有许多变异,而这种变异又被视为等位基因型,因此在象征该类型的同源染色质单位上就有相似的变异。
2.产生真杂交种的首次杂交
真杂交种的生殖细胞内显然不能有质的退化。在正常情况下,应该用母本和父本的同源染色体间存在的亲和力来说明联会问题。相反,退化则是失去这种亲和力或因某种巨力而造成中性化所致。目前,在山柳菊属(Hieracium)方面,杂交种的性状往往就是两个亲本性状的中间性状,表明双方的等位基因(或染色质单位)都在进行着工作。但在自体受精情况下,就不会减少等位基因(减数分裂)。相反,所有的组合都产生相似的后代,而它们又与亲本相似,这一事实说明了所有的生殖细胞都是相等的。Bateson在另一处提出一种看法,“如果一个等位基因单独来自父本,另一个等位基因来自母本,那么我们能得到的只能是一个由杂合子组成的品种”,这种情况粗粗一看在逻辑上过得去,似乎只要按此培育就会产生这种结果。然而,根据这样一种概念,我们无法从细胞学角度找到根据,因为,如果发生减数分裂,那么,父本和母本生殖细胞内的两部分染色体也都分裂成数目相等的两个部分。以后,因减数分裂而分离的各种各样的成对细胞融合起来,由于母本与父本生殖细胞内染色体的组合形式极其多样,致使杂种的确切染色体组很难重复。根据细胞学观点更为合适的解释是联会中的染色体联合十分牢固,牢固得无法产生减数。比如说有这么一种情况,父本和母本的染色体进行永久性融合,形成一种新的染色体,这种新的染色体以后只进行均等分裂。结果应该是,生殖细胞彼此相同,也与亲本的生殖细胞相同;因此,自体受精实际上会产生无变异的后代。如这一解释系属正确,那么联会过程显然是病态的,因此Bateson所指出的情况将往往表现“一定程度的不育性”,这就不奇怪了。
3.Millardet“假杂种”
Millardet、弗德里斯和贝特森都有过这样的实验报道:由不相似的个体杂交产生的后代只表现出亲本一方的性状,亲本另一方的性状在其后实验中永久消失。对这一现象如何从细胞学角度作出明确的解释,贝特森曾在下面一段话里有过提示:“也许可将这些现象看做是对Strasburger和Boveri观点的补充,就是说受精可能包括两个明显的过程——刺激发育与合子中性状的联合。”卵分裂而不进行原核的融合,是一种众所周知的现象,这种现象已在用氯醛(Hertwig氏兄弟)或乙醚(Wilson)处理的卵中观察到,并可设想在某些异常的自然条件下发生。然而,在所述及的实验中,两个原核都各自继续分裂。为了从细胞学上说明假杂种的产生,必须设想的不只是核的接合失败,而且还得设想其中一个完全消失。这种情况可与化学诱导的孤雌生殖或无核卵断片受精(留存的核为父本或母本)相比,但是,这段推测除非能用以指导研究,否则将是无益的。仔细研究诸如Millardet研究的草莓、弗德里斯研究的月见草属(Oenothera)以及Bateson研究的紫罗兰属杂交中的受精问题,无疑能直接得出肯定的结果。
4.嵌合体
第四种非孟德尔式情况,即嵌合体或镶嵌集合成一个组,或认为对这种组进行细胞学研究只能得到相反的证据。对于这类情况,贝特森和Saunders获得的曼陀罗嵌合体果实是一个好例子,它在一般情况下虽然表现无刺的隐性性状,但也例外地显示出刺斑。针对这种情况,贝特森说:“除非这个刺斑是这一个体的某个部分原有的斑点,否则这种现象可认为生殖细胞也许就是嵌合的。”我在这里把它理解为嵌合生殖细胞,或该承认或失败了。我试图表明,生殖细胞十之八九都是母本和父本的染色体的嵌合体,但很显然这并不是贝特森的意思。
我愿按照染色体学说的观点提出一种解释:我们已经假设体细胞染色体组具有与卵裂核为数相似的组成部分,并由后者经均等分裂而产生,它是和形成成对的同源染色体相同的方式形成的。按照这个看法,每个体细胞在它应表现的每个性状方面都应有双重的基础。按严格的孟德尔遗传,表现性状的这个生物的各个部分,两种同源染色体之一都是显性。然而,正如已经指出的,显性的染色质单位要在所有的后代中都呈显性是很不可能的事。弗德里斯的甜菜实验说明了这一点,甜菜在正常情况下为二年生植物,但往往出现少数一年生植株或“走私的”,后者被视为隐性。将这些“走私者”在不利的条件下进行培育可以增加它们的百分数,这样做证明了在这种条件下,隐性的等位基因可能变成显性。
如果每个细胞都含有母本和父本表现每一性状的潜力,如果显性并不是这些潜力中某一种潜力的一种普通功能,那就没有什么东西可以说明为什么由于某种干扰因素在一些细胞中,某一染色质单位不起作用;而在另一些细胞中,其同源染色质不起作用。这正好是贝特森和Saunders在曼陀萝中看到的那种嵌合体或Tchermak以Telephone豌豆与黄色品种杂交获得的黄绿斑豌豆。科伦斯(Correns)称之为poecilodynamous,据我所知,他对造成这种现象的原因的看法与我在前面所勾画的情况相符。贝特森在提到逻辑上的可能性时认为,像嵌合的豌豆这样的隐性斑点,可能由于配对的隐性等位基因与那些等位基因的染色体学说不相符合,因为表现隐性性状的细胞和表现显性性状的相邻细胞,这两者染色体组都是由原始卵裂核染色体的纵向分裂或均等分裂而来,因而彼此很相似。
这里提到的学说的应用情况可通过一项实验加以检验,在这项实验中,把不同的纯种杂种再进行杂交,并产生了“四分之一血统”式的第三代。如果这个学说是正确的话,那么,在这样的有机体里产生的嵌合体将表明:有一种性状像母系方面的外祖父母的性状,同时有一种性状像父系方面的原始纯种的性状。如果嵌合体的两个性状明确表明为父本性状或母本性状,那就说明这里所勾画的学说理由不充分,因为在各个世代中通过减数分裂,每对染色体之一和与之相应的性状被否定了。
在考虑两个染色体作为特定性状基础的作用时指出,在某些情况下,由不相似的等位基因组合产生的杂合子性状有时与两种等位基因都不相同,孟德尔发现高度为1英尺和6英尺的豌豆进行杂交,后代高度在6~7.5英尺之间。在讨论一些类似情况时,贝特森提请注意下列线索。如果我们大胆设想两个等位基因各自关系到一种化合物,他把等位基因的行动比喻为形成食盐时的钠和氯。化学分析结果表明,染色质的最具特征的现象是它具有大量高度复杂、变化多样的化合物——核蛋白,并如所想象的一样,如果染色体是遗传性状的特定基础,那么贝特森的设想便远远超过了有趣味的比喻范围。
从杂种不分离的现象出发,我们有理由怀疑纯杂种传递杂合子性状是由于同源染色体的永久联合。照此说来,可以很快得出结论,在染色体实际上没有永久联合的情况下,它们在同一液态解质中可能进行某种程度的化学反应。事实上这就是联合。在一般情况下,这种联合是轻微的,在一代中看不出效果,但是由于一再重复进行联合而出现的最轻微的变异——虽然方向各不相同——在长期自然选择的引导下,在染色体与其直系后裔之间,终于在某一方向出现相当程度的差异,因此在性状方面也出现联合。我们设想了同源染色体发生个别变异的原因,其理由前已述及。
最后,我们简单谈一谈某些现象,乍一看来,这些现象似乎排除了普遍应用上述结论的可能性。假如把孟德尔发现的、失去某些性状的现象看做是染色体丢失的表现,那么染色体必然是根据孟德尔定律在分裂过程中以各种各样的方式丢失的。然而在脊椎动物和有花植物——绝大多数孟德尔遗传现象是在各种脊椎动物和有花植物中得到的——经过多次实验并未见有丢失分裂。这是一个矛盾,对这种矛盾,在当初由Fick、最近由Montgomery提出的建议中,我曾大胆地提出一种可能的解释。在脊椎动物和其他生物发生联会时,染色体是边靠边的,而不是头接头的,有染色体环,就和节肢动物一样。在脊椎动物中,其初级精母细胞前期染色体平行纵裂为二,以后才进行两次分裂。两者都是纵向的,以前称为均等分裂。如果追溯到联会时精母细胞两套染色体边靠边联结的原始边线,就可以想象丢失分裂了。在我研究Brachystola的工作中,有很多现象与这一论点相符。
再者,假如正常的遗传现象取决于有丝分裂过程中完成精确的染色质分裂,那么在种质周期的任何阶段发生非有丝分裂的异常处理,其后果只能是从根本上偏离正常遗传。Meves、McGregor及其他人在两栖动物初级精母细胞方面的确已报道过这种现象。在他们所报道的这些事例中,核经过非有丝分裂之后,细胞本体似乎并不需要裂开。因此我认为这一过程在遗传上也许并不重要,因为第一次有丝分裂的片子上核膜不见了,恢复到原来的状态,染色体排列在赤道板上,就和没有受非有丝分裂的干扰一般。
在染色体作为遗传因素的论述中。没有报道过副染色体与遗传有关的结果。这种副染色体总是纵向分裂,因而很可能是均等的。它在第一次成熟有丝分裂时不分开,而是整个进入子细胞之一,普通染色体则是均等分裂的。在第二次成熟分裂时,普通染色体进行减数,副染色体此时才纵向分裂 【19】 。
我对副染色体的观察,可用来支持McClung的假说。由一个初级精母细胞产生的4个精子,其中两个有副染色体,以后参与形成雄性后代;另外两个没有副染色体,以后参与产生雌性。假如这个假说是正确的,那么性别这一性状显然是在第一次成熟有丝分裂时减数的,因为正是这一次分裂,把只能产生雄性的细胞和只能产生雌性的细胞区分开来。因此就有这么一种可能性:某一性状是在这一次成熟分裂中进行减数;而所有其余的性状则是在另一次成熟分裂中减数。作出这种安排的意义虽然不易理解,但显然是意义重大的。由每一种减数有丝分裂产生的两个细胞又成对配合,所以从各个性状的观点来看,染色体组是相对的。我们设想它有8个染色体,4个来自父系A、B、C、D,4个来自母系a、b、c、d。通过减数分裂,一个细胞含有A、b、c、D,另一个姊妹细胞则含有相对的a、B、C、d。就每一种可能的性状来说,这种相互配合的方式显然各不相同,因此一个生物所产生的精子极其多样。假如决定性别的染色质也如此减数,那么雌雄相对的两系要进入性别不同的个体中去;假如在第一次减数时,随着副染色体的不对称分布,性别减数就已完成,那么互相配对的两个成员都参与产生雄性或雌性后代,因而各种可能的染色体组合限于每一种性别之内。
后记
Guyer关于“杂交与生殖细胞”的有意义的重要报道对本文的全文来说,收到得太晚了。这位研究人员也用细胞学研究的结论来解释某些遗传现象,他对可育与不育的杂交种精子发生的比较观察,对本专题的细胞学研究来说是一大贡献。所得结论很有意义,但我想在某些方面是可以评议的。他假设“母本和父本染色体分离到不同的细胞中去,可以把它们看做是‘纯’生殖细胞,只具备一个品种的特征”。他在做这样的假设时,重犯了本文前面提到过的Cannon的错误。文中没有提到孟德尔法则,但考虑到了杂种鸽经过近亲交配得到的母本类型。作者说了这样一段话,说明他是精通孟德尔原理的,他说:“第三代一般都回复祖代的原有颜色。”有时所有的特征与祖代的一方很相似,那么纯生殖细胞的概念得以适用,这是很明确的;而作者对于杂种近交后代表现混合性状显然也很熟悉。他倾向于用以下两个方面加以解释:①“分别代表两个原始种的两个细胞的联合,将产生混合型后代。”②“除通过刚才说的混合方法外,在某些情况下,个别染色体的不均等分裂并非罕见,也可能引起变异性问题,通过这种情况,在某些成熟的生殖细胞中,可能出现不同比例的两个亲本品种的染色质。”
上述第一种解释与孟德尔实验的结果相符,可是它错在用于(不具备细胞学依据)全部性状或染色体、而不是用于个别性状或染色质。至于引用的第二段话,可能没有多大问题,就是说,染色体的不正常分裂会产生变异;正如Guyer本人所观察到的,这些不正常现象会根据不育性的程度而增加。因此,可以很自然地得出结论说,它们不仅是病理性的,而且可能部分由于不育的缘故。此外,根据Guyer所接受的染色体个体性的假设,由于不正常分裂而使染色体失去一部分,将会是永久性的;而且,一再丢失一部分染色体,将使单个染色体组(他把它看做是一个整体)的染色物质明显衰退,必然迅速导致功能恶化,终于不育。
正如已经指出的那样,对形成变异的两种解释中的第一种,对一对染色体的后代来说,只有4种可能的染色体组合。但我们知道,尽管有无数后代,但实际在后代当中从不出现重复,同卵双生的情况除外。因此,不管后代的数量有多少,所有的变异,除4种正常染色体组合的情况外,都应按照明显趋于不育这种病态分裂程序来考虑。然而我们从Bateson和Saunders给进化委员会(Evolution Committee)的报告中发现有这么一句话:“我们还没有听说孟德尔式遗传中可育性受到损伤的情况。”我们认为,这些研究人员的大量实验系属孟德尔遗传,我们将这一证据与Cannon的论证联系起来,说明各种棉花杂种的成熟过程若不是正常,就必定是明显地不正常,以致必然不育;还使之与Guyer自己的认可联系起来,Guyer认为,有丝分裂中的不正常现象随不育性程度的加深而增加,而平衡状态是杂种变异的正常因素,病态有丝分裂的效应则相反。
参考文献
[1]Sutton, Walter S., "On the Morphology of the Chromosome Group in Brachystola magna,” Biol. Bull., IV., 1, 1902.
[2]Mendel, Gregor Johann, "Vesuche über Pflanzen-Hybriden,” Verh. naturf. Vers in Brünn IV., and in Osterwald's Klassiker der exakten Wissenschaft. English translation in Journ. Roy. Hort. Soc., XXVI., 1901. Later reprinted with modifications and corrections in Bateson's "Mendel's Principles of Heredity,” Cambridge, 1902, p. 40.
[3]Wilson, E. B., "Mendel's Principles of Heredity and the Maturation of the Germ-Cells,” Science, XVI., 416.
[4]Bateson, W., "Mendel's Principles of Heredity,” Cambridge, 1902, p.30.
[5]Sutton, W. S., loc. cit.
[6]The conclusion that synapsis involves a union of paternal and maternal chromosomes in pairs was first reached by Montgomery in 1901. Montgomery. T. H., Jr., "A Study of the Chromosomes of the Germ-Cells of Metazoa," Trans. Amer. Phil. Soc., XX.
[7]Cannon, W. A., "A Cytological Basis for the Mendelian Laws," Bull. Torrey Botanical Club, 29, 1902.
[8]Absolute proof is impossible in a pure-bred form on account of the impossibility of distinguishing between maternal and paternal members of any synaptic pair. If, however, such hybrids as those obtained by Moenkhaus (Moenkhaus, W. J., "Early Development in Certain Hybrid Species," Report of Second Meeting of Naturalists at Chicago, Science, XIII., 323), with fishes can be reared to sexual maturity absolute proof of this point may be expected. This observer was able in the early cells of certain fish hybrids to distinguish the maternal from the paternal chromosomes by differences in form, and if the same can be done in the maturation divisions the question of the distribution of chromosomes in reduction becomes a very simple matter of observation.
[9]Boveri, Th., "Ueber Mehrpolige Mitosen als Mittel zur Analyse des Zellkerns," Verb. d. Phys.-Med. Ges. zu Würzburg, N. F., Bd. XXXV., 1902. It appears from a personal letter that Boveri had noted the correspondence between chromosomic behavior as deducible from his experiments and the results on plant hybrids, as indicated also in reference 1, 1.c., p.81.
[10]Cannon, W. A., loc. cit.
[11]Bateson and Saunders, Experimental Studies in the Physiology of Heredity (Reports to the Evolution Committee, I., London, 1902)p.81, paragraphs 11 and 12.
[12]Cf. Bateson and Saunders, loc. cit.
[13]Ibid.
[14]Cf. Mendel's experiments on Hieracium.
[15]Bateson and Saunders, loc. cit., p.154.
[16]Cf. Bateson and Saunders, pp.135,136.
[17]Bateson and Saunders, p.156.
[18]Fick, R., "Mittheilung ueber Eireifung bei Amphibien," Supp. Anat. Anz., XVI.
[19]Montgomery, T. H., Jr., loc. cit.
[20]It is of interest in connection with this question that there occurs regularly in each of the spermatogonial generations in Brachystola a condition of the nucleus which suggests amitosis but which in reality is nothing more than the enclosure of the different chromosomes in partially separated vesicles. Cf. Sutton, W. S., "The Spermatogonial Divisions in Brachystola Magna," Kans. Univ. Quart., IX., 2.
[21]McClung, C. E., "The Accessory Chromosome—Sex Determinant?" Biol. Bull., III., 1 and 2, 1902. "Notes on the Accessory Chromosome," Anat. Anz., XX., pp.220-226.
[22]Guyer, M. F., "Hybridism and the Germ Cell," Bulletin of the University of Cincinnati, No.21, 1902.
[23]Cannon, W. A., loc. cit.
昆虫染色体与性决定的关系 【20】
埃德蒙·威尔逊(E. B. Wilson) 【21】
(1905年)
去年夏天所得到的材料十分清楚地证明,半翅目昆虫的性别在染色体组中表现出稳定和特定的差别,这种差别无疑是这样一种本性:这些动物在染色体和性决定之间存在着某种特定的联系。这种差别有两种,一种是雌性细胞比它的雄性细胞多一个染色体;另一种是雌雄性细胞染色体数目相同,但雄性细胞有一个染色体比雌性细胞相应的那个染色体要小得多(这同Stevens对粉虫属的观察是一致的)。为方便起见,把这两种类型分别称之为A和B。每一种类型都有三个属,已确定这是确切的事实。A类为Protenor belfragei、南瓜缘蝽(Anasa tristis)和蛛缘蝽(Alydus pilosulus);B类为Lygoeus turcicus、蝽Euschistus fissilis和Coenus delius。染色体组是用雌性分裂卵原细胞和卵巢卵泡细胞以及雄性分裂精原细胞和裸细胞来检验的。
A类(自从Henking 1890年在Pyrrochoris方面的文章发表后已知道)精子有两种,一种精子比另一种精子多一个染色体(所谓副染色体或异向染色体)。这一类雌性体细胞染色体数目为偶数,而雄性体细胞染色体数目要少一个,为奇数。Protenor和蛛缘蝽(Alydus)的实际数字为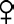 14、
14、 13,南瓜缘蝽(Anasa)为 22、 21。研究两性染色体组还看到以下事实:在雌性细胞中,所有的染色体双双成对排列,每对由两个大小相等的染色体组成,这在Protenor漂亮的染色体组看得最清楚,其染色体大小差别十分明显。雄性细胞的全部染色体,除一个染色体无配偶外,其他都对称地配对。这个无配偶的染色体是副染色体或异向染色体,由于它无配对,只能进入一半精子 【22】 。
13,南瓜缘蝽(Anasa)为 22、 21。研究两性染色体组还看到以下事实:在雌性细胞中,所有的染色体双双成对排列,每对由两个大小相等的染色体组成,这在Protenor漂亮的染色体组看得最清楚,其染色体大小差别十分明显。雄性细胞的全部染色体,除一个染色体无配偶外,其他都对称地配对。这个无配偶的染色体是副染色体或异向染色体,由于它无配对,只能进入一半精子 【22】 。
B类的全部精子都含有相同数目的染色体(雌雄两性的染色体都是体细胞染色体数的一半)。但尽管如此,它们仍有两种:一种有一个大的性染色体,另一种有一个小的性染色体。两性的体细胞染色体数目一样(上述三个例子为14个染色体),但有以下方面的区别:雌性细胞(卵原细胞和卵泡细胞)的全部染色体像A类一样,双双成对,数目一样,不出现小的性染色体。雄性细胞除两个染色体外,也是双双成对。这两个染色体是不相等的性染色体,而在成熟过程中它们的分配情况是小的一个性染色体分到一半精子中,大的一个性染色体分到另一半精子中。
这些事实使我相信只能有一种解释。既然雌性细胞(卵厚细胞)的全部染色体可以对称成对,毫无疑问,这个性细胞的联会产生对称的二价染色体的减半数目,其结果是所有的卵都收下同样数目的染色体。这个数目(Anasa为11,Protenor或Alydus为7)同含有副染色体的精子的情况是一样的。显然,这两种精子都是有功能的,A类则雌性细胞是由卵同含有副染色体的精子受精产生的,而雄性细胞是由卵同没有副染色体的精子受精产生的(McClung所作推测的颠倒)。这样,如果n为雌性体细胞染色体数,n/2是所有成熟卵染色体数,那么有一半精子(含有副染色体的精子)的染色体数为n/2,而另一半精子为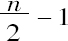 。因此在受精中:
。因此在受精中:
首先由Montgomery指出的Protenor的情况完全说明这样解释是可靠的,Protenor的副染色体在每一个时期都能根据其巨大的体积而能毫无错误地予以识别。精原细胞分裂总是出现只有一个大染色体,而卵原细胞分裂则看到一对完全相似的染色体。雌性细胞的这对染色体中的一个在受精时必然来自卵核,而另一个染色体(显然指那个副染色体)来自精子核。所以,很清楚,所有成熟卵在受精之前必须有一个作为雄性副染色体的母本配偶的染色体。而雌性细胞是由卵同含有相似染色体组的精子(即含有副染色体)受精产生的。雄性体细胞核只有一个大染色体(副染色体),只能意味着雄性是由卵同没有这个副染色体的精子受精得到的,以及雄性的这个单个副染色体是在受精时来自卵核。
B类的所有的卵必须有一个相当于雄性巨大性染色体的染色体。当含有巨大性染色体的精子同卵受精产生雌性,而当含有小性染色体的精子同卵受精则产生雄性。
上述有区别的A、B两类很容易变成一类。如果B类的小性染色体消失掉,所发生的现象就同A类一样。有点怀疑这种现象是否为A类的真正起源,以及副染色体最初为一巨大性染色体,其较小的配偶染色体消失了。这样,副染色体的不配对特性就找到一个完整的解释,而副染色体的配对行为又使其明显地丧失掉异常的特性。
以上事实必然地得出结论:在染色体和性决定之间存在着某种因果关系;头一个想到的结论自然是McClung在副染色体情况下推测的,性染色体和异向染色体实际上是性的决定子。然而,分析将表明,不论哪一种有关这些染色体明确地是雄性或雌性决定子,会碰到巨大的、即使并不是不可克服的困难。更可能的是,由于将来会提出的种种理由,染色体组在两个性别中的活动、卵和精子间的差别主要为活动程度或强度上的差别,而不是活动种类上的不同,我们或许在这里找到有关性决定一般理论的一点线索,它同半翅目昆虫 【23】 所看到的实际情况是一致的。提出这一问题的一个重要事实是在这两种类型中,两性的差别在于联会和生长期间性染色体或副染色体的行为,这些被认为是雄性的染色体形成浓缩的染色体核仁,而雌性染色体则像其他染色体一样,核仁呈扩散状态。这表明,在联会和生长期间雌性染色体比雄性染色体在细胞新陈代谢中作用更活跃。因而,生殖细胞分化的首要因素或许是一种新陈代谢,或者是一种生长。
细胞核的化学成分 【24】
阿尔布雷希·科塞尔(A. Kossel) 【25】
(1910年)
19世纪有机化学的发展,主要建筑在建立原子空间排列的概念。人们都知道,有机化学家既能提出有机物成分及其化学反应方面的知识,又能站在化学系统的地位上,建造出一幅种类不同的原子空间分布图,从而清楚又准确地提出他的观点。
人们一旦把这些概念用于动物或植物组织的研究中,就会推导出该有机物的化学结构图。以这种方式开辟的科学领域能在许多方面同生命物质的解剖学研究相媲美。
解剖学和化学这两门学科,看来首先是仅仅致力于描述有机物。但是在这两种情况下,这种描述都远远不是真正的研究目的。据我们看来,解剖学和化学结构的知识是唯一有价值的,因为从这些知识我们渴望了解各部分的功能、它们的发育机理或其他的生物学重要问题。
因此我们只能作为取得更多知识的起步来估价细胞和原生质成分的经验。截至目前,我将要报告得到的结果,确实更适合于激励我们渴望得到更多的知识,而绝不是高兴一场而已。从考虑器械的各个片断到懂得它的活动方式,还要走很长的路。
比较观察得出这样的概念:某些化学的生命过程为动物和植物所共有;某种程度上,有一种化学机理以其共同的原理在各种生命物质中起作用。这些基本的生理过程必须位于这样一种物质,看来无论在什么地方它都是生理氧化过程的主要集中点,与此同时,身体的其余部分即原生质从这一点出发的。
显然,必须把这种结构的化学研究看成是生物化学最重要的问题之一,但这类研究在第一次分析时,首先是材料的选择和制备方面,就显出其困难性。细胞的生理活性、细胞的可见或不可见的营养产物,几乎都是指活细胞而言的。成分和内含物间的差别、有机体物质和化学代谢物之间的差别都很难规定,而只能把某种程度上可予肯定的结果寄希望于仔细的组织学鉴定和比较研究的基础上。这样就研究了多种多样的细胞结构和无一定形状的原生质,并把成分一览表内反复出现的各个化合物组规定下来,鉴于Hoppe-Seyler在核素、卵磷脂、胆固醇和钾盐方面的工作,除蛋白质外,把这些化合物都增添在一览表中。
当人们试图把细胞核引入这些研究的范围时,从此开辟了新的前景。这里我们有一个细胞器,其结构和功能必然同生命的普遍过程有联系。从以下各方面说明上述论点已经清楚,即它的结构状态,它在细胞分裂前和伴随着细胞分裂过程在形状上的变化;它在动物、植物世界不同地区的再现;以及它基本上与种和群或有机界系统的位置无关。现在又对这个器官的形态学特性以及化学特性,它把该器官的特点规定得更加明显,因为即使在核的结构没有确定的细胞里也能把它识别出来。现在我将扼要地概括一下这些化学特性。
1860年,Hoppe-Seyler实验室对脓细胞核的研究开始了这一领域的第一个观察。Hoppe-Seyler的一名学生米舍尔(Miescher)能分离出这些核,他发现核里有一种非常富有磷的物质,他称这种物质为核素。在一种组织里发现有一个进一步开展这项工作的适宜对象,即精子的头部。这种结构是通过细胞核的转化而发育,并保持其化学成分,它显然也是其生理功能的一个重要部分。已经积累的证据说明核素或核材料的确是细胞核所特有的。还发现别的研究对象在某种程度上可以分离细胞核,例如鸟类的红细胞,其细胞体可溶于水。对上述分离出来的一批适量的核也能进行化学研究,并再一次发现核物质的明显特点,而显微化学测验则证实了这一点。它们同时证明核材料属于核物质的一种规定良好的部分,这部分在转化过程中以一种非常引人注目的方式而显得突出,它的量在不同核中有变化,并因为它的某些染色反应而取得染色质之名。这方面唯一的困难是在那些没有细胞核的动物产物,如卵的卵黄囊、牛奶的酪蛋白中寻找核物质,在用更精确的化学研究予以澄清前,的确曾尝试用专门的假说来解释这些事实。
这些核物质的化学结构,在原生质的许多有机成分中,特别在那些积极参与代谢过程的有机成分中,曾发现过它们的一些特点。曾观察到这种成分很容易分解成一定数量的封闭的原子基团,它可与积木相比拟。这种“积木”以大量数目和多样化的方式装配在一起,并显然按照一特定的计划形成蛋白质和淀粉,糖原的分子,以及用较少的数目装配成脂肪和磷脂的分子。营养物复杂的有机成分,当它们准备被消化以便进入体内时,分解成这种积木,然后在有机体内再把这些积木重新装配成大分子。
核物质也是这种组成,化学分析表明,首先在许多情况下核物质分解成两部分,其中之一有蛋白质特性。这部分除正常的蛋白质外,不具有其他原子团。然而,另一部分有特殊的结构,已给它命名为核酸。我们从核酸成功地得到一些碎片,这些碎片即使用温和的化学作用也能把它部分地从分子中溶解出来,并根据一种相当特异的氮原子集中而能予以识别。这里一起出现4种含氮基因:胞嘧啶、胸腺嘧啶、腺嘌呤、鸟嘌呤。
这四种物体之一的鸟嘌呤,在各动物组织中已为人们所知道,并为Picard在鲑的精子中发现,虽然这位研究工作者的确没有怀疑它同核素有任何遗传关系。早先时候普遍采纳鸟嘌呤和其他类似物质来自蛋白质分子,米舍尔则想到这些物质可能从鱼精蛋白产生,而Picard提出的意见是“在鲑精子中它们是预先一起存在的”。它们起源于核酸的见解出乎意料地从一开始就碰到了强烈的反对意见,却在同时弄清了一种尚未找到解释的特殊现象。例如,曾注意到在白血病中,鸟嘌呤和有关物质在血液中大量存在。现在这种疾病的特点是含有核的成分取代了无核的红细胞,但前者分解成很大的数目,从而使得体液为这种核物质的分解产物所淹没。从此,上述碱基或同它们关系非常密切的转化产物大量存在于体液中。还有前面提到的矛盾,即假定在卵黄和牛奶中有核物质,这一点现已得到解决。一项更精确的研究证明,这些成分由于其表面行为和含有磷,过去被认为是核素,却具有不同的化学结构。我称之为富有氮的积木完全不存在,因此它们不属于真正核物质一类,而形成特别的一类。
越是弄清楚含氮丰富物质与细胞核的关系,氮和碳原子在分子中的排列问题也越显得突出。4个物体中有2个,腺嘌呤和鸟嘌呤,属于当今通常包括在称之为四氧嘧啶衍生物或嘌呤衍生物的一类化合物。这类化合物各个成员的被发现及其化学性质的得到阐明是同Schecle、Torbern、本杰明(Bergmann)、Wöhler、李比希(Liebig)、Strecker和阿道夫(Adolf)、拜尔(Baeyer)的名字联系在一起的,而费歇尔(Emil Fischer)的著作已对这一系列杰出的研究作了总结,该著作最终确立了令人满意的化学式。另外两个胸腺嘧啶和胞嘧啶组成较简单;分解和合成实验得出结果:胸腺嘧啶必须符合下列方式的碳原子和氮原子的组合:
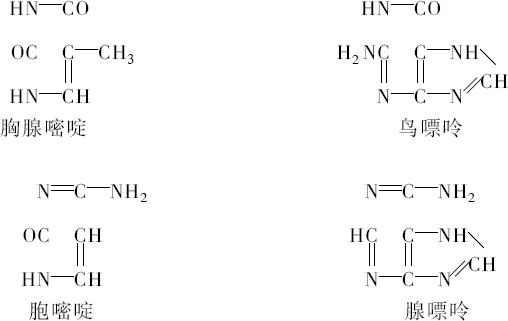
从上式看出,必须认为胸腺嘧啶和胞嘧啶是一种碳和氮原子的环状系统。可以确定原子在胞嘧啶里的位置,因为在一种氧化剂的作用下,这种物质分解成双缩脲和草酸,并可以立即用它的合成阐明其组成。同这一称之为嘧啶环的简单环相反,腺嘌呤和鸟嘌呤的化学式是一种双环,即所谓嘌呤环,说明它的氮原子集中得还要多些。
在这4种核酸分子碎片中,看到碳原子和氮原子是按照同一个基本计划进行搭配的。嘌呤环的出现就好像它是嘧啶环的一种结构增建的结果。如果现在把已知的这4种嘧啶和嘌呤衍生物置于较强烈的化学反应,或者在体内追踪它们的行为,能看到那些连接在一起形成环的碳和氮原子,要使它们彼此分开相当困难,相反,额外连接在环上的其他原子,例如NH2 基,则可通过引进水的成分把它们拆开。在这种方式下产生的衍生物称为次黄嘌呤、黄嘌呤和尿嘧啶,而尿嘧啶常常发现在腺嘌呤、鸟嘌呤和胞嘧啶的附近。此外,还有其他物质作为动物新陈代谢的最终产物出现。
从现在起,我们对核酸分子的一部分,即含氮部分,已有某种程度的了解;还有一个由两个不相似的成分组成的剩余物,其中之一含有6个碳原子,它由氧和氢;以碳水化合物所特有的方式相连,另一个不含碳的成分是磷酸。
一旦对核酸中存在的这样一个大分子结构各积木块的性质得以肯定,就出现两个新问题:每一块积木的相对量是多少?它们是如何相互排列的?第一个问题已由H. Stendel的研究得到解决。根据他的分析,我们已能认为这4个富氮基团中的每一个基团都有两个碳水化合物分子和一个磷酸分子。目前第二个问题尚未得到恰当的答案。只有一次观察的结论是碳水化合物团和富氮物体间有一种联系,即如核酸小心地分解,仍然发现这两个碎片连接在一起,植物新陈代谢中也发生这种结合。
根据我们就当代的见解和意见所作的这一肤浅的评述,核酸看来是一种复合体,它至少有12个积木,但在活细胞里,它的结构很可能要大些,因为从一些观察提出,在器官中这样的复合体有几个是彼此结合在一起的。
我打算谈一种在动物机体的某些细胞中含有的核酸,但它不是典型核酸基团的唯一形式。对不同有机体和同一个体不同器官的研究,证明了这类物质的结构有重大差别。犹如我们在蛋白质、脂肪、胆酸和许多其他生物产物已经知道的那样,在核酸中反复出现的相同现象,即用以证明同一结构思想的各种物质的整个系列,是通过多种多样的途径发展起来的。
我所概括的核酸结构在其他器官中以更简单的方式重复出现。例如,酵母的细胞中发现有一种核酸缺少4个含氮基团中的一个基团,即胸腺嘧啶,并且同6个成员的碳水化合物环相反,它含有的碳水化合物是5个成员。次黄核苷酸和鸟核苷酸的组成更简单。前者早为李比希发现,但Haiser首先弄清楚它的化学性质,它存在于肌肉中,在4个含氮物质的地方只有单独一个含氮物质,而且它的形状有点改变,碳水化合物也只有5个碳原子。鸟苷酸的结构相似,Olof Hammarsten和Jvar Bang首先发现了这种物质。它也只有一个含氮基团,即鸟嘌呤,并也有一个由5个碳原子成员的链作为一个碳水化合物同鸟嘌呤和磷酸连接起来。
生物学家们把兴趣放在这些物质上,这由于它们已被承认是核酸组的最简单的成员,这一点完全可以理解。现在的见解仍然是,在顺利地具备种种简单而易于掌握的方法之前,首要的是研究方法错综复杂并难以辩论。我们并不知道,次黄苷酸和鸟苷酸是否像复杂的核酸那样对细胞生命起同样重要的作用;尤其是目前,这最后提到的两个酸,其位置是否坐落在细胞核的染色质中,尚未确定下来。
我在前面已经提到,复杂的核酸是在这种同蛋白质结合的形态上如此重要的结构中发现的,而这种结合的方式则多种多样。在有些器官中,发现这两种成分结合得比较松散,其行为像一盐类,并容易把酸和蛋白质从它那里分离出来。另外一些细胞,核酸和蛋白质之间结合牢固,它对化学分离物强烈抵抗。在鸟的红细胞的核里可找到这种盐状物,而我已经说过,当红细胞溶于水时,能把核分离出来。然后这种细胞核物质同一些结合的基质像一团不溶的物质一样留在后面。如使这团核同稀释酸接触,大部分蛋白质就被溶解,而把核酸留下来。胸腺、淋巴腺和脾腺之类腺组织的细胞,也发现相似的松散结合,并且所有这些组织也有一部分同蛋白质牢固地结合,另一部分则松散地结合。精子头部从其起源和组织学性状来看,的确是细胞核,它的行为引人注目。可以设想,不同的动物种,在它们具有同一功能的一种器官中,将会找到相似的化学关系,但蛋白质-核酸的结合方面则并非如此。在一些至今仍然只在为数不多的热血动物种所作的研究中,发现同无脊椎动物的核酸与蛋白质松散结合的情况相反,热血动物的精子则牢固结合,可能大多数情况都是如此。鱼的精子很像鸟的红细胞的核,至今经常发现的只是一种松散的结合,虽然还不能就此断定是否也有结合牢固的。
具有核酸松散结合的核将说明另外一个值得注意的现象,例如,同核酸结合的蛋白质的特殊安排。它们具有一种有机碱的性质。蛋白质被牢固结合的核,其化学反应敏感性要差得多,下面不再另作介绍。
为了把蛋白质分子变换成一个可以理解的基础,我将扼要介绍这类对有机界如此重要的物质在其化学结构方面的主要特性。
如上述细胞的碳化合物一样,蛋白质是由许多连接基因,即所称积木制成,这里所说的积木我指的是一种直接相连的碳原子复合物。当这些碳原子相连接的地方为其他的原子打断时,这些积木就常常拆开,这时候大分子就在有机体内或有机体外分解。这种大积木的蛋白质,其总数迄今确实肯定的最高数为9个,和它们直接牢固结合的碳原子数可能是12个,但在多数情况下,这种基因要小些。通常由一个氮原子把这些基因彼此联合起来,与此同时,这个氮原子同一个氢原子相连接而形成一个所谓“酰亚胺”基。这种连接方法主要是通过费歇尔的工作确定下来的。在特殊情况下会发生别的连接法,例如,E. Baumann发现的二硫连接,它是由两个相互连接的硫原子把两个碳链连接起来。根据K. A.H. Mörner的工作已知为蛋白质分子的一个成分——胱氨酸,就是这种情况。如果现在蛋白质分子分解了,这常常随着水分子的引入而发生。
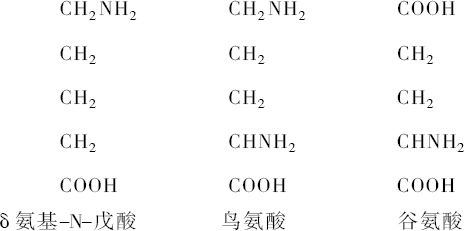
把这些积木本身从整个分子的结构中释放出来,在它们中间至少能识别出19种。在涉及其内部结构之前,这些积木或碎片的大多数都遵循一个共同的原理。几乎所有这些碎片都具有氨基酸的特性。人们可以拿氨基戊酸作为这种物质的一个例子。它含有一个同氢、氧和氮原子连接起来的碳原子链。这些物质的第一个特点是COOH基,它给物质以酸的性质;第二个特点是NH2 基,它的存在使物质具有碱的性质。现在我们所知道的氨基酸,如所举例的δ-氨基-N-戊酸,其COOH基和NH2 基的数目相等;也有其他物质多一个NH2 基或多一个COOH基。后者为酸的性质,前者称二氨基酸,碱的性质占优势。
然而,构成蛋白质的氨基酸的多样性,不仅由于它们在COOH基和NH2 基数目上的变动,也由于连接在一个链上的碳原子数不一而造成。我们从蛋白质分子能得到2、3、5或6个碳原子的链。通过引入一个氧原子或硫原子,使氢原子同碳原子分开,或因为有一个复杂的有机基团如3C、2N和3H取代了H原子的位置,这些都可带来更多的变异。

在构成蛋白质的一系列积木中,发现这些氨基酸也有种类大不相同的原子基因,这种氨基酸含有一个碳原子和两个氮原子,并且这一基团在分子中总是同前面提到的二氨基戊酸相结合。这种脒基同二氨基戊酸或鸟氨酸相结合而命名的精氨酸,是由E. Schulze发现,并由S. G.Hedin证实它是蛋白质的一个成分。
蛋白质就是由这样一些积木构成的。我们不清楚为什么每一个积木在整个结构中如此经常地反复出现,但我们能确定形状不同的积木其总数之间的相对比例。例如,我们能确定同一氨基酸相比,二氨基酸的量有多大,以及在二氨基戊酸形式中存在的总氮量百分率是多少。尽管从这些比例不能得出有关积木排列的任何想法,但它早已表明迄今所研究的蛋白质之间的重要差异,它还说明,在它们中间,前面提到的细胞核结合松散的蛋白质占有相当特殊的地位。
这些核蛋白质的特性由以下事实决定,即在它们的结构方面,某几种积木如含氮丰富的基团的量更多些。这就是说,例如同其余的蛋白质相比,它们含二氨基酸较多,特别是二氨基戊酸和同它连接的脒基;在它们中间也能找到大量的组氨酸。
这些含氮基团之插入蛋白质分子还使碱基因强烈地呈现自由反应的状态。
例如在鸟的红细胞核中发现有这种蛋白质,而且我已经提到,用稀释的无机酸很容易把它分出来,这种蛋白质称之为组蛋白。相似的物质以一种同核酸结合在一起,像盐类那样广泛地分布于高等动物和低等动物的组织里面。无脊椎动物,如头足网海胆的精子和一些鱼类的精子也有这种物质。我可以引用不同种的鳕作例子,从鳕的精巢里我们得到一种组蛋白,其化学性质和成分同那些从鸟类红细胞或胸腺中得到的组蛋白非常相似。
这些同核酸结合松散的组蛋白进一步说明通常结构复杂的蛋白质的性质。它们唯一的区别之处是一个特异的性质,即游离的碱基团占优势。
如取其他鱼的精巢作同样研究,得到的是组成简单得多的物质,它在精子的头部取代了组蛋白的位置,这些物质是鱼精蛋白。
这里我不准备谈从整套观察所形成的意见,这些碱性蛋白是在发育过程中通过正常蛋白质的转化而产生的,在这个过程中,含氮少的基团逐渐从它们中被分解掉。这种转化或多或少是广泛存在的,它把正常的蛋白质首先转化成组蛋白,如果这种消失过程继续进行下去,我们就得到鱼精蛋白。这样鱼精蛋白同组蛋白相比,其一氨基酸少,二氨基酸相对多。但鱼精蛋白彼此间也有差别,并显然是通过一些中间步骤把它同组蛋白连接起来。例如从鲟卵得到的鲟精蛋白,含有前面提到过的4种蛋白质分子的富氮基团:两个二氨基酸中有一个同脒基相结合,而另一个同组氨酸相结合。其他的鱼精蛋白至少含有两个或3个已知的碱基团。在某些鲑(Salmonidae)的精子头部,蛋白质分子组成的变动显著减少,其整个分子限制在5种不同的积木。其中两个,即二氨基戊酸和脒基,是氮的主要载体,其数量大大超过其余的载体,约携带总氮量的88%。
这样,在这个特有的转化中,越来越多的碳链消失掉,这种碳链对建立大多数蛋白质十分重要,从而形成其主要部分的含氮量减少,与此相反,出现了一种C和N交替排列的基因。我们已经看到这种排列也存在于细胞核的另一个成分即核酸中。
如果我们现在对结合松散的核素物质进行小结,其结果如下:细胞核染色质的组成来自两种成分,一种成分富有磷酸并具有酸的性质;另一种为具有碱基性质的蛋白质。这两种成分在化学结构方面明显的相似之处是氮原子积聚惊人,并由于这种化学结构,染色质组成物同其余的细胞成分能区别清楚;显然,必须把这种性质同染色质物质的功能联系起来。这些富有氮和含磷的原子团是染色体里的贮存物,在细胞分裂期间它最先活动起来,而它对别的细胞的传递将构成繁殖过程中的一个重要部分。
在这一点上,我们已经到达只有通过用各种研究方法一起进行工作,才能解决问题的时候了。形态科学的代表性工作是在显微镜下观察细胞内的结构,并研究它的形状同基本的有机体状态的依赖关系。生物化学家们则规定这个结构的组成,它在化学系统中的地位,以及它与细胞的其他化学成分的关系。但这项工作需要结构化学的理论并借助于合成方法。
这样,我今天要描述的结果是从不同研究单位得到的,如果要对全体作出贡献的人表示谢意的话,则将提到许多人的姓名。
孟德尔遗传的随机分离与相引 【26】
托马斯·摩尔根(T. H. Morgan) 【27】
(1911年)
孟德尔遗传定律在于假定单位性状因子的随机分离。孟德尔遗传所特有的两个或两个以上性状的典型比例,诸如9∶3∶3∶1等是根据这种假定作出的。近年来看到一些情况,当涉及两个或两个以上的性状时,其比例同孟德尔随机分离的假定不符合。其中最突出的例子是在Abraxas和果蝇,以及几个家禽品种的性限制遗传中发现的,必须认为这种性限制遗传,在雌性因子同另一个因子之间发生一种相引,豌豆花粉的颜色和形状也是如此。除这些例子外,Bateson和他的同事(Punnett, DeVilmorin和Gregory)最近发表了一些新的例子。
为了说明其结果,Bateson认为生殖细胞不仅有相引也有相斥。这些事实看来完全可以同我在果蝇方面发现的情况相比拟,此后这些结果使我得出非常简单的解释,我想尝试着把Bateson的假设同我提出的假设作一比较。
Bateson解释所依据的事实,用他自己的话来说可以简述如下:“如果A、a和B、b,是属于相引和相斥的两个等位基因对,那么由Ab×aB结合产生的杂合子的配子发生,A和B将彼此互斥;但由AB×ab结合产生的杂合子的配子发生,A和B又将相引。”“我们还不能推测出这一特性的重要性质,而至今全部能说的是,在这些特殊情况下,性状在杂合子中的分配受原始纯合亲本中性状分配的影响。”Bateson进一步指出,由于“鸡的性别至少是三个其他因子的排斥者……可以发觉其中有些因子比其余因子领先,其方式是作为一种结果,用下一次的相引来取消这一次的相斥”。
在吸引、相斥和领先的次序以及相引的精细系统方面,我根据果蝇眼色、体色、翅突变和雌性因素的遗传结果提出了一个比较简单的解释。如果代表这些因子的物质包含在染色体里面,又如果这些相引的因子彼此紧挨在一个直线系统上,那么当亲本对(在杂合子中)接合时,相似区域就会处于对立状态。有充分证据支持下述观点:在绞线期,同源染色体彼此缠绕在一起,但当染色体分开(分裂)时,分裂是在Janssens主张的一个单平面上进行的。其结果是,相距近的原始物质更可能落在同一个半面上,而相距更远的区段则可能最后落在同一个半面上,另一边的情况也是如此,因此,我们只发现某些性状的相引,而很少或根本没有找到其他性状相引的证据;这种差别取决于代表因子的那些染色体物质彼此间在直线上的距离。这种解释将说明我所看到的所有的许多现象,而且我想,将同样能阐明截至目前谈到的其他情况。这种结果是染色体里物质的位置和同源染色体联合方法的简单的机械结果,它所产生的比例用数字系统来表示,同染色体因子相对位置表示一样都不大。除了孟德尔观念的随机分离外,我们发现染色体位置相近的因子联合。细胞学将提供实验证明所需要的机理。
果蝇的六个性连锁因子由其联合方式所表示的直线排列 【28】
埃利弗利德·斯特蒂文特(A. H.Sturtevant) 【29】
(1913年)
历史
瑟顿(Sutton)首先指出染色体在减数中的行为同孟德尔因子在分离中的行为,两者之间存在着一种平行关系(1902),而同年早些时候Boveri(1902)已提到一种可能的联系,Boveri在这篇和另几篇文章中,从实验胚胎学的领域里引来了重要的证据,说明染色体在发育和遗传中起着重要的作用。McClung(1902)关于副染色体是一个性的决定者的假设,是第一次尝试把某个体细胞性状同一个特定的染色体联系起来。Stevens(1905)和Wilson(1905)通过以下证明证实了这一点,性染色体形状多种多样,它存在于所有的卵和产生雌性的精子里面,但在产生雄性的精子里面,或没有这个性染色体或由一个较小的同源染色体替代它。当摩尔根证明果蝇(Drosophila ampelophila)的眼色因子,遵循Stevens(1908)在这同一个种里面已经发现的性染色体的分配时前进了一大步。以后,关于出现果蝇性连锁的翅突变,摩尔根(1910、1911)弄清楚了一个新的起点。把白眼、长翅果蝇同红眼和痕迹翅(新的性连锁性状)果蝇杂交,他在子二代得到了白眼、痕迹翅的果蝇。这只能在交换是可能的情况下发生;这意味着,根据这些因子都在性染色体上的假设,发生同源染色体之间物质的互换(只在雌性发生,因雄性蝇只有一个性染色体)。当时没有引起注意,以后才提出的一个要点是同果蝇的其他性连锁因子有联系(Morgan,1911)。显然,有些性连锁因子是联合在一起的。这就是说,交换并不是在一些因子之间随意发生的,事实证明子一代果蝇出现同一性状的新组合,较之子二代果蝇多得多。从染色体的观点来看,这意味着这个染色体,或至少是该染色体的某些节段在减数期间比它们互换材料的那部分更可能保持其完整性 【30】 。摩尔根(1911)根据这些事实提出了一个有关相引的生理基础的假设。他采用Janssens(1909)的交叉型假说为机理。他是这样说的(Morgan,1911):
如果代表这些因子的物质存在于染色体中,而那些相引的因子又在一直线排列上彼此紧挨在一起,那么当亲本染色体对(在杂合子中)结合时,相似的区段将保持着方向相反的位置。有足够的证据支持这种观点,这就是在绞线期同源染色体彼此缠绕在一起,但当染色体分开时(分裂),如Janssens主张的那样,它是在一个单平面上进行分裂的。其结果是相距短的原始物质更可能落在同一个半面上,而距离更远的区段则可能是最后一个掉在同一个半面上,另一半面的情况也是如此。总之,我们发现某些性状有相引的现象,而另一些性状则很少或根本没有看到相引的证据,这种差别取决于代表遗传因子的染色体物质在直线排列上的距离。这样一种解释能说明我所观察到的所有大量的现象,并且我想,将能同样解释至今谈到的其他情况。这种结果是染色质物质位置和同源染色体联合方法的一种简单的机械结果,而所得的比例用数字来表示不如染色体里面因子的相对位置那么多。
研究范围
看来如果这个假说是正确的话,可以把交换的比例作为任何两个因子之间距离的一项指标。然后通过测定A和B及B和C之间的距离(上述含义),能预测AC。因为,倘若交换的比例的确能代表距离,AC必定或是AB+BC或是AB-BC,而不会是任何一个中间值。然而,根据纯粹数学方面的考虑,A和B之间交换比例同B和C之间交换比例的总和及差数,只是A和C之间交换比例的限制值。通过使用几对因子人们定能在一些例子中使用这样测验。此外,同时包括3个或3个以上性连锁等位因子对的实验,将对这种观点提供另一种测验,它或许是一种更为重要的测验。本文是对这些问题所作研究的初步报告。
我希望借此对摩尔根博士慷慨地给我提供本研究的材料和在工作进展中所给予的鼓励和建议表示感谢。在这一问题的理论方面,同缪勒(H. J.Muller)、E. Altenburg、布里奇斯(C. B.Bridges)等诸位先生们的讨论,也使我得益匪浅。Muller先生的建议在本文的实际准备中特别有用。
6个有关的因子
我将在本文处理6个性连锁因子及其相互关系。我将讨论的这些因子,其次序看来已排列好。
B为黑色因子。同它有关的果蝇隐性因子b具有黄色的身体。摩尔根(1911)首先介绍了这个因子和它的遗传。
C是一个使眼睛有色的因子。白眼果蝇(摩尔根在1910年第一个介绍)现已知它对C和第二个因子都是隐性。
同O有关的果蝇隐性因子(o)为曙红眼。C和O之间的关系已由摩尔根在一篇正在付印并准备在费城自然科学院会议录上发表的文章作过解释。
P带有p的果蝇为朱红眼,替代了正常红眼(Morgan,1911)。
R这一个因子同第二个因子都对翅有影响。正常翅为RM。已知rM为小翅,RM为痕迹翅以及rM为痕迹小翅。这个R因子是一个被摩尔根(1911)以及摩尔根和Cattell(1912)称为L的因子。摩尔根在更早些时候文章(1911)里面的L是第二个因子。
M这个因子在上面已讨论过,它在R之下,由摩尔根叙述过小的和痕迹翅(1911)。
这些因子的相对位置是B、C/O、P、R、M。C和O因其完全连锁而位于同一点上。在弄清楚有两个相关的因子之前,从CO(红眼)和co(白眼)的杂交得到了数千个果蝇。因为有一个突变而不是通过任何交换才发现了这两个因子。但很明显,除非相引的强度不一,否则联系到别的等位因子对,不管用的是CO(红眼)对co(白眼),Co(曙红眼)对co(白眼),还是CO(红眼)对Co(曙红眼)(cO的组合尚不清楚),都必定得到相同的配子比率。
计算联合强度的方法
我用P和M因子来举例说明计算配子比率所用的方法。在这种情况下使用的是长翅、朱红眼的雌性同痕迹翅、红眼的雄性来杂交。分析和结果见表1。
当然,从图表中明显地看出有一些地方是痕迹翅果蝇所特有的。它们出现的数目是如此之少。这一点在这里不会难住我们,因为它的出现经常同痕迹翅杂交有联系,而摩尔根正在对此进行研究。目前感兴趣的要点是连锁。在F2 ,原始组合红眼痕迹翅和朱红眼长翅在雄性中(允许痕迹翅的低生活力),比两个新的或交换组合即红眼长翅和朱红眼痕迹翅要多得多。从这一分析看出,在雌性中发现联合的证明,由于一旦发生联合,在所有产生雌性的精子中存在的M就把m掩盖起来。但通过F2 的雄性可毫不复杂地得出配子的交换比率。鉴于F1 雄性果蝇的产生雄性的精子没有性连锁的基因,在这种情况下有349个雄性果蝇属于无交换一类,而交换类的有109个。根据本文一开始就提出的理论,看来能最令人满意地表示因子相对位置的方法如下:取距离单位为这种长度的染色体的一部分,即平均来说,每产生100个配子会发生一次交换。这就是说,把交换的百分数作为距离的一个指标。就P和M来说,405个配子中发生109次交换,其比率为100次中26.9次;把交换的百分率26.9看成是P和M之间的距离。
表1

因子的直线排列
表2说明那些已经研究清楚的交换比例。有关杂交的详细结果列于本文的结尾部分。B和CO的16287例取自Dexter的资料(1912)。由于C和O完全连锁,我把C、O以及C和O的数目加在一起,使以(C、O)P、B(C、O)等为起点的线上可得出总的结果,并在我的计算中用这些数字来代替各别的C、O或CO的结果。交换比例一栏上的分数表示交换数(分子)对被研究的总配子数(分母)。
正如以后将要说明的,如果因子的距离短,即如果联合有力,更可能得到距离的精确数字。
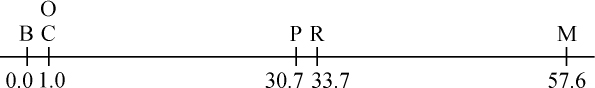
为此,我尽可能用相邻点之间的交换百分率来绘制不同因子间的距离。这样,B(C、O)、(C、O)P、PR和PM就成了图解的基础。图上的数字表示计算出来的同B相距的距离。
表2

当然,所划的距离是否代表因子间相对的空间距离,这一点还不知道。这样,CP的距离实际上或许比BC要短些,但就我们所知,C和P之间比B和C之间发生断裂的可能性要大得多。因此,CP或者是一长段距离,或由于某些原因它也是一段弱的地方。这里我希望提出的一点是我们无法知道染色体是否具有均匀一致的强度,而如果染色体有强的和弱的地方,那就会妨碍我们的图解代表实际的相对距离,但我想它不至于贬低作为一个图解的价值。
我们的理论在多大程度上站得住脚,可见表3所示的测验,它提供所观察到的交换百分率和根据染色体图上的数字计算出来的距离。表3包括列于表2但没有用在绘制图解的全部因子对。
表3
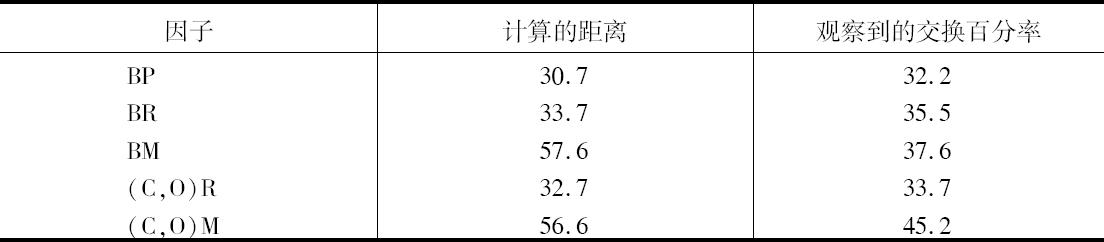
我们立刻就想到,距离长的BM和(C、O)M,其交换百分率比计算所要求的小些。这一点是可以猜想到的并将在后面加以讨论。现在根据以下见解可对此不予考虑,即在同一个染色体里面可能发生两个断裂,或双交换。但是在距离比较短的情况下,其预期符合度同现在的小的数目方面的预期结果很接近。因此,BP较BR少3.2,预期差数为3.0。(C、O)R较BR少1.8而不是1.0。实际上已发现用这个方法有可能预测两个因子之间的结合强度,作杂交之前,对BR和某些在本文中没有处理的有关因子的组合,得到了相当不错的近似值。
双交换
根据交叉型假说,如Dexter(1912)表示,并为摩尔根(1911)明确指出,有时候会发生一种情况,有一段母本染色体其两端都带有父本染色体的成分,或者除此之外,有更多的母本染色体节段。如若发生这种情况,将使结果复杂化。例如,当B和P之间发生一个断裂,和P和M间发生另一个断裂,则除非我们也能追踪P,不然就无法证明B和M之间的关系,而从这类配子孵化出的果蝇将被列入无交换一类,尽管在事实上这种果蝇代表两次交换。为了弄清楚双交换是否确实存在,必须在同一个实验中使用3个或3个以上的性连锁等位的因子对。摩尔根(1911)以及摩尔根和Cattell(1912)对B、CO和R因子报道过这种情况。他们作了这样一些杂交,如长翅、灰体、红眼同小翅、黄体、白眼杂交,长翅、黄体、红眼同小翅、灰体、白眼杂交,等等。其详细情况和分析见原文,从我们现在的目的出发,只对那些观察到的双交换的果蝇感兴趣。表4以图解表示10495例中发生的情况。
双交换的确是发生的,但注意到B和CO间发生断裂,就会阻止CO和R之间发生断裂(或反之)。这样,当B和CO不分开时,CO和R发生断裂的配子比率约为1至2,但是,当B和CO分开时,其比率则为1∶6.5左右。
本人的研究结果得到了3个相似的例子,虽然这是在数量较小的水平上作出的,这3个例子见本文结尾部分的表。其结果列于表5、6和7。
表4
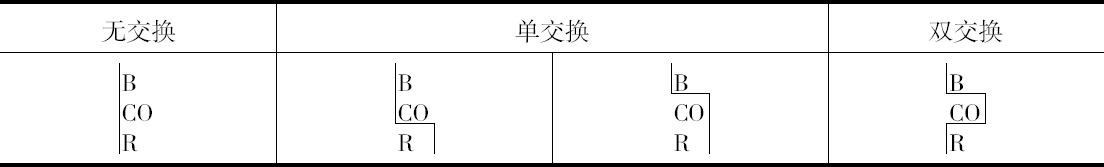
表5
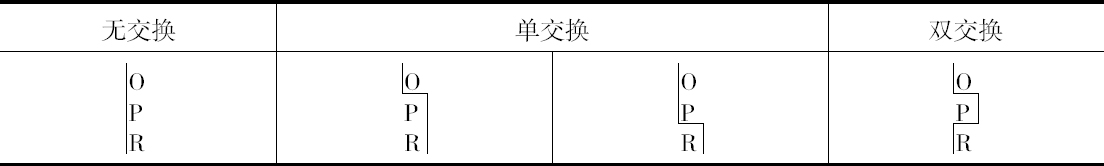
表6
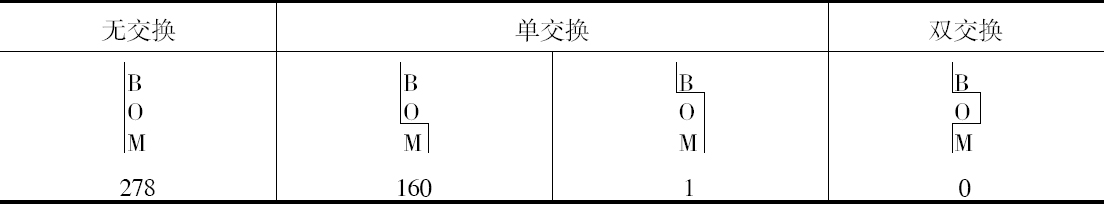
表7
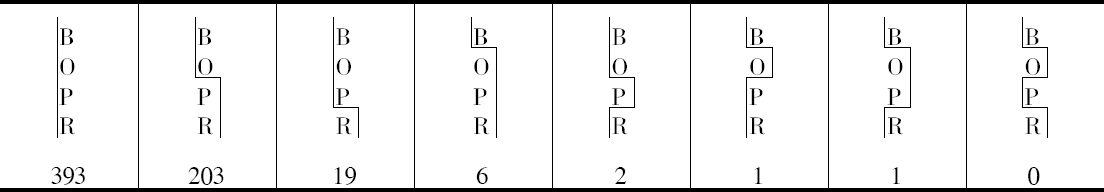
人们将注意到,就目前已有的证明而言,它表明在同一个配子里面,发生一次交换就使另一次交换不大可能发生。以BOPR来说,有3次交换的机会,但它并不发生。当然,如果得到足够数目的果蝇,根据本文提出的观点,没有理由说为什么它一定不发生。对这些数字进行检验将证明像本文提供的这样小的数目是不能指望它发生的。目前,就我所知,还没有3次交换的证据,但发生3次交换将得到证明,这是非常可能的 【31】 。
遗憾的是,上述4种例子中没有一个涉及两个比较长的距离,而只有一个例子的数字足以构成作计算用的比较好的基础,所以似乎还很难确定,在推翻BM和(C、O)M例子中所观察到的交换百分率方面,双交换究竟起了多大的作用。这种效应是否为3次交换所引起的反平衡,这一点至今未得解决。现正进行的工作应当对这两个问题作出答复。
表8




对这些结果可能提出的反对意见
人们将会注意到,在相引强度上看来有些变化。这样,我发现(CO)R是36.7;Morgan和Cattell得到的结果为33.9;OR我得到的是34.0,而CR则为28.5。(CO)R(全部数字)与CR间差数的标准差是1.8%,它是指5.5%的差数可能是显著的(Yule,1911,第264页)。观察到的差数是6.1%,这说明存在着一些复杂现象。同样地,BM为37.6,而OM为54.0,以及BOM对BM来说是36.7,对OM则为36.5。在这些例子中显然有些复杂的情况,但我倾向于认为,下面谈到的干扰因子(生活力)将对此作出解释。然而,现正在进行实验以测验某些外部条件对相引强度的影响。人们将看到,从整体上来看,如果从不同实验得到大量数目并加以平均的话,就得到了一个相当一致的图解。然而,在这一问题能通过进一步的实验予以澄清之前,以暂不作出最后的定论为宜。
在这方面必须考虑到的另一点是生活力不同的影响。就以上作为举例说明的P和M来说,痕迹翅的果蝇,其发育要比长翅果蝇差得多。现假定红眼和朱红眼的生活力是不同的,那么长翅果蝇将不能测出连锁,而出现数目如此少的痕迹翅又不能同长翅数目拉平。由于这个原因,除痕迹翅杂交的情况外,可能没有什么重大的错误,因为这两边的数字倾向于拉平,一方的存活力大大低于另一方的情况则除外,而这只有在痕迹翅的情况是这样。在痕迹翅杂交[BM和(CO)M]情况下碰到的观察和计算之间唯一明显的不一致性,没有什么参考价值。摩尔根现正付印的某些资料和早已计划的进一步的工作,将可能对M这个因子的位置和行为的有关问题作出重要的揭示。
小结
已发现把每100例的交换数目作为任何两个因子之间距离的一项指标,有可能将果蝇的6个性连锁因子作直线排列。此计划总的说来得到了一致的结果。
在预测未经测验的因子之间的联合强度方面,发现双交换中有一个错误来源。此现象的发生得到了证实,并证明从纯数学观点来看,出现这种现象不会像预期的那样多,但至今未研究清楚控制其发生频率的条件。
根据摩尔根采用Janssen的有关联合遗传的交叉型假说对这些实验结果作了解释。它们形成有利于染色体遗传观点的一种新证据,因为它们强有力地表明所研究的因子是直线排列的,至少在数学上是如此。
参考文献
[1]Boveri, T., 1902. Ueber mehrpolige Mitosenals Mittel zur Analyse des Zellkerns. Verh. Phys-Med. Ges Wurzburg., N. F., 35 , 67.
[2]Dexter, J. S., 1912. On coupling of certain sexlinked characters in Drosophila. Biol. Bull.,23 , 183.
[3]Janssens, F. A., 1909. La théorie de la chiasmatypie. La Cellule, 25 , 389.
[4]Lodk, R. H., 1906. Recent progress in the study of variation, heredity, and evolution. London and New York.
[5]McClung, C. E., 1902. The accessory chromosome—sex determinant? Biol. Bull., 3 , 43.
[6]Morgan, T. H., 1910. sex-limited inheritance in Drosophila. Science, n. s., 32 , 1;
1910a. The method of inheritance of two sex-limited characters in the same animal. Proc. Soc. Esp. Biol. Med., 8 , 17;
- The application of the conception of pure lines to sex-limited inheritance and to sexual dimorphism. Amer. Nat., 45, 65;
1911a. The origin of nine wing mutations in Drosophila. Science, n. s., 33 , 496;
1911b. The origin of five mutations in eye color in Drosophila and their modes of inheritance. Science, n. s., 33 , 534;
1911c. Random segregation versus coupling in Mendelian inheritance. Science, n. s.,34 , 384;
1911d. An attempt to analyze the constitution of the chromosomes on the basis of sex-limited inheritance in Drosophila. Jour. Exp. Zoöl., 11 , 365.
[7]Morgan, T. H. and Cattell, E., 1912. Data for the study of sex-linked inheritance in Drosophila. Jour. Exp. Zoöl., 13 , 79.
[8]Stevens, N. M., 1905. Studies in spermatogenesis with special reference to the "accessory chromosome." Carnegie Inst. Washington Publ.,36 . 1908 A study of the germ-cells of certain Diptera, Jour. Exp. Zoöl., 5 , 359.
[9]Sutton, W. S., 1902. On the morphology of the chromosome group in Brachystola magna. Biol. Bull., 4 , 39.
[10]Wilson, E. B., 1905. The behavior of the idiochromosomes in Hemiptera. Jour. Exp. Zoöl., 2 , 371.
- The sexual differences of the chromosomegroups in Hemiptera, with some considerations on the determination and inheritance of sex. Jour. Exp. Zoöl., 3 , 1.
[11]Yule, G. R., 1911. An introduction to the theory of Statistics. London.
性别对染色体和基因的关系 【32】
卡尔文·布里奇斯(C. B.Bridges) 【33】
(1925年)
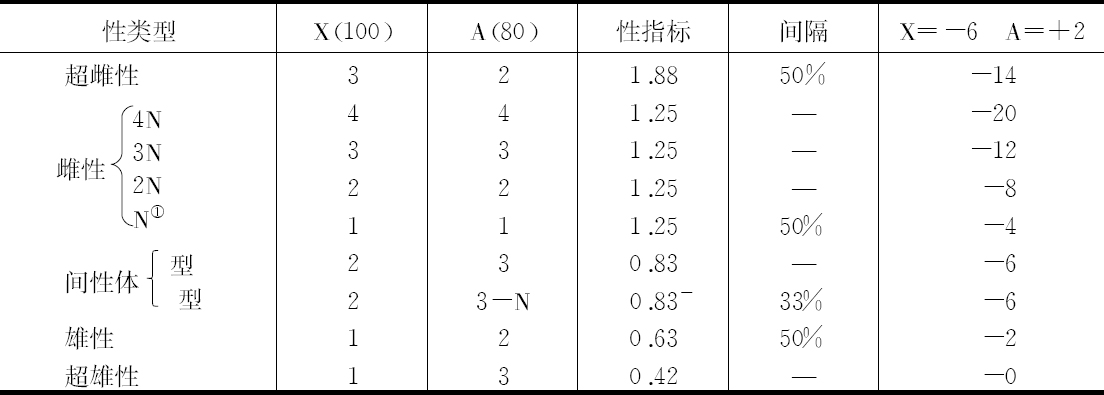
自从在多伦多会议上报告(Bridges,1922)后,3年来在果蝇育种工作中得到了一批不同的性类型,从中积累了重要的新资料(见表1)。这些不同的性类型,其中每一个都是染色体的一种特殊组合的结果。它们主要发生在三倍体的雌性后代中。这就是说,它们有3个X染色体和3套常染色体。具有一个额外的X染色体和同时具有额外的一套常染色体组,并不使这个体在其性的性状方面同正常的雌性类型有什么不同。然而,在配子发生时,3N组是一种不稳定的染色体组。每个卵接受一整套染色体组,另一整套染色体组到极体。额外的一套染色体的成员则在卵和极体之间以各种可能组合的方式进行分配。这样,一种普通的3N雌性卵是有一套额外的染色体组。如我们用A表示一套常染色体组,这个卵可以写成X+A+A或X,2A。当这样一种卵被一个正常的可写成X+A的精子受精,其接合子为2X,-3A。这种接合子发育成一个间性体,即发育成一个个体,它既不是雄的也不是雌的,而是一个中间性的,或是雌、雄部分的混合体,它与Goldschmidt在舞毒蛾(Lymantria dispar)方面进行过广泛研究的那种间性体(Goldschmidt,1920)非常相似。
表1 果蝇性别对染色体的关系
①未发现单倍体类型。
由于在测定正常的性状方面作了非常广泛的研究,特别是研究了一方面丢失小而圆的染色体之一的第四染色体,同时,另一方面取得此染色体所引起的可对此进行比较的性状变异,就能用染色体上携带基因的论点来解释这些间性体。根据这些研究产生了一种观点,即一个个体的每个性状是大量的、但数目未知的基因发生作用达到平衡点的指标,其中有些基因朝着一个方向改变发育,另一些则朝着相反的方向起作用(Bridges,1922)。把基因平衡的概念应用于间性体的性性状的情况如下:在染色体组成方面,间性体与雌性之差别仅在于前者多了一套额外的常染色体组。这证明常染色体同性别决定有关。此外,它们的作用是决定雄性,因为增加一套常染色体使雌性看起来像雄性的特性。这就是说,在常染色体中,有倾向于产生我们称之为雄性性状的基因,当这些基因比倾向于产生另一些即我们称之为雌性性状的常染色体基因总数,数目更多或潜力更大时,它们的作用就更有效。另一方面,X染色体有一净雌性趋向,证明这一点的事实是给雄性果蝇增添一个X染色体,就使个体变成雌性。一组常染色体的净雄性趋向不如一个X染色体的净雌性趋向。实际上看到,常染色体组和X染色体各有两份的个体,即2X,2A。雌性基因在分量上超过了雄性,而结果是一个雌性。假如我们用100代表X染色体里面雌性趋向基因的净效应,然而用小一些的数目字代表一组常染色体的净雄性效应,比如80。在一个2X,2A的个体,雌性效应对雄性效应之比为200∶160,或1.25∶1。根据这个公式,1.25的性指标相当于正常的雌性。在X,2A的个体,雌性效应对雄性效应之比为100∶160;或者说一个正常雄性的性指标是0.63。在2X,3A性间体,其比率为200∶240,性指标是0.83,介乎雌性和雄性指标之间。3N雌性的比率为300∶240,性指标为1.25,同正常雌性完全相同。3N和2N形式性指标的一致性,同它们之间没有看到明显的性差别是相符的。3N个体体积较大,眼结构较粗糙等,这可直接归结于核体积的改变而不是性的性质。
另一种3N 卵是X+X+A;这种卵同一个正常的XA精子受精得到的一种3X,2A的个体,其性指标为1.88,它比正常雌性高50%。这个结构在事实上符合在这些繁殖体和其他地方所碰到的超雌性。这种超雌性发育大大推迟,很少能存活,并可能是完全不孕。
与此相反,一个X+A+A的卵,被不带X染色体的那种精子受精,就得到一个X,3A的接合子,其性指标只有0.42。可以指望这种个体比一个正常的雄性个体更雄性化;在3N雌性后代中曾找到这类个体。最初什么也没有找到,但不久发现,繁殖很晚时遇到一个偶然的例子,它是一种明显不同的雄性类型。这些所谓超雄性同样是不孕的。最近得到的细胞学证明说明了这种性别的构造为X,3A,它同以前得到的遗传学证明是一致的。
据观察,间性体有明显的变异,并且似乎形成一种双峰极。鉴于细胞学研究已证明一些间性体有3个小而圆的第四染色体,而另一些只有两个这样的染色体,据猜测这种更雄性化的模式同完全的三重第四染色体相符,而更雌性化的模式则与缺少一个第四染色体的细胞学类型相符。在这一点上致力于获得细胞学证据。但这种证据说服力不够;通过使用第四染色体的突变体无眼性状,试图对存在第四染色体的数目作遗传学测验,结果也不肯定。目前用3N母本对已知有一个额外的第四染色体的雄性个体作连续杂交,正在把额外的第四染色体人工插入到间性体里。相反,别的间性体品系则用3N母本同已知两个第四染色体中缺少一个的雄性个体连续交配,使第四染色体的数目减少。同样,3X,2A个体的超雌性可通过同具三倍第四染色体的雄性或单倍第四染色体的雄性交配,使之减少或增加。此试验所用的雌性,它们的两个X染色体一直彼此相连(L. V.Morgan,1922),由此通过不离开而得到比例很高的3X超雌性。从尚未完成的试验来看,现在的说法尚有矛盾之处,但倾向性的结论同早些时候可能根据当时提供的少数证据所作的报道(Bridges,1922)相反。如第四染色体的数目是3,间性体更像雌性;当这个数目为2时,它们就更像雄性。第四染色体有一种与X染色体相似、却又与其他常染色体有别的净雌性趋势。通过变换第四染色体数,有可能对每一种性差别的主要类型的次要性类型略知皮毛。
四倍体或4N个体的发现,扩大了性的种类。这些性类型为雌性,它们的性性状同正常的雌性十分相像。四倍体是在一个三倍体原种里得到的;并只有通过所得后代差别极大才能探查出来。从3N原种选出一个假定为3N的雌性体,再用一个正常雄性体同它外交。所有后代都是三倍体雌性体(约30),或三倍体间性体(约20)。没有2N后代或超性。据知,如果母本是4N,而不是3N,就可以得到这个结果。因为在那种情况下,所有减数的卵将是2N;而它们与X精子受精将产生3N雌性,同Y精子受精将产生2X,3A的间性体。
在发现这个4N个体前,从一些事实推测它是会发生的。这样,在发现三倍数性以后的3年内,已发现新发生的三倍数性不少于25例。这个频率很高,与细胞学观察是平行的,它对三倍体的起源作出了解释。在3个分开的正常2N雌性体的制片中,发现一个卵巢的一部分由明显增大的细胞组成;有两个个体有两个正在分裂的巨型细胞,计算其染色体数为4N。显然,在一些卵原细胞中有一种染色体分裂,这种分裂以后不发生核和细胞质的分裂。由此产生的组织是四倍体,而任何一个减数的配子将是2N。这种2N配子被一正常精子受精,将对25次出现三倍数性的每一例作出说明。
此外,在检查间性体的切片时,发现两个个体出现比细胞还要大的相似的囊。其中一个囊正在发生分裂;染色体很清楚是6N。一个3N雌性里面的一个6N囊,经过减数将产生3N的囊,它同X精子受精将产生预期的4N雌性类型。
出现这第一例4N雌性后不久,发现了第二个相似的例子。L. V.Morgan还发现了第三例,并能用遗传学测验证明,存在着4个分开的X染色体(正在付印)。
事实是4N个体都是雌性,且对性别不作修饰,这对我们形成基因通过这种途径相互作用以发挥其效应这一思想有着重要意义。此处采纳的观点是:一般说来,有效性同基因的数目成比例,而要点为那些倾向于产生正反两种效应的基因组之间的比率。根据这一观点,我们发现对这种实际情况很容易解释,即2N、3N和4N个体这样一些不同的形式,在它们的性性状方面是精确相似的,因为所有这些形式,两种相互抗衡影响的效应都得到了加倍、三倍或四倍,而其比率则保持不变。
但是公式系统同Goldschmidt在他的卓越的舞毒蛾族间杂交工作中研究所产生的间性体时采用的比率类型有所不同。他对某一个族的雄性趋向指定一肯定的值,这个值同雄性决定基因成比例,他指定雌性趋势另一个值,也同雌性决定基因的强度成比例。然后他假设,当一个个体里面雄性值大于雌性值时,该个体是雄性,反之,当雌性值以相同的单位数大于雄性值时,该个体就是雌性。雄性趋向基因(M)的位点在“Z染色体”上,该染色体雄性有两个,雌性有一个。雌性趋向是严格的母体遗传,因此,F(雌性) 【34】 基因的位点位于从母亲传给女儿的W染色体上。假定F基因对发育卵的细胞质起作用,那么尽管雄性无W染色体,可认为它有卵对细胞质施加的一定的雌性趋向,并保持在发育的始终。对一个弱族来说,F这个值被指定为80,M是60。在WZ个体,细胞质F 值80超过M值60,上位最小值20个单位,因而这个个体是雌性。同样地,在ZZ个体,F 值为80,但净M值为60加倍成120,其雄性方向要大40个单位。一个强的族则F和M都高些,例如,分别为100和80,但F和M值之间的数字关系仍控制着个体的性别。当一个弱的雌性和一个强的雄性作杂交,ZW个体从母亲那里接收一个80值的F ,从父亲那里接受80值的M。这个值在一方为雌性所需的超量和另一方为雄性所需的超量之间中途就达到了平衡,其结果就成为一间性体。这个公式远不能令人满意,如不对大批杂交进行比较并通过整个试验范围来得出每一个族的F和M值,这个尝试很难成功。据我看来,对比率作重新设计以取代代数基础,将得到一批连贯的指标,而又不会同Goldschmidt发展起来的非常有价值的生理概念相违背。
在果蝇性类型的表中,单倍体个体的指标为1.25、2N、3N和4N雌性的指标相同。不幸的是,没有发现过单倍体个体。但根据上述观点看来很清楚,对一个单倍体果蝇(Drosophila melanogaster)的预测它将是雌性的。因此,必须认为,果蝇的性决定机理同蜜蜂和相似形式的情况大不相同,在它们那里单倍体个体是一个雄性个体。对我来说,蜜蜂的性决定是突出的没有解决的谜题,虽然在发展出基因平衡的概念之前它似乎是一个最清楚和最简单的例子。如雄性确是一单倍体个体,那么人们将设想二倍体个体必将同样是雄的,因为性决定基因中间的比率在这两种情况下并无差别。
Schrader和Sturtevant曾设法用Goldschmidt的代数公式来协调果蝇和蜜蜂的情况。他们对每个A定一个正值即+2,给每一个X定一个负值-6。然后假定其有效关系为X和A值的代数和(见表1右方一栏)。根据这个观点,单倍体或许是雄的。但这个系统有一个难题,这就是连续指标间的区间与性等级间看到的差异不能很好地吻合。这样,实际上在3N和2N个体间观察到的最小的区间,可用4个单位的差数来表示,而雄性和雌性之间非常大的区间却只有6个单位来表示。那时候还不知道4N型;如把它加进去,符合度在代数系统方面就很差,而在比率系统方面却很好。我重申,只要现在对蜜蜂机理的报道没有受到挑战,我不把蜜蜂的例子解释成同果蝇相同的基础。目前,单倍体和二倍体性别间的差别必须涉及同更大的体积、眼的更粗糙的结构和其他能从2N个体中辨别3N个体的轻微变化有关的同一种决定类型。
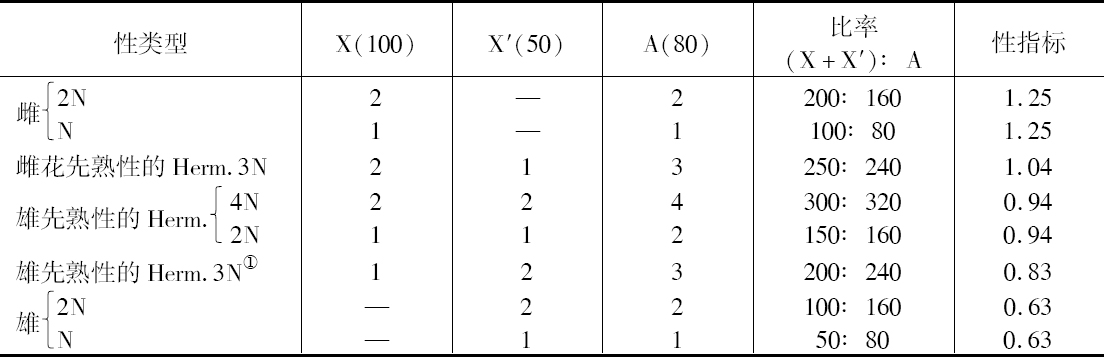
但除了像蜜蜂这类例子,看来很可能相互作用的比率是普遍的模式。为证明这一点,可引用一大批雌雄同株植物,其性关系二倍体、三倍体和四倍体都一样。当然,三倍体形式高度不孕,这因为3N组在减数分裂时不稳定,并由此产生无生活力的配子或接合子。最有力地支持这种基因平衡的比率观点之一,是通过Marchals、Schweitzer和von Wettstein的杰出工作在藓中看到的。例如,他们发现有一种藓,它具有分开的性别:一个结合两组雌性染色体的2N配子体是一个纯的雌性,相似于单倍体雌性植物(表2),同样地,结合两组雄性染色体的2N配子体是一个纯的雄性,相似于单倍体雄性植物。但一个结合一套雌性染色体一套雄性染色体的2N配子体就不再是一个单性植物,而是一种雄性先熟的两性体。此外,结合两套雄性染色体组和两套雌性染色体组的4N配子体(FFMM)是一个两性体,它们相似于FM两性体。但由两套雌性一套雄性染色体形成的三倍体是一个两性体,它是雌花先熟性的,而非雄花先熟性的。另一方面,在研究单倍体组是一两性体的雌雄同株藓时,那么所有的单倍体、二倍体、三倍体和四倍体植株都毫无区别地是两性体,这因为从它们具有雌性对雄性决定子的相同比率来看,它们必然如此。
从表2可见,一套性指标,像果蝇那样对雌雄异株的藓是适合的。这里假设有一对染色体X和X′,它们的差别说明是雌性和雄性类型之间的差别。据设想在这两种性别中,或可用A表示其他染色体的净效应都是雄性决定。那么,由于X, A型为一雌性,A值必定小于X,例如,X=100,A=80。同样地,由于X′A为一雄性,X′值必须小于A值,例如,X=50。还有,由于FM植株是一个两性体,它不像正常的雌性而更像正常的雄性,X+X′<2A。并由于FFM植株为一两性体,它更像正常的雌性,X+X+X′>3A。这样如表2中所示,3个值X, X′和A,其指定值为100,50和80,就可能有5种限定的等式,尽管或许有别的轻微差异的值,它能得出一套指标,其区间对观察到的差数比上述情况甚至更为密切符合。
表2 一雌雄异株的藓的性类型
①已知此类型未报道。
就我能搜集到的材料来说,在单倍体、二倍体、三倍体和四倍体曼陀萝中,比例规律看来是同样符合的。但以基因平衡观点来看,曼陀萝的12种染色体,其中每一个可以像果蝇第四个染色体的不平衡一样,具有性控基因可予区别的内部不平衡。曼陀萝通过增加一个特定的额外染色体有一整套同2N有区别的形式。如果这12种染色体中的任何一个,其雄性趋向基因比雌性趋向基因更为有效,或相反,那么人们完全可以指望发现Blakeslee的一些“Apostles”和“Acolytes”具有非典型的性别关系。
参考文献
[1]Blakeslee, A. F., 1922. Variations in Datura, due to changes in chromosome number. Am. Nat., 56 , 16~31.
[2]Bridges, C. B., 1922. The origin of variations in sexual and sex-limited characters. Am. Nat., 56 , 51~63.
[3]Goldschmidt, R., 1920. Untersuchungen uber Intersexualitat. Zeit. f. ind. Abst. u. Veror., 23 , 1~197.
[4]Morgan, L. V., 1922. Non-criss-cross inheritance in Drosophila melanogaster. Biol. Bull., 42 , 267~274.
[5]Schweitzer, J., 1923. Polyploidje und Geschlechterverteilung bei Splachnum sphericum Schwartz. Flora. 116 , 1~72.
[6]Wettsteinm F. V., 1924. Morphologie und Physiologie des Formwechsels der Moose auf genetischer Grundlage. Zeit. f. ind. Abst. u. Verer., 33 , 1~236.
基因的人工蜕变 【35】
赫曼·缪勒(H. J.Muller) 【36】
(1927年)
大多数现代遗传学家将同意基因突变为有机体进化以及生命物质极其错综复杂的主要基础。然而,对遗传学家来说,不幸的是在正常条件下很少发生基因突变,以及试图用一种肯定和可以探查的方法来有力地改变这一缓慢的“自然”突变率未获成功,致使基因突变以及通过突变对基因本身的研究受到了非常严重的障碍。当然,从更直接的实用目的出发,这些局限性也影响到改造有机体原有的本性,因而实际育种工作者不得不把育种内容停留在只对现成的材料,加上自然界由于偶然和稀有的机会,产生无法预测的突变所恩赐的材料进行重组。正由于这种情况,它促使一部分生物学家,非常希望获得一些控制基因内遗传性发生改变的方法。
人们曾一再报道X射线或镭射线能诱发生殖变化,大概是诱发突变,但正如与此相类似的公开主张,使用其他诱变剂(酒精、铅、抗体等)的情况一样,在进行这些工作时,从现代遗传学观点来分析,这些资料的含义至多引起争论纷纷而已。此外,还争议哪些是反复得到负的或相反结果最明显的例子。尽管如此,从理论上来说,作者感到波长短的射线特别容易产生突变变化,鉴于这一点和其他理由,去年用果蝇(Drosophila melanogaster)开展了一系列同这一问题有关的试验,试图提供精确的资料。众所周知,果蝇这一物种适合于作遗传学研究,作者8年来在果蝇突变率(包括温度方面)的深入工作中采用了专门方法,这终于使我能找出使用X射线而得到的一些肯定的效应。这里所指的效应是真正的突变,同已知的以不离开的、不遗传的交换修饰等,表示X射线对染色质分布的效应决不混淆。本短文为这些工作的摘要,只能列举一些概括的事实及其结论,提出一些问题,而不能详尽涉及所用的遗传学方法或所得到的个别结果。
已十分肯定地发现,用较高剂量的X射线处理精子,能诱发受处理的生殖细胞发生高比例的真正的“基因突变”。这种方法在短时间内得到了几百个突变体,并经过四代或四代以上,接着发现一百个以上的突变基因。它们(总之,几乎是全体)在遗传上是稳定的,并且大多数表现为,一般在有机体中发现的孟德尔染色体突变基因的典型方式。同其他染色体相比,杂交的性质更适于探查X染色体的突变,所以大多数涉及的突变基因是性连锁的;但有大量证明,突变同样地通过染色质发生。当精子接受最高剂量处理时,由这些精子得到并繁殖的后代中约有1/7的后代,在它们的X染色体上具有一个个能探查出来的突变。由于X染色体形成的1/4的单倍体染色质,因而,如果我们假定,所有的染色体的突变率相等(按它们的长度为单位),那么,几乎精子细胞“每隔一个”能产生一个可育的个体,这个个体在这一些染色体或另一些染色体上,具有“可以分别探查出来”的突变。数千个未经处理的亲本果蝇作为对照,以相同于受处理果蝇的方法进行繁殖。在两组条件下突变率的比较表明,高剂量处理的突变率要比未受处理的生殖细胞,高出约15000%。
关于所产生的突变类型,如预期的那样,作者发现不论在理论上,或是根据Altenburg和作者过去的突变研究,致死因子(致死效应为隐性,尽管有些为显性,其效应可见)在数量上,大大超过产生可见的形态上畸形的非致死因子。也有一些“半致死因子”(规定突变体的存活力通常约为正常个体0.5%~10%之间的为半致死)。但幸运的是,对于把致死因子作为突变率的一项指标来说,半致死因子不像致死因子那样多。那种使存活率降低得更少,且不易与致死因子混淆的、渗漏的“不可见”突变,看来比半致死因子要更多些,但没有对它们进行研究。此外,这些实验也可能首先得到证明,在X染色体和别的染色体中,都发生了显性致死因子的遗传变异。由于接合子接受了这些显性致死因子,它们就不能再发育到成熟,这类致死因子不能一个个地探查出来,但它们的数目是如此之多,以致通过卵的计数和对性比率的影响,可以证明它们是大量存在的。已发现其数目同隐性致死因子为同一个等级。由这些显性致死因子造成受处理的雄性果蝇的“部分不孕性”,至少已达到了可观的程度。发现了另一种数量大,而过去并不知道的突变,当它们为杂合时,导致不孕性,但在外貌上看不出有什么变化,它们发生的数目也很相似于隐性致死因子,并且当发生频率高时,以后可作为一般突变的最方便的指标之一。当然,它对受处理个体后代所造成的不孕性,同受处理个体本身由于显性致死因子所造成的“部分不孕性”,是一种不同的现象。
谈到“一个个能探查出突变”的比例,X染色体有1/7左右,因此,差不多所有染色质的一半仅仅是指隐性致死、半致死和“可见的”突变体。如果考虑到显性致死、显性和隐性不孕基因、或稍微降低存活力或可孕性(或别的影响)的“不可见”的基因,突变体的百分率远远高得多,因此,这就证明,在事实上,受处理精子的绝大多数具有这几种或另几种突变。看来用X射线处理后,基因的突变率占基因的总数的比例方面是相当的高,以致甚至在单个座位(locus)上进行研究,以探查等位性等问题也是切实可行的。
回过头来考虑产生可见效应的诱发突变。需要指出,本实验条件可以探查出许多接近于正常类型或与正常类型相重叠的突变体,而这些突变体通常在观察时会被忽略掉。另有明确的证据,这类突变变化与更明显的突变相比,其频率相当高。这种意见已多次在果蝇的文献中提到过,这种生物在“自然”突变的情况下也是如此,但只是作为“一般印象”来发现。然而,Baur证实,在金鱼草中的确是这种情况。总的说来,已发现射线产生的可见的突变,就其总的特性而言,从Bridges和其他人早先用非射线物质在果蝇方面的探查,可见突变所作的大量观察是相似的。的确,有不少诱发的可见突变是在过去从未看到的座位上发生的,而其中有些突变的形态效应,同过去看到过的并不完全相似(如“斑翅”、“无栉性”等),但另一方面,也有许多为过去已知突变的重现。事实上,果蝇X染色体大多数已知突变,如“白眼”、“小翅”、“带叉的毛”等,都得到过,其中有些多次重新得到。在发现的可见突变中,绝大多数为隐性突变,同其他工作一样,还有“一点点”显性突变。然而首先毫无疑问,由X射线产生的变异,至少其中有许多同那些不加这些处理所得到的“基因突变”完全是同一种,只是后者得到的机会要稀罕得多,而我们相信它是进化过程中的一砖一瓦。
除基因突变外,发现X射线处理也能造成基因在直线上的次序重新排列,且发生比例很高。这一点在一般情况下,是通过经常对交换率发生遗传干扰而得到证实(单X染色体就探查出至少为3%,许多伴随着致死效应,有些则不伴随致死效应),也可以通过各种例子来专门证实这一点,即用其他方法包括一个染色体各部分的倒位、“缺失”、断裂、易位等等来证实。这些例子使人们有可能揭示至今还难以接近的一些遗传学问题。
X射线对基因的诱变作用,不是仅限于对精子细胞,当它处理未受精的雌性细胞时,所产生突变的情况,像处理雄性细胞一样容易。对卵母细胞和早期卵原细胞都有作用。必须特别指出,X射线(在所用的剂量范围内)使哺乳动物一个时期高度不孕,这种不孕在处理后不久就开始,以后又部分地得到恢复。可以肯定地说,孕性的恢复并不意味着新生产的卵不受影响,本实验发现它们像可以存活下来的成熟卵一样,含有高比例的突变基因(通常主要为致死的)。现代X射线治疗常用的照射处理实践,肯定不会造成永久性的不孕,主要是站在一种纯粹理论性的概念上来防护的,这种理论概念认为孕性恢复后产生的卵必定代表“未受损伤”的组织。由于这个假设在这里证明是错误的,所以医学实践有义务作出相应的修改,至少哺乳动物大量遗传学实验可能证明其结果是完全相反之前,应当这样做。同以上果蝇方面的实验相比,用哺乳动物开展这样的工作需要极其仔细地进行。
从生物学理论的观点来看,本实验的主要意义在于,它同染色体和基因的组成、行为问题有关系。通过专门的遗传学方法,有可能得到经处理后第一个和以后的合子世代细胞中,那些发生改变的基因分布方式的资料。已发现,突变并非总是在处理的时候,使该染色体座位上存在的全部基因物质,都发生永久性的改变,但它们或是以这种方式影响到一部分物质,或是作为一种后效应,紧接着只对受处理的基因产生了两个或两个以上的子代基因中的一个发生作用。目前正在设计一系列大量试验,以便在这两种可能性之间作出最终的判断,这一点是必要的,但手头已掌握的证据则倾向于前一种可能性。这是指精子的基因(或从整体来说,是染色体)是一种稍微复杂的物质。另一方面,突变组织分布的方式同一般采用的“基因成分”的理论有矛盾,此理论首先由Anderson根据玉米的杂色种皮提出,以后由Eyster,最近又由Demerec在果蝇(Drosophila Virilis)方面进一步强调指出的。
染色体早期加倍(或以后增殖)的状态(准备下一步有丝分裂),足以说明上述X射线对特定座位的断裂效应。但是这种关于每一个基因分裂成为若干个最初相同的“小单位”,这些小单位在有丝分裂时是否分开还不太肯定的理论,同本工作所得到的结果有出入。根据这种理论,应当在本工作经常发现在杂色玉米和果蝇(D. Virilis)变种方面看到的情况,即突变组织通过经常性的“回复突变”而成为正常的组织;还有,在最初没有显示出突变的受处理的组织,在处理后数代,通过“挑选出”对立的小单位而可以经常产生正常组织。这些效应都未发现。前已提及,发现突变体经数代后变得稳定,至少在绝大多数情况下是这样。数百个受处理生殖细胞没有变化的后代,也让它们通过几个世代,证明在第一代以后的世代中没有产生突变。然而,这样做需要数目更多的材料,而且还要进一步安排各种实验以揭示基因的结构问题,此问题或许能得到肯定的答复。
上述诸点有些已经得到肯定,特别是X射线与产生显性致死有关的断裂效应,看来这对X射线对诸如肿瘤、胚组织和表皮组织中细胞不断进行分裂的专一性破坏作用提供了线索(虽然其他因素的作用,如异常的有丝分裂会得到同样结果而不能予以排除);此外,在偶尔产生的肿瘤中,X射线的逆效应也可以同它们在产生突变中的作用联系起来。然而,现在要详细考虑过去被认为是“生理的”各种X射线效应尚为时过早,目前或可接受的一种可能的解释为X射线的基因诱变的特性;这里,或许更恰当的是把我们自己局限于那些能更严格地被证明为遗传的物质。
从研究X射线的不同剂量以及在生命周期的不同时刻和不同条件下,使用X射线的相对效应,会得到有关基因本性更多的事实。此处报道的实验使用了几种不同的剂量,但结果尚不能十分肯定,可能是在使用剂量的范围内,隐性致死因子并不是直接随吸收的X射线的能量而变化,但更接近于随X射线的能量的平方根而变化。然而,正如Irving Langmuir博士向我指出的,这种缺乏确实的比例关系必须予以肯定,我们必须得出结论,这些突变并不是由在一些特定部位所吸收的X射线能量的单独的量所直接造成的。如果蜕变效应相对来说是间接的,那么更有可能受到其他理化因素的影响,但我们的问题就变得更加复杂化了。然而,把X射线产生的致死突变的总数,作为单个座位上发生基因突变的一个指标,却有些危险,因为有些致死因子包括交换率的改变,可能同染色体区的重排有关,而这种改变与“点突变”相比,由X射线能量的单独的量来决定的可能性更要小得多。因此,必须对不同剂量的效应重新进行检验,在检验时各种突变类型彼此间要清楚地区别开。如果这个问题得到了解决,对于范围很广的剂量和发育时期来说,我们仍然有责任去确定,自然界中存在的微量γ射线,究竟是否在野生和家养的生物中在没有人工施加X射线处理的情况下造成正常的突变。
在研究变换其他条件对X射线产生的突变率的效应方面,作为一个起点,把雄性精子和雌性生殖器照射后的突变率,同雄性生殖系统不同部分生殖细胞照射时的突变率作了比较,没有看到明显的差别。此外,在处理后受精前使精子变老的做法,并没有使可探查的突变频率发生什么明显的改变。因此,突变精子的死亡率并不比未受影响的精子高多少;此外,不能把突变看成是任何一种半致死生理性变化的次生效应,可以认为这种半致死生理性变化,有些精子(“敏感性更高”)比另一些精子发生得更强烈些。
尽管刚刚提到的是一些“负结果”,然而,已经肯定X射线的影响是不同的,而这种差别本身尚不足以说明突变率的全部变化,现在接踵而来的X射线工作是确定决定于温度的突变率(该工作尚未发表)。Altenburg和作者在1918年的工作或许是第一次确定这种关系,但直到1926年完成几项试验后,才得以最后确立温度与突变率的关系。这些试验第一次肯定地证明,基因突变无论到什么程度都是可以控制的,虽然总的说来,热效应同化学反应所发现的情况相似,但联系到几乎不可觉察的“自然”突变率,它的突变率也是太小了。因为人们想通过它为突变研究提供有用的工具。然而,这些结果足以表明,除X射线以外的各种因素的确可以影响到基因的组成,而测定它们的效应,至少在与X射线联合使用时,是行之有效的。这样,我们可希望从各种新的角度来缩短解决基因的组成和行为问题,并在这些研究中发现新的线索,因此,如果不是说基因物理学和基因化学,那么至少在谈到“基因生理学”这一课题是完全合法的。
总之,从事于经典遗传学范围的人们,可以把注意力放在通过使用X射线为他们提供的机会,在他们所挑选的有机体中,创造出一系列可用于研究遗传和“表现型遗传”现象的人工亚种。总的看来似乎有以下可能性:如果这种效应对大多数有机体是共同的,就应当可以“下令”在他们所选择的物种中,产生足够数量的突变体,以提供相应的遗传图,并通过使用这些绘制成图的基因,来分析同时得到的染色体的畸变现象。同样地,对实际育种工作者来说,则希望最终证明这种方法是有用的。要在这里讨论有关人类方面的可能性,时机尚不成熟。
人类血液的个别差异 【37】
卡尔·兰德施泰纳(K. Lansteiner) 【38】
(1930年)
鉴于研究大分子物质的困难,要确定作为有生命机体的主要成分蛋白质的化学特性和组分,我们还要走很长的道路。因此,不是用常规的化学方法,而是用血清试剂所得出蛋白质化学的一个重要的普遍结果,即知悉各动物和植物种的蛋白质是不同的,以及每个种有其自己的特性。事实上不同器官含有特殊蛋白质使这种差异性还在进一步扩大,因此,看来在有生命的机体中,对每一个特定的类型或功能都需要专门的建筑材料,而进行多种多样操作的人造机器却能够从数目有限的物质制造出来。
发现一个物种所特有的生化特异性所提出的问题,即我们将要谈到的研究课题,是确定这种差别是否超越种的范围,以及一个种内的个体是否有相似的但较小的差异。由于在这方面从未作过观察,我选择了可提供的最简单的实验安排和前景远大的材料。为此,本实验是把不同人的血清和红细胞作彼此间的反应。
只得到一定的预期结果。许多样品无明显的变化,换句话说,结果完全像血细胞同它们自己的血清混合一样,但经常发生一种称之为凝集的现象,在凝集中血清使外来个体的细胞聚合成丛状物。
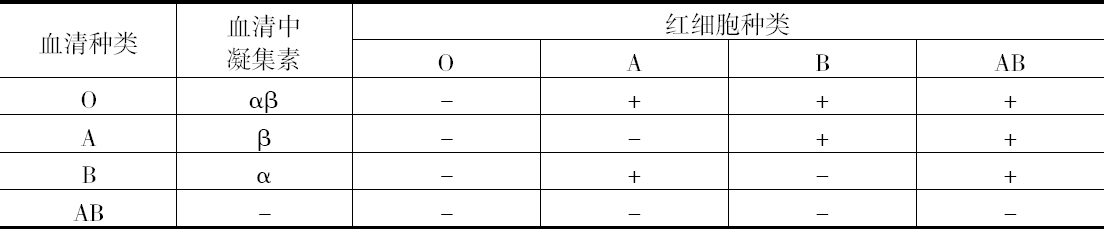
惊人的事情是凝集作用,它一旦发生,就如同不同动物种的血清和细胞间相互作用所发生的那种早已熟知的反应一样明显,而在另一些情况下不同人的血液则看不到差别。因而,首先必须考虑个体间所发现的这种生理差异,是否在实际上就是我们正在寻找的差异,以及尽管这是在健康人血液中看到的现象,但这种现象会不会由于不得病而产生。但不久就弄清楚,此反应遵循一种对全体人类血液都有效的模式,而所发现的这种特性,犹如一个动物种所特有的血清情景一样,恰恰是个体的特性。实际情况是基本上有4种不同类型的人血,即所谓血型。从以下事实得出血型的数目:红细胞显然含有两种结构不同的物质(同种凝集原),在一个人的红细胞中或许两种物质都没有,或者有一种,或者两种都有。单用这一点仍不能对这种反应作出解释;血清的活性物质同种凝集素,也必须有一种特殊的分布。它的确是这种情况,由于任何一种血清具有同细胞中不存在的凝集原起反应的那种凝集素,这是一个值得注意的现象,其原因还不清楚。这种现象使血型间产生某种关系,它使得血型测定很容易,可以下页表说明。根据细胞中含有的凝集原命名各血清(表中符号“+”表示凝集)。
现在提出一个问题:正常血清的同种凝集作用是限定于人血呢?还是在动物中也发生这种作用?事实上这种反应只在少数物种发现,但有区别,且从来没有像人那样有规则。只有最高级的人、猿有血型特征,虽然它们的血球蛋白质同人无甚区别,这种迄今我们尚未确立的血型特征同人的血型特征完全相符。
可以设想,对许多动物种进行比较检验将有助于说明血型这一尚未为人们充分了解的现象是如何形成的。检验动物血已经得到了引人注目的结果。对同种凝集作了第一次观察后不久,Ehilich和Moigenioth介绍了采用血液溶剂抗体(异细胞溶素)的实验,他们证明当山羊被注入同一个种的其他个体的血液时,它的血液就不同了。但是,在这种情况下,发现它不是典型的血型,而是许多明显的随机的差异,其结果除了可能是反应强度外,它大体上同人们预期的情况相符。相似的研究,特别是由Todd在牛和小鸡方面进行的研究(Landsteiner和Miller;Todd)表明它几乎完全是个别的特异性。
在人和动物方面,观察之间存在的明显矛盾最近已得到了解决。在这方面早有几位先行者,以及我同Levine一起进行的工作,通过把人血注射到兔中产生特异的免疫血清,得到了明显的结果;这些结果发现在所有4种血型中存在3种新的可凝集的因子。这样,当考虑把A和AB型各自分成两个亚型时(V. Dungern和Hirszfeld;Guthrie等)(此项工作最近由Thomsen在本实验室作了充分的研究),就发现至少有36种不同的人血。此外,还看到那种并不遵循血型规律,其特异性又不同的微弱的同种反应,这种反应比早先认为的不规则反应更常见,它的确很容易同典型的反应相区别,并绝不影响4种血型的有效性。这些发现肯定了人类中存在许许多多个别的血液差异,以及肯定还有其他尚未探查出来的差异。是否每一个个体的血液确实是它本身的一种特性,或者经常要到什么程度才算完全相似,这一点我们还不能肯定。
在那时候,至少这些因素在考虑血型治疗应用方面不占重要位置,这一点将在后面讨论,但它们很可能同外科的一个重要领域,即组织移植有密切关系。
人们早已熟知,如皮肤的移植,如果被移植的材料来自同一个体,则成功率高得多,由Jensen首先介绍的把可移植的肿瘤转移到一个动物种的不同品系时,其结果相似。动物方面的研究肯定了外科医师的经验,其中由L. Loeb进行的一系列重要的实验特别引人注意。Loeb的实验包括从一个动物自己的身体、从同一个原种的有亲缘关系和无亲缘关系的动物以及从不同品种和物种的成员,分别取下不同组织进行移植。总的说来移植成功同亲缘程度有关,总之,借助于这些观察,可得出结论:不同个体的组织必须具有特异的生化特性。
两个彼此独立的方法所得结果却惊人的一致,不免使人立刻想到,这种一方面用血清反应能探查出的个体差异,另一方面移植组织的个体特异行为的差别,实质上是同一回事。提出这种设想的理由是器官细胞如同血液一样,能证明其血型特点。然而,根据这一设想,即在组织移植时考虑到血型,结果却并不明显。但这是可以理解的,因为血型只说明一些确实存在的血清差异,而很明显,即使轻微的偏差也能影响到组织的永久的痊愈。这将消除由这些实验所产生的怀疑,而最可能的假设是个体间血清学差异和移植特异性,它们基本上是有关系的,并取决于种类相似的化学差异。总之,把血清反应使用于移植治疗的重要工作的可能性,肯定不能予以排除,但现在的知识还只能停留在抱有希望而已。
对于现在我将检验的个体特异物质的化学性质的问题,回答完全是否定的。但尽管如此,它并非毫无意义。我在演讲中一开始提到的沉淀素反应揭示了蛋白质之间的物种差别,由此产生一种观点:所有血清反应的底物是蛋白质或同蛋白质密切有关的物质。血抗原的研究首先使这种观点发生了动摇。特异物质在有机溶剂中的溶解度,特别是对异源绵羊的血抗原的研究(血抗原由瑞典病理学家Forssman在绵羊血液和不同动物器官中发现),发现一种特别结合而又不直接起抗原作用的物质,能用酒精提取并把它分离出来,这使我产生一种想法:许多细胞抗原的组分并非蛋白质一类物质,而只是同蛋白质结合在一起,从而成了抗原,可恰当地把它称之为复杂抗原。这一理论得到如下事实的强有力的支持,这就是当我把这种特殊物质同含有蛋白质的溶液混合时,能保持它的抗原活动。
一项关于细菌中存在某些特异物质的研究(Zinsser)得到了相似的结果。而就细菌来说,这种特异结合的物质(半抗原)的化学性质经测定的确是胶体多糖(Avery和Heidelberger),在动物细胞抗原方面尚未得到肯定的结果。尽管如此,可以这样说,动物中的生化特性在于存在着两类不同的物质——特异物质(Landsteiner和Van der Scheer;Bordet和Renaux),这种物质在它们发生的性质上有着基本的差异。
关于在本演讲中所要谈的课题,其实际情况是聚集的特异物质也能用酒精从血液细胞中提取出来,并且在这种情况下通常使抗体只在一带有抗原蛋白质的混合液中形成。从而得出结论,在一个物种里面的半抗原是有变化的,且允许对蛋白质之间相似的血清学差异产生怀疑,但不能令人信服地得到证明。另一个特点是实际上在动物学系统相距很远的动物种里面,经常碰到同它们的反应有关的半抗原。因此,同种凝集原A在血清学上同在绵羊血液中含有的Forssman抗原有关系,所以免疫血清对绵羊血和A型与AB型的人血都有反应,但对O型或B型人血不起反应(Schiff和Adelsberger)。更值得注意的是细菌里面存在相似的结构。这是从以下事实得出的:在许多抗细菌的血浆,如对Paratyphus bacilli的免疫血清中存在绵羊血的细胞溶素和A型血的凝集素,一种使人血凝集的痢疾血清(最近由Eisler介绍)含有一种抗体,这种抗体对于A型的两个亚型中对同种凝集素不太敏感的那一个影响较大。
根据人造复杂抗原方面的研究结果,发生属于个体差异的免疫同种抗体可能由以下事实造成:由于同其他物质结合在一起,该物种所特有的蛋白质就不能诱导抗体的形成。如反过来,把同该动物相同的或关系密切的半抗原,与外来蛋白质一起注入该动物,看来通常不产生抗体。Witebsky的实验提供了一个例子,它证明注入A型血后,只在兔子的器官里没有同A凝集原相像的物质才形成组型特异免疫血清。然而,Sachs和Klopstock所进行的实验,用兔子器官的混合酒精提取液的外来血清注射后,兔子就发生Wassermann反应,该实验说明这个规律不能普遍使用。
然而,在这种抗体只同器官提取液起反应的情况下,O. Fischer用注射兔血混合提取液的外来血清进行试验,成功地使兔产生同源抗体,这种抗体对完整的血液细胞起作用,但在预冷后只有溶血作用,犹如我同Donath一起发现,溶血作用是血液在阵发冷血红蛋白尿中溶解的原因。这一结果和一方面从O型和B型红细胞提取液,同另一方面从完整细胞所得到的免疫血清之间的差别,说明把细胞所含有的物质结合一起的性质,对抗原的性质也有影响。
对血液的个别差异和细胞抗原的各个特性进行扼要的概述后,现在我必须谈谈血型反应的应用。
自从L. Loeb和H. Hirschfeld作了引人注目的观察,即在人的不同种族发现各个血型的相对频率有特定的差异后,在这一方面进行了大量的通信来往。他们最重要的发现是,在北欧,A型比B型多,而在一些亚洲人种,这种位置却颠倒过来,另一个惊人的例子是美洲印第安人,当他们在人种上是纯的,几乎都是O型(Coca;Snyder),据此推论,在少数情况下确有A型和B型存在,这是人种混杂的结果。
我不适合在这里讨论血型的人类学研究和由此得出的结论,且无论如何,不同作者在关于进行解释所必须依赖的普遍原理和个别问题方面,他们的意见总是不同的。但尽管如此,多数观点认为,血型的行为同其他人类学特点一起使我们能对人类种族的关系和起源,作出结论,并对人类学的研究有一定的重要性。
不久,对血型特性本身提出的一种实际应用,是在法医学上辨别人的血斑。用沉淀反应(Kraus;Border;Uhlenhuth)不难确定一个血斑是人的还是动物的,但法医无法区别不同人的血斑。鉴于同种凝集素及其相应的凝集原都可在干燥状态下保持相当一段时间,这问题在某些案件,特别当有问题的血液,即被告和原告人的血液属于不同血型时,可得到解决。当然,使用这种方法的理由不是经常碰到的,尤其在你们国家,使用它的机会很少,但尽管如此,根据Lattes的一篇报告(他是第一个把它用于法庭案件),证明这种测验在一些案件中是有用的,并可作为法官裁决和开释被告人的基础。
血型反应以更大范围被法医用在确立父方身份方面。这类案件能不能作出决议,就靠血型遗传传递的研究了;我们把这方面主要的真实结果归功于Von Dungern和Hirszfeld的工作。由于他们的研究结果,确立A和B凝集原是显性遗传特性,这些特性的传递遵循孟德尔法则。这个特性的重要性在于,事实上在人中间很少有其他任何一种具有如此简单的遗传行为被鉴定起来而又明白无误的生理特性。由上述作者提出的两个独立基因对的遗传理论,经过Bernstein的统计研究后,不得不予以放弃。假若一个群体相当混杂,可根据某种遗传假设能把这个遗传特性的频率计算出来。Bernstein作了这个计算并发现所观察到的数字同根据Von Dungern和Hirszfeld所提出的理论计算所得的数字常有差别。另一方面,当根据在染色体的一个位置上有3个等位基因的假说所作的计算,则发现它们完全相符。这个假设也使AB双亲的孩子们得到可靠的推断。除极少数个别例子外,这个推断也可按照Bernstein的理论来解释,正如Thomsen、Schiff、Snyder、Furuhata和Wiener通过大量研究指出,经验证同样证实了这个推断,因而这个新理论目前已几乎被普遍采纳。
一旦受到法庭的重视,A和B的显性法律就得到确定。这样,在所有那些案件中,当小孩属于A或B型血,而母亲和主张是自己孩子的父亲又都不是这种血型,就能否决父亲一方。这种测验在一些国家,尤其在德国 【39】 和奥地利,还有斯堪的那维亚用得相当多。在去年出现的一份调查报告,Schiff报道了约5000例法医研究,其中父亲一方被否决的案件在8%以上,这期间计算有可能被否决的案件,其比例约为15~100。在赞成这种方法方面,可以提到它也是一种手段使一些父亲能认出他们的私生子。
说明父系诊断如何进一步发展,这或许是有兴趣的。Lansteiner和Levine研究了上述两种血型特性的传递,这些特性可用免疫血清探查出来,并用字母M和N代表它们。根据这些初步结果,最可能的设想是这些性状的出现,是由于有一对彼此都不是显性的基因,所以当两个基因都存在时,就产生一混合型。然后把出现的3种表型M+N+、M+N-和M-N+,实际上解释成第三种为杂合形式,而第一和第二种都是纯合形式。因此,能直接把这种杂合形式看做是一种特殊的表现型。这种假说的结果见下列图解:
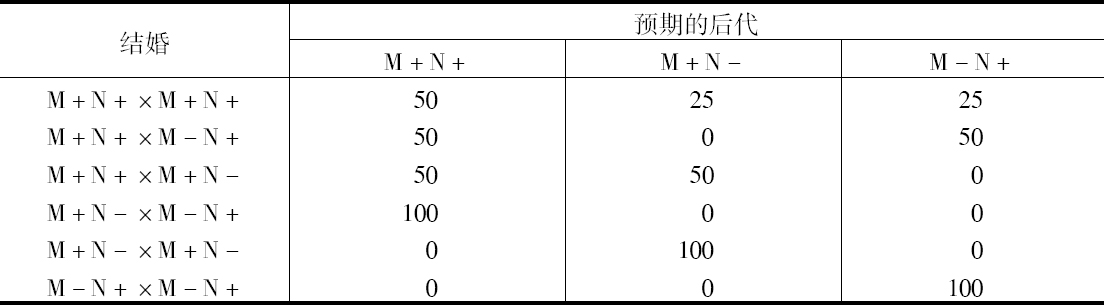
我们自己的观察有几个同这些规律不符的例外,这妨碍我们最终接受这个假说,但这种偏差可能由于私生子或测定血型不是那么简单,这些在实验方法上不完备造成的结果,而事实上Schiff在最近发明的遗传和群体统计方面的观察,经发现同预期情况完全符合一致。Wiener未发表新的结果也很好。
如果这个假设进一步证明是正确的话,排除父亲的可能性必将提高约一倍,即3个案件中有一个案件可以作出判决。然而,即使根据已有的资料,要作出判断还带有很大程度的概率。如果将来的经验能证实这些人们还怀疑的法则是正确的话,那么使用A型和B型的亚型有可能进一步发展。
在医学实践方面,较之上述课题更重要的是把血型反应用在输血上。细述输血的颇饶兴趣的历史显得太长了。它要追溯到几个世纪以前,即哈维(Harvey)发现血循环的时期。在此之前早对进行输血的可能性进行过争论,但是在哈维的伟大发现启示下进行的第一次成功的输血,是1666年Lower在英国用狗实现的,次年Denys在法国,Lower和King在英国首次进行了把动物血输给人。以后致力于发明特殊的器械,而经验证明不需要从血管到血管输血,但只能用去纤维蛋白的血液(Bischoff, 1835年)。在19世纪上半叶可能是Blundell第一次进行了人的输血。
我可以列举引自Snyner的两种评论,用以说明对输血前景的估价是如此的不同。在皇家协会的一段往事中,Sprat(1607)说:“由此引起了新的实验,而主要是输血实验,它很可能以取得卓越的成就而告终。”还是皇家学会的一段往事,我们发现Thompson(1812)的一段话:“早就知道想从这个实践得到预期的好处是幻想而已。”尽管对这个问题作出了巨大的努力和活跃的讨论,却没有达到把这个方法引入到常规的医学实践,这因为纵然它在手术时非常有用,但有时造成严重的症状,甚至导致病人死亡而终于放弃了这个念头。
有关动物血液的注射,Landois的观察是解答为什么发生这种灾难的前提,直到1875年他发现了凝集和溶血作用现象,当血液同一个外来种的血清接触时,这种现象是经常发生的。然而,为什么把人血带入循环也有危险性,这仍然是一个未解之谜,因为很明显,就相同物种的细胞来说,血清或血浆是一种惰性介质,在组织学观察中使用这类血浆这一事实增加了这种信念。
血液个别差异和血型的发现,使这个问题很简单地解决了。动物实验以及特别是在血型测定错误情况下更多的临床试验,肯定了这种关系,且毫无疑问,把能凝集的人血作输血,通常伴随着有害的结果。但是,输血休克的病理发生还没有得到充分的解释。
Ottenberg是第一个考虑到在输血时有凝集素反应,但只有在爆发伟大战争的期间,给体血清选择的输血方法才被广泛采纳,从此这个方法就成为常用的手术。
对血型测定错误的起源,通过直接比较受体和给体的血液可加以控制,对于开始输血时,先注射少量血液等预防措施谈得太详细就离题了。必须提到一点,即不是绝对需要用同一种血型,例如O型血(参见Otterberg)受体血清对它的细胞不发生影响,因而也能使用。然而,在这种情况下,必须小心排除血清具高含量凝集素的给体,因为高含量凝集素有危险性,特别对高度贫血和体弱的患者更是如此。使用这种属于O型的所谓“全能给体”的血液或任一种外来血型的无凝集能力的血液,对防止意外事故和稀有血型的给体,都有巨大的价值。
最明显需要输血的是急性或慢性贫血,如伤口出血或肺出血、产科接生情况下出血、胃溃疡和十二指肠溃疡。在出血情况下输血通常意味着对病人生命的抢救,其效果当然首先要归结于血液的替换,这方面有一个重要的因素,即被转移的红细胞在循环中保持它们功能上的能力可达数周之久。另一个重要的效果是由于提高可凝集能力和也可能刺激血液在骨髓中的再生而产生的止血作用,这一点根据组织学血液图变化已得到肯定。然而,对恶性贫血广泛使用的输血治疗,因发明肝脏治疗现已使前者几乎成为多余的了。
另一个使用输血的广阔场所是重伤和手术引起的休克,并认为在这些病例中输血的效果比注射等渗溶液如含有普通盐溶液的阿拉伯树胶更有效,阿拉伯树胶是由Bayliss在战争期间推荐的。根据这一点,经常在大手术后进行输血,并卓有成效。输血的目的不仅仅是替换血液,也当做一刺激剂来使用。美国的外科医师也推荐对体弱病人在大手术前施行输血。
对血友病、血小板减少性紫癜和一定程度上对粒细胞缺乏症、一氧化碳中毒和烧伤,也都得到了良好的结果,而对败血症等一批其他疾病,曾试用过输血治疗,结果大可怀疑。
我在巴黎微生物学会议上的一次报告中所引用的一些情景,可提供有关使用输血的频率和使用这一方法达到的相对安全性的资料,虽然必须记住这种成功部分地要归功于外科方法的巨大的进展。统计学上有小的变动,这因为同别人相反,有几位作者仍报道个别失败的例子。鉴于这种差别很可能同输血的技术有关,我想,我的判断是根据赞同意见的报告占大量病例而作出的,在这一点上我是公正的。
输血数目大得惊人,似乎使用这种技术太快了。根据医学科学院Corwin博士向我善意提供的统计资料,1929年在纽约作了数万次输血。最新发表的纽约Bellevue医院Tiber的通讯,报道了截至1929年7月的3年半期间,进行了1467例输血。其中有两例死亡。一例因不正确的血型测定,另一例(也能避免的)是用一种所谓“万能给体”O型血给一个处于虚弱状态下的A型婴孩输血。Pemberton在Mayo临床教授班的一次报告中,提到1036例中有3例不幸致死是由于血型测定错误造成的。在Kiel、Beck博士告诉我,约在5年期间进行了2300例输血,没有一次致死的医疗事故。约有2%~3%的病人感到轻微的后效应,如颤抖和发热。Beck报道的一个值得注意的病例是有一个患有恶性贫血症的病人,在3年半内给他输血87次而无任何严重症状。
虽然输血结果良好,但正如我已经说过,仍有报道个别严重的、甚至死亡的医疗事故(它或许不是由技术错误所造成)和经常发生的小干扰。不可能是一种非典型的同种凝集素所表示的血液差异为发生这种情况的一个重要因素,因为如果是这样的话,它们是很容易避免发生的。曾经设想过,但还没有充分肯定,是否强烈的假凝集作用是渗入受体血浆的一种有害效应。所观察到的一些干扰可能是对注入血液中存在的营养物质过敏所致,而另一些则由于过早输血而形成抗体作用的结果。另一个尚未得到充分研究的问题为是否因为各个蛋白质之间存在着差异,而真若如此,则是否这些差异可导致抗体形成。
总之,输血的结果已经令人十分满意,而我们有理由希望,通过对具有不理想后效应的病例做彻底的研究,将有助于我们估价所推测原因的意义和(或)可揭示一些未知的原因,而将最终从本质上消除输血时仍然碰到的这种小的危险性。
除了解决这个实际问题外,我们所谈到的这个课题也可以通过以下方面得到发展:从总的方面研究个别血清差异的生物学问题,特别是通过进一步改良测定人血更细微的个别差异的技术,以及继续进行人和动物血清学方面的血液差异的遗传分析。从已经作过的工作结果来看,人的染色体对中,除了性染色体外,至少有两对具有一种特别的特性(也参见F. Bernstein)。
玉米细胞学和遗传学交换的关系 【40】
哈里特·克赖顿(H. B. Creighton) 【41】
巴巴拉·麦克林托克(B. McClintock) 【42】
(1931年)
交换的遗传学研究要求在同一个连锁群中两个等位因子处于杂合状态。分析在两个点上形态上可予区别的同源或部分同源染色体的行为,将得到细胞学交换的证据。本文目的为证明细胞学交换是随同遗传学交换而发生的。
某些玉米系的第二个最小的染色体(第九染色体),在其短臂的末端有一个明显的结。染色结在以后各代中的分布同一个基因的情况相似。如果把一株第二个最小染色体的两端都有染色结的植株,同一个两端没有染色结的植株杂交,细胞学观察表明,在它们产生的子一代个体中,同源染色体对中只有一个成员有染色结。当把这种个体同一个两个染色体成员都没有染色结的植株作回交,后代中有一半在染色结中是杂合的;有一半完全没有染色结。因此,染色结是该染色体的一个连续的特征。当染色结出现在一个染色体而不出现在其同源染色体时,就明显地使该染色体成为异型的。
早先报道过,某些玉米系在第八染色体和第九染色体之间会发生互换。互换片大小相等;第九染色体的长臂相对地增长,而第八染色体的长臂则相应地缩短。如一个具有这两个互换的染色体的配子同一个具有正常染色体组的配子交配,它所产生的个体的减数分裂,其特点为同源部分并排地联会。因此,这就有可能在染色结和互换点之间发生交换。
过去的报道也指出,这样的个体,只有具有两个正常染色体(N、n)或两个互换染色体(I、i)的配子,即这一种或另一种排列都是完全的染色体组的配子,才是有功能的。所以有功能的配子或具有较短的正常的有结的染色体(n),或具有较长的互换的有结的染色体(I)。因而,当把这种植株同一个具有正常染色体的植株进行杂交,而所得个体的减数分裂前期出现10个二价染色体,就表明前者有功能的配子具有正常的染色体。如另一些子一代个体,在减数分裂晚前期出现8个二价染色体,加之4个染色体形成的一个环,则说明另一种有功能的配子具有互换的染色体。
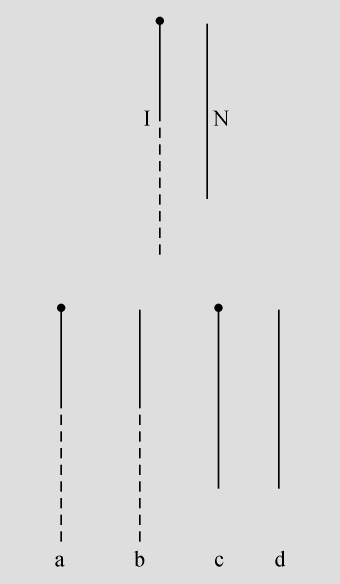
图1
上图:研究交换的染色体图解;
下图:在一个具有上述构造的植株的配子中发现的染色体类型图解。
a:带染色结的交换过的染色体;
b:无染色结的交换过的染色体;
c:带染色结的正常染色体;
d:无染色结的正常染色体。
a和d为非交换类型;b和c为交换型。
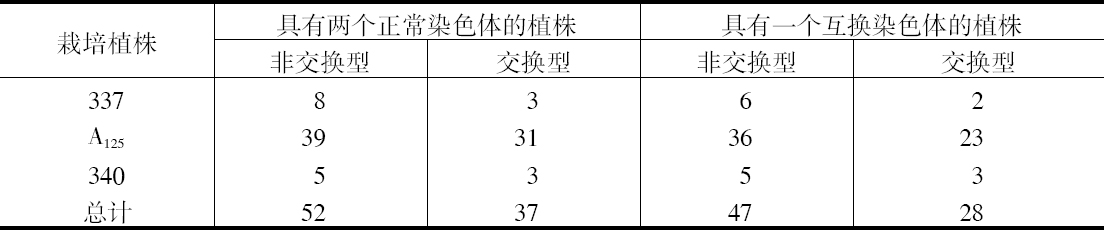
如把一第九染色体为正常,无结的配子同一个互换的具染色结的配子交配,显然,由此产生的个体在减数分裂前期,这两个沿着其同源部分联会的染色体,在它们的两端是明显不同的。如果不发生交换,这种个体形成的配子将具有或有染色结的交换过的染色体(见图1,a),或是无染色结的正常染色体(见图1,d)。由于交换会形成含有带染色结的正常染色体(见图1,c),或无染色结的交换过的染色体(见图1,b)。如把这种个体同具有两个无染色结的正常染色体的植株杂交,所产生的个体将有4种。无交换的配子产生的个体有两种:①在减数分裂前期为10个二价染色体,第九染色体无染色结,说明带有d型染色体的配子是有功能的;②4个染色体形成一个环,带有一单个的明显的染色结,说明带有A型染色体的配子是有功能的。交换类型的个体有以下两种:①10个二价染色体和二价的第九染色体带有一个单染色纽;②4个染色体连成一个环,无染色纽,说明c和b交换配子都是有功能的,这种杂交的结果见表1中的337系。同样地,如果这种植株同一种在第九染色体的两端都有染色结的正常植株杂交,并发生交换,它们产生的个体就有4种。代表非交换类型的有两种植株:①染色结是纯合的并具有互换过的染色体;②染色结是杂合的并具有两个正常染色体。在染色结和互换染色体之间进行交换而产生的有功能的配子,产生以下两种个体:①染色结是杂合的并具有互换染色体;②染色结是纯合的并具有两个正常染色体。这类杂交结果见表1 A125 和340系。尽管数据少,但它们是一致的。从这些数据测出的染色结和互换染色体之间的交换量大约为39%。
表1  ×无染色结的正常染色体337系植株,和有染色结的正常染色体A125 和340系植株
×无染色结的正常染色体337系植株,和有染色结的正常染色体A125 和340系植株
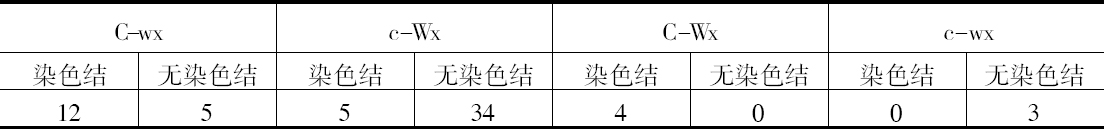
前一篇文章指出,具染色结的染色体携有带色糊粉(C)、皱缩胚乳(sh)和蜡质胚乳(wx)的基因。此外,看到以互换点为起点,这些基因的次序为wx-sh-C。还有,有可能所有这些基因都位于具染色结的染色体的短臂上。因此,可以指望染色结和这些基因之间为一连锁。
一个具有正常染色体组的植株,其第九染色体有一个染色结并携有C和wx基因。它的同源染色体无染色结并携有c和Wx基因。非交换型配子应具有一带染色结的C-wx染色体,或一无染色结的c-Wx染色体。在第1区(染色结和C之间)的交换得到无染色结C-wx的染色体和有染色结的c-Wx染色体。在第2区(C和wx之间)交换则得到带染色结的C-Wx染色体和无染色结的c-wx的染色体。这种植株同一无染色结的c-wx型植株的杂交结果见表2。在相互干扰的基础上,可以预期,当C和wx之间发生交换时,染色结和C会仍然在一起;因此,从有色淀粉(C-Wx)子粒长成的个体应具有染色结,而从五色、蜡质(c-wx)子粒长成的个体则应当不带染色结。虽然数据不多,却令人信服。显然在染色结和C之间有着一种非常密切的联系。
表2  ×无染色结-c-Wx
×无染色结-c-Wx
为了取得细胞学和遗传学交换之间的相关性,必须有一种植株,它在染色结,基因c和wx以及互换方面是异型的。338号植株(17)一个染色体有染色结,基因C和wx以及第八染色体的互换片。另一个染色体是正常的无染色结并具有c和Wx基因。把这个植株同另一个植株杂交,该植株具有两个正常的但分别带有c-Wx和c-wx基因的无染色结的染色体。这一杂交图解如下:
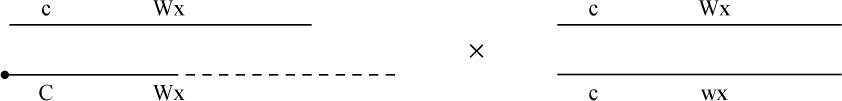
杂交的结果见表3。在这种情况下所有带色的子粒都长成有染色结的个体,而全部无色子粒则长成无染色结的个体。
表3 
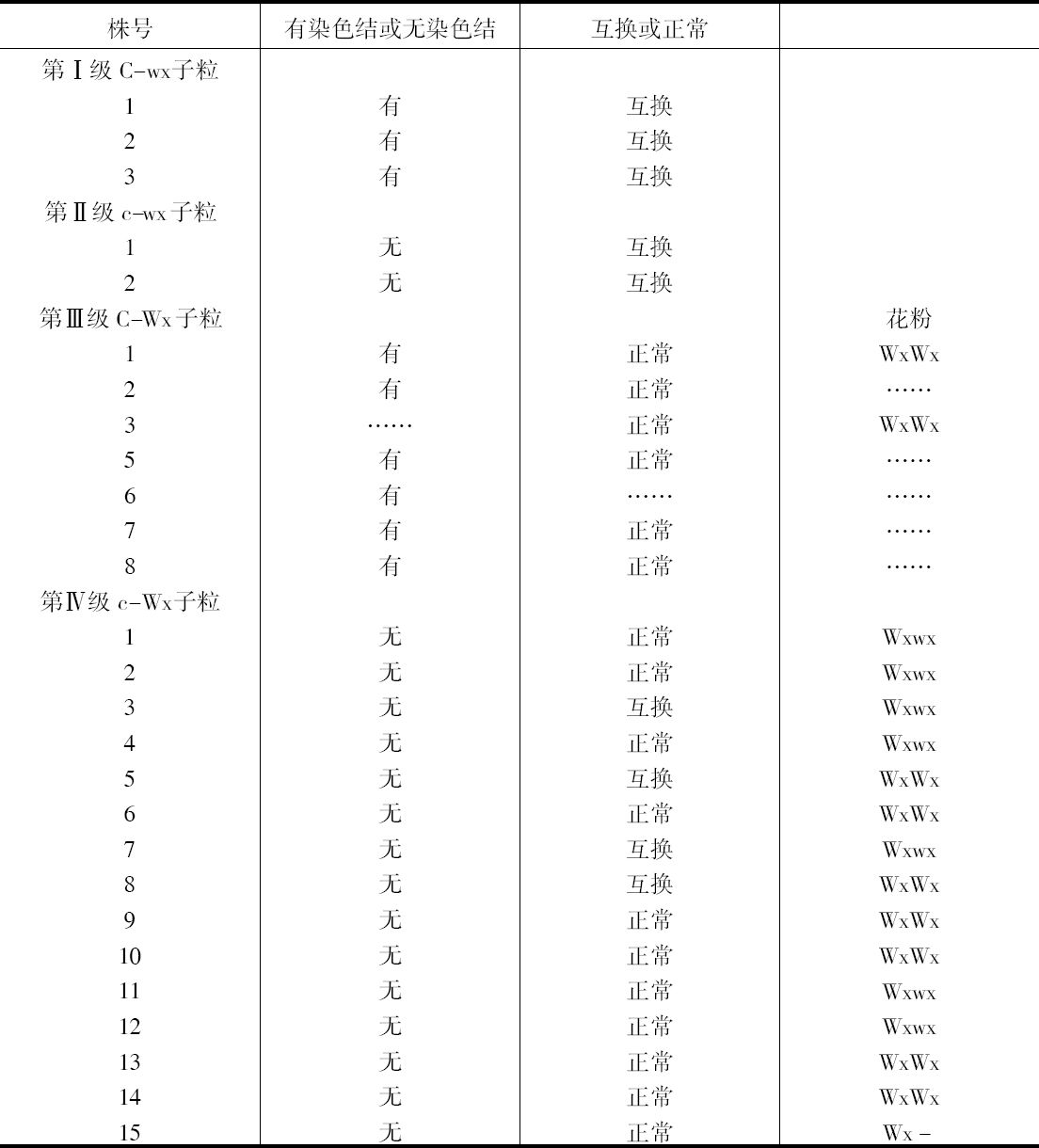
染色结和互换点之间的交换量为39%左右(见表1),c和互换点之间约为33%,wx和互换点之间则为13%。记住这些资料,就能对表3所得的数据进行分析。由于果穗上只结少数子粒,数据必然很少。第Ⅰ级的3个个体显然是非交换类型。第Ⅱ级的个体是第2区即c和wx之间交换产生的。在这种情况下第2区的交换决不同第1区(染色结和C之间)或第3区(wx和互换点之间)的交换一起发生。第Ⅲ级的全部个体都具有正常的染色体。遗憾的是,经检验有染色结的6个个体中只有一个得到了花粉。这一个个体显然可猜想到它是通过第2区的交换所产生的配子长成的。第Ⅳ级更难以分析。第6、9、10、13和14号是正常和带有WxWx基因的植株,因而它们代表非交换类型。在正常、Wxwx级中预期非交换类型数目相等。第1、2、4、11和12号植株或许是这种类型。通过一个c-Wx配子同一个在第2和第3区上双交换所产生的配子结合,而得到这种类型是可能的,但却未必能发生。第5和第8株是第3区上发生的单交换,而第3和第7株则或许代表第2区或第3区的单交换型。
上述证据说明,发生着细胞学交换,而且它是随同遗传学交换的预期类型一起发生的。
结论 已经证明在两个区上异型的配对染色体,在交换染色体部分的同时,交换位于这些区的基因。
一种研究染色体重排和绘制染色体图的新方法 【43】
西奥菲勒斯·喷特(T. S.Painter) 【44】
(1933年)
人们早就知道,许多双翅目幼虫的有功能的唾腺,其染色体为一伸长和有环的结构。去年,作者主要用醋酸洋红法对果蝇(Drosophila melanogaster)幼虫的唾腺染色体进行了研究。根据这一研究,得出如下结论:
(1)每一个染色体有其明确而稳定的形态特性,并由节段组成,每一个节段的特有模式是染色质线或较宽的带盘绕在非染色质基质的外面。因此,相同的染色体或特定的部分,在一个个体的细胞内,或一个物种的不同个体中,都容易被识别出来。如由于某种形式的离位(易位、倒位等),一个或更多的节段的位置发生了变迁,能测定出断裂确切的形态学上的点(或几个点),并能在它们新的位置上鉴定出这种节段。这一发现首次使我们持有一种染色体分析的定性方法,并只要通过研究可知遗传性状的染色体重排,从而知道任何一个成分正常的形态学,我们就能得出基因位点的形态学位置,并能作出比迄今为止更为大大精确的染色体图。
(2)在老龄幼虫,同源染色体进行体细胞染色体联会。这种联合绝不是一种简单的并列,因为染色体成分以最确切的方式线对线地配对;并形成一明显清楚的结构。如果同源染色体之一有一段位置颠倒,我们就得到像在减数分裂中预期的典型的倒位图像。如果同源染色体之一在一些点上发生缺失,则除缺失点上正常成分常常被扣住外,两个配偶对进行联合。这样我们就很容易精确地决定一个染色体缺少了多少。在唾液腺中使同源染色体联合起来与减数分裂的情况相同,这一点是可能的,而就我们所知,这些特别的染色体从不分裂,我们至少能研究在联会时畸形染色体是如何联合的,这一事实对遗传学家有巨大的价值。
(3)在唾液腺中,V形常染色体的两臂看来是在它们之间无明显联系的独立成分。其结果是在体细胞染色体联会后,我们发现核里面有6个成员,而不是单倍体数。
(4)X染色体的惰性区域,看来不像这个成分的有机部分,它也不像核里任何别的已弄清楚的形式。同样地,Y染色体经鉴定为一短片的这个唯一的部分,它在形态学上同X染色体右方末端节段的那一部分是同源的。X染色体的这一部分(见图1)携带着成串的正常等位基因。X和Y染色体的惰性物质,在个体发育中都通过减少或一些相似的过程而消失掉,或者这种物质以某些同该染色体无明显联系的尚未认出的形式,存在于唾液腺核中。惰性面积约占卵原细胞中期染色体体积的3/8。
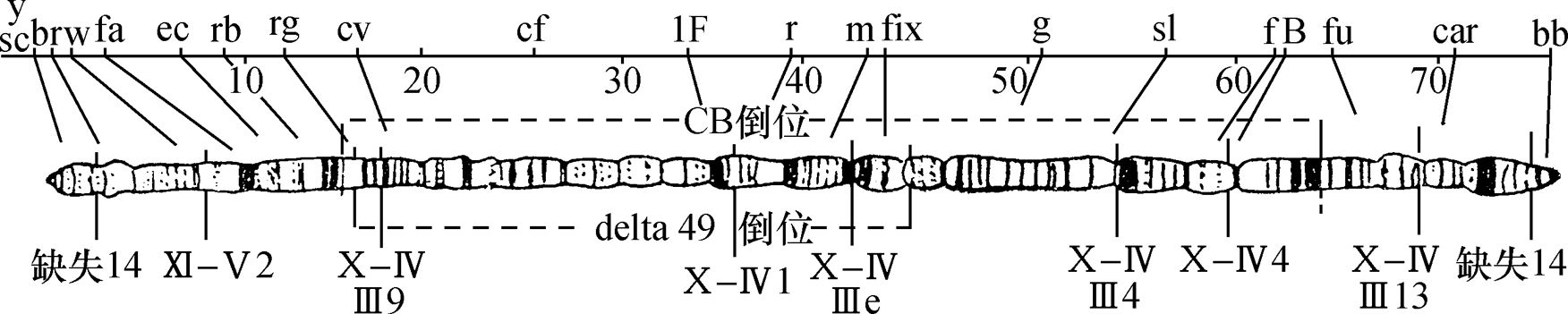
图1
图1是通过把各区照相清晰的草图连起来绘出的X染色体图。省去其细节部分。图1上方为长度同X染色体相同的交换图。基因定位位点的符号在线上一起表示它们近似的形态学位置。X染色体上所表示的断裂点,其断裂名称列在下方。这样,缺失14(在左方)使X染色体在盾片和宽度的位点之间断开。图中可见断裂的形态学上的点,当然,盾片必定在断点的左方,宽度在右方。以相似的方法已测出其他基因位点的位置。遗传学家将有兴趣注意到图中表示的C1B和△49倒位的形态学(和遗传学)限制,并且细胞学和交换图之间完全相符。
作者有两篇论文正在付印,一篇涉及唾液腺染色体的技术及其总的形态学性状;另一篇为X染色体的细节研究,从中得出此处提出的X染色体图。相似的常染色体研究已进行了一段时候,一些研究人员正从事于用这个新的方法来揭示各细胞学和遗传学问题。
链孢霉生物化学反应的遗传控制 【45】
乔治·比德尔(G. W.Beadle) 【46】 E. L. 塔图姆(E. L. Tatum)
(美国 斯坦福大学生物系,1941年)
从生理遗传学的观点来看,一个生物体的发育和功能主要是由一个完整的生化反应系统构成的,这些生化反应以某种方式受到某些基因的控制。据推测,这些基因本身就是这个系统的一部分,它们或者是以酶的方式直接起作用,或者是决定着酶的特异性,从而控制或调节这个系统中的特异反应[1 ] 。这种推测看来是可靠的。这个系统中的各个成分好像是以复杂的方式互相起作用的,单个基因各部分的合成也似乎依赖于其他基因的作用;基因对生化反应的控制,有简单的一对一的关系,也有更复杂的关系。在研究基因作用时,生理遗传学家总想决定已知遗传特性的生理和生化基础。对植物花青甙色素[2 ] 、酵母菌的蔗糖发酵[3 ] 以及一些其他情况[4 ] 的研究所进行的探讨,事实上已经证实:很多生化反应是由特异的基因、以特异的方式控制的。而且这类研究趋向于支持酶的特异性是在基因水平上控制的这一假说[5 ] 。然而,这个方法有一定的局限性,其中最大的局限性,可能在于研究者一般只局限于研究非致死性的遗传特性。这些特性在某种程度上与非主要的、所谓末端反应有关[5 ] 。对遗传学研究的这些选择,可能与当时认为基因只能控制表面现象的看法有关,这种看法现在已经消失了。其次还有一些困难,即研究这个问题的标准方法,所选用的特性必须具有明显的表型。这样的特性大都与形态变异有关,而且与它们有关的生化反应系统,似乎都是相当复杂、很难分析的。
考虑到刚才提到的那些问题,在研究发育和代谢反应遗传控制的一般问题时,为了阐述已知的生化反应是否由基因控制,以及基因如何控制,我们不是去解决已知遗传特性的化学基础,而采用了逆转正常程序的方法。子囊菌链孢霉为这种方法提供了很多有利之处,它很适于作遗传学研究[6 ] 。因此,我们围绕着链孢霉制定了研究方案。这个方案是以下述假定为基础的,即X射线处理将诱发控制着已知特异性化学反应的那些基因发生突变。如果菌体必须完成某种化学反应才能在一种特定培养基中生存下来,那么一个不能完成这种反应的突变体,在这种培养基中显然会死亡。但是,如果在这种培养基中,加入这个遗传阻断反应的主要产物突变体就能生长的话,突变体就能保存下来,并用于研究。根据这个理由,我们打算选一个适当的例子来说明这个实验程序。粗糙链孢霉的正常菌株能利用蔗糖做碳源,因此菌株能完成与这种糖的水解有关的特异性酶促控制反应。假定这个反应是受遗传控制的,就有可能诱发一个基因突变,导致在与原来同样条件下不再能水解蔗糖。一个携带这种突变的菌株,在以蔗糖做唯一碳源的培养基上不能生长,但在含有另外一些可利用碳源的培养基上是可以生长的。换句话说,在含有葡萄糖的培养基上就可以得到和保持这样的一个突变菌株,再把它转移到蔗糖培养基上,根据它不能生长,就可以知道它丧失了利用蔗糖的能力。
在很多代谢过程中都建立起了基本相似的程序。如果我们的推测是正确的,那么合成生长因子(维生素)、氨基酸和其他必须物质的能力,通过基因突变也可能丢失。从理论上讲,只要缺少的物质可以从培养基中得到,而且这种物质能透过细胞壁和原生质膜的话,任何一种代谢缺陷都是可以“克服”的。
依据这类实验的经验,我们曾设计了一种程序,在这种程序中,X射线处理过的单孢子培养物,可以在一种完全培养基上生长起来(完全培养基所含的成分实际上和有机体正常合成的成分一样多)。然后再把它们转移到一种无机培养基上进行检验(无机培养基是一种要求有机体能完成它能够完成的所有必需的合成作用才能生长的培养基)。实际上,完全培养基是由洋菜、无机盐、麦芽汁、酵母浸出液和葡萄糖组成的。无机培养基含有洋菜(任意的)、无机盐、生物素以及一种双糖、脂肪或更复杂的碳源。生物素是一种生长因子,连野生型的粗糙链孢霉菌株也不能合成它[7 ] ,使用的生物素是一种商品的浓缩液,每毫升含生物素100微克。如果一个菌株失去了合成任何一种必需物质的能力,若这种必需物质在完全培养基中含有,而在无机培养基中没有,那么这个菌株在前一种培养基上能够生长,而在后一种培养基上则不能生长,据此可以得知它不能合成某种物质。然后再用一系列的方法进一步测定该菌株不能合成哪种物质或哪几种物质。以后的测定包括检验各突变菌株在含有下列成分的无机培养基上的生长情况:①加入已知的维生素;②加入已知的氨基酸;③用葡萄糖代替无机培养基中较复杂的碳源。
从单个子囊孢子得到的各个菌株是分别从减数分裂前经X射线照射过的粗糙链孢霉和好食链孢霉的子囊壳中分离出来的。在大约2000个这样的菌株中,找到了3个突变体,它们在完全培养基上生长基本正常,在用蔗糖做碳源的无机培养基上完全不能生长。其中一个菌株(从好食链孢霉中得到的)不能合成维生素B6 (吡哆醇)。第二个菌株(从好食链孢霉中得到的)不能合成维生素B1 (硫胺素);进一步的测定说明,这个菌株能合成B1 分子的一半,即嘧啶,但不能合成它的另一半,即噻唑。假如在无机培养基中单独加入噻唑,菌株生长就基本正常。第三个菌株(从粗糙链孢霉中分离到的)不能合成对氨基苯甲酸。这个突变菌株在添加对氨基苯甲酸的无机培养基上,生长完全正常。在诱发引起代谢缺陷的菌株中,我们只对维生素B6 缺陷菌株进行了遗传分析 【47】 。硫胺素缺陷和对氨基苯甲酸缺陷菌株的详细情况,将在以后给予说明。
定性的研究情况清楚地表明:维生素B6 缺陷突变体,培养在一种含有合成维生素B6 盐酸盐(每25毫升中,含有1微克或1微克以上)的培养基中,它的生长速度和特性与培养在同样的、但不加B6 的培养基中的正常菌株是很接近的。B6 的浓度较低时,生长速度会减慢。突变体的生长对培养基中维生素B6 定量的依赖关系研究的初步结果总结于表1。其他实验也得到了基本相似的结果,在数量上与表1的结果是大体一致的。很清楚,对培养条件进行详细的研究之后,可以根据这个突变体的重量增加的速率,精确地测定维生素B6 的含量。
表1 好食链孢霉的维生素B6 缺陷菌株
(培养在含无机盐① 、1%蔗糖和每毫升0.004微克生物素的液体培养液中的生长情况。
温度25℃。生长期,从用分生孢子接种起共6天。)
①在进行链孢霉的研究工作中,全都采用Fries命名的第3号盐混合物,其成分如下:
酒石酸铵5克;NH4 NO3 1克;KH2 PO4 1克;NaCl 0.1克;MgSO4 ·7H2 O 0.5克;CaCl2 0.1克;1%的FeCl3 溶液10滴;水1升。
链孢霉不能用酒石酸盐作为碳源。
我们发现粗糙链孢霉菌丝体边缘,沿着加了一半洋菜培养基的水平玻璃培养管的方向向前扩展,这为研究生长因子的定量效应提供了一个简便的方法。所用的试管,内径约为13毫米,长度约40厘米。两头各有长5厘米的向上翘起的部分,大约成45°角。向管子中加入约一半的洋菜培养基,把管子的主要部分放成水平位置。管子向上翘起的两头用棉塞堵住。从一头的洋菜表面接种,每隔一定的时间记录菌丝体前沿的位置。向前扩展的菌丝体形成的边缘是很好确定的,不难精确到1毫米以上,菌丝体沿着这种管子向前扩展与时间成严格的线性关系,而扩展速率与管子长度无关(在1.5米以下)。把管子内径减到9毫米,或者封闭一头,或两头都封闭,都不能改变其速率。由此看来,好像在这种管子中,气体的扩散不是限制因子。
维生素B6 缺陷菌株在装有含不同数量B6 的洋菜培养基的水平管中的生长结果见图1和图2。很明显,菌丝体向前扩展的速率是培养基中维生素B6 浓度的函数 【48】 。同时还清楚看出,突变体在加了B6 的培养基上的生长速率,和正常菌株培养在不加B6 的这种培养基上的生长速率没有明显差异。这些结果与下述假设是一致的:假设认为维生素B6 缺陷菌株和正常菌株之间最根本的生理学差异,就在于前者不能合成维生素B6 。这个合成过程肯定不止一个步骤,据推测,维生素B6 生物合成过程中发生突变的这个基因只与一个特异的步骤有关。
②原文中0.05,系错误。——译者注
图1 在水平管子中,好食链孢霉的正常菌株(上面的二条)和维生素B6 缺陷菌株(其余的)的生长曲线
在这个图中,每一条连续曲线的纵坐标标度都移动了一定的量。每条曲线右边的数字是表示维生素B6 的浓度,即在每25毫升培养基中B6 的微克数。
为了确定维生素B6 缺陷的遗传特性,进行了正常菌株和突变体之间的杂交。杂交的技术和分生孢子的分离方法已经建立,见Dodge和Lindegren[6 ] 的叙述。分离了杂交过的24个子囊的分生孢子,并记录了它们在子囊中的位置。由于一些仍未搞清的原因,这些分生孢子多数不能萌发。但其中的7个子囊,每个中有一个或多个孢子能萌发。把它们培养在一种含有葡萄糖、麦芽浸出液和酵母浸出液的培养基上,它们都能正常生长。把它们培养在一种不含B6 的培养基上,正常的和突变体的培养物是有差别的。在这个培养基上,突变菌株几乎不能生长,而非突变菌株能正常生长。此结果总结于表2。从这些相当有限的资料可以清楚地看出,如果维生素B6 缺陷与正常菌株只有一个基因之差,那么不能合成维生素B6 的能力是可以遗传的。
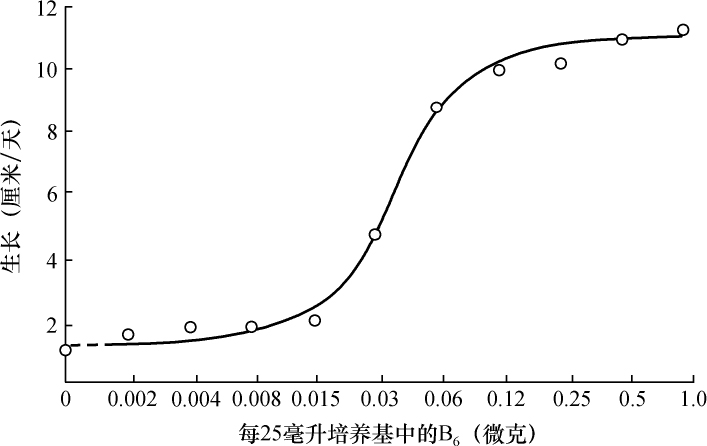
图2 生长速度(厘米/天)与维生素B6 浓度之间的关系
表2 好食链孢霉的正常菌株和维生素B6 缺陷菌株杂交,得到的单个子囊孢子培养物的分类结果
①—线表示从外观可以看出孢子没萌发。
②pdx表示在不含维生素B6 的培养基上生长很弱。
③N表示在不含维生素B6 的培养基上正常生长。
④*表示分离出来的孢子2,3,4位置搞乱了。其中2个孢子萌发了,经证明都是突变体。
以上总结的初步结果向我们指出:概述的这个方法可以为进一步研究基因如何调节发育和功能,提供了一种很有希望的方法。例如,通过寻找不能完成一个已知合成过程的某个特定步骤的许多突变体,就可以检验通常是否只有一个基因与一个指定的特异性化学反应的直接调节有关。
从生物化学和生理学的观点来看,这种方法作为发现具有生理学意义的其他物质的一种技术来说,也是很有价值的。因为所用的完全培养基可由酵母浸出液来制备,也可用正常的粗糙链孢霉浸出液来制备。很明显,如果一个菌株通过突变失去了合成一种必需物质的能力,它就可以用做分离这种物质的指示菌株。也有可能这种物质是以前尚不了解,而且对任何菌体的生长都是必需的一种物质,这样,我们可以去发现新的维生素。按同样的方法,也可以去发现其他的必需氨基酸(如果它存在的话)。事实上,我们已经找到了一个突变菌株,它可以在含有Difco酵母浸出液的培养基上生长,但却不能在至今为止我们测定过的任何一种合成培养基中生长。显然,在酵母中存在一种我们至今尚不了解的生长因子,它是粗糙链孢霉生长所必需的。
摘要
用粗糙链孢霉研究出一种方法,用它可以发现和保存X射线诱发的突变菌株,这类突变菌株由于不能完成特异的生化过程而被鉴别出来。
采用这种方法得到了3个突变菌株。其中一个突变菌株合成维生素B6 的能力已完全或大部分丧失了。第二个突变菌株不能合成维生素B1 分子噻唑部分。第三个突变菌株不能合成对氨基苯甲酸。因此,清楚地说明这些物质对粗糙链孢霉来说,都是必需的生长因子 【49】 。
把维生素B6 缺陷突变体(一个不能合成维生素B6 的突变体)培养在含有B6 的培养基中,它的生长是培养基中B6 含量的函数。本文还介绍了一个方法,可以用于测定菌丝体沿着加了一半洋菜培养基的水平管子向前扩展的线性生长情况。
显然,菌体能否合成维生素B6 这种生长必需因子的能力,只有一个基因之差。
参考文献
[1 ]Troland, L. T., 1917 Amer. Nat., 51 321~350; Wright, S., 1927 Genetics, 12 , 530~569; and Haldane. J. B. S., 1937 In perspectives in Biochemistry, Cambridge Univ. press, 1~10.
[2 ]Onslow, Scott-Moncrieff, and others, 1940 See review by Lawrence, W. J. C., and Price, J. R., Biol. Rev., 15 , 35~58.
[3 ]Winge, O., and Laustsen, O., Compt rend. Lab. Carlsberg, 1939 Serie Physiol., 22 , 337~352.
[4 ]Goldschmidt, R., 1939 Physiological Genetics, McGraw-Hill, p.1~375 and Beadle, G. W., and Tatum, E. L., Amer. Nat., 75 , 107~116.
[5 ]Sturtevant, A. H., and Beadle, G. W., 1931 An Introduction to Genetics, Saunders, p.1~391, and Beadle, G. W., and Tatum, E. L., loc. cit., footnote 4.
[6 ]Dodge, B. O., 1927 Jour Agric. Res., 35 , 289~305; and Lindegren, C. C., 1932 Bull. Torrey Bot. Club, 59 , 85~102.
[7 ]Buler, E. T., Robbins, W. J., and Dodge, B. O., 1941 Science, 94 , 262~263.
对引起肺炎球菌类型转化的物质化学特性的研究 【50】
从肺炎球菌Ⅲ型分离出来的脱氧核糖核酸引起的转化
奥斯瓦尔德·埃弗里(O. T.Avery) 【51】
C. M. 麦克劳德(C. M. MacLeod)M. 麦卡蒂(M. McCarty)
(美国 洛克菲勒医学研究所附属医院,1944年)
生物学家很久以来就希望用化学方法,按照人们的需要,引起高等生物的特异性变化,并使这些变化能作为遗传性状连续传递下去。微生物中,细胞结构和功能上能够遗传的特异性变化,最明显的例子就是肺炎球菌类型的转化。上面这些变化都是用实验方法可以引起的,而且在限定的、适当控制的条件下能重复发生。转化现象首先是格里菲斯(Griffith)[1 ] 提出的,他把从一种特异类型的肺炎球菌分离出来的无毒而且能形成荚膜的(R)变种,成功地转化成为一个完全不同的、能很好形成荚膜的、强毒的(S)特异类型。肺炎球菌种间的转化现象是一个典型例子,它足以说明最初所采用的转化技术,而且还可以用来说明转化现象所引起的巨大变化。
格里菲斯发现了用少量活的、从肺炎球菌Ⅱ型得到的R培养物和少量经热处理杀死的Ⅲ型(S)肺炎球菌一起,从皮下注射到小鼠中,常常引起感染,而且从感染的小鼠心脏血液中,经过纯培养得到了Ⅲ型肺炎球菌。R型菌株是无毒的,它自己不能引起致死的菌血症;另外,热处理过的Ⅲ型细胞悬浮液中也不含活的细菌;这些事实令人信服地证明:在这种条件下生长的R型菌,重新获得了Ⅲ型肺炎球菌的荚膜结构和生物学特异性。
格里菲斯最初的发现,后来又被国外的Neufeld和Levinthal[2 ] ;Baurhenn[3 ] ,以及我们实验室的Dawson[4 ] 进一步证实了。此后,Dawson和Sia[5 ] 又成功地进行了离体转化,他们是把R细胞培养在含有抗R血清和热处理杀死了的、能形成荚膜的S细胞的液体培养基中完成这项转化的。他们指出:根据这个反应系统中所用的S细胞的类型特异性,在试管中和动物体内一样,也可以选择性地引起转化。后来,Alloway[6 ] 把S细胞经过Berkefeld滤器过滤,除去产生的各种成分和细胞碎片,得到一种无菌提取液,用它在体外也引起了特异性转化。据此他提出:溶有活性转化物质的粗提液在引起特异性转化上,和制备提取液所用的完整细胞是同等有效的。
在病毒中,还有与肺炎球菌类型转变相类似的另一个转化的例子。Berry和Dedrick[7 ] 成功地把兔纤维瘤病毒转变成了有侵染性的黏液瘤病毒。他们用活化的纤维瘤病毒与热处理失活的黏液瘤病毒悬浮液一起给兔子接种,使兔子产生感染了多发性黏液瘤的症状和病理损伤特征。此后,转化了的病毒再感染动物,所产生的感染和天然发生的多发性黏液瘤感染是一样的。Berry[8 ] 把经过洗涤的黏液瘤病毒加热杀死,用其上清液也成功地引起了同样的转化。就这些病毒的情况来说,所使用的方法与Griffith在肺炎球菌类型转化中所使用的方法是相类似的。以后的研究者进一步证实了这些结果[9 ] 。
本文对肺炎球菌的类型转化现象进行了更详细的分析。最主要的目的是试图从细菌的粗提取液中分离出活性物质,并弄清其化学特性,或者至少要充分确定它属于哪一大类化学物质。从这个研究目的出发,我们选用了我们了解最多、可能也是最适于进行分析的一个典型转化系统作为工作模型,这个特殊系统就是从不形成荚膜的肺炎球菌Ⅱ型R变种到肺炎球菌Ⅲ型的转化。
实验
肺炎球菌不同类型的离体转化实验,除了需要有效提取液外,还需要充分满足某些培养条件才能转化成功。转化不仅需要有最适于细菌生长的肉汤培养基,而且还必须补加血清或具有某些特殊性质的血浆液。后面还将进一步讲到,R变种还必须处于一种能接受转化刺激的活化时期。为了方便起见,在转化实验中,这些因素统称为反应系统。对这个系统的每一个组分都必须弄清楚,实验才能获得一致的、可以重复的结果。这个系统的各种组分,按下列顺序逐一叙述:
1.营养肉汤
把含1%新蛋白胨、但不加葡萄糖的牛心肉浸汤,调pH到7.6~7.8,用做基本培养基。各批肉汤在支持转化的特性上表现出明显的、预先没料到的差异。但发现,按Macleod和Mirick[10 ] 在除去磺胺抑制物时所叙述的方法,把肉汤经过炭吸附,可以大大消除这些差异。以后,在制备效果恒定的、用于测定提取液转化活性的肉汤时,将这一步骤定为常规操作。
2.血清或血浆液
在离体转化的第一次成功实验中,Dawson和Sia[5 ] 发现,在培养基中加入血清是非常重要的。实验中用了抗R肺炎球菌的兔血清,因为曾经发现,生长在含有抗R血清的培养基中的一种R型肺炎球菌被诱导回复成了其同源的S型。后来Alloway[6 ] 发现,腹水液、胸液以及正常的猪血清都含有R抗体,因此在这个反应系统中都可以代替抗肺炎球菌的兔血清。某种形式的血清是很重要的,就我们所知,离体转化在没有血清或血浆液的条件下从未成功过。
在我们的研究中,几乎无例外地应用了人的胸膜液和腹水液。不同批号的血清,效果是不同的,观察到的这些差异与R抗体浓度无关,因为发现许多高效价的血清并不能促进转化。这些事实表明,并不是R抗体,而是另外一些因子起了作用。
发现各种动物的血清,不管它们的免疫性如何,都含有一种酶,这种酶能破坏有效提取液中的转化因素,它的性质及其作用的专一性底物将在本文后面提到。把血清在60~65℃加热,这种酶就失活了。在能破坏这种酶的温度下,热处理过的血清在转化系统中常常表现出是有效的。进一步的分析还说明,能分解转化物质的酶已被失活、但还含有R抗体的一些血清仍不能促进转化。这一事实说明,血清中还有一种很重要的因子,这种因子的浓度随着不同的血清而有变化。目前对其特性尚不清楚。
目前,在挑选合适的血清或血浆液时,除了实际测定它们促进转化的能力外,没有其他可遵循的标准。幸好,所需要的特性在一个相当长的时间内是稳定的,而且不会受损害。发现把血清放在冰箱中保存几个月后再重新测定时,它促进转化的效果降低很少,或根本不降低。
对血清中的这些因素及其在转化系统中作用的了解,使我们更容易确定标准的培养条件,以获得一致的、可以重复的实验结果。
3.R菌株(R36A)
研究中所用的不能形成荚膜的R菌株,是从肺炎球菌Ⅱ型的一种强毒S培养物分离得到的。肺炎球菌各种变种,不管是哪种类型,只要它们不形成荚膜,也就丢失了类型的特异性和感染动物的能力。据此进行鉴定,从而选出R菌株。这些变种之所以定名为R型,只是根据它们在人工培养基上形成的菌落表面是粗糙的;而能形成荚膜的S细胞的菌落表面是光滑、发亮的。
把肺炎球菌Ⅱ型的亲代S培养物,在含有抗肺炎球菌Ⅱ型兔血清的肉汤中连续培养36代,然后分离变异株,从而获得R36A这个菌株。菌株R36A已完全丧失了亲代S菌种的典型特性,成为一种无毒的、不能形成荚膜的R变种。由S→R的变化常常是可逆的,这说明R细胞退化得不是太远,通过在动物体内连续传代或在抗R血清中连续多次继代培养,常常可以使R型回复成原来的特异类型。因此,在这样的条件下,R培养物必定回复成原来亲本能形成荚膜的那种特异类型[11 ] 。但菌株R36A却已变成稳定的R型,从未自发回复成SⅡ型。而且,试图在上述条件下引起它回复的想法,都未获得成功。
在同一种类型中,S R之间的相互转变,与通过R型从一种特异类型到另一种特异类型之间的转化,二者是完全不同的。类型间的转化现象从未发现自发产生,但用本文前面所概述的特殊技术进行实验,可以诱发产生。在这样的条件下,用来提取转化因素的S细胞的特异性类型,选择性地、专一地决定着一种化学上、免疫学上十分不同的荚膜多糖的酶促合成过程。
R之间的相互转变,与通过R型从一种特异类型到另一种特异类型之间的转化,二者是完全不同的。类型间的转化现象从未发现自发产生,但用本文前面所概述的特殊技术进行实验,可以诱发产生。在这样的条件下,用来提取转化因素的S细胞的特异性类型,选择性地、专一地决定着一种化学上、免疫学上十分不同的荚膜多糖的酶促合成过程。
在我们的研究过程中,曾观察到R36的原种培养物在血液肉汤中通过连续传代,它自发分离,因而产生了许多其他的R变种,这些变种可以根据其菌落类型彼此区分开来。这一现象的重要性在于:从亲本R培养物中分离出来的4个不同的变种,只有一个(R36A)对于有效提取液的转化作用表现敏感,而其他的都无反应,完全没有转化活性。不同的R变种对同一种特异性刺激具有不同的反应,这一事实强调指出:转化实验中所用的R变种,必须经过仔细选择。菌株R36A对各种不同转化因素的反应已搞清楚了,根据它对各种转化因素的不同反应,可以把它转化成Ⅰ型、Ⅲ型、Ⅵ型或 型,以及它原来的类型(Ⅱ型),前面已提到,这是从未自发回复过的。
型,以及它原来的类型(Ⅱ型),前面已提到,这是从未自发回复过的。
肺炎球菌细胞具有一种酶,它能破坏转化因素的活性,这一事实的重要性以后会明白,不过这里还要再提一下。的确,发现这种酶存在于许多不同菌株的自溶产物中,而且有很高的活性。Dawson和Sia[5 ] 发现,采用少量幼龄而旺盛生长的R细胞进行接种,对完成在试管中的转化实验起到很重要的作用。细胞在自溶过程中释放出胞内酶的事实,至少可以部分地解释这一发现。当接种量过大时,转化不易成功,或得出的结果没有规律性,这可能是由于自溶细胞释放出的酶量足以破坏这个反应系统的转化因素。
为了得出一致而又能重复的结果,必须记住以下两点:①R培养物能自发分离,并产生别的变种,这些变种已丧失了对转化刺激有反应的能力;②肺炎球菌细胞含有一种胞内酶,当它释放出来时,就会破坏转化因素的活性。因而,挑选敏感的菌株和尽可能防止与自溶有关的破坏性变化都是很重要的。
4.测定转化活性物质的方法
从肺炎球菌粗提取液中分离和纯化活性转化因素时,最好对各部分的转化活性有一个定量的测定方法。
所用的实验程序如下:用乙醇将待测活性材料灭菌,已知乙醇对转化活性无影响。把量好体积的提取液放在无菌的离心管中,加入4~5倍体积的无水乙醇,使其沉淀,把这种混合物在冰箱中静置8小时以上,能更有效地灭菌。把乙醇沉淀的物质离心,弃去上清液,把含有沉淀物的管子倒过来静置一会,除去乙醇。然后,把管口谨慎地经火焰灭菌,并用一个干的无菌棉塞堵住。把沉淀物重新溶于原体积的生理盐水中。活性物质用这种方法灭菌,效果稳定。采用这种方法不会使活性物质丢失,如果溶液经Berkefeld滤器过滤或在高温下加热灭菌,会引起待测活性物质的丢失。
在用炭吸附过的肉汤中,加入10%的无菌腹水液或胸膜液,这些体液都预先在60℃加热30分钟,以破坏能使转化因素失活的酶。把这种营养丰富的培养基在无菌条件下分装到15×100毫米的试管中,每管装2.0毫升。把灭过菌的提取液在pH7.2~7.6(用0.1克当量 【52】 NaOH调)的生理盐水中连续稀释,或者在0.025克分子 【53】 、pH7.4的磷酸缓冲液中同样稀释,每一稀释度取样0.2毫升,加到装有血清培养基的试管中,每个稀释度至少3管或4管。然后,把R36A的5~8小时血液肉汤培养物进行稀释,取其10-4 稀释液在每管中加0.05毫升,在37℃保温培养18~24小时。
在这种培养基中,血清的抗R特性引起R细胞在生长过程中发生凝集,而且凝集的细胞团离开上清液沉到管底。如果发生了转化,能形成荚膜的S细胞不受这些抗体的影响,在整个培养基中分散地生长。相反,在没有发生转化时,上清液仍然是清的,只有R型菌在底部生长。这种生长上的差异,使我们只要通过肉眼观察,就可以区分正负结果。按照常规,把所有培养物都涂布在血液琼脂平皿上,以便确证并进一步进行细菌学鉴定。由于本研究中所用的提取液是从肺炎球菌Ⅲ型分离得到的,因此,原来的R菌和转化了的S细胞菌落差异非常大,后者菌落大、闪光、呈黏液状,是典型的肺炎球菌Ⅲ型菌落。图1表示这两种菌落的形态差异。一种高度纯化制品的转化活性的典型数据列于表4。
图1 两种肺炎球菌的菌落
1.从肺炎球菌Ⅱ型分离到的R变种(R36A)的菌落。在不含转化因素的血清肉汤中培养的菌涂布在血液琼脂平皿上。(放大3.5倍);
2.R36A细胞在加入了肺炎球菌Ⅲ型转化因素的血清肉汤培养基中发生了转化之后,在血液琼脂平皿上形成的菌落。这种光滑、明亮、黏液状菌落是肺炎球菌Ⅲ型的典型特性,与图1所示亲代R菌株的小而粗糙的菌落是很容易区分的。(取自Joseph B. Haulenbeek拍摄的照片)
制备方法
1.材料
本研究中从肺炎球菌Ⅲ型的一株实验室原种菌株(A66)提取活性转化因素。将菌在50~75升牛心肉浸汤中,于37℃保温培养16~18小时。用一个蒸汽开动的、能够灭菌的Sharples离心机收集菌体。离心机装有浸在冰水中的冷凝管,因此培养液在流入机器之前已经充分冷却。这种程序可以抑制离心过程中的自溶作用。沉淀下来的细胞通过收集管取出,重新悬浮于大约150毫升冷的生理盐水中(0.85%NaCl),并仔细地把所有菌块都充分乳化。把盛有稠乳状的细胞悬液浸在水浴中,并将悬浮液的温度迅速升高到65℃。边加热边搅动,在65℃保温30分钟,把能破坏转化因素的胞内酶失活。
2.热杀死的细胞提取液
用过的提取方法很多,这里只介绍最好的一种。把加热杀死的细胞用生理盐水洗涤3次,洗涤过程的主要作用是除去大量过剩的荚膜多糖及一些蛋白质、核糖核酸和体细胞的C多糖。对转化活性的定量测定表明,在洗涤过程中,转化活性物质丢失不超过10%~15%,这与洗涤过程除去的非活性物质的量相比是很小的。
最末一次洗涤后,把细胞放在150毫升含有去氧胆酸钠(最终浓度为0.5%)的生理盐水中进行提取,提取时,把混合物机械振荡30~60分钟,通过离心把细胞分离出来,提取过程重复2~3次。用这种方法制备的去氧胆酸钠提取液是清亮无色的。合并提取液,并加入3~4倍体积的无水乙醇使提取液沉淀。因为去氧胆酸钠溶解在乙醇中,留在上清液里,通过这一步骤就把它除去了。沉淀形成丝状物,浮在乙醇界面上,用一个刮勺就可以直接把它挑出来。把沉淀物上剩余的乙醇倒去,然后再重新溶解在大约50毫升生理盐水中。得到的溶液通常是黏滞的,呈乳白色,是不透明的。
3.荚膜多糖的去除和脱蛋白的过程
按Sevag所叙述的氯仿法[12 ] 将溶液脱蛋白,这一步骤要重复2~3次,直到溶液变清为止。经过这一预备处理,然后再用3~4倍体积的乙醇将材料重新沉淀。得到的沉淀物溶解于150毫升的生理盐水中,再加入3~5毫克能水解肺炎球菌Ⅲ型荚膜多糖的、纯的细菌酶制品[13 ] 。将这种混合物在37℃下保温,然后用肺炎球菌Ⅲ型的抗体溶液来进行血清学试验,从而测定出荚膜多糖的破坏情况。抗体溶液是按Liu和Wu[14 ] 所介绍的方法,通过免疫沉淀物的解离制备出来的。采用抗体溶液的好处有以下两点:①它不与提取液中的其他活性物质起反应。②即使在1∶6000000的稀释液中,它也可以选择性地检查出荚膜多糖的存在。从丧失血清学活性的有关证据来看,酶解荚膜多糖通常在4~6小时完成。再用3~4倍体积的乙醇沉淀酶解物,得到的沉淀重新溶解于50毫升生理盐水中。氯仿法脱蛋白还用来除去加入的酶蛋白和残留的肺炎球菌蛋白。这一步骤要反复进行,直到在界面上不再出现蛋白质-氯仿凝胶薄膜为止。
4.乙醇分离
材料经脱蛋白和酶解荚膜多糖之后,再按下述方法用乙醇反复分离。一边搅动溶液,一边逐滴加入乙醇,在加入的乙醇接近临界浓度时(0.8~1.0倍体积),活性物质就呈纤维状细丝分离出来,并缠绕在搅棒上,然后再把它们从搅棒上冲洗到50%的乙醇和生理盐水混合液中。虽然大部分活性物质通过乙醇分离,在临界浓度时析出来了,但也有相当数量仍然留在溶液中。进一步将乙醇浓度增加到3倍,剩余的活性物质就与非活性物质一同呈絮状沉淀物沉降下来,把这些絮状沉淀物收集到5~10毫升生理盐水中,再用0.8~1.0倍体积的乙醇分离一次,又可得到一些丝状的活性物质,把它与第一次得到的活性物质合在一起。乙醇分离要重复4~5次。用这种方法可以从75升培养物中得到丝状活性物质10~15毫克,而且大部分是从最初的粗提取液中得到的。
5.温度的影响
除有特殊说明外,所有纯化步骤,常规操作都在室温下进行。但在理论上讲,制备生物活性物质在低温下进行更为有利。因此,有一批制品(制品44)的纯化工作是在低温下进行的。这种情况下,除了去氧胆酸钠提取和酶处理两步之外,其他所有步骤都是在保持0~4℃的房间中进行。在低温下制备的转化活性物质比在室温下同样制备的物质活性高得多。
用去氧胆酸钠提取加热杀死的细胞,如果在低温下进行,并不那么有效,而且得到的活性物质的量还要少些。对此现象曾有这样的解释:虽然就转化活性来说,最好是保持在低温下,但是高温使得活性转化因素的提取比较容易。
对提纯的转化物质的分析
1.一般性质
含纯转化物质为0.5~1.0毫克/毫升的生理盐水溶液,在漫射光下是清亮无色的,但在强透射光下不是完全无色的,搅动时,发出丝绸般的光泽。这种浓度的溶液是高度黏稠的。
提纯的物质溶于生理盐水中,在2~4℃下保存,至少3个月内活力不降低。但如果溶于蒸馏水中,其活力迅速降低,在几天之内就完全没有活性了。如果将生理盐水溶液在CO2 冰箱(-70℃)中冰冻保存,几个月中可以完全保持其活力。同理,用乙醇从生理盐水溶液中沉淀出来的活性物质,贮存在这种上清液中,其活力可以保持相当长的时期。纯化不完全的活性物质可以经过冷冻干燥保存在低压冻干器中。同样的程序用于保存高度纯化的材料时,发现材料会发生变化,引起溶解度降低,活性丧失。粗提取液中,转化因素的活性可以抵抗在65℃加热30~60分钟。高度纯化的活性物质不那么稳定,在此温度下,活性有所降低。高温加热对于纯化物质影响的定量研究现在尚未完成。Alloway[6 ] 用从肺炎球菌Ⅲ型细胞中提取出来的粗提取液进行实验,发现在90℃的水浴中加热10分钟后,偶尔还可以表现出活性。
因为氢离子浓度在酸性范围内引起转化物质活性逐步降低,因此,上述程序必须在中性溶液中进行,在pH5或更低时,失活迅速发生。
2.定性的化学检验
浓缩液中的提纯物质,对缩二脲和米隆(Millon)检验都是负结果。用干物质直接进行这些检验也是负结果。对脱氧核糖核酸的Dische二苯胺反应是强的正结果。对核糖核酸的苔黑酚检验(Bial)是弱的正结果。还发现:用不同方法从动物制备的DNA纯制品,在相同浓度时产生相同强度的Bial反应。
纯化物质中是否存在类脂,虽未进行专门的检验,但发现粗提取液可以经乙醇和乙醚在-12℃反复提取而不丧失活性。此外,正如在制备程序中指出的,用乙醇反复沉淀,和用氯仿反复处理,不引起生物活性的降低。
3.化学元素分析
分析了4种纯化制品的氮、磷、碳、氢的含量,结果列于表1。氮/磷比值在1.58~1.75,平均值为1.67,这个比值与根据DNA钠盐(四核苷酸)的理论结构推算出来的比值是一致的。单凭分析数字还不能确定分离出来的物质是纯的化合物。但是,根据氮/磷比值来看,似乎是蛋白质或其他含氮或磷的物质很少以杂质形式存在于这种制品中,否则这个比值会有较大的波动。
表1 转化物质纯化制品的化学元素分析
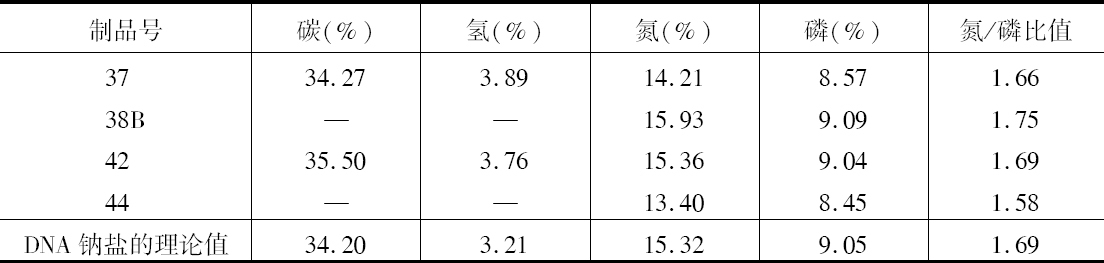
4.酶学分析
我们对各种粗酶和结晶酶破坏细菌提取液转化活性的能力进行了测定。在最适pH缓冲液中保存的提取液,加入结晶的胰蛋白酶、胰凝乳蛋白酶,或者两者都加入,经过这些酶处理后,活性不丧失。由于胃蛋白酶的作用需要在低pH下进行,但在低pH时提取液迅速失活,所以对胃蛋白酶没有测定。在最适条件下,用结晶核糖核酸酶长时间处理,并不引起转化活性明显降低。胰蛋白酶、胰凝乳蛋白酶及核糖核酸酶对转化因素没有影响,这些事实进一步证明这种物质不是RNA,也不是对胰蛋白酶类敏感的蛋白质。
除了各种结晶的酶之外,还测定了从各种动物组织得到的血清和酶类,以了解它们对转化活性的影响,发现有的能完全破坏转化因素的生物活性。测定过的各种酶制品,包括有用Martland和Robison[15 ] 的方法从兔子骨头,以及按H. Albers和E. Albers[16 ] 的方法从猪的肾脏中得到的高活性的磷酸酶。此外,还用了一种制品,这种制品是按Levene和Dillon[17 ] 的方法从狗的肠黏液中制备出来的,它含有胸腺核酸的一种多核苷酸酶。还测定了肺炎球菌的自溶物和商品的胰酶。这些制品的碱性磷酸酶活性是根据它们对β-磷酸甘油和磷酸苯酯的作用测定出来的,酯酶活性是根据它们裂解甘油三丁酸酯的能力测定出来的。因为发现从肺炎球菌提取液中分离出来的高度纯化过的转化物质含有DNA,因此也测定了这些酶对已知的一些DNA样品的解聚活性,这些DNA样品是Mirsky从鱼精子和哺乳动物组织中提取出来的。测定的结果列于表2。在表2中,对这些粗酶的磷酸酶、酯酶以及核酸解聚酶活性与它们破坏转化因素的能力进行了比较,分析这些结果可以看出,不管磷酸酶或酯酶的存在与否,发现只有一类制品能使转化因素失活,这类制品都含有一种能解聚DNA的酶。
表2 酶的粗制品对转化因素的失活作用

Greenstein和Genrette[18 ] 曾经指出,组织提取液以及几种哺乳动物的奶和血清,含有一种能使DNA解聚的酶。后来Greenstein把这种酶叫做DNA解聚酶[19 ] 。这些研究者们用降低DNA钠盐溶液黏度的方法测定了解聚酶的活性。把核酸盐与酶在黏度计中混合,在30℃保温,在不同时间间隔测定黏度。本实验中,把这种方法用于测定多聚酶活性,只是保温改在37℃下进行。除了用黏度降低法外,还根据核酸盐在酶解过程中酸沉淀能力逐步降低的特性,进一步测定了酶的作用。
新鲜的、正常的狗和兔子的血清对于转化物质活性的影响,是通过下面的实验说明的。
从正常的狗和正常的兔子得到的血清,用等体积的生理盐水稀释,把稀释过的血清分成相等的三部分:一部分在65℃加热30分钟;另一部分在60℃加热30分钟;第三部分不加热作为对照。一种提纯不完全的转化物质,预先在低压冻干器中经过干燥,然后在生理盐水中稀释,使浓度为3.7毫克/毫升。取这种溶液1.0毫升与加热过和没加热的各种血清混合,调混合物pH为7.4,然后在37℃保温2小时。血清和转化物质作用之后,把所有试管都在65℃加热30分钟,终止酶反应。然后用生理盐水进行连续稀释,按照测定方法中所叙述的步骤,测定三部分的转化活性。表3给出的结果表示能使转化因素失活的狗血清和兔子血清中的各种酶的不同热失活情况。
从表3的资料可以看出,未经热处理的狗血清和兔血清都能完全破坏转化活性,狗血清在60℃或65℃,加热30分钟后,不能再使转化活性丧失。这样看来,在狗血清中,能破坏转化因素的酶在60℃就完全失活了。而兔血清中能破坏转化因素的酶,则需要在65℃,加热30分钟才能完全失活。
表3 能使转化物质失活的狗血清和兔血清中的酶的不同热失活情况
①血清和转化物质的水解混合液的稀释度。
在上述实验中所用的狗血清和兔血清,我们还测了它们对于Mirshy从鱼的精子中分离出来的DNA钠盐制品的解聚酶活性。
将核酸钠盐的高黏性溶液在蒸馏水中调其浓度为1毫克/毫升备用。把热处理过的和未经热处理的血清,按前面所说的方法在生理盐水中稀释,然后取1毫升稀释好的血清溶液和4毫升核酸钠盐的水溶液在Ostwald黏度计中混合,并立即测定其黏度,然后在37℃保温24小时,在这段时间的不同间隔,分别进行黏度测定。
这个实验的结果在图2中表示出来。狗和兔子未经热处理的血清,在5~7小时,黏度都降低到和水的黏度一样。经60℃、热处理30分钟的狗血清,在22小时后其黏度没有明显降低。相反,经60℃热处理的兔血清只是降低了解聚酶的作用速率,24小时后,其黏度和未加热的血清降到相同水平。65℃热处理,完全破坏了兔血清解聚酶。
图2 狗和兔血清的DNA解聚酶的不同热失活情况
从狗和兔子血清的情况来看,解聚酶和能破坏转化因素的酶,失活温度之间有着明显的平行关系。不同的酶失活的温度不同,这是血清中所有酶的普遍性质。用上述同种血清样品中的甘油三丁酸酯酶所做的热失活实验,进一步证实了这个事实。后面这个实验中的结果与用多聚酶所观察到的结果是相反的,兔血清的酯酶在60℃几乎完全失活,而狗血清的酯酶在这个温度下受到的影响很小。
我们曾对很多物质抑制能够破坏转化因素的酶的能力进行了测定,其中只有氟化钠具有明显抑制效果。从肺炎球菌细胞、狗肠黏液、胰酶、或正常血清中得到的能破坏转化因素的酶,其活力都受到氟化物的抑制。与此相似,还发现相同浓度的氟化物也能抑制DNA的酶促解聚作用。
我们认为转化活性因素是脱氧核糖类型的核酸,证据是:①只有含DNA解聚酶的那些制品才能破坏转化活性;②在这两个例子中,与转化活性有关的酶 【54】 都是在相同的温度下被失活的,而且都受到氟化物的抑制。
5.血清学分析
发现在转化活性物质的化学分离过程中,当粗提取液被提纯后,肺炎球菌Ⅲ型的血清学活性逐渐降低了,而相应的生物学活性并不丧失。用高效价的Ⅲ型抗肺炎球菌兔血清对高度纯化过的转化物质溶液进行沉淀检验时,它本身只产生微弱的痕迹量的反应。已知,肺炎球菌的蛋白质即使稀释到1∶50000,也可以用血清学方法检查出来,荚膜和体细胞多糖即使稀释到1∶5000000以上,也可以用血清学方法检查出来。从这些事实看来,血清学活性的丧失说明在最后的制品中,已把上述的各种细胞组分 【55】 完全除去了。纯化状态的转化物质,血清学活性极少,或根本没有血清学活性,但它诱发肺炎球菌转化的生物学特异性与此是完全相反的。
6.物理化学研究
我们用分析超速离心机测定了一种纯化的、有活性的转化物质(制品44)。这种物质只产生一条单一的、清楚的带,这说明它是均一的、分子大小一致、并且非常不对称。发现生物学活性和光学上观察到的带以相同的速率下降,这个事实说明转化物质的生物学活性与其大小有关。只有测定出了扩散常数和某些特殊的体积,才能准确地测定出其分子量。Tennent和Vilbrandt[20 ] 已经测定了胸腺核酸的几种制品的扩散常数,其沉降速率与本研究中所观察到的数值一致。假定在这两种情况下分子的不对称性相同,那么可以估计出肺炎球菌转化物质的分子量在500000数量级。
制品44在Tiselius电泳仪上电泳,只出现一种电泳成分,其泳动力很高,与核酸的泳动力一致。转化活性与泳动快、而且形成清晰带的那种组分有关。这样看来,在电泳和离心两个方面,这种纯化物质的行为都与下述概念一致,即这种生物学活性是高度聚合的核酸的一种特性。
这种物质的紫外吸收曲线,最大值在2600埃(1埃=0.1纳米),最小值在2350埃,这正是核酸的典型特性。
7.生物学活性的定量测定
发现分离出来的这种物质,在其高度纯化状态时,能引起转化的数量范围是0.02到0.003微克。制品44是在低温下提纯的,其氮磷比值为1.58,它有很高的转化活性。这种制品的活性滴度在表4中给出。
表4 制品44的转化活性
①稀释前,原液每毫升中含有纯化物质0.5毫克,每个稀释度取0.2毫升溶液加到含有2毫升标准血清肉汤的试管中,共4个试管。取R36A血液肉汤培养物的10-4 稀释液0.05毫升加到各试管中。
把每毫升含纯化物质0.5毫克的溶液按规定作一系列的稀释。每种稀释液取0.2毫升加到4个试管中,每个试管中都含有2毫升标准血清肉汤。然后将R36A的5~8小时血液肉汤培养物稀释到10-4 ,再取0.05毫升接种到各试管中。按照滴度测定方法中所叙述过的程序测定转化活性。
表4给出的资料说明,活性物质引起转化的最低干重为0.003微克。由于在这个反应系统中,是在2.25毫升中含0.003微克,这表明纯化物质的最终浓度是1/6×108 。
讨论
本文对引起肺炎球菌特异性转化的物质之化学特性进行了讨论。从肺炎球菌Ⅲ型中分离出来的DNA,能够把从肺炎球菌Ⅱ型分离出来的不形成荚膜的R变种,转化成为完全形成荚膜的肺炎球菌Ⅲ型细胞。Thompson和Dubos[21 ] 已从肺炎球菌中分离出了一种核糖类型的核酸。就作者所知,在这以前从肺炎球菌中没有发现脱氧核糖类型的核酸。用一种已知的化学物质在离体实验中诱导成功特异性转化,以前也没有见到过。
虽然这些发现只局限在一个样品中,但前人的工作已经阐明了各种肺炎球菌类型的相互变化,并指出,每种情况下,引起变化的特异性是由引起这种变化并能形成荚膜的细胞的特殊类型决定的。从普遍现象的观点来看,所研究过的高度纯化、不含蛋白质而含有大量DNA的这种样品,它能促使肺炎球菌Ⅱ型不形成荚膜的变种产生荚膜多糖,所产生的荚膜多糖与分离这种诱导物质的细胞的荚膜多糖在类型特异性上是一致的,这一点具有重要的意义。另外,很明显引起这个反应的物质与随之而产生的荚膜物质在化学上是不同的,分别属于完全不同的两类化合物。
这种诱导物质,根据其化学和物理特性来看,可能是一种高度聚合、而且很黏滞的DNA钠盐。由它引起合成的Ⅲ型荚膜物质,主要是由不含氮的多糖组成的,这种多糖是由葡萄糖和葡萄糖醛酸通过配糖键连接而成的[22 ] 。新形成的这种荚膜所含的多糖具有类型特异性,因此使转化了的细胞具有肺炎球菌Ⅲ型的所有典型特性。由此看来,这种诱导物质和由它产生的物质在化学上虽然是不同的,它们的作用各有其生物学特异性,但对于决定所形成的细胞的特异性类型都是必需的,它们都是这种新形成的细胞的一部分。
本文所列举的实验资料有力地说明,转化因素的选择性作用,证实了核酸(至少是脱氧核糖类型的核酸)携带着不同的特异性。以前曾有人提出过[23 ,24 ] 核酸的生物学行为可能存在着特异性差别,但一直没有被实验证实,至少其中一部分原因是由于缺少合适的生物学方法。转化研究中所用的这种技术,可能为检验这种假说的确实性提供一种敏感的方法,而且到现在为止,所得到的结果都支持这种观点。
前面所说的这种物质的转化活性,实际上是核酸的一种遗传特性,这已经完全被证实了,但人们还必须进一步阐明其作用具有生物学特异性的化学基础。乍一看,免疫学方法可能为测定这类生物学上有重要意义的物质不同特异性,提供理想的方法。虽然核酸分子的组成单位和一般构型已经明确,但分子构型的细微差别对这些物质的生物学特异性会起到什么影响,至今所知甚少。由于纯的核酸以及与组蛋白或精蛋白结合在一起的核酸,都没有抗原性功能,因此我们预料用免疫学技术不能揭示出核酸分子构型的这些细微差别。不含蛋白质的高度纯化DNA,虽然诱导转化很有效,但用抗肺炎球菌Ⅲ型的兔血清进行沉淀素检验时,仅有微弱的、痕迹量反应,这是不足为奇的。
根据这些有限的观察,就对核酸的免疫学重要性作出任何结论都是不明智的,除非将来对这个问题有了进一步了解。Lackman及其同事[25 ] 的发现说明,从溶血链球菌和动物、植物中得到的酵母型和胸腺型的核酸,都能与某些抗肺炎球菌的血清产生沉淀。反应随不同批号的免疫血清而有所变化,用抗肺炎球菌的马血清比用相应的免疫兔血清出现的变化更多。遇到的这些不规律性和广泛的交叉反应使研究者们对于这些结果的免疫学意义产生了怀疑。目前,转化技术是测定核酸生物学行为差异的唯一有效方法(除非将来设计出更特异的免疫学方法来,能像证明简单的非抗原物质的血清学特异性所用的方法那样有效)。
应该承认,转化问题还有许多方面需要进一步研究,还有许多问题仍然不清楚,这主要是因为技术上的困难造成的,例如,弄清转化物质的浓度和反应速率的相关性是很有意义的。反应系统中,转化了的细胞与未转化细胞的相对比例,从细菌学观点来看,根据菌落进行计数比用光电比色计数更不准确。因为R细胞能被培养基中的抗血清凝聚产生集结和沉淀。把细菌放在不能生长和繁殖的条件下保存一段时间,然后用这种静止的细胞悬液进行转化,都未获得成功。由此看来,很可能只有当细胞处在旺盛繁殖时期才能发生转化。这方面有一个重要的事实是肺炎球菌的R细胞、转化了的细胞、可能还有所有其他变种及各种类型的肺炎球菌都含有一种胞内酶,它在细胞自溶过程中释放出来,处于游离状态,它能迅速而完全地破坏转化因素的活性。因此,很可能在细胞处于分裂最活跃、自溶最小的对数生长期时,培养条件对于维持R细胞的最大反应能力和因自溶酶释放而引起转化因素的最小失活之间的平衡关系是最合适的。
就目前的知识水平来说,对转化机制的任何一种解释都只能是纯理论的。在转化过程中发生的生化事件说明,转化因素与R细胞相互作用,产生一系列的酶反应,这些反应的结果合成了Ⅲ型荚膜抗原。这些实验的结果清楚地说明,引起的变化不是随机的,而是可以预料的,总是与提取转化因素的那些能形成荚膜的细胞的类型特异性一致。一旦发生了转化,以后在人工培养基上不需要再加转化因素,新获得的性状就可以无数代的传下去。在转化了的细胞中,发现一种物质与转化因素具有相同的活性,但数量比原来为引起这种变化所加入的量大得多。因此,很明显,转化了的细胞在连续的繁殖过程中,不仅产生了荚膜物质,而且控制着荚膜形成和荚膜特殊性的基本因素也在子代细胞中复制了。产生的这些变化不是暂时的,而是永久性的,倘若培养条件有利于保持荚膜形成能力,那么这些变化便能一直持续下去。转化了的细胞,不仅可以通过血清学反应,还可以根据它具有明显的新形成的荚膜,从而很容易地与亲代R型细胞区分开来。荚膜是肺炎球菌类型特异性的免疫学单位,也是决定这种细菌能够感染动物所必需的附属结构。
实验诱发的变化与形成一种新的形态结构以及随后获得新的抗原和侵染特性是密切相关的,就肺炎球菌的情况来说,这是非常重要的。这些变化是可以预料到的、具有类型特异性、又是可以遗传的,这些事实也是比较重要的。
已经提出了几种假说来解释引起的这些变化的性质。格里菲斯[1 ] 在对这种现象的最初叙述中提到,接种物中的死细菌可能提供了某些特异性蛋白质作为食料,使R型细胞能制造荚膜碳水化合物。
最近从遗传学的观点对这种现象进行了解释[26 ,27 ] ,这种诱导物质被比喻为一个基因,随着它而产生的这种荚膜抗原被认为是一种基因产物。在讨论转化现象时,Dobzhansky[27 ] 曾说:“如果把这种转化现象说成是一种遗传突变——回避这种说法也很困难——我们现在所讨论的正是通过特殊处理引起特殊突变的真实事例……。”
Stanley对这种现象提出了另一种解释[28 ] ,他曾经详述过转化因素的活性和病毒的活性之间的相似性。另一方面,Murphy[29 ] 曾对引起鸡肿瘤的因素和肺炎球菌的转化因素进行过比较,他建议称这两类因素为感染性诱变因素,以区别于病毒。不管你怎样证明解释的正确性,但观点上的这些不同足以说明了相对于遗传学、病毒学和癌的研究领域中同类问题来讲,对转化现象的了解还是不清楚的。
当然也有可能,前面谈到的这种物质的生物学活性并不是核酸的一种遗传特性,而是由于某些微量的其他物质所造成的,这些微量物质或者是吸附在它上面,或者与它密切结合在一起,因此检查不出来。但是分离出来的这种生物学活性物质,经过高度纯化就以DNA钠盐形式存在,有可靠的证据充分说明它实际上就是转化因素。因此,这种类型的核酸,不仅在结构上非常重要,而在决定肺炎球菌细胞的特异性和生化活性上也是很重要的。如果DNA钠盐和活性转化因素的确是同种物质,那么我们可以说,转化现象是通过一种已知的化合物直接地、专一性地诱发出来的变化。如果本文有关转化因素化学性质的研究结果被进一步证实,那么核酸必定还具有化学基础上目前尚不清楚的生物学特异性。
摘要
(1)从肺炎球菌Ⅲ型分离出一种具有生物学活性的成分,这种成分在其高度纯化形式时,只要极少数量,在适宜培养条件下,就可以引起肺炎球菌Ⅱ类不形成荚膜的R变种,转化成能形成荚膜的细胞,这些能形成荚膜的细胞与发现这种转化物质的、经热处理杀死的细菌具有相同的类型特异性。
(2)叙述了分离和提纯活性转化物质的方法。
(3)从化学、酶学和血清学分析得出的结果,及以前电泳、超离心、紫外线光谱学研究结果可以说明,在现有测定方法范围内,活性物质不含蛋白质、不含类脂、也不含具有血清学反应的多糖,而主要是(即便不是唯一的)由高度聚合的、黏滞的DNA组成的。
(4)列举了一些证据,说明化学诱发的细胞结构和功能上的变化是可以预料的,是具有类型特异性的,而且是可以遗传的。评论了这些变化的性质的各种假说。
结论
提出的证据支持了认为脱氧核糖类型的核酸是肺炎球菌Ⅲ型转化因素的基本单位这种看法。
参考文献
[1 ]Griffith, F., 1928. J. Hyg., Cambridge, Eng., 27 , 113.
[2 ]Neufeld, F., and Levinthal. W., 1928. Z. Immunitätsforsch., 55 , 324.
[3 ]Baurhenn, W., 1932. Centr. Bakt., 1. Abl., Orig., 126 , 68.
[4 ]Dawson, M. H., 1930. J. Exp. Med., 51 , 123.
[5 ]Dawson, M. H., and Sia, 1931. R. H. P., J. Exp. Med., 54 , 681.
[6 ]Alloway, J. L., 1932. J. Exp. Med., 55 , 91; 1933 57 , 265.
[7 ]Berry, G. P., and Dedrick, H. M., 1936. J. Bact., 31 , 50.
[8 ]Berry, G. P., 1937. Arch. Path., 24 , 533.
[9 ]Hurst. E. W., 1937. Brit. J. Exp. Path., 18 , 23; Hoffstadt, R. E., and Pilcher. K. S., 1941 J. Infect. Dis., 68 , 67; Gardner, R. E., and Hyde, R. R., 1942. J. Infect. Dis., 71 , 47; Houlihan, R. B., 1942. Proc. Soc. Exp. Biol. and Med., 51 . 259.
[10 ]MacLeod, C. M., and Mirick, G. S., 1942. J. Bact., 44 , 277.
[11 ]Dawson. M. H., 1928. J. Exp. Med., 47 , 577; 1930. 51 , 99.
[12 ]Sevag, M. G., 1934. Biochem. Z. 273 , 419; Sevag, M. G., Lackman, D. B., and Smolens, J., 1938. J. Biol. Chem., 124 , 425.
[13 ]Dubos. R. J., and Avery, O. T., 1931. J. Exp. Med., 54 , 51; Dubos. R. J. and Bauer, J. H., 1935. J. Exp. Med.,62 , 271.
[14 ]Liu. S., and Wu, H., 1938. Chinese J. Physiol., 13 . 449.
[15 ]Martland, M., and Robison, R., 1929. Biochem. J., 23 , 237.
[16 ]Albers, H., and Albers, E., Z. 1935. physiol. Chem., 232 , 189.
[17 ]Levene, P. A., and Dillon, R. T., 1937. J. Biol. Chem., 96 , 461.
[18 ]Greenstein, J. P., and Jenrette, W. Y., 1940. J. Nat. Cancer Inst., 1 , 845.
[19 ]Greenstein, J. P., 1943. J. Nat. Cancer Inst., 4 , 55.
[20 ]Tennent, H. G., and Vilbrandt, C. F., 1943. J. Am. Chem. Soc., 65 , 424.
[21 ]Thompson, R. H. S., and Dubos, R. J., 1938. J. Biol. Chem, 125 , 65.
[22 ]Reeves, R. E. and Goebel, W. F., 1941. J. Biol. Chem., 139 , 511.
[23 ]Schultz, J., 1941. In Genes and chromosomes. Structure and organization, Cold Spring Harbor symp. on quant. biol. Cold Spring Harbor, Long Island Biological Association, 9 , 55.
[24 ]Mirsky, A. E., 1943. In Advances in enzymology and related subjects of biochemistry, (F. F. Nord, and C. H. Werkman, editors), New York, Interscience publishers, Inc., 3 , 1.
[25 ]Lackman, D., Mudd, S., Sevag, M. G., Smolens, J., and Wiener, M., 1941. J. Immunol., 40 . 1.
[26 ]Gortner, R. A., 1938. Outlines of biochemistry, New York, Wiley, 2nd edition, 547.
[27 ]Dobzhansky, T., 1941. Genetics and the origin of the species, New York Columbia University Press, 47.
[28 ]Stanley, W. M., 1938. in Doerr, R., and Hallauer, C., Handbuch der Virusforschung, Vienna, Julius Springer, 1 , 491.
[29 ]Murphy, J. B., 1931. Tr. Assn. Am. Physn., 46 , 182; 1935. Bull. Johns Hopkins Hosp., 56 , 1.
在细菌生物化学突变型混合培养物中出现的新基因型 【56】 【57】
乔舒亚·莱德伯格(J. Lederberg) 【58】 E. L. 塔图姆(E. L. Tatum)
(美国 冷泉港实验室,1946年)
Hershey曾报道过[1 ] ,在一种细菌病毒中出现了遗传性状的新组合。从一些简单的实验叙述中可以看出,在大肠杆菌中有一种情况与上述报道在某些方面是相似的。
Tatum[6 ] 在评述大肠杆菌生化突变时,曾谈到过这些生化突变特性为遗传分析提供的方便。尤其是人们通过把冲洗过的细胞浓悬液涂布在只有原养型才能形成肉眼可见菌落的基本琼脂培养基上,很容易把这些特性检查出来,从而认识到了这种方法的简便和确切性。有一种假说认为,原养型是由于两个或多个座位(locus)的回复突变巧合地出现在同一个细胞之中所产生的结果。但是在生化突变型混合培养物中原养型出现的频率,事实上比根据这种假说预计的频率要大得多。而且在相同条件下培养和鉴定出来的相同的多重突变型,发现它们的培养物彼此是相同的,没有相对独立的特性。由于大肠杆菌许多生化突变型以大约10-7 数量级的频率产生回复突变[3 ,5 ] 。因此,我们采用了多重突变型,这些多重突变型是参照以前的介绍用多次突变的方法获得的[6 ] 。
把几种多重突变型的混合培养物在完全培养基(酵母膏-蛋白胨-葡萄糖)中进行培养,曾多次发现有相当数量的原养型[4 ] (或者叫营养野生型)细胞出现。但到现在为止,发现原养型出现的频率都低于10-7 ,当然也发现了少数细胞在单一座位上发生了回复突变。
本实验室已经证实,不同的生化突变型,通过在培养基中的交换作用,可以彼此提供生长所需因素。由此看来,这些原养型很可能是不同突变型的异源集合体。我们试图通过生物学和物理学方法来检查或诱导这种想象集合体组分的解离,但正如下面指出的那样,都未获得成功。因此,很可能这些原养型在基因型上是一致的。
通过单菌落分离可以看出,这些原养型都是十分稳定的,从在完全培养基上长成的培养物中分离出来的几百个菌落,经证明都是原养型的。
用适当剂量的紫外线照射生长在完全培养基中的原养型培养物,使之涂布在完全琼脂培养基上后,出现的菌落数降低到1∶105 。很显然,在这样的致死率下,所假定的那种集合体大多数都不能存活。未杀死的细菌中,在大多数情况下,只有单细胞存活者才有可能存活下来形成菌落。检查过的几百个菌落也都是原养型的。
幸好,用来进行这些实验的菌株(K-12)对于细菌病毒T1[2 ] 是敏感的。在下面的实验中,所用的多重突变型分别需要生物素和蛋氨酸(B- M- P+ T+ )、苏氨酸和脯氨酸(B+ M+ P- T- )。此外,以低频率发生的抗(R)T1的自发突变是按Luria和Delbrück[2 ] 叙述的程序挑选出来的。
把混合培养物按前面谈到的方法涂布在培养基上,分离出原养型来,并测定它对病毒的抗性。研究了各敏感菌株的混合物,发现都是敏感的原养型。同样,检验各抗性菌株混合物时,得到的原养型全是抗性的。但当用(B- M- P+ T+ R)和(B+ M+ P- T- )的混合培养物时,分离出来的10株原养型中,有8株是抗性的、有2株是敏感的。当用(B- M- P+ T+ )和(B+ M+ P- T- R)的混合物时,分离出来的10株原养型中有3株是抗性的、有7株是敏感的。如果这些原养型是由原来各突变型的集合体组成的,那么,在这些情况中,每种原养型都应该有大部分细胞是抗病毒的。我们以前曾提到过发现9种原养型培养物都能被T1完全溶菌。正如人们预料的那样,显性性状可以是抗性,但也可以是敏感性;所以,抗性原养型和敏感原养型都能出现,这也证明了原养型内部的同质性(见表1)。
表1 从单一培养物和混合培养物中分离出来的各种基因型
(在虚线上的字母表示实验中所用的突变型。)
①原养型。②生化变异见A。
R抗病毒T1。
下面各字母分别表示各种必需代谢物:
B 生物素; M 蛋氨酸;  苯丙氨酸; P 脯氨酸; C 胱氨酸; T 苏氨酸。
苯丙氨酸; P 脯氨酸; C 胱氨酸; T 苏氨酸。
其他突变型的组合也形成了原养型。尤其是把脯氨酸-苏氨酸缺陷型(B+ + C+ P- T- )和生物素-苯丙氨酸-胱氨酸缺陷型(B- - C- P+ T+ )的混合物,涂布到含有生物素、苯丙氨酸和脯氨酸的琼脂平皿上,除了分离出了原养型外,还分离到了一些单一回复突变型,例如,生物素缺陷型(B- + C+ P+ T+ )和生物素-脯氨酸缺陷型(B- + C+ P- T+ )。
参考文献
[1 ]Hershey. A. D., 1946. Spontaneous mutations in bacterial viruses. Cold Spring Harbor Symp. Quant. Biol.11 : 67~77.
[2 ]Luria, S. E. and Delbrück, M., 1943. Mutations of bacteria from virus sensitivity to virus resistance. Genetics 28 : 491~511.
[3 ]Ryan, F. J. 1946. Back-mutation and adaptation of nutritional mutants. Cold Spring Harbor Symp. Quant. Biol. 11 : 215~227.
[4 ]Ryan, F. J. and Lederberg, J. 1946. Reverse-mutation and adaptation in leucineless Neurospora. proc. nat. Acad. Scl. 32 : 163~173.
[5 ]Ryan, F. J. and Lederberg, J. Unpublished experiments.
[6 ]Tatum, E. L. 1946. Induced biochemical mutations in bacteria. Cold Spring Harbor Symp. Quant. Biol. 11 : 278~284.
沙门氏菌的遗传交换 【59】
诺顿·津德尔(N. D.Zinder) 【60】 乔舒亚·莱德伯格(J. Lederberg)
(1952年)
许多不同细菌的遗传学研究揭示出它同更高级的生物有共同之处,也有不同之处。把选择性富集培养技术成功地应用于大肠杆菌基因重组的研究(Tatum和Lederberg,1947;Lederberg等,1951),使人设想到应当对别的细菌使用一种相似的手段。本文介绍在鼠伤寒沙门氏菌和其他沙门氏菌血清型方面所进行的这类实验的结果。这些实验中发现的遗传交换机理在许多方面同大肠杆菌的有性重组是不同的,从而有理由提出一个新的说明名词,即转导。
材料和方法
Lilleengen(1948)提供了鼠伤寒沙门氏菌的大多数菌株,这些菌株代表了他的21个噬菌体类型,从LT-1到LT-22鼠伤寒沙门氏菌的大多数菌株(即使不是全体)是溶源的(Boyd,1950),它们有12个细菌噬菌体品系。从F. Kauffmann、E. K.Borman和P. R.Edwards得到其他培养物。全部培养物都保存在斜面肉汤琼脂上。
把紫外线照射过的细胞悬浮液,用青霉素法处理作选择性分离,得到了对特异生长因子依赖的突变体(营养缺陷型)(Davis,1950a;Lederberg和Zinder,1948)。Plough等(1951)和Bacon等(1951)在沙门氏菌得到了相似的突变体。对营养缺陷和发酵突变体的分离和说明其特性的其他方法,在别的文章中已有介绍(Lederberg,1950;Lederberg和Lederberg,1952)。通过把稠密的未照射的细胞悬浮液接种于每升含500毫克双羟链霉素的琼脂平皿上选出抗链霉素的突变体。
用与大肠杆菌相同的公式(Lederberg,1950),制成完全的指示培养基(EMB)。规定的伊红美蓝培养基(EML琼脂)在每升中含有:乳酸钠2.5克、(NH4 )2 SO4 5克、NaCl 1克、MgSO4 1克、K2 HPO4 2克、美蓝盐酸0.05克、Y伊红0.3克和琼脂15克。采用Difco产物,Penassay肉汤和肉汤琼脂为完全培养基。
除另有说明外,全部培养物都在37℃下培养,经24和48小时后记载平皿培养情况。
试验结果
直接杂交:混合培养物的平皿培养
在大肠杆菌中,把各营养缺陷型一起涂布在最低琼脂培养基上,有选择性地检出其重组体。两亲本在这个培养基上都受到抑制,并除去各种实验误差外,只有原养型重组体细胞形成菌落。用适宜的对照能查出这类误差,但使用双营养突变体(二营养缺陷型)可使这类误差大大减少。这些双营养突变体是在过去确立的营养缺陷型系中再分离突变体得到的。
我们的突变体分离的技术对Lilleengen菌株中有一个菌株不适用。剩下的20种菌株,每一个都准备了营养要求上彼此互补的两步突变体。在包括自身杂交在内的200个可能配对的组合中,测验了100个组合。通过混合并在一最低琼脂平皿培养基上涂布两个亲本109 洗涤过的细胞,对每一个组合进行了研究。每次试验每个亲本接种15个混合平皿培养和5个对照平皿培养。同对照相反,15个组合产生原养型。特别与LA-2相比较,LA-22菌株是最为多产的(见表1)。这个杂交每接种10万个细胞产生的一个原养型细胞。LA-22以外的杂交产生原养型少得可怜,从而使它显得毫无意义。以后弄清楚,LA-22虽然来自两步突变并具有复杂的营养,但它在遗传上是一个单独的稳定突变体。
表1 突变体菌株及所用符号

LT-22对一个作用于LT-2的病毒(以后称之为PLT-22)来说,是溶源性的。这个病毒能诱导LT-2的溶源性。在LA-2的溶源衍生物中,发现有3组相互作用各不相同:大多数不再同LA-22相互作用产生原养型;少数以不配对的效应起相互作用;更少数在这方面毫无影响。这些试验表明,的确发生着遗传交换,而潜在的细菌噬菌体在这个相互作用中起到了一定的作用。
间接杂交:细胞和滤液的涂布
为测试滤过性因素在这种相互作用中的可能作用,按照Davis(1950年)的设计制备了一种具有一“极细”的熔结Pyrex滤器分配的U形管。通过管臂之间交替抽气,可以在同一个培养基上得到两个完整的正在生长的细菌群体。留下一部分不接种作对照试验以验证滤器的完整性。然后取每个亲本108 细胞接种到20毫升的肉汤中,再分别置于管的两臂。当培养物生长到饱和时,4小时内每20分钟用10毫升肉汤从这一边到那一边进行冲洗。漂洗出两个群体并把它涂布在最低培养基上。LA-22平皿上出现原养型细胞,但LA-2平皿上则不出现原养型细胞。LA-2肉汤培养物的消毒滤液不能从LA-22诱导出原养型细胞。但LA-2和LA-22混合培养的滤液,大约每一百万个LA-22细胞诱导出一个原养型细胞。这样,在LA-22的刺激作用下,LA-2产生了一种滤过物(以后简称为FA),它能从LA-22诱导出原养型细胞。含有相当量对LA-2有作用的噬菌体(PLT-22)的LA-22培养物滤液,也刺激LA-2产生FA。这种噬菌体的作用将在后面讨论。
为有助于进一步说明我们的测验,我们将使用转导一词作为遗传物质单方面的转移,以有别于受精中相等成分的联合。沙门氏杆菌FA是一种遗传转导物这一工作假说,对我们的讨论,提供了一个可供参考的有用基础。
FA的测定
使LA-22和LA-2在肉汤中混合培养生长制备成FA原汁。48小时后,沉淀细胞,并使上清液通过一熔结的Pyrex滤器。通过在制备时把样品接种到肉汤中并涂布在琼脂中作为特定试验的对照以验证滤器的无菌。要采取这种预防措施,虽然对大多数试验来说,由于在间接杂交中同LA-22相互作用产生原养型细胞需要每平皿一百万个以上的LA-2细胞,因而完全的无菌并不是苛求的。这些制备物可贮藏在冰箱中达数月之久而不丧失活性。
为了进一步工作,发展了一种测定FA的标准方法。LA-22生长在肉汤琼脂平皿上,并以浓缩的含盐悬浮液收集。把这种悬浮液适量稀释,再涂布在肉汤琼脂上,计算成活细胞。取细胞的各种稀释液,分别加上体积恒定的FA制备物涂布于基本培养基上。24小时出现原养型细胞,48小时后计数。图1表示用每平皿大约109 ~1010 个细胞所发现的稳定的反应。在高细胞浓度处的下降可能由于过于拥挤而抑制菌落的形成,低浓度处的下降则可能由于细胞和滤过物的物理性散开或饱含敏感细胞所致。
用LA-22 109 细胞同各组FA稀释液进行平皿培养,发现原养型细胞的产量和FA的量之间有着明显的直线关系(见图1)。高浓度FA的效应将在下一节讨论。
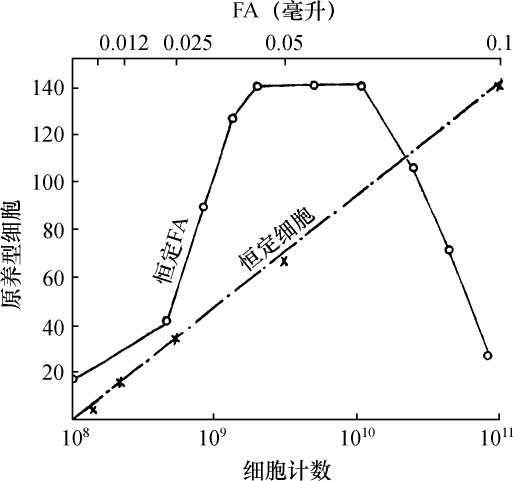
图1 FA的测定
以各种稀释度混合FA(LT-2)和细胞(LA-22)并涂布于基本培养基平皿上。48小时后计数原养型细胞。
FA的一个单位可规定为滤液从最适浓度的LA-22细胞中诱导出单个原养型细胞的含量。混合细胞制备物的滤液通常是每毫升含有约2500个这种单位。
FA的化学反应
随着发展出一种标准的测定法,就可能比较各种处理对FA和细菌细胞的效应。前者加上氯仿、甲苯、乙醇和福尔马林这类试剂,予以震荡消毒。其中只有福尔马林能使FA失活。细菌消毒是加热到56℃,30分钟。为探查出FA的效应,需要70℃的温度。只有到达100℃,它才迅速失活。
用1到2份体积的冷乙醇或半饱和的硫铵可从肉汤中定量沉淀FA。在这两种情况下都出现累累絮凝,其大部分仍然是非水溶性的;但FA重新分散。
供试的一些酶对FA都没有什么影响。把酶直接加入活性滤液并保温2小时。试验包括胰酶(100毫克/毫升)、胰蛋白酶(100微克/毫升)、Taka淀粉酶制剂(100毫克/毫升)、核糖核酸酶(10微克/毫升)和脱氧核糖核酸酶(20微克/毫升)。脱氧核糖核酸酶之不能使FA失活特别有意思。通过测试这种反应混合液的样品在降低胸腺核酸(由R. D.Hotchkiss博士慷慨提供)的黏度,来验证酶的活性。其他的酶不作相似的对照。
FA的诱发作用
Burnet和McKie(1929)以及Boyd(1951)总结了沙门氏杆菌的潜伏噬菌体的特点。溶源噬菌体,即从溶源细菌得到的噬菌体,对敏感培养物的溶菌能力很差,并易于诱发第二种抗溶源类型。仅在少量侵染时才看到敏感细胞可见的溶解。当把噬菌体加到肉汤培养物中,试管就污染,而细菌的生长速度就明显地放慢,PLT-22是这类典型的噬菌体。
为了确定是否只有PLT-22具有FA的诱发活性,对LT-2菌株的休眠和生长细胞进行了各种处理。在幼龄培养物的滤液,或用苯自溶后,提取干细胞,用高浓度的抗菌素(青霉素、杆菌肽和金霉素)处理,或完全的噬菌体溶菌,都觉察不到有FA。稀释的抗菌素、氯酸锂和结晶紫产生不同的FA。用溶源噬菌体处理的培养物滤液最容易探查出复活性。这些结果说明,FA不是由于细胞机械化学的或生物学的瓦解而释放出来。但各种有害作用物所造成的情况,其方式或许同潜伏噬菌体的作用相平行。这些试剂最有效的浓度仅仅对细胞稍有抑制。如老龄培养物发生自溶也可探查出FA。这可归结为突变体溶源噬菌体的作用。
FA生产对化学刺激剂的反应方面还不足以控制得到实验用途所需的前后一致的产量。然而,当使用结晶紫或青霉素这类试剂处理LA-2后制备成一种不含或含有一点点FA的滤液,再把它接种到含有LA-2的肉汤中,就释放出大量的FA。这个步骤一直进行5个周期:FA明显的再生可能是第一次处理所释放的溶源噬菌体所起的作用。这个噬菌体缺乏一个可信赖的指示菌株,有碍于对这一反应进行分析。但是,在没有导入外源细菌或病毒的情况下从一个单一菌株诱发FA,它是一个有用的工具。
形态学和物理学研究
用Spinco超速离心机100000g 30分钟可定量沉淀并重新获得FA。用具有多速转头的国际离心机20000g产生部分沉降。因此,在这些制备中,FA超过大分子尺度。也能用一系列定孔胶膜(由S. E.Luria好意赠送)过滤来估测颗粒的大小。用A. P.D.420纳米的膜获得10%~20%的FA,230和170纳米则获得70%的FA以及120纳米获得99%的FA。这些结果表明一个颗粒的大小略小于0.1微米(Bawden,1950)。
在相差显微镜下,FA制备物呈现为一些小的勉强溶解的细粒。电子显微镜看到颗粒大小同过滤实验对FA的估测大致相符(见图2)。有些颗粒同抗O血清发生凝聚。通过离心可以除去在反应管中出现的絮状沉淀。然而,在上清液中,其活性仍然是完整的。用抗血清保温培养,一些颗粒增大,并在4小时后到达5~8微米的大小(Lederberg等,1951)。这些巨体同补加的细胞形成混合的絮状物。

图2 一个部分提纯的活性滤液×40000(由Paul Kaesberg进行电子显微摄影)
用诱发噬菌体或青霉素的FA处理沙门氏杆菌,经保温一小时和一个半小时产生了链,3小时后出现唯一的中央部分呈膨胀球状的蛇状物(Fleming等,1950)。也看到碎片和小颗粒。就在这个时候还产生FA。用常规方法难以对这些培养物的上清液消毒。用8或14磅测试的Mandler candles过滤得到每毫升约有100个计算成活的细胞的滤液。经证明未作处理的培养物相类似的滤液照例是无菌的。发现熔结的UF滤器对活性滤液的消毒过滤是合适的。
这些观察使人回想起Klieneberger-Nobel(1951)、Dienes和Weinberger(1951)和Tulasne(1951)所特别阐述的细菌的L类型。然而,在L类型和转导之间无功能关系的证据。从我们的培养物中使L-菌落生长以便作更直接的测验未获成功,其他工作人员对L类型生长所作的遗传分析,也未能支持关于它在生命史中所起作用的推测。
FA活性的起源和范围
迄今为止,规定FA是一种具有单独转导LT-22特殊突变体能力的LA-2菌株的特异性产物。但是,包括LA-22在内的其他直接杂交都得到了原养型细胞。为了确定能否从其他菌株得到FA,采用了一种最简单的测试办法,其中包括选择抗链霉素原养型细胞SRP(Lederberg,1951)。把SW-435(LA-22Sr )同鼠伤寒沙门氏杆菌50种不同的野生型(链霉素敏感原养型)的每一种混合培养生长,再把一些混合培养物涂布在每升含500毫克链霉素的最低琼脂上。28次杂交得到了明显的重组体,它说明有许多菌株都能产生FA。
当找到正常的刺激剂,25个鼠伤寒沙门氏杆菌的测验菌株的每一个菌株都能提取出FA。PLT-22用在许多对它敏感的菌株,它可能对SRP杂交的取得成功作出解释,而其他的溶源噬菌体(来自Lilleengen系列)则刺激别的菌株对PLT-22有抗性。总之,在10毫升的新鲜肉汤中接种109 个鼠伤寒沙门氏杆菌细胞和108 ~109 个对它敏感的溶源噬菌体颗粒,接种后4个小时将产生FA。低浓度的青霉素(每毫升1~5个单位)对某些培养是成功的。
借助于从营养缺陷型的混合平皿培养发现原养型,首次找到了沙门氏杆菌重组的证据。典型性别更完全的证明依赖于出现“非选择标志者”的新组合(Lederberg,1947)。在分别含有各种糖的EML琼脂上使SW-478(LA-22Gal- ,Xy- ,Mtl- ,Sr )与SW-414(LA-2Gal+ ,Xy+ ,Mtl+ ,Ss )杂交,从而可以在杂交平皿上直接记下一个未经选择的发酵性状。所筛选的约20000原养型细胞中,除它们的营养要求外,与SW-478毫无差别。除突变差异外,LA-22和LA-2在利用苹果酸、丙氨酸或琥珀酸作为生长必需的唯一碳源的能力方面有着本质的差别。所有的原养型都像LA-22。总共8个未选择的标记,都没有共同分离的证据。用来自LT-2的活性滤液重复了这些试验,所得结果相同。
如果安排试验以便对3个标记性状(一个营养的、两个发酵的)进行选择,则观察到每一个性状都有遗传转移。把SW-435(营养缺陷型,Gal- ,Xyl- ,Sr )同FA(来自LT-2的原养型,Gal+ ,Xyl+ ,Ss )一起涂布在最低的EMB半乳糖和EMB木糖琼脂上。在EMB培养基上首先出现一薄薄的生长层(粉红色,因此是非发酵的),然后出现使半乳糖或木糖发酵的小的乳头突起。这些突起长得很大(见图3),因为当其他营养耗尽时它们能利用糖。木糖缺陷突变体由于自发回复而产生一些突起,但不足以干扰试验记载。半乳糖缺陷突变体比较稳定,仅偶尔有回复。EMB上的突起数目或最低琼脂上的原养型细胞数(见表2)几乎相同,因而对不同选择性状的转导效率可说是一致的。然而,未选择的标记仍然不变;这就是说,所有原养型选择都是非发酵者,而突起选择只对一种糖有作用并为营养缺陷型。全部转导细胞仍为抗链霉素。
图3 涂布在EMB半乳糖琼脂上的SW-435,(1)用热失活和(2)用FA激活
表2 在不同选择培养基上的SW-435和FA①

①数字为每平皿上菌落或突起数。
在双重糖琼脂上重复上述试验。个别突起或使半乳糖、或使木糖发酵,且都是营养缺陷型。由于它们在质地上稍有差别,有可能直接在指示平皿培养上直接区分出这两种突起。挑出整个突起并转到另一种糖和最低琼脂上。在许多测试的乳头突起中,没有发现混合的突起。用这种严格的选择可以探查出任何一种这样的突起。
根据这些试验,我们得出结论,一种FA滤液有许多活性,产生多种多样的转导(但每个细胞不超过一种),它单独地产生转导菌落。
我们没有观察到像在大肠杆菌重组中发现的那种连锁分离。如不能证明连锁是由于亲本染色体结构上的差异,FA的独一无二的活性或许仍然同配子的解释相一致。另一种观点是,或可把FA当做一种非特异性诱变剂,它对不同因子具有独立的作用。进一步试验无疑对这两种观点都否定了。
LT-7作为一个有效的给体,FA为受体,并选用它们来研究菌株内转移和检验这些考虑(有关它们的标记,见表1)。为使所采用的FA来源肯定起见,要在无外部细菌或病毒的影响下进行制备(如上所述)。从SW-184(脯氨酸缺陷)、SW-188(蛋氨酸缺陷)和SW-191(亮氨酸缺陷)制备出FA。每一种制备物用来对3个LT-7营养缺陷型的每一种类型和LA-22(存在每一种活性的对照)测定其从营养缺陷型转导到原养型。制备物对LA-22具有相当一致的活性。然而,从3种LT-7营养缺陷型分别得到的FA,各自能转导另外两种培养物,但不能转导其起源的培养物(见表3)。因此,FA同产生它的细胞的基因型是一致的。由3种营养缺陷型分别得到一些半乳糖缺陷突变体。数千个被转导的原养型细胞没有一个是利用半乳糖的。从SW-184(脯氨酸缺陷)得到的FA,把它与SW-188(蛋氨酸缺陷)一起涂布在补加脯氨酸的最低琼脂上,只产生对脯氨酸独立的菌落(原养型)。分别用3种营养缺陷型所作的相似试验都得到了类似的结果。在转导过程中,没有发生连锁分离,或这3个营养标志彼此间或同发酵标志间的联合。抗链霉素仍然是另一种标志,它在细胞转导成另一种性状时仍然保持不变。
表3 来自LT-7及其衍生菌株的FA对LT-7衍生菌株的效应
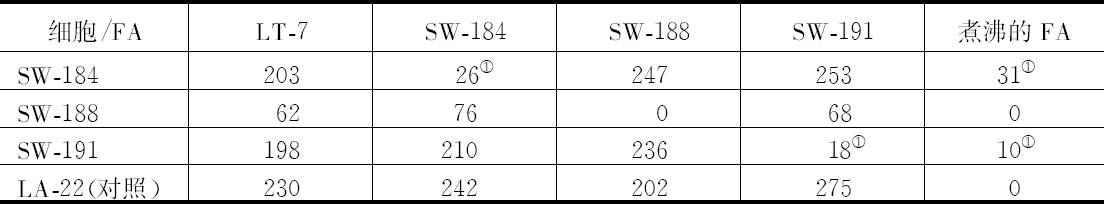
①大概是自发回复突起。数字为每平皿上从营养缺陷型转导成原养型的细胞数。
用它们的亲本野生类型得到的FA,使一些半乳糖缺陷突变体转导成能利用半乳糖的。从这些突变体得到的FA得到相反的结果。突变体绝不能用它们自己的FA转导,但用别的一些突变体得到的FA则能使它转导。这些相互作用对于把突变体在等位性或基因相同性方面进行归类提供了依据(见表4)。
表4 来自几个半乳糖缺陷突变体的FA对这些相同突变体的效应
①产生能利用半乳糖的乳头突起。
②突起不多于对照。
迄今讨论的全部转导都属于突变野生类型的范围。照例,由于缺乏适宜的选择方法而难以在其他方面筛选变异。用链霉素抗性能做到这一点,因为野生型状态对链霉素是敏感的(Ss ),而突变体是抗链霉素的(Sr )。把刚收获的细胞,用抗链霉素和对链霉素敏感的亲本得到的FA处理,然后涂布在EMB半乳糖上。经保温2小时后(Davis,1950),在平皿上喷上浓缩的链霉素溶液(每毫升0.1克)。表5说明只有当所使用的FA,其来源是抗链霉素的突变体时,才发生这种转导。没有发现有联系的变化。通过把许多子菌落反复涂布于正常的和含有链霉素的培养基上进行试验(Lederberg和Lederberg,1952),来验证转导细胞的稳定性。
表5 从抗链霉素和对链霉素敏感细胞得到的FA,对敏感细胞效应的比较①

①表中数字为每平皿抗链霉素菌落的数目。
现在很清楚,已对这种特定的FA规定出一种测定方法,这种FA恰恰是某种滤液的几种共存功能之一。有人建议我们把FA称之为截至目前所研究的任何一种遗传因子,且可以用同样的方法把某种滤液的作用范围叫做培养物的基因型,而滤液就是从这种培养物得到的,例如SW-514、原养型、半乳糖(Gal)+,木糖(Xyl)+,Sr (见图4)以及从这种培养物得到FA。但除非另有证明,将继续把FA称之为对LA-22测出的转导
FA(SW-514,原养型,GAL+ ,XYL+ ,Sr )
SW-351(缺陷型,GAL- ,XYL- ,Ss )
图4 一种活性滤液的复合潜力
FA的吸附作用
转导的第一步必须使FA吸附于足够多的细胞。从肉汤琼脂平皿培养获得LA-22。每隔一定时间将取样悬浮在1毫升的活性滤液。沉降细胞并把它涂布在基本培养基上以确定交换的数目。在56℃条件下进行热击以破坏所有未沉降的细胞后,用LA-22在未吸附的FA方面测定悬浮液。在离心所需的时间(15分钟)内,适量的FA完全有吸附作用,而在沉淀细胞中数量重新恢复。
鼠伤寒沙门氏杆菌所有测验的光滑型菌株都吸附FA。给体菌株细胞像其他菌株一样能有效地吸附,并自始至终在菌株内转移方面取得了成功。用煮沸或紫外线照射进行消毒(留下极少量存活的碎片)并不影响吸附作用,在肉汤中延长培养所选出的粗糙培养物(Page等,1951)没有吸附作用。这些结果表明,吸附的部位是热稳定的,不受细胞死亡的影响,并可能同体质抗原有关。
用以上所用的量,FA测定与FA浓度直接成比例。从肉汤琼脂得到LA-22细胞。10个离心管,每管沉降出含有1010 细胞的样品,然后废弃上清液。添加FA复合样品(1~10毫升),并在37℃下放置15分钟使其吸附。从EML半乳糖中收集上清液和细胞并进行测定,在原养型细胞中没有观察到一致的变化(即能利用半乳糖)。图5表示使用8毫升左右的FA时所发生的最高数目的转导。饱和的沉淀物不再有更多的样品部分吸附FA。除了可能因机械原因而有一小部分有规律的丢失外,上清液或沉淀物占有FA的所有单位。
图5 FA的吸附
LA-22 1010 细胞接受FA(LT-2)处理15~30分钟。经离心,分别测定残余的FA和已开始的转导后收集上清液和沉淀物。
饱和对吸附的干扰在一项阻断试验中得到了更明确的证明。SW-188(蛋氨酸缺陷,M- )用过量的来自M- 细胞的FA处理15分钟,然后在细胞再次沉降前对被沉降的细胞加入LT-7(M+ )的FA再处理15分钟。M+ FA既没有结合,也没有发生SW-188转导。这证实断阻概念并表明在15分钟到达饱和后,吸附作用是不可逆的。
鉴于FA的吸附作用如此迅速,看来可以放心地认为,各个细菌的大部分是能够吸附它的。用细菌饱和所需的毫升数(每个细菌一个颗粒)除以细菌总数,我们能得出每毫升这种滤液可吸附颗粒数的近似的最低估测为1010 /8。可以指望活性滤液每毫升超过1011 颗粒而不出现混浊,每毫升最高的颗粒数就是根据这一事实来估测的。
要用这么许多无法检验的假设在这里详尽讨论吸附作用动力学的可能模式是不太合适的。但可以指出,双交换的低频率或频率等于0并不意味着一种FA颗粒排除另一种FA颗粒。如果大多数细菌可以被转导,那么一种特定转导的频率将是一种概率,即每一个被吸附的颗粒都有一特定的效应。双重转导作为单交换的绝对频率,其比率与单交换相同,而这个频率(据计算为10-5 )要在我们的试验中探查出双交换则太低了。但是,如果转导局限在少数有转导能力的细胞,则双重转导没有独立的概率,并需要诸如相互排斥之类的另一些假设,对所观察到低频率的双重事件作出说明。
看来下述情景最符合于迄今观察到的结果。一个活性滤液是由许多种颗粒组成的总体,每一种颗粒相当于一种遗传效应,虽然有一些在本质上是惰性的。每个细菌可吸收数目有限的颗粒,其可能的范围从一个颗粒到大约100个颗粒。每一个被吸附的颗粒有一固定的独立的概率发挥其特定的转导效应。单转导,尤其是双转导的频率之低是受到可能被吸附的颗粒总数的限制,也可能受到一个被吸附的颗粒就能完成其效应这种极罕见的概率所限制。
连续转导
单项试验从未看到过双重转导。其原因为以上所谈的一些考虑,而不是连续转移所出现的一些固有的局限性。一旦转导出一个细胞,它就能长大,并把它重新置于FA中,再就其他方面的变异进行选择。对SW-351(营养缺陷型、半乳糖- 、木糖- )连续转移从营养缺陷型到原养型,从半乳糖- 到半乳糖+ ,和从木糖- 到木糖+ 。完成转移的次序无差别。与SW-351在每个性状方面单一转导相比较,重复转导的效率没有消失。
FA吸附作用的特异性
吸附试验表明吸附能力和免疫特异性有相关性。用大约12个沙门氏杆菌属血清型所作的初步试验肯定了这种相关性,并把它缩小到出现菌体抗原Ⅻ上。使供试的血清型肉汤培养物沉降,再加入1毫升FA。吸附进行15分钟,然后在56℃条件下使反应管热击一个小时使细胞消毒。用已知吸附细胞所作的初步试验证明,一旦FA被吸附后,用这个方法就不能把它洗提出来,以LA-22测定混合物游离的FA。用这种方式测试了约50种不同的血清型。尽管有几个具有Ⅻ的类型是惰性的,但没有一个类型不吸附Ⅻ。Salmonella coli类型保持了这种相关性。吸附的Ⅻ菌株有:乙种副伤寒沙门氏杆菌、鼠伤寒沙门氏杆菌(25个菌株)、斯坦利沙门氏杆菌、海德尔堡沙门氏杆菌、切斯特沙门氏杆菌、圣地亚哥沙门氏杆菌、绵羊流产沙门氏杆菌、伤寒沙门氏杆菌W、伤寒沙门氏杆菌V、肠炎沙门氏杆菌、莫斯科沙门氏杆菌、布利丹沙门氏杆菌、伊斯特本沙门氏杆菌、仙台沙门氏杆菌、奥博尼沙门氏杆菌、大肠杆菌3、大肠杆菌4、卡普士得沙门氏杆菌、萨林纳斯沙门氏杆菌、雏白痢沙门氏杆菌和鸡沙门氏杆菌。以下Ⅻ型不吸附:甲种副伤寒沙门氏杆菌和牛流产沙门氏杆菌,大概因为它们缺乏Ⅻ2 组分。测验过的不吸附的非Ⅻ型则有:鼠伤寒沙门氏杆菌(粗糙变体),猪霍乱沙门氏杆菌,纽波特沙门氏杆菌、伦敦沙门氏杆菌、桑夫顿堡沙门氏杆菌、阿柏丁沙门氏杆菌、浦那沙门氏杆菌、沃信顿沙门氏杆菌、非丁伏斯沙门氏杆菌、肯塔基沙门氏杆菌、威奇塔沙门氏杆菌、厄班那沙门氏杆菌、哈瓦那沙门氏杆菌、阿尔腾道夫沙门氏杆菌、瓦伊勒沙门氏杆菌、梦得维的亚沙门氏杆菌、大肠杆菌1、大肠杆菌2、大肠杆菌5、大肠杆菌K-12、波那雷恩沙门氏杆菌、佛罗里达沙门氏杆菌、马德利亚沙门氏杆菌。
类型间转导
FA的吸附作用是否足以表明遗传转移的敏感性,这一点尚不清楚,但初步的资料确证有一可能的受体组,或许有可能进行类型间转导。
伤寒沙门氏杆菌和鼠伤寒沙门氏杆菌在一些培养和血清性状方面是各异的。后者能发酵阿拉伯糖和鼠李糖,而前者则不能使这两种糖发酵,且这两种糖对它皆有抑制作用。伤寒沙门氏杆菌用来自鼠伤寒沙门氏杆菌的FA处理,并接种到含有百分之一阿拉伯糖或鼠李糖的肉汤的Durham发酵管中,24小时后FA处理的培养物生长得更加旺盛,约48小时产生出酸。从这些发酵管分离出的培养物,它们与伤寒沙门氏杆菌的差别,仅仅是对这些糖的发酵能力不同而已。无FA处理的对照培养物,生长得很少,无发酵证据。虽然鼠伤寒沙门氏杆菌从鼠李糖和阿拉伯糖产生出气体,但这些新的类型仍然是典型的厌氧性。试验也在琼脂上进行。把处理细胞涂布在EMB阿拉伯糖和EMB鼠李糖上。伤寒沙门氏杆菌偶尔突变成一种非抑制类型(Kristensen,1948),其特点为一白色乳状突起,它在试验和对照平皿上都可以观察到。然而,只在试验平皿上看到紫红色(发酵)的乳状突起。它们在培养中很像在肉汤中转导后分离来的发酵菌株。用伤寒沙门氏杆菌的另外两个菌株重复得到这些结果。用鼠伤寒沙门氏杆菌的一个抗链霉素突变体作为FA来源,把这个性状转移给伤寒沙门氏杆菌是可能的。曾试图用FA处理伤寒沙门氏杆菌以产生对葡萄糖的厌氧发酵,但都失败了,这可能因为要探查出这个性状被转导的细胞的选择条件不够的缘故。
伤寒沙门氏杆菌的抗原特性是Ⅸ、Ⅶ: d——(单相),而鼠伤寒沙门氏杆菌是Ⅰ、Ⅳ、Ⅴ、Ⅶ: i——1、2、3。伤寒沙门氏杆菌接受来自鼠伤寒沙门氏杆菌FA的处理,并选择鞭毛抗原的转导。取一个以真菌生长管为基础的试管(Ryan等,1943),一半装有含稀释抗-d血清(1/200的血清滴定到1/5000)的软琼脂。在试管的一边大量接种细胞,并注视其移动情况。一项试验,4个试验管中有两管出现移动,而3个对照管则接种体完全固定不变。移动细胞和固定的接种之间界限分明。从管的没有接种的一端掏出移动细胞,并进行培养和血清学测定。两个分离体在培养上都像伤寒沙门氏杆菌。它们中有一个与抗i血清起反应,而另一个则对伤寒沙门氏杆菌或鼠伤寒沙门氏杆菌鞭毛抗血清都无反应,从而判断它是j相(Kauffmann,1936)。P. R.Edwards肯定了这两个菌株的分析。从108 FA饱含伤寒沙门氏杆菌细胞的31个供试接种体中有12个得到了i抗原的转导。试验和对照管中都偶尔出现“j”相。50个无FA的对照测验中都没有发觉i相。“杂种”的完全抗原分析为Ⅸ、Ⅻ: i——。与鼠伤寒沙门氏杆菌衍生出i鞭毛不同,这些“杂种"都没有发现相的变异。现在继续进行试验以探究别的鞭毛和体质抗原的转导。
转导细胞
通过转导[FA(LA-2)对LA-22]产生的原养型细胞,测试了它们在无性生殖和进一步转导方面的稳定性。从试验平皿上分离出以后,用划线分离予以纯化。5个单菌落在完全肉汤中生长并涂布于平皿培养。每一种各挑200个菌落,并在最低琼脂培养基上再测试:全部都是原养型。这种被转导的培养物再接受FA处理,以选择别的变异(半乳糖- 变为半乳糖+ )。通过重复涂布平皿培养,测试了大约1500个菌落,全部菌落都保持了成为原养型的最初的转导。
转导培养物在其生长期间不释放FA,除对亲本培养物所用的方法外,采用任何其他措施都不能得到FA。在这方面由于菌株内转导的产物而碰到了一些困难。它们全都是与活性滤液有联系的噬菌体的抗性载体,而需要一些新的噬菌体从它们中间诱发出FA。噬菌体的抗性也使重复诱导的效率降低,这大概因为减弱了FA的吸附作用。
自发回复突变使转导其突变亲本的能力得到恢复,其情况犹如转导回复变异那样。这是说,当不管用哪一种方法使一个细胞从A-变为A+,它能重新产生A+物质。对游离FA的突变尚未进行研究。
细菌噬菌体和FA之间的相互关系
一些最新收集的证据指出FA颗粒和细菌噬菌体的同一性。FA和噬菌体具有一共同的滤过终点;用一个120纳米的A. P. D. 膜,两者都能保留99%。与菌体抗原Ⅻ相联系,它们对沙门氏杆菌属血清型的吸附具有共同的特异性。在吸附鼠伤寒沙门氏杆菌时两者都在同一点上达到饱和,且噬菌体对FA的比率保持恒定不变。提纯过程中,FA和噬菌体仍然在一起。在短期试验,从侵染细菌的噬菌体中同时释放出FA和噬菌体。电子显微镜下呈现出正常大小颗粒在形态上的相似性。
下述试验说明噬菌体颗粒只能是转导遗传物质的一个被动的载体。从生长在细菌细胞上的单一噬菌体的颗粒,有得到高滴定度的噬菌体和包括亲本细胞整个基因型,但只能使每个细菌细胞发生一次转导的FA总体。从这种滤液得到的单一噬菌体颗粒,能在亲本起源相同,但基因型不同的细菌细胞上生长。所产生的FA相当于第二给体的基因型。
在FA诱发作用一节中,曾提到过借助于转移使FA明显地再生。其解释是FA同噬菌体的联合,噬菌体不断刺激FA的产生。为测试这一点,用青霉素处理A-、B+、C+细胞。用相同的细胞转移滤液以产生FA(A-、B+、C+)和一种能在这些相同细胞上作测定的噬菌体。当添加到A+、B-、C+细胞(来自相同起源的亲本)时,所得到的FA是A+、B-、C+。所有B+制剂都被吸附而消失掉,从而得到了与B+细胞基因型相平行的制剂。FA不是这样来繁殖,而是为了进一步生产,同必须的刺激剂噬菌体相结合。
讨论
鼠伤寒沙门氏杆菌的遗传交换是由一细菌产物,即我们所说的FA(滤过性物质)来进行传递的。一种个别的活性滤液能把许多遗传特性从一个菌株转移(转导)到另一个菌株。虽然这种滤液的全部活性包括其亲本培养物的基因型,但每一次转导只传递每一个细菌的一个单一的特性。这与大肠杆菌K-12菌株的遗传交换不同,那里是几个区分两个亲本的遗传标志进行不受限制的重组。
可以把FA看做是进入被转导细胞固定遗传的遗传物质。我们可以提问,究竟这种转移是一种单纯的添加物还是固定遗传因子的一种可以取代的交换和替代?如果根据大肠杆菌杂合二倍体研究(Lederberg,1951b)来推理链霉素抗性是一隐性突变,那么抗性的转导与简单添加的机理就不符。
必须小心辨别FA的两种特点:颗粒本身的生物学性质及其遗传功能。有充分理由要用细菌噬菌体来鉴定颗粒。然而,噬菌体颗粒在功能上是把遗传物质从一个细菌转导到另一个细菌的被动载体。这种遗传物质只相当于细菌基因型的一部分。例如,当把一个标志原养型的FA同一个营养缺陷型一起涂布在最低琼脂培养基上,在被转导的原养型中,看不到假设为“给体核”的基因型。假如所产生的这种独一无二的效应取决于偶尔有机会从一个复合的颗粒释放出某种特定的活性,或者细胞中发生一些局部化的非遗传性事件,它通常只留下一种对转导敏感的功能,那么可以坚持主张FA是一种遗传复合体而不是一个遗传单位的假说。这种最初单个转导的细胞发育成一个独立的菌落。由于这种菌落包括约107 个细菌,人们可以指望,一个FA颗粒的复合残留物,如果是存活的,将在菌落生长期间,使一些子细胞向别的性状转导。但是,每一个FA颗粒只产生一个单独的转导菌落。这说明它的组成同其遗传效应一样的简单。
当LA-22从营养缺陷型(苯丙氨酸缺陷和酪氨酸缺陷;色氨酸缺陷)转导成原养型,我们就有一个明显的双重变异。如把这个突变体涂布在辅加苯丙氨酸和酪氨酸的最低琼脂培养基上,它偶尔会回复到第一步的营养缺陷状态。然而,当LA-22在这个培养基上被转导的话,所发现的第一步营养缺陷型绝不会超过用自发回复所能解释的范围。大多数选出的菌落是原养型。在其他任何哪一种菌株间或菌株内转导,我们都不能使转导的影响超过一个特性。LA-22的营养像是由相同遗传位置上两个连续突变决定的。Davis(1951)关于芳香族生物合成的设计同这一见解相符。虽然LA-22这个突变体能自发回复到一个中间性的等位基因,但转导使野生型基因取代成充分的合成。
对FA细粒似乎最有理的假设为,它们是核素的一个异质群体,而每一种核素有它自己的能力,换句话说,每一种核素负载一“单基因”或小的染色体片段。
不管FA颗粒的性质如何,对于把转导的遗传物质引入受体细胞固定的遗传,必须假定一些机理。这里顺便说一下Muller(1947)对肺炎球菌转化类型的分析:“……实际上,还有存活的细菌染色体,或部分染色体,自由地流入所用的培养基中。据我所见,这些染色体可侵入无荚膜的细菌,并或者同寄主染色体进行一种交换后,在那里部分地扎下了根。”
在一篇有关沙门氏杆菌重组系统的初报中提到(Lederberg等,1951),FA可能同细菌的L类型(Klieneberger-Nobel,1951)有关。对某些制剂起反应而发生膨胀的“蛇”,可滤的颗粒和巨体,皆为FA和L类型所特有的。除了提出经过滤器成活的颗粒外,我们不能重复所报道的周期。这种可见的凝集颗粒和诱发膨胀形状的抗血清对FA的活性并非必需。但是,不能用一个简单的设计使全部成员都适用,这可能因为它比我们目前所知的系统更要复杂得多。
细菌学文献中有许多报道结果可以解释成转导(参见Luria,1947和Lederberg,1948的评论)。这些试验由于它们繁殖和定量方面的困难而受到批评或被忽视,但现在借助于所提出的发现可予以重新研究。目前引用一些更恰当的试验应该可以得到满足了。Wollman(1925)报道了用大肠杆菌经滤过性物质获得沙门氏杆菌的免疫特异性。相似的材料(它能通过细菌体溶菌作用得到)牵涉到抗青霉素的葡萄球菌和链球菌对相对的青霉素敏感性的改变(Voureka,1948;George和Pandalai,1949)。曼彻斯特型副痢疾志贺氏菌(Weil和Binder,1947)当用异源类型的提取液处理,就得到了新的免疫特异性。Boivin(1947)报道了大肠杆菌的一种相似的变异。不幸的是他的菌株已丢失而不能再予以论证。Bruner和Edwards(1948)在一篇关于沙门氏杆菌生长在特异血清中引起体质抗原变异的报道中评论了这种可能性,即在血清中溶解的细菌产物与这种变化有关系。
这些有争议的系统,不足以为详尽比较沙门氏杆菌属的转导提供证明。肺炎球菌(Avery等,1944;McCarty,1946)和流感嗜血杆菌(Alexander和Leidy,1951)的转化研究得更完整。
肺炎球菌荚膜性状的遗传转化取决于一种特殊的细菌产物(肺炎球菌的转化原理,PTP)。最初,把它解释为一种直接突变,现在认为是一种遗传交换(Ephrussi-Taylor,1950)。这样,完全的荚膜性状(Griffith,1928),一系列中间性荚膜性状(Ephrussi-Taylor,1951),M蛋白质性状(Austrian和Macleod,1949)和青霉素抗性(Hotchkiss,1951)都得到了转化。如同沙门氏杆菌一样,每一个性状都是独立转化的。但这两个系统之间有一些差别。FA必须是诱发的,而PTP则可从健康细胞中提取。FA对各种化学处理的抗性只得出与其化学性质相反的证据。脱氧核糖核酸在PTP中的作用已通过脱氧核糖核酸酶使之失活而予以证实。用定孔胶膜保持活性,得到对两个不同性状有影响的FA颗粒大小(约0.1微米)的可比较的估测值。而另一方面,经各种估测,PTP的颗粒大小从平均500000个离心团(Avery等,1944)到相当于具有高度不对称性的18000000分子量(Fluke等,1951)的一个电离辐射敏感体积。它比起FA颗粒要明显小得多。用一复合血清系统,肺炎球菌必然在吸附PTP方面是敏感的。转化频率低而不易测定,看来是因为细菌的低能力的缘故。在缺乏吸附试验的情况下,不能排除有一种相似于沙门氏杆菌属的系统。这两个系统还缺乏重要的资料,而时间会解决这些明显的差别。
沙门氏杆菌转导与大肠杆菌有性重组的关系尚模糊不清。在可杂交的大肠杆菌或沙门氏杆菌的有性重组中都没有发现过转导。这两个属在分类学上关系密切,但似乎有着完全不同的遗传交换的模式。
在大肠杆菌K-12菌株首次证实了有性重组。随着发展出一种有效的筛选措施,已证明有2%到3%的大肠杆菌分离体与K-12菌株杂交(Lederberg,1951)。大肠杆菌重组的作用物差不多肯定是细菌细胞。细菌明显地配对,形成接合子,亲本和重组体细胞从接合子起开始以后的减数分裂,其中连锁为一突出的特点(Lederberg,1947)。意外地发生不分开而继续分离为单倍体和二倍体染色体组(Zelle和Lederberg,1951),证实了在一个单细胞内染色体组的结合。虽然在沙门氏杆菌的转导中,溶菌性起了重要的作用,但大肠杆菌培养物溶菌和非溶菌的所有组合杂交都是同样方便的(Lederberg,1951)。
鉴于缺少未选择标志者的重组,对某些类型的遗传分析来说,转导与有性重组相比,是一个用途较差的工具。但由于FA或许同细胞外遗传物质有关,诸如基因繁殖,新陈代谢和突变等问题可能更容易得到解决。有性系统通常能提供遗传物质的再分配,并且是有机界进化中实现自然选择的一个重要的变异来源。有性重组和转导,由于其频率太低都只能局限于细菌的遗传互换。转导交换则受频率和范围两方面的限制。
要估价转导在发展免疫上复合的沙门氏菌种所起的作用尚嫌为时过早。White(1926)曾猜测,从一个具有所有许多可能的抗原的单一菌株丧失变异可得到许多血清类型。Bruner和Edwards(1948)得到了使当今物种丢失变异的特殊例子。转导对一些在“后代”系之间自发和独立产生的变异如何进行转移提供了机理。沙门氏菌属包括一批血清型,它们是鼠伤寒沙门氏杆菌FA的一种受体。正在寻找其他的受体种类。在这类受体中,有可能在实验室发展出其他新的可与伤寒沙门氏杆菌和鼠伤寒沙门氏杆菌抗原杂种相媲美的血清类型。
几个不同的细菌属已就其遗传交换的模式进行了深入的研究。这几个已知的系统,每一个在细节上都有所不同,这扩大了我们对细菌繁殖和遗传的见解。
小结
当鼠伤寒沙门氏杆菌生长在具有种种适度的有害物质,特别是微弱的溶菌噬菌体时,它产生出一种滤过性物质(FA),能使遗传特性从一个菌株转移到另一个菌株。
各滤液能转导许多不同的特性,但一个单细胞不超过一种。滤液的活性同给体细胞的特性是平行的。营养、发酵、药物抗性和抗原性状都进行了转导。新的性状经多代继代培养后成为稳定。
FA对于氯仿、甲苯和乙醇一类的消毒剂和胰酶、胰蛋白酶、核糖核酸酶和脱氧核糖核酸酶等一类酶是有抗性的。FA的颗粒大小,经定孔胶膜过滤测定,约为0.1微米。FA吸附作用迅速,且在供试的各种血清型中与存在体质抗原Ⅻ有关。
对任何一个性状转导的最高频率为2×10-6 ,吸附期间为饱和所限制。观察到一些类型间的转移。例如,来自鼠伤寒沙门氏杆菌的i鞭毛抗原转导到伤寒沙门氏杆菌而产生一些新的血清类型:Ⅸ、Ⅻ;i,——。把沙门氏菌属的遗传转导同嗜血杆菌和肺炎球菌的“类型转化”以及大肠杆菌的有性重组作了对照和比较。
噬菌体生长过程中蛋白质和核酸各自的功能 【61】
阿尔弗雷德·赫希(A. D.Hershey) 【62】 玛莎·蔡斯(M. Chase)
(美国 冷泉港实验室,1952年)
Doermann(1948)Doermann和Dissosway(1949)以及Anderson和Doermann(1952)的工作说明,噬菌体T2、T3和T4,是以一种非侵染形式在细菌细胞中增殖的。这和某些溶源性细菌携带的噬菌体是同样的情况(Lwoff和Gutmann,1950)。对这些病毒的营养生长期尚不了解。本文报道的实验表明,T2生长的最初阶段之一是病毒颗粒的核酸,从它的蛋白质外壳中释放出来,此后,这些含硫蛋白质就没有其他作用了。
材料和方法
本文中噬菌体T2指的是一个叫T2H的变种(Hershey,1946);T2h指的是T2的一个寄主范围改变的突变体;uv- 噬菌体指的是噬菌体用一灭菌的紫外灯(通用电器公司)照射,使存活率为10-5 。
敏感细菌指的是大肠杆菌的一个对T2及T2的h突变体敏感的菌株H;抗性菌B/2指的是一个抗T2、但对T2的h突变体敏感的菌株;抗性菌株B/2h指的是一个对这两者都抗的菌株。这些细菌不能吸附它们所能抵抗的噬菌体。
低盐肉汤培养基,每升含有bacto-蛋白胨10克、葡萄糖1克、NaCl 1克。肉汤培养基除了这些成分外,还含有bacto-牛肉浸汁3克、NaCl 4克。
甘油-乳酸盐培养基每升含有乳酸钠70mM 【63】 、甘油4克、NaCl 5克、KCl 2克、NH4 Cl 1克、MgCl2 1mM、CaCl2 0.1mM、明胶0.01克、磷(如正磷酸盐)10毫克、硫(如MgSO4 )10毫克、pH7.0。
吸附培养基每升含有NaCl 4克、K2 SO4 5克、KH2 PO4 1.5克、Na2 HPO4 3克、MgSO4 1mM、CaCl2 0.1mM、明胶0.01克、pH7.0。
佛罗那(Veronal)缓冲液每升含有二乙基巴比土酸钠1克、MgSO4 3mM、明胶1克、pH8.0。
本文中提到的HCN是由氰化钠溶液组成,使用时用磷酸中和。
把培养18小时的细菌,在37℃热处理10分钟,并用吸附培养基洗涤,然后与同位素样品在吸附培养基中混合,测定细菌吸附的同位素,混合物在37℃保温5分钟,再用水稀释、离心。分别对沉淀和上清液进行分析。
用抗血清沉淀的同位素测定方法,是在每毫升约含1011 无放射性噬菌体的0.5%盐溶液中,把同位素样品和稍多于最低量的抗噬菌体血清(最后稀释度为1∶160)混合,会产生明显的沉淀。混合物在37℃保温2小时后离心。
DNase试验的做法是:用佛罗那缓冲液稀释过的样品,经过预热,每毫升加0.1毫克晶体的DNase(华盛顿生化实验室),37℃保温15分钟。样品冷却后,用5%TCA(每毫升中含有1毫克血清白蛋白)沉淀,离心。然后测定酸溶性同位素。
在离心分离过程中得到的所有沉淀都未经洗涤,因此含有5%的上清液。沉淀和上清液都进行分析。
用一个在末端开口的Geiger计数器来测定放射性,为了避免样品本身吸附作用造成的损失,采用的样品要少,而且要充分干燥。在测定32 P绝对值时,使用的32 P溶液,以及标准溶液,都是从国家标准局得到的。在测定35 S的绝对值时,我们采用了供给同位素单位(美国橡树岭国家实验室)提供的分析(±20%)。
用甘油-乳酸盐培养基培养细菌,在磷和硫的浓度低时,pH比较稳定,这种培养基对于本文中提到的其他一些实验也是有用的。
在这种培养基中,敏感菌的18小时培养物,每毫升约有2×109 个菌,在对数生长时没有延迟期,而且在同种培养基中进行继代培养时,不管接种量的大小,每个细胞都没有光散射的改变。在37℃增殖一代的时间是1.5小时。这种细胞要比在肉汤中培养的细胞小些。在这种培养基中,T2表现出有22~25分钟的潜伏期。用氰化物和UV-噬菌体(在文章内容中将叙述)的溶菌作用得到的噬菌体产量:在15分钟的时候,每个细菌1个;在25分钟的时候,每个细菌16个。在稀释培养物中,50分钟的时候,达到最终释放量是每个细菌有30~40个噬菌体。在每毫升为2×108 个细菌时,培养物慢慢地溶菌,每个细菌可产生140个噬菌体。在这种培养基中,细菌和噬菌体的生长都和在肉汤中一样,重复性很好。
为了制备同位素标记的噬菌体,把比活性为0.5毫居里/毫克的32 P或比活性为8.0毫居里/毫克的35 S加入甘油-乳酸盐培养基中,先让细菌至少生长4个小时,然后加进噬菌体。噬菌体感染后,将培养物通气培养过夜,用低速(2000×g)和高速(12000×g)交替离心各三次,分离出同位素标记的噬菌体,噬菌体悬液贮存时,浓度不要超过4微居里/毫升。
这种制品中每个活的噬菌体颗粒含有(1.0~3.0)×10-12 微克的硫和(2.5~3.5)×10-11 微克的磷,有时制品含硫量过多,可用热处理杀死了的、不能吸附噬菌体的细菌的吸附作用来改良。由于存在失活的噬菌体颗粒和空的噬菌体“外膜”,影响了制品的放射化学纯度。制品中的硫(约20%)可以被抗噬菌体血清沉淀(见表1),也可以被抗噬菌体的细菌吸附,但不能被对噬菌体敏感的细菌吸附(见表7),这说明制品中污染有“外膜”物质。表1的资料表明,就我们的研究目的来看,细菌原来的污染可以忽略不计。为了证实我们的主要发现反映的确实是活噬菌体的特性,在本文末尾引用了用失活噬菌体做的一些实验。
表1 质壁分离的噬菌体的“空胞”和溶液的组分
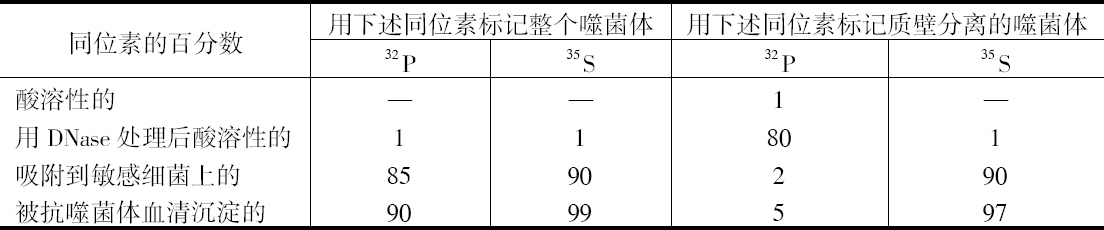
静止噬菌体颗粒的化学形态
Anderson(1949)发现将T2噬菌体颗粒悬浮在高浓度的氯化钠中,再用水把悬浮液快速稀释,可使噬菌体失活。在电子显微镜下可见这种失活的噬菌体好像是蝌蚪形的“空胞”。假定慢慢稀释,就不会引起噬菌体失活,因此认为这种失活是由于渗透压骤变造成的。并且推论,这种颗粒具有一种渗透膜。Herriott(1951)发现渗透压骤变能使噬菌体颗粒的DNA释放到溶液中,“空胞”可以吸附到细菌上,并使细菌溶解。他指出这是用病毒物质鉴定病毒功能的一个开端。
室温下,将噬菌体(1011 个/毫升)在3M的NaCl溶液中悬浮5分钟,并迅速在悬液中注入40倍体积的蒸馏水,能使同位素标记过的T2发生质壁分离。这种质壁分离的噬菌体,存活者不到2%,然后用表1的几种方法分析磷和硫。结果从以下几点证实并发展了以前的发现:
(1)质壁分离将噬菌体T2分离成“空胞”和溶液两部分,整个噬菌体颗粒所含的硫几乎全在“空胞”中,而DNA则几乎全在溶液中。
(2)“空胞”含有噬菌体颗粒的主要抗原,这种抗原用抗血清可以检查出来。DNA释放出来后成为游离酸,也可能与不含硫、也无抗原活性的物质结合在一起。
(3)“空胞”能特异性吸附到对噬菌体敏感的细菌上去;DNA则不能。
(4)“空胞”是蛋白质外壳,它裹在整个噬菌体颗粒DNA的外面,能与抗血清起反应,能保护DNA免受DNase破坏,它带有能吸附到细菌上去的“器官”。
(5)噬菌体的质壁分离是由于渗透压骤变造成的,因为如果将噬菌体悬浮在盐溶液中缓慢地稀释,就不能使它失活,而且它的DNA也不会暴露在DNase中。
噬菌体吸附到细菌上后,其DNA变得对DNase敏感了
上述的静止噬菌体颗粒的结构说明,噬菌体的增殖可能首先要改变或除去这种病毒颗粒的保护性外壳。估计这种改变本身就说明噬菌体DNA变得对DNase敏感了。表2列出的实验说明了这一点。这些结果可归纳为以下几点:
(1)噬菌体吸附到加热杀死的细菌上后,其DNA变得对DNase非常敏感。
(2)噬菌体吸附到活细菌上后,放在80℃加热10分钟,其DNA也变得对DNase敏感了。在这个温度下不发生吸附的噬菌体,对DNase是不敏感的。
(3)吸附到未经加热的细菌上去的噬菌体DNA,对DNase是有抗性的,推测是因为酶透不过细胞结构,因而起了保护作用。
表2 噬菌体吸附到细菌上后,其DNA变得对DNase敏感了①
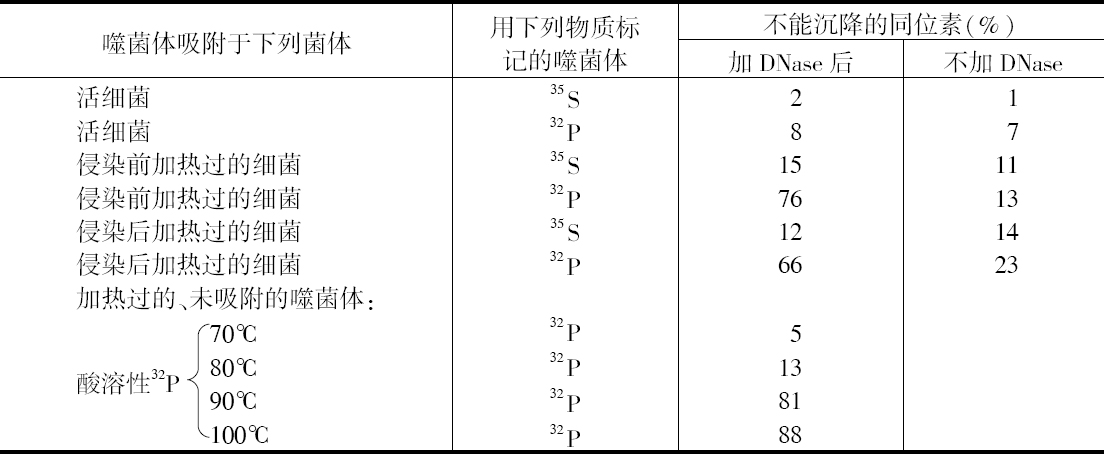
①在吸附培养基中,37℃ 5分钟,使噬菌体吸附到细菌上去,随后就洗涤。
细菌在吸附培养基中(侵染前)或佛罗那缓冲液中(侵染后),80℃热处理10分钟。
不能吸附的噬菌体是在佛罗那缓冲液中加热,并用DNase处理,用TCA沉淀。
分离供试样品,都是在1300×g离心10分钟。
噬菌体吸附到加热杀死的细菌上后,其DNA变得对DNase敏感了。这一现象首先是Graham和他的同事(私人通信)发现的。
通过冻融交替(随后用甲醛固定,使细胞的酶失活),可以使被侵染细胞中的DNA容易受到DNase的影响。只用甲醛固定也有一定作用,这可由下面的实验来说明。
将细菌培养在肉汤中达5×107 细菌/毫升,离心,再悬浮在吸附培养基中,每个细菌用大约2个32 P标记过的噬菌体来侵染。吸附5分钟后,用每升含有MgSO4 1.0mM;CaCl2 0.1mM;明胶10毫克的水溶液来稀释此悬浮液,再次离心收集菌体。菌体再悬浮在刚才提到的这种溶液中,浓度为5×108 个细菌/毫升。置于-15℃冰冻,而后用最小热量使之融化,这样冰冻、融化连续进行3次。第三次融化后,立刻加进0.5%(V/V)的甲醛(35%甲醛),将菌固定。在室温下30分钟后,透析除去甲醛,悬液在2200×g离心15分钟,用经过冻、融、固定、透析的32 P标记噬菌体和经过固定、透析的被侵染细菌作为对照。
分析表3给出的资料可以看出,冰冻和融化的效应是:使胞内的DNA对DNase不稳定了,而不是使大量的DNA滤出细胞。冻融和甲醛固定对未被吸附的噬菌体没有什么影响,单独的甲醛固定对于被侵染过的细胞只有温和的影响。
表3 冻、融和甲醛固定使胞内噬菌体变得对DNase敏感了①

①数字表示在原来的噬菌体或其被吸附的部分中,32 P的百分数。
胞内的32 P变得对DNase敏感,以及DNA不透出细胞,这两点都是这类试验的恒定特性,和见到的溶菌现象无关。在刚才叙述过的实验中,冰冻过的悬液在透析过程中变清了。相差显微镜检查看到:这种细胞基本上都是空的,显然大多数都破裂了。另一个实验中,在低盐肉汤中培养的细菌,经噬菌体侵染后,在噬菌体生长潜伏期的不同时间,经过反复冻、融,并用甲醛固定,然后离心洗涤。清楚可见的溶菌作用和在显微镜下才能看到的溶菌作用,只发生在潜伏期后半期的冰冻菌悬液中,在第一次或第二次融化时,从菌悬液中都可以看到这种现象;在这种情况下,溶解的细胞全是由完整的外膜组成的,除了附着在细胞壁上的几个小的、相当有特色的折射体外,其他显然都是空的。从表3看出,不论在溶菌或不溶菌的细胞中,胞内的32 P对DNase的行为都没有很大差异,只是溶菌之后,32 P的含量稍有降低。本实验中,对冻融期间释放出来的噬菌体的滴度,也进行了测定。在噬菌体侵染后,第16分钟及16分钟以前冰冻的菌悬液,溶菌时基本上不释放噬菌体;第20分钟冰冻的菌悬液,溶菌时每个细菌只产生5个噬菌体;第30分钟用甲醛处理的菌悬液,在常温下离心,不能沉降的部分中含有66%的32 P;第30分钟冰冻的菌悬液,每个细菌产生的胞外噬菌体是108个,沉降下来的物质大部分是不成型的碎片,但也含有很多完整的外膜。
用32 P标记的噬菌体侵染细菌,随后进行冻融处理,用这样的菌体所做的实验可以得出如下结论:
(1)噬菌体吸附到细菌上去之后,在不引起生长的缓冲液中,噬菌体DNA会变得对DNase敏感了(Benzer 1952;Dulbecco 1952)。
(2)在不允许胞内32 P或细胞内含物跑出去的条件下,外膜可变得对DNase有透性。
(3)即使由于冻融造成细胞溶菌,细胞的其他成分逸出胞外,但由噬菌体带进来的32 P大部分仍留在外膜内,形成成熟的子代噬菌体。
(4)在发生自发溶菌时,随着噬菌体的释放,由噬菌体带进细胞的32 P也大部分释放出去了。
我们对这些事实所做的解释是:从噬菌体带进细菌细胞内的DNA,不单单是能在溶液中存在,而且在整个潜伏期中都是组织结构的一部分。
噬菌体吸附到细菌的碎片上后,其DNA从噬菌体颗粒中释放出来
噬菌体吸附到细菌上去以后,其DNA对特异的解聚酶变得敏感了,这可能意味着在吸附后,噬菌体DNA就从其保护外壳中排出。下面的实验证明:实验中,当噬菌体吸附到破碎的细菌细胞上去时,确实会发生这种情况。
在吸附培养基中,每个细菌用4个T2噬菌体颗粒进行侵染,来制备细菌碎片。在37℃,把这些细胞转到低盐肉汤中,培养物通气培养60分钟,加入0.02 M HCN,继续保温培养30分钟以上,这时,胞外噬菌体产量可以达到每个细菌有400个颗粒,只是因为电解质的浓度低,所以不能吸附。溶解细胞的碎片在1700×g离心洗涤,再悬浮在吸附培养基中,使其浓度相当于3×109 个溶解细胞/毫升。这种悬液主要含有崩溃的、破碎外膜。同位素标记的噬菌体吸附到这些材料上面去的情况见表4。下面几点值得注意。
表4 噬菌体吸附到细胞碎片上后,其DNA就释放出来①
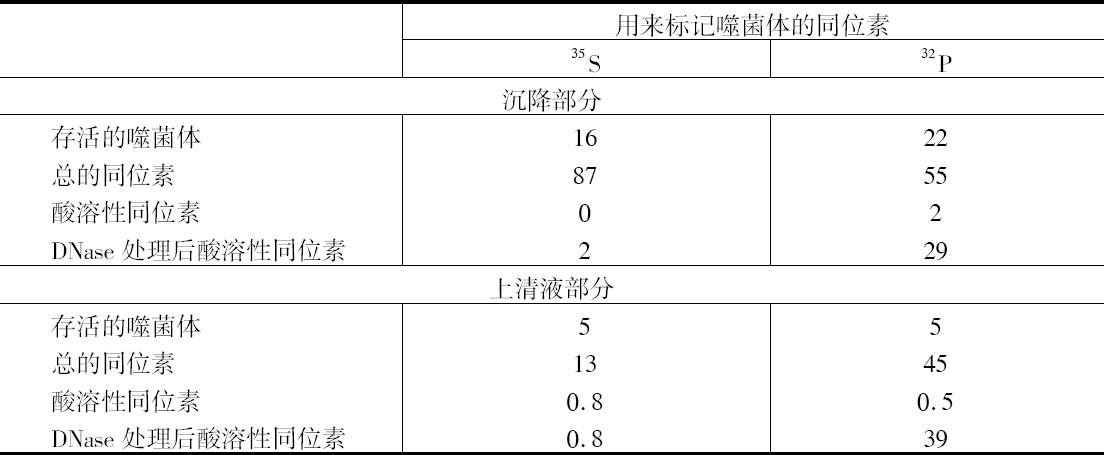
①35 S和32 P标记过的T2噬菌体与相应的细胞碎片样品,在吸附培养基中混合,在37℃保温30分钟,混合物在2200×g离心15分钟,然后对沉淀和上清液部分分别进行分析,结果用进入细胞碎片的噬菌体或同位素的百分数来表示。
(1)在未吸附部分中,只含有5%原来的侵染型噬菌体颗粒,占13%的总硫量(这些硫多数是在不能被完整的细菌吸附的物质中)。
(2)约有80%的噬菌体失活了。这种噬菌体的大部分硫,以及大部分存活的噬菌体是在沉淀中。
(3)上清液中除了含有未被吸附的存活噬菌体外,还含有总噬菌体DNA的40%(以对DNase敏感的形式存在)。这种对DNase敏感的DNA,数量约为失活噬菌体颗粒的DNA数量的一半,它们的硫与细菌碎片一起沉降。
(4)大多数可沉淀的DNA可能是存活的噬菌体,也可能是对DNase敏感的DNA,后者的数量约相当于失活颗粒DNA的一半。
这类实验有一个方面是不能令人满意的,人们不清楚,释放出来的DNA是代表失活颗粒的全部DNA呢?还是只是其中的一部分?
当细菌(菌株B)用大量的、紫外线杀死的噬菌体T2或T4溶菌后,再用32 P标记的T2和T4来测定,也得到了同样的结果。这个实验中,最有意义的一点是:用紫外线杀死的T2饱和的细菌碎片,吸附T4要比吸附T2好;用T4饱和的细菌碎片吸附T2要比吸附T4好。和前面的实验一样,某些噬菌体在吸附之后并不失去活性,某些失活噬菌体的DNA也没有从这些细菌碎片中释放出来。
这些实验表明:细菌对T2的某些接受器和对T4的某些接受器是不同的。而且,吸附到这些特异性的接受器上的噬菌体和吸附到非选择性接受器上的噬菌体的失活机制是一样的。显然,这种机制是很有效的,它的作用不单单是封阻噬菌体吸附到细菌上去的位点,可能还有其他作用。
从受侵染的细菌上去掉噬菌体外壳
Anderson(1951)得到的电子显微照片表明:噬菌体T2靠它的尾部吸附到细菌上去。假定在侵染过程中,这种不稳定的吸附受到保护;再假定上面得到的结论是正确的,那么噬菌体把DNA注入细菌细胞内,空的噬菌体外壳断离受侵染细胞,应该是一件简单的事。
下面的实验表明,用很强的、就像剪刀剪东西般的力量作用于受侵染的细菌悬液,就很容易从受侵染菌体上去掉噬菌体外壳;而且实验还说明:去掉了亲本病毒80%的硫的受侵染细胞,仍能形成子代噬菌体。
把在肉汤中培养的细菌,放在吸附培养基中用35 S或32 P标记的噬菌体进行侵染,离心除去未被吸附的噬菌体,把菌重新悬浮在含有下列物质的水中:MgSO4 1mM;CaCl2 0.1mM;明胶0.1克;水1升。悬液放在Waring捣碎器(半微量)中搅拌(10000rpm)。搅拌时,每隔60秒,把悬液放在冰水中冷却一会儿,隔一定时间取样,用抗噬菌体血清进行滴定,测出细菌产生的噬菌体数目,并离心测定细胞释放出的同位素的比例。
图1表示用同位素所作的一次实验结果。被侵染细菌的35 S和存活率,是从同一实验得到的,从直接滴定法测出:在这个实验中加入的噬菌体与细菌的比例为0.28,细菌的总浓度为9.7×108 /毫升,其中受到侵染的为2.5×108 /毫升。用32 P标记噬菌体所作的实验与这个实验十分相似。从这些结果,会使人回想起Anderson(1949)的发现,即:快速搅拌菌悬液可以防止噬菌体吸附到细菌上。
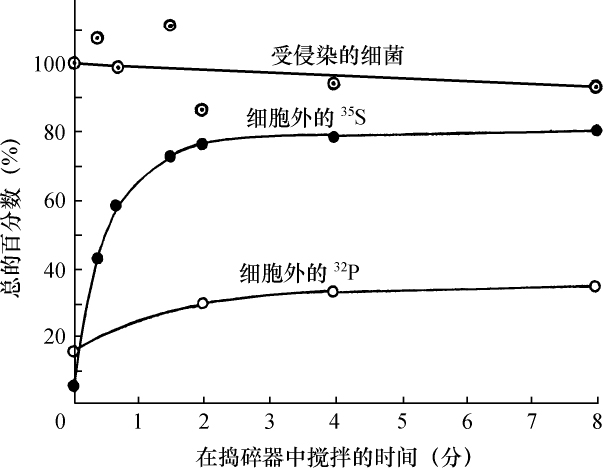
图1 在Waring捣碎器中搅拌时,同位素标记噬菌体侵染过的细菌中,35 S和32 P的去除以及受侵染细菌的存活曲线
虽然32 P的洗脱和受侵染细胞的存活不受侵染倍数的影响,但在这些实验条件下,当侵染倍数较高时,有相当数量的噬菌体硫从这些细胞中自发地洗脱下来(见表5)。这表明了各个噬菌体颗粒通过互相协作引起细菌细胞膜产生改变,从而减弱了噬菌体的吸附。用这种方法检查出来的这些细胞变化,可能与从被侵染的细菌中释放出细菌成分的那些变化是有关的(Prater,1951;Price,1952)。
表5 侵染倍数对从侵染细胞中洗脱噬菌体外壳的影响①
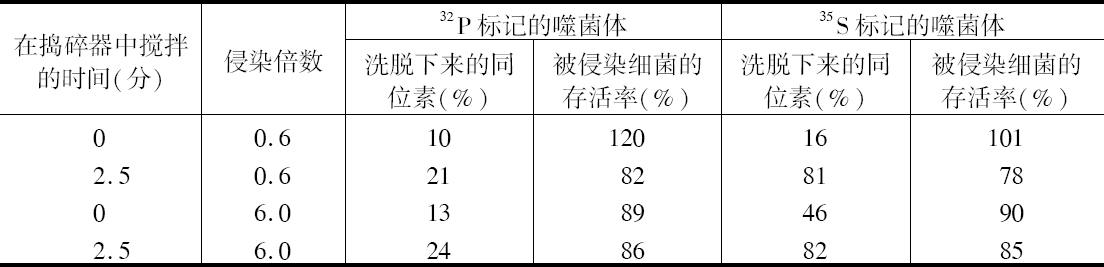
①被侵染细菌以109 /毫升的浓度悬浮在每升含有:MgSO4 1mM;CaCl2 0.1mM;明胶0.1克的水溶液中。取样,分析胞外同位素及悬液在搅拌前后被侵染细胞的数目。在这两种情况下,细胞都要在洗脱液中停留15分钟(室温)。
为了测定在噬菌体生长后期细菌的情况,对以前的实验作了某些改动。从这个目的出发,把被侵染细菌在肉汤中通气培养5或15分钟,加入0.5%(V/V)的商品甲醛进行固定、离心,再次悬浮于0.1%的甲醛水溶液中,其后的处理同以前所述。结果与以前十分相似,只是从细菌中释放出来的32 P稍微少些,受侵染细胞的滴度没有测定。
按照前面叙述的方法,从被侵染细菌剥离下来的这种35 S标记的物质,具有下列性质:在12000×g离心时,它沉降得不如完整的噬菌体颗粒那么完全;在用完整噬菌体做载体时,它能全部地被抗噬菌体血清沉淀;在37℃5分钟,它的40%~50%能够再次吸附到敏感细菌上去,当细菌浓度为2×108 ~2×109 /毫升时,它的吸附几乎与细菌浓度无关,这种吸附不是十分特异的,在同样条件下,有10%~25%能吸附到抗噬菌体的细菌上去。吸附作用需要盐,为此,只要在一种缺乏电解质的液体中,便可以从被侵染细菌上有效地除去35 S。
这些实验的结果总结如下:
(1)激烈振荡悬液可以把噬菌体75%~80%的硫从受侵染细胞上剥落下来。在侵染倍数高时,大约有50%的35 S能自发的洗脱下来。35 S标记材料的这种性质表明,它在不同程度上含有完整的噬菌体外壳。这些35 S标记的材料,大部分已失去特异性地吸附到细菌上去的能力。
(2)在这些硫释放出来的同时,噬菌体磷的释放只有21%~35%,其中半数在不用机械搅拌的情况下就可自发释放出来。
(3)实验所用的处理不会造成胞内噬菌体的明显失活。
(4)这些事实表明:在侵染过程中,噬菌体所含的硫,大部分仍留在细菌的表面,不参与胞内噬菌体的复制。相反,噬菌体的大部分DNA,在噬菌体吸附到细菌上后,很快就进入细胞。
硫和磷从亲代噬菌体向子代噬菌体的传递
上面我们已经推断:静止期噬菌体颗粒蛋白质所含的大部分硫不参与噬菌体的复制,事实上也不进入细菌。因此,亲本噬菌体的硫也不可能传递给子代。下述实验说明这个推测是正确的,这种传递的最大值不超过1%。
把细菌在甘油-乳酸盐培养基中培养过夜,然后再在同种培养基中,在37℃继代培养2小时,接种量调到能使继代培养物产生2×108 个细菌/毫升。把细菌沉淀下来,再悬于吸附培养基中(浓度为109 细菌/毫升),再用35 S标记的噬菌体T2进行侵染。37℃5分钟后,用2倍体积的水来稀释悬浮液,离心,除去未被吸附的噬菌体(占滴度的5%~10%)和35 S(约15%)。随后把细菌悬浮在甘油-乳酸盐培养基中,浓度为2×108 /毫升,37℃通气培养。在指定时间,快速连续地加入0.02mM HCN和2×1011 /毫升紫外线杀死的噬菌体,停止噬菌体的生长。氰化物能停止胞内噬菌体的成熟(Doermann,1948),紫外线杀死的噬菌体能吸附到细胞碎片上,使子代噬菌体损失减到最小,并促进细菌的溶解(Maaløe & Watson,1951)。正如在别处提到的,在这些实验中也注意到了,为了防止由于吸附到细菌碎片上造成子代噬菌体失活,溶菌噬菌体与经过增殖的噬菌体必须有密切关系[例如:在这种情况下应该用T2H(T2的h突变体)或T2L,但不应该用T4或T6]。
为了得到通常所说的噬菌体的最高产量,把被侵染细菌放在培养基中培养25分钟后,再加入溶菌噬菌体,而氰化物是在第二小时末尾加入。在这些条件下,侵染细胞的溶菌作用是相当慢的。
加入氰化物时立即停止通气,培养物在37℃放置过夜。然后经过离心分离把溶菌产物分离成以下几部分:第一次低速离心(2500×g,20分钟)沉淀物;高速离心(12000×g,30分钟)上清液;高速离心沉淀物重悬在吸附培养基中,再进行低速离心,得到第二次低速离心的沉淀和澄清了的高速离心沉淀。
表6表示这种类型的3种培养物的各部分中,35 S和噬菌体的分布情况。这些结果是在肉汤培养基和在甘油-乳酸盐培养基中得到的溶菌产物的代表性结果。
表6 用35 S标记的T2侵染细菌后,溶菌产物经离心分离得到的各部分中,噬菌体和35 S的分布百分比①
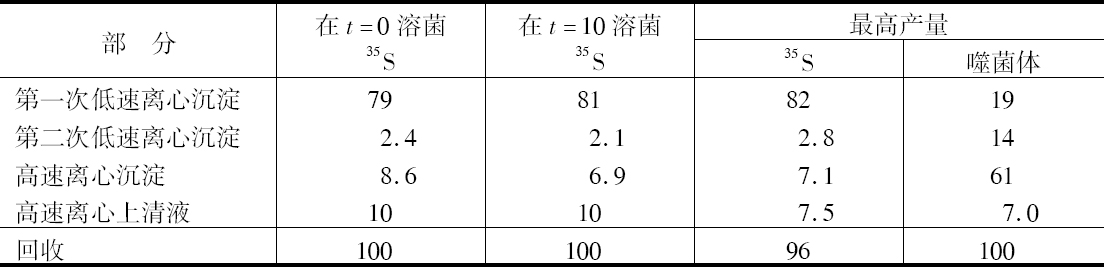
①用35 S标记的噬菌体T2侵染时,每个细菌平均为0.8个噬菌体颗粒。溶菌噬菌体是紫外线杀死的T2的h突变体。在t=0溶菌后,每个受侵染的细菌产生的噬菌体数量<0.1;在t=10时,溶菌后,则为0.12;最大值为29。35 S的回收指的是在4个部分中,回收量和吸附了的加入量的百分比;噬菌体的回收指的是根据各部分滴度来测定回收噬菌体量和总噬菌体量(分离前用噬菌斑计数)的百分比。
这个实验的显著结果是:不带有子代噬菌体的早期溶菌产物和含有子代噬菌体的晚期溶菌产物各部分中35 S的分布是一样的。这说明在成熟的子代噬菌体中几乎不含35 S。通过子代噬菌体吸附到细菌上去所起到的进一步分离作用,更证实了这一说法。
为此目的制备的吸附混合物,每毫升吸附培养基中含有:18小时肉汤培养物经热处理(70℃,10分钟)杀死的细菌约5×109 个,噬菌体约1011 个(紫外线杀死的溶菌噬菌体和测定噬菌体的总量)。37℃加热5分钟后,混合物用2倍体积的水稀释,并离心。对上清液和未经洗涤重新悬浮的沉淀分别进行分析。
表7表示35 S和噬菌体吸附到下面几种细菌上去的试验结果:①既能吸附T2子代噬菌体也能吸附h突变体溶菌噬菌体的细菌(H);②只能吸附溶菌噬菌体的细菌(B/2);③两种噬菌体都不能吸附的细菌(B/2h)。用35 S标记噬菌体做出可靠平行试验的结果也列于表7中。
表7 用统一的、35 S标记的噬菌体,以及它们在不含同位素培养基上的培养物所做的吸附试验①
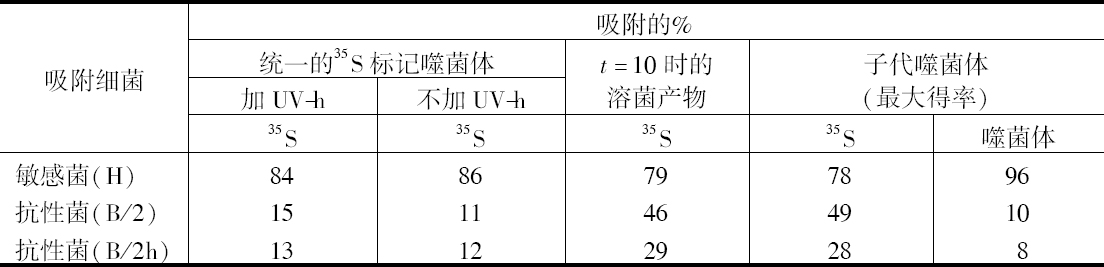
①统一的标记噬菌体和由它们得到的培养物,分别是种子噬菌体和表6实验的高速沉淀部分。测定统一的标记噬菌体是在噬菌体和细菌的比例较低的情况下进行的。加UV-h意思是加入紫外线杀死的h突变体(浓度和在其他试验中是相同的)。按常规方法,噬菌体的吸附作用是根据上消液噬菌斑计数来测定的;在抗性细菌的情况下,也用沉淀物噬菌斑计数来测定。
吸附试验表明:接种噬菌体中的35 S与这种噬菌体特异地结合在一起,而这种噬菌体侵染过的细菌溶菌产物中的35 S,表现出的行为更为复杂。它与既能吸附子代噬菌体也能吸附溶菌噬菌体的细菌牢牢地结合在一起;但与不能吸附这种噬菌体的细菌则结合得很微弱;它与能吸附溶菌噬菌体而不吸附子代噬菌体的细菌的结合情况,居于前二者中间。后面的测定表明:在子代噬菌体中没有35 S,而且还说明在早期溶菌产物中的35 S没有子代噬菌体的行为。
污染有35 S标记物质的子代噬菌体吸附作用的特异性,显然是由于溶菌噬菌体造成的,这种溶菌噬菌体吸附到菌株H上要比吸附到B/2上强烈得多,这可以从对它们各自的测定以及混合物上清液的Tyndall散射明显降低(由于溶菌噬菌体引起)看出来。下列事实进一步证实了这个结论。
(1)如果细菌经35 S标记噬菌体侵染,然后在潜伏期的中点附近只用氰化物溶菌(在低盐肉汤中,防止35 S再吸附到细菌碎片上去),这时高速沉降部分含有的35 S,只能微弱地、非特异性地吸附到细菌上去。
(2)假定溶菌噬菌体和35 S标记的侵染噬菌体是相同的(T2),或者假定在低盐肉汤中的这种培养物能自发溶菌(因此能得到大量的后代),这样,在高速沉淀物中的35 S与子代噬菌体就能特异性地结合在一起(微弱的非特异性吸附除外)。表7中吸附到H和B/2h上去的情况说明了这一点。
必须注意:一个从35 S标记噬菌体培养得到的、而且或多或少有放射性的子代噬菌体与一个真正的35 S标记的噬菌体,用已知的方法是不能区别的,只有通过吸附到抗噬菌体的细菌上去,才能把这种少量的污染物除去。除了已经提到的这些性质外,污染的35 S能与噬菌体一起被抗血清完全沉淀,不管在高浓度还是低浓度电解质中,用进一步分离沉淀的方法不能把它和噬菌体分离开。另一方面,这种来源的化学污染,在适宜环境条件下是很小的,因为一个噬菌体颗粒产生的子代是很多的,而污染物显然只能来自亲本。
35 S标记污染物的性质表明,它是由亲本噬菌体颗粒外壳的残留物组成的,推测它和在Waring捣碎器中能够从未溶菌细胞上除去的物质是一样的。它没有发生化学变化,这是不足为奇的,因为它根本就没进入被侵染的细胞。
叙述的这些性质说明,最初有关35 S从亲本噬菌体传递到子代噬菌体的报告(Hershey等,1951)是错误的。
用32 P标记的噬菌体做了和表6、表7所示相似的实验,实验结果表明:有30%的磷从亲代噬菌体传递到子代噬菌体中去了,每个受侵染细菌产生约30个噬菌体,在过早溶菌的培养物中,32 P几乎全部都是不能沉淀的,而在成熟后溶菌的培养物中,32 P事实上变成酸溶性的了。
Putnam和Kozloff(1950)等人也发表了有关32 P转移的类似测定结果。Watson和Maaløe(1952)总结了这项工作,并报道了磷和腺嘌呤的转移相等(接近50%)。
35 S标记的子代噬菌体几乎没有其亲本的标记
下列实验清楚表明:从亲代噬菌体传递给子代噬菌体的硫低于1%,还可能更少。作实验的时候,在一个Waring捣碎器中,将35 S标记的噬菌体外壳剥落下来,再从这样的受侵染的细菌获得噬菌体,直接测定其35 S含量。
在肉汤中培养的敏感菌,每个菌用5个35 S标记的噬菌体颗粒来侵染,从分析目的出发,必须采用这种高比例的侵染。侵染了的细菌,去掉吸附的噬菌体,再悬在每升含有MgSO4 1mM、CaCl2 0.1mM、明胶1克的水溶液中。从这种悬液取样,第一个样品在Waring捣碎器中搅拌2.5分钟,离心除去胞外的35 S。第二个样品放在捣碎器中,但不搅拌,在同样的时间也离心。两个样品的细菌都重悬在保温的低盐肉汤中(浓度为108 个细菌/毫升),通气培养80分钟。然后每毫升培养物中加入HCN 0.02mM、紫外线杀死的T2噬菌体2×1011 个、NaCl 6毫克进行溶菌。在这时加入盐,使35 S不容易被洗脱(Hershey等1951),仍旧吸附在细菌碎片上。按前面叙述的方法分离和测定溶菌产物,结果见表8。
表8 用35 S标记的T2噬菌体侵染过的细菌和在Waring捣碎器中再进一步剥落噬菌体外壳的细菌的溶菌产物①
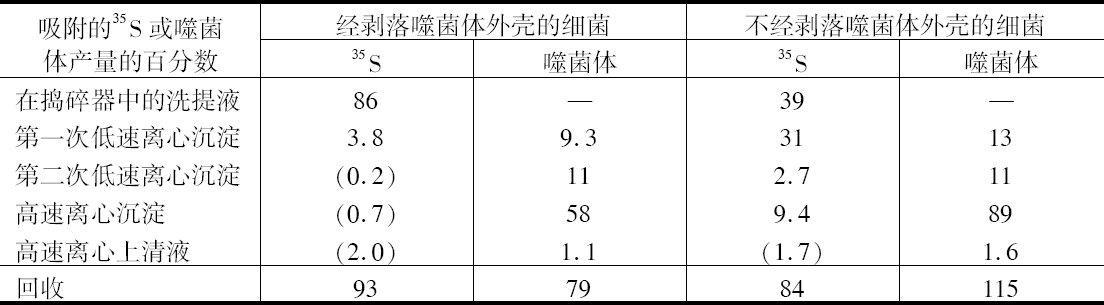
①在这两种培养物的潜伏期测定受侵染细胞时,回收了全部加入的细菌。在离心分离前测出每个细菌产生的噬菌体产量,分别是270(剥落了噬菌体外壳的细菌)和200个。括号中的数字是从接近于本底的计算比率得到的。
这些资料表明:剥落噬菌体外壳,能在不同程度上降低各个部分中35 S含量,尤其是含有大量子代噬菌体的部分降低最为明显,从最初所吸附同位素的大约10%降低到1%以下。这个实验说明:在溶菌产物各个部分中的35 S,都是来自保留在亲代噬菌体颗粒外壳中的35 S。
用甲醛失活的噬菌体的性质
噬菌体T2在含有0.1%(V/V)商品甲醛(35%的HCHO)的吸附培养基中,37℃保温1小时,然后透析除掉甲醛,这时噬菌斑滴度会降低1000倍或更多。这种失活的噬菌体具有下列性质:
(1)这种噬菌体约有70%能被吸附到敏感菌上去(这可用35 S或32 P标记来测定)。
(2)这种吸附的噬菌体,杀死细菌的效能只有原来的噬菌体原种的35%。
(3)这种失活噬菌体颗粒的DNA,在噬菌体吸附到热处理杀死的细菌上去后,对DNase仍然不敏感,在噬菌体吸附到细菌碎片上去后,不能被释放到溶液中去。
(4)在Waring捣碎器中搅拌,可以使70%吸附到细菌上去的噬菌体DNA与被侵染的细胞分离开。分离下来的DNA几乎全部抗DNase。
这些性质表明:用甲醛失活的噬菌体T2,大部分不能将它的DNA注入它所吸附的细胞中去。在概述的这个实验中,失活噬菌体的这些行为,有力地支持了我们对用活噬菌体做的有关实验所作的解释。
讨论
我们已经指出,当一个T2噬菌体颗粒吸附到细菌细胞上去后,它的大部分DNA进入细胞,留在细菌细胞表面的残留物含有噬菌体的含硫蛋白质80%以上。这种残留物是由形成静止噬菌体颗粒保护膜的物质所组成,在噬菌体吸附到细菌上后,在侵染中它没有进一步的作用。
还有20%含硫蛋白质,可能进入细胞,也可能不进入细胞,至于它们的作用是什么?还有待研究。我们发现:它几乎不参入侵染性子代噬菌体的颗粒,至少它的一部分可能是留在细胞外面的另一种残留物。另一方面,从侵染性噬菌体颗粒DNA得来的磷和腺嘌呤(Watson和Maaløe,1952),则以相当大的量、以同等程度传递给子代噬菌体。我们推断:含硫的蛋白质在噬菌体增殖中不起作用,而DNA有些作用。
现在回想起来,下列问题还没弄清楚:
(1)除了DNA之外,还有其他种类的不含硫的噬菌体物质进入细胞吗?
(2)假定有的话,那么它传递给子代噬菌体吗?
(3)磷(也许还有其他物质)传递给子代是直接的还是间接的?所谓直接的,就是在任何时间,它都保持着一种特异的、和噬菌体物质一样的形式。
我们的实验清楚地表明:用物理方法把噬菌体T2分离成遗传的和非遗传的两部分,这是可能的。在同一种噬菌体中,表现型和基因型具有一定独立性,由此可以看出,在相应的功能上也有所分工(Novick和Szilard,1951;Hershey等,1951)。然而,遗传部分的化学鉴定必须等上面提的一些问题得到回答后才能实现。
对于解决病毒培养问题的免疫学方法具有重大意义的两个事实,在这里必须强调一下,第一,在被侵染细胞中,T2噬菌体侵染性颗粒的主要抗原保持不变。第二,这种主要抗原依然吸附在细胞溶解形成的碎片上。Rountree(1951)在对T5噬菌体生长期间的病毒抗原进行研究的过程中,曾经预计过这些可能性。
摘要
(1)渗透压骤变使T2噬菌体颗粒分解出一种物质,它含有几乎全部的噬菌体硫,它能被抗噬菌体血清沉淀,还能特异地吸附到细菌上去,它向溶液中释放出几乎全部的噬菌体DNA,这种噬菌体DNA不能被抗血清沉淀,不能吸附到细菌上去。噬菌体颗粒的含硫蛋白质显然是构成一种膜,这种膜保护着噬菌体DNA免遭DNase破坏,它含有单一的或主要的抗原物质,它担负着使噬菌体吸附到细菌上去的责任。
(2)T2吸附到热处理杀死的细菌上去、受侵染的细胞经过加热或冻融交替处理,都可能使吸附了的噬菌体DNA变得对DNase敏感。这些处理不能使未吸附的噬菌体DNA变得对DNase敏感。虽然加热或冻融交替处理能从受侵染的细胞中提取出其他细胞成分,但不能使细胞释放出噬菌体DNA。这些事实说明:在整个噬菌体生长期间,噬菌体DNA成为组成细菌胞内结构的一部分。
(3)噬菌体T2吸附到细菌细胞碎片上去后,引起溶液中出现部分噬菌体DNA,留下的噬菌体硫吸附在碎片上。另一部分噬菌体DNA(大约相当于留下的失活噬菌体DNA的一半),仍然吸附在细胞碎片上,但可用DNase将它分离下来。尽管噬菌体T4和T2吸附位点不同,但噬菌体T4的行为与T2相似。噬菌体被细胞碎片失活,显然是和其外膜的破裂伴随发生的。
(4)在Waring捣碎器中搅动受侵染细菌细胞的悬浮液,因为受到强大的剪刀剪东西般力的结果,使75%的噬菌体硫和15%噬菌体磷释放到溶液中。这种细胞仍能产生子代噬菌体。
(5)列举的事实表明:在侵染时,噬菌体的大部分硫留在细菌表面上,噬菌体的大部分DNA进入细胞。除了DNA外,是否还有其他不含硫的物质进入细胞,尚不能肯定。从含硫残留物的性质可以看出:它主要是噬菌体颗粒未发生变化的外膜。各种类型的证据都证明:噬菌体DNA进入细胞只能在一定条件的非营养培养基中发生,在这种条件下,病毒生长的其他已知步骤都不发生。
(6)用放射性硫标记的噬菌体侵染过的细菌,产生的子代噬菌体所含的放射性,还不到亲本的1%。用放射性磷标记的噬菌体颗粒所产生的子代噬菌体所含的磷达到亲本的30%或30%以上。
(7)用稀甲醛失活的噬菌体能够吸附到细菌上去,但不能把它的DNA释放到细菌细胞中。这表明:噬菌体DNA从它的保护膜中释放出来是由于噬菌体和细菌之间的相互作用引起的,这种作用取决于噬菌体颗粒中的一些不稳定的成分。相反,对这种相互作用不可缺少的细菌成分则是十分稳定的。这种相互作用的性质在其他方面还不知道。
(8)静止期噬菌体颗粒的含硫蛋白质被证实是一种保护外壳,它担负着使噬菌体吸附到细菌上去的任务,还要起到一种能把噬菌体DNA注入细胞的装置的功能。这种蛋白质对胞内噬菌体的生长可能是没有作用的。而DNA却有一些作用。从提出的实验来看,还不能做出进一步的化学论断。
参考文献
[1]Anderson, T. F., 1949. The reactions of bacterial viruses with their host cells. Bot. Rev., 15 , 464.
[2]Anderson, T. F., 1951. Tr. New York Acad. Sc., 13 , 130.
[3]Anderson, T. F., and Doermann, A. H., 1952. J. Gen. Physiol., 35 , 657.
[4]Benzer, S., 1952. J. Bact., 63 , 59.
[5]Doermann, A. H., 1948. Carnegie Institution of Washington Yearbook, No. 47 , 176.
[6]Doermann, A. H., and Dissosway. C., 1949. Carnegie Institution of Washington Yearbook, No 48 , 170.
[7]Dulbecco,R., 1952. J. Bact., 63 , 209.
[8]Herriott, R, M., 1951. J. Bact., 61 , 752.
[9]Hershey, A. D., 1946. Genetics, 31 , 620.
[10]Hershey, A. D., Roesel., C., Chase, M., and Forman, S., 1951. Carnegie Institution of Washington Yearbook, No. 50, 195.
[11]Lwoff, A., and Gutmann, A,, 1950. Ann. Inst. Pasteur, 78 , 711.
[12]Maaløe, O., and Watson, J. D., 1951. Proc. Nat. Acad. Sc., 37 , 507.
[13]Novick, A., and Szilard, L., 1951. Science, 113 , 34.
[14]Prater, C. D., 1951. Thesis, University of Pennsylvania.
[15]Price, W. H., 1952. J. Gen. Physiol., 35 , 409.
[16]Putnam, F. W., and Kozloff, L., 1950. J. Biol. Chem., 182 , 243.
[17]Rountree, P. M., 1951. Brit. J. Exp. Path., 32 , 341.
[18]Watson, J. D., and Maaløe, O., 1952. Acta Path. et Microbiol. Scand., (in press).
核酸的分子结构 【64】
脱氧核糖核酸的结构
詹姆斯·沃森(J. D.Watson) 【65】 弗朗西斯·克里克(F. H.C. Crick)
(英国 剑桥卡文迪什实验室,1953年)
我们想提出脱氧核糖核酸(DNA)盐的一种结构,其特色是具有重要的生物学意义。
Pauling和Corey[1 ] 曾经提出过一种核酸结构模型,他们的模型是由3条缠绕的链组成,靠近轴线的是磷酸,碱基在外侧。我们认为这个结构不十分令人满意,原因有两点:
(1)从材料的X射线图像看来它属于盐类,而不是游离的酸。没有酸性的氢原子,那么又是什么力量使这个结构聚集在一起的呢?特别是靠近轴线而且带有负电荷的磷酸将会互相排斥。
(2)某些van der Waals距离看来似乎是太小了。
Fraser还提出了另一种3条链的结构。在他的模型中,磷酸是在外侧,碱基在内侧,并通过氢键连在一起。这个结构说得很含糊,因此,我们不评论它。
我们想对DNA盐提出一种根本不同的结构。在这种结构中,围绕着同一根轴线有两条螺旋状的链(见图1)。按通常的化学推测,每条链都是由磷酸二酯基团和β-D-脱氧呋喃核糖基团组成的,二者通过3′,5′键连接起来。两条链上的碱基,通过氢键形成碱基对,使两条链连在一起,碱基对的平面与轴线垂直。两条链均为右手旋转的螺旋,但由于成对排列的缘故,因此原子在两条链上的排列次序是反向的。每条链都有点像Furberg[2 ] 的模型No.1;即碱基在螺旋的内侧,磷酸在外侧。糖和靠近它的原子的构型与Furberg的“标准构型”很相似,糖与附着在糖上的碱基近于垂直。在每条链的“Z”形长轴方向上,每隔3.4埃,有一个碱基。我们推测,在同一条链上,相邻的碱基之间差36°角,所以,在每条链上,10个碱基后,也就是隔34埃,结构将重复一次。磷原子与轴线的距离是10埃。由于磷酸是在螺旋外侧,阳离子容易接近它们。
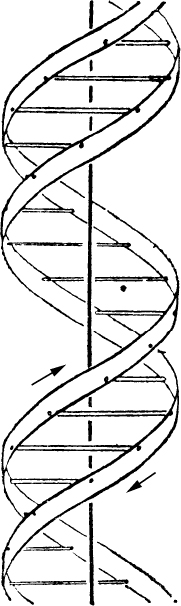
图1 DNA结构模式图
两条带表示两条糖-磷酸链,水平短线表示把两条链连在一起的碱基对。垂直线表示轴线。
由于这种结构是开放式的,因此含水量相当高;在含水量低时,预计碱基将倾斜,使结构变得更加致密。
这种结构的新特色是:两条链是由嘌呤和嘧啶碱基连在一起的。碱基的平面与轴线垂直,碱基成对地连在一起,一条链上的一个碱基与另一条链上的一个碱基,通过氢键连接起来,这样,在“Z”型的长轴方向上,两条链是紧靠着的。碱基配对时,必须一个是嘌呤,另一个是嘧啶。氢键组成如下:嘌呤位置1对嘧啶位置1;嘌呤位置6对嘧啶位置6。
假如在这种结构中,碱基只以通常的互变异构型式出现(即以酮式结构而不是烯醇式结构出现),那么只有特异的碱基可以结合在一起形成碱基对。这些碱基对是:腺嘌呤(A)与胸腺嘧啶(T),鸟嘌呤(G)与胞嘧啶(C)。
换句话说,如果腺嘌呤充当了一个碱基对中的一个成员,不管它是在哪条链上,按照这种假说,这个碱基对的另一个成员必定是胸腺嘧啶;对于鸟嘌呤和胞嘧啶来说也是同样的情况。在同一条链上,碱基的排列顺序可能不受任何限制。但是,如果只形成特异性的碱基对的话,当一条链上碱基的顺序定下来后,那么另一条链上的顺序便自动地决定了。
实验证明[3 -4 ] ,在DNA中,A/T的比值和G/C的比值总是十分接近于1。
用核糖来代替脱氧核糖就不可能得出这种结构,因为额外的氧原子将造成太紧密的van der Waals接触。
以前发表的有关DNA的X射线的资料[5 -6 ] ,对于严格测定我们的结构来说,尚不充足,我们现在只能说它与实验资料是一致的,但这种结构在未经确切验证之前,只能认为是推理性的。其中有些结果将在以后的文章中发表。然而,我们在设计这个结构时,并不知道那些结果的详细情况,这种结构的提出主要是(虽然不是全部)以发表过的实验资料和立体化学的证据为根据的。
在提出碱基特异性配对的看法后,我们立即又提出了遗传物质进行复制的一种可能机理。
整个结构的详细情况(包括推测它时所用的条件与原子的坐标位置)将一起在下面的一篇文章中发表。
参考文献
[1 ]Pauling, L., and corey, R. B., 1953. Nature, 171, 346; 1953 Proc. Nat. Acad. Sci. USA, 39 , 84.
[2 ]Furberg, S., 1952. Acta Chem. Scand., 6 , 634.
[3 ]Chargaff, E., for references see Zamenhof, S., Brawerman, G., and Chargaff, E., 1952. Biochim. et Biophys. Acta, 9 , 402.
[4 ]Wyatt. G. R., 1952. J. Gen. Physiol., 36 , 201.
[5 ]Astbury, W. T., 1947. Symp. Soc. Exp. Biol. 1, Nucleic Acid, 66(Camb. Univ. Press).
[6 ]Wilkins, M, H. F., and Randall, J. T., 1953. Biochim. et Biophys. Acta, 10 , 192.
脱氧核糖核酸的结构 【66】
詹姆斯·沃森(J. D.Watson) 弗朗西斯·克里克(F. H.C. Crick)
(英国 剑桥卡文迪什实验室,1953年)
在这本关于病毒的论文集中,收入这篇关于脱氧核糖核酸(DNA,下同)结构,讨论它对病毒繁殖的重要性方面的文章,似乎是多余的。但我们不仅认为DNA是很重要的,而且还认为DNA是病毒遗传特异性的载体(有关这方面的讨论,可参考本文集Hershey的文章),因此在某种意义上讲,DNA必然具有精确的自我复制能力。在本文中我们将叙述一种DNA结构,这种结构暗示了DNA自我复制的机制,并使我们能首次从原子水平上提出遗传物质自我复制的详细假说。
首先,我们讨论说明DNA是一条很长的纤维状分子的有关化学和物理化学方面的资料。其次,我们解释为什么结晶学证据说明DNA结构单位不是由一条而是两条多核苷酸链组成的。然后讨论我们认为能满意地解释化学和结晶学资料的一种立体化学模型。最后我们提出这种结构的某些明显的遗传学含义。对其中某些资料的初步看法已在英国《自然》杂志上发表了(Watson和Crick,1953a、b)。
说明DNA是纤维状的有关证据
DNA的基本化学式现在已经确定了。如图1所示,它是由两条很长的链构成的,它的主链是由糖和磷酸基团交替组成的,彼此以3′,5′磷酸二酯键连在一起。每个糖上结合着一个含氮碱基,通常在DNA中只有4种不同的碱基,其中两种是嘌呤,即腺嘌呤(A)和鸟嘌呤(G);另外两种是嘧啶,即胸腺嘧啶(T)和胞嘧啶(C)。在某些生物中,第五种碱基,即5-甲基胞嘧啶(MeC),也少量出现;第六种碱基,5-羟甲基胞嘧啶(HMC),在T系偶数列噬菌体中代替了胞嘧啶(Wyatt和Cohen;1952)。
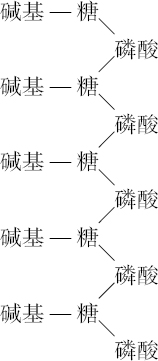
图1 一条DNA单链的化学式(图解)
应当指出,由于核苷酸之间的连接是有规律的,因此链是不分枝的。另一方面,就目前已经确定的情况而言,不同核苷酸的顺序完全无规律可循,因此,DNA的某些性质是有规律的,而另一些则是没有规律的。
从物理-化学分析,包括沉降、扩散、光散射,以及黏度等方面的测定,得出一个共同的概念,即DNA的分子是细长的纤维状结构。这些技术说明,DNA的结构是很不对称的,粗细大约为20埃,长度达数千埃,分子量估计在5×106 1×107 道尔顿(大约3×104 个核苷酸)。幸运的是各种测定都趋于说明DNA是相当牢固的,从糖-磷酸主链中含有大量单键来看(每个核苷酸有5个单键),这是一个令人费解的发现。最近这些间接推论被电子显微镜观察所证实了。Williams(1952)和Kahler等人(1953)采用高分辨技术,在DNA制品中都看到了细长的DNA纤维,这些纤维粗细均匀,大约是15~20埃。
说明DNA有两条链的证据
这类证据主要是来自X射线研究。所用的材料是DNA钠盐(DNA通常是从小牛胸腺提取的),经过纯化并拉成纤维,这些纤维是高度双折射的,表现出明显的紫外线和红外线的双色性(Wilkins等,1951;Fraser和Fraser,1951),而且能形成很好的X射线图像。Wilkins, Franklin以及他们在英国皇家学院的同事,根据这些初步研究,推导出关于DNA结构的某些一般性结论(Wilkins等,1953;Franklin和Gosling,1953a、b、c)。从他们的工作可见有两点是非常重要的:
DNA有两种不同形式
第一种是晶体型,即A型结构(见图2),在相对湿度为75%时出现这种结构,其含水量为30%。在湿度更高时,这些纤维吸收更多的水分,长度约增加30%,变成第二种结构,即B型结构(见图3)。B型结构没有A型结构那么整齐,可能是类晶体;也就是说各个分子都彼此平行地堆垒在一起,而不是以另外的方式有规律地排列在空间中。我们把区分这两种形式的某些特性列于表1。A、B间的转换是可逆的,因此这两种结构可能是以某种简单方式联系起来的。

图2 DNA A型结构的X射线图像
(H. M.F. Wilkins和H. R.Wilson没公开发表的资料。)
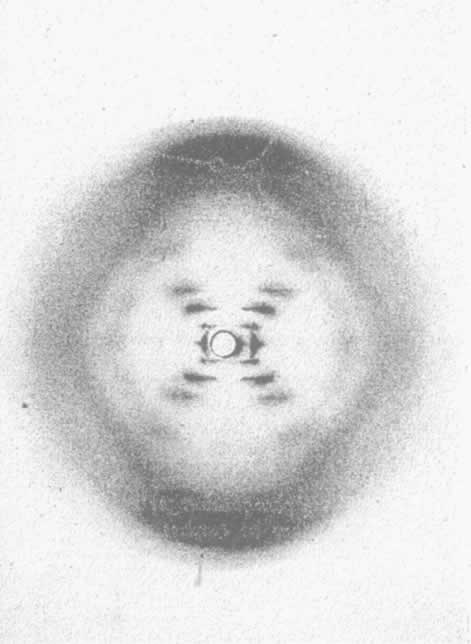
图3 DNA B型结构的X射线图像
(R. E.Franklin和R. Gosling,1953a.)
表 1

(引自Franklin和Gosling,1953a、b、c。)
晶体单位含有两条多核苷酸链
由于结晶学上有异议,因此只能给出一个概貌。B型结构在子午圈3.4埃处有一个很强的反射。正如Astburg(1947)最初指出的那样,这只能说明核苷酸基团在轴线方向上,每隔3.4埃的距离出现一次。从B型结构到A型结构,纤维缩短30%。这样,在A型结构中,核苷酸基团在轴线方向上每隔2.5埃 【67】 出现一次。根据A型结构的密度(Franklin和Gosling,1953c),以及相邻碱基对之间空间大小,可见每有这样一个基团,必有两个核苷酸。因此很可能,晶体单位是由两条不同的多核苷酸链构成的,对此要作最后证明,只能在这种结构完全弄清以后。
A型结构有一个拟六角形的格子,各角点相距22埃。这个距离大致相当于电子显微镜下看到的这些纤维的直径,这里应当指出,A型结构是很干燥的。这样看来,晶体单位和这种纤维很可能就是同一种东西。
对结构的描述
从上述资料可以得出两个有用的结论。第一,DNA的结构是很有规律的,足以形成一种三维晶体。在这一点上,没有考虑组成它的两条链可能会有不规则顺序的嘌呤和嘧啶核苷酸这个事实。第二,由于这种结构具有两条链,因此它们必定是彼此有规律地排列着。
考虑到这些发现,我们提出了一种结构(Watson和Crick,1953a),在这种结构中两条链围绕着一条共同的轴线缠绕,并通过核苷酸碱基之间的氢键彼此连接起来(见图4)。两条链都是右手旋转的螺旋,但原子在糖-磷主链上的顺序是反方向的,并成对地垂直于螺旋轴线。糖和磷酸基团在螺旋的外侧,而碱基在内侧。磷原子与轴线的距离是10埃。我们创建的模型相当于B型结构,B型结构的X射线资料表明,在轴线方向上每隔34埃结构就重复一次,在X射线图像的子午圈上3.4埃的空间处有一个非常强的反射。我们的结构中,在轴线方向上,每隔3.4埃有一个核苷酸,10个这样的间隔,也就是34埃后,完成一个整周,这与上述发现是一致的。我们的结构说得很清楚,所有的键距和角度,包括van der Waal距离,都是立体化学上可以接受的。
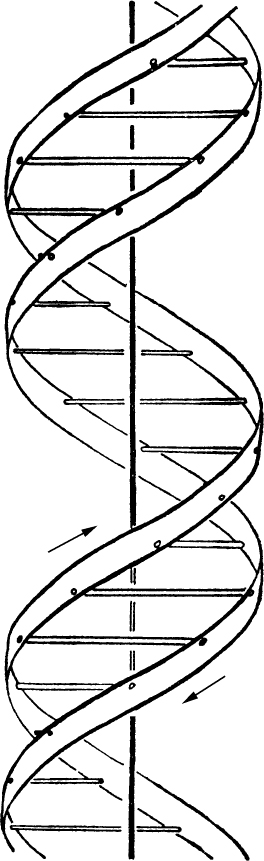
图4 DNA结构模式图
两条带表示两条糖-磷酸链,水平线表示连接两条链的碱基对,垂直线表示轴线。
我们这种结构的特色,是组成它的两条链通过碱基之间的氢键彼此连在一起。碱基与轴线垂直,并且成对地连在一起。这些碱基对的排列是非常特异的,而且只有某些碱基对能适合于这种结构,其主要原因就是这种结构中,每条多核苷酸链的主链都是一种有规律的螺旋形式。这样,不管是出现哪种碱基,配糖键(连接糖和碱基)都是以很规则的空间方式排列着的。特别是已经结合到一个碱基对上的任何两个配糖键(每条链一个),总是以固定的距离出现,这是因为它们连接的两条主链是有规律的。结果,一个碱基对中的一个成员总是一个嘌呤,另一个成员是一个嘧啶,只有这样才能在两条链之间搭成桥。例如,如果一个碱基对是由两个嘌呤组成,那么就会因占的空间太大而放不下;如果是由两个嘧啶组成,那么又会因相距太远而不能形成氢键。
在理论上讲,一个碱基能以多种互变异构形式存在,在各种形式下,结合氢原子的确切位置是不同的。但是在一定的物理条件下,对每种碱基来说,都有一种特殊形式比其他任何一种形式出现的可能性都大得多。假定,碱基总是以其常见形式出现,那么配对的要求是很局限的。A只能与T配对,G只能与C配对(或者与5-甲基胞嘧啶或与5-羟甲基胞嘧啶配对)。这种配对方式详细示于图5和图6。如果试图让A和C配对,就不会形成氢键,因为在一个键位上有两个氢原子,而在另一个键位上,由于被替代,所以一个氢原子也没有。
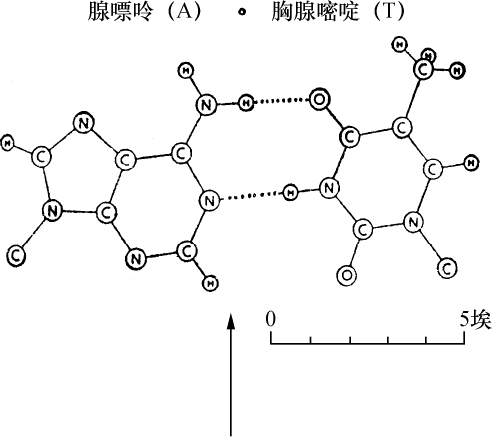
图5 A和T的配对
氢键用虚线表示。每个糖只表示出一个碳原子。
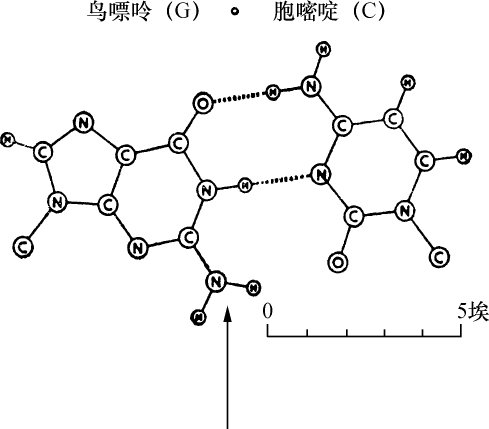
图6 G和C的配对 【68】
氢键用虚线表示。每个糖只表示出一个碳原子。
一个已知的碱基对能以任何一种方式来形成。例如,A可以出现在任何一条链上,但是,一旦它定下来了,那么在另一条链上和它配对的必定是T。这可能是因为一个碱基对中,两个糖苷键彼此是对称的(见图5、图6),因此,如这个碱基对颠倒过来,两个糖苷键就会出现在同一个位置上,因此不能配对。
应当强调指出,由于每一个碱基可以在几个位置上形成氢键,因此,分离出来的核苷酸可以通过多种方式配对,我们限制某些条件就可以得到特异性碱基对,在我们的模型中,这是直接由糖-磷主链的规律性所决定的。
还应强调的是,在这种DNA结构的某一个点上,不管碱基对是怎么产生的,对相邻的碱基对都没有限制作用,各种碱基对顺序都可能出现。这是因为所有碱基都是扁平的,一个堆垒在另一个上面,就像堆垒起来的铜钱一样,哪个靠近哪个是没有差别的。
虽然任何碱基顺序都适合于我们的结构,但是特异性碱基配对方式决定了两条链上碱基顺序有着明显的相关性。也就是说,如果我们知道了一条链上碱基的实际顺序,那么自动地就可以写出另一条链上碱基的顺序。因此,我们的结构是由两条彼此互补的链组成的。
支持这种互补模型的证据
有些实验证据为我们的结构提供了有力的支持,但应该强调指出,目前尚未证实这种结构是正确的。支持这种模型的证据有以下三类。
1.X射线图像的一般外形有力地说明这种基本结构是螺旋状的(Wilkins等,1953;Franklin和Gosling,1953a)
假定螺旋是存在的,我们马上就能从B型结构(见图3)的X射线图像推断出它的螺距是34埃,它的直径大约为20埃。而且这种图像还表明了在螺旋的周围原子密度很高,这与我们模型中糖-磷主链在外侧的看法是一致的;这种图像还说明了两条多核苷酸链不是沿着轴线留下相等空间,而是以纤维轴间距的3/8互相移位的,这种推论在性质上与我们的模型也是一致的。
对A型结构(晶体型)的X射线图像的解释不那么明了。这种构型在3.4埃处不产生子午圈反射,而在子午圈外25°周围,在3~4埃的空间处产生一系列的反射(见图2)。这向我们提示了在这种形式中,碱基不再与轴线垂直,而是从垂直位置倾斜大约25°,因此使纤维缩短30%,并使每个核苷酸的纵向平移减少到2.5埃左右。应当指出A型结构的X射线图像比B型结构的要详细得多,因此,如能正确解释,可能获得有关DNA的更确切资料。提出的任何一种DNA模型,它必须或者能相当于A型结构,或者能相当于B型结构。这进一步说明了我们对于A型结构所做的这些解释虽然是很粗略的,但仍是很必要的。
2.未经酸和碱降解的DNA有不规则的滴定曲线,有力地表明形成氢键是DNA结构的一个典型特性
如果用酸或碱处理一种DNA溶液,最初当pH5~11时,通过滴定检查不出有什么基团来,但超过此范围;溶液迅速发生电离(Gulland和Jordan,1947;Jordan,1951)。从pH12用酸回滴,或者从pH2.5用碱回滴,得出了不同的滴定曲线,这说明可滴定的基团比未处理的溶液更容易受到酸和碱的影响。在pH11.5和pH3.5~4.5的范围中,随着基团开始释放,黏度明显降低,强烈的流动双折射消失了。这种降低最初被认为是由于一种可逆的解聚过程引起的(Vilbrandt和Tennent,1943),Gulland、Jordan和Taylor(1947)曾指出,观察到次生磷酸基数量可能有所增加。他们指出碱基的某些基团在不同的碱基之间形成了氢键,但还不能确定连接碱基的氢键是在同一个结构单位上,还是在相邻的结构单位上。如果氢键是在同一结构单位内的碱基之间形成,那么,大多数可以电离的基团起初不受酸和碱的影响也就容易解释了。当相邻结构单位之间相互影响很小时,如果这种最初的滴定曲线与低浓度DNA的滴定曲线形状相同的话,那么就可以断定氢键是在同一个结构单位内的碱基间形成。
3.对各种碱基相对比例的分析资料表明,A与T的数量接近,G的数量与C+MeC的数量接近,而A与G的比例则随不同来源而有所变化(Chargaff,1951;Wyatt,1952)
实际上,碱基测定技术证明,A/T、G/C+5-MeC的比值都接近于1。这些结果是很明显的,尤其是当一条链上碱基顺序不规律的情况下就更明显,这说明碱基在结构中是配对的。这些分析资料完全支持DNA具有两条链这一具有生物学意义的特性,从而为支持我们的模型提供了非常重要、非常有用的证据。
因此,我们认为目前的实验证据证实了我们的工作假说(即我们模型的基本特性)是正确的,所以使我们能够考虑它的遗传学含意。
这种互补模型的遗传学含义
首先,我们应该说明,获得X射线图像的这种DNA纤维并不是由于制备方法引起的人工产物。第一,Wilkins及其同事(见Wilkins等,1953)曾指出,从这些分离出来的DNA纤维,与从某些完整的生物学材料(例如精子和噬菌体颗粒),所得出的X射线图像是相似的。第二,我们提出的模型是极其特异的,因此我们认为:它不可能是在从活细胞提取DNA的过程中形成的。
一种遗传物质,必须以某种方式行使两种功能,即自我复制和对细胞的高度特异性影响。我们的DNA模型为第一个过程提出了一种简单的机制,但目前还不知道它如何完成第二个过程。但我们相信,其特异性是由准确的碱基对顺序表达的。在我们模型中DNA的主链是非常有规律的,碱基顺序是能够携带遗传信息的唯一特性。不能认为,在我们的结构中,因为碱基在里面就不能与别的分子接触了。由于我们的结构是开放性的,实际上这些碱基很容易受到影响。
DNA复制的机制
我们结构的互补特性揭示了它是如何自我复制的。要想象出DNA如何自我复制,这是较困难的,曾有人提出(见Plauling和Delbrück,1940;Friedrich-Freksa,1940;Muller,1947)自我复制可能和各部分与其相对或互补部分的结合有关。在这些讨论中,普遍提到蛋白质和核酸是彼此互补的,自我复制过程与这两种成分的交替合成有关。相反,我们提出了实现特异的DNA自我复制不依赖于特异的蛋白质合成,我们模型中的每一条互补DNA链,在形成它的一条互补新链时,都可以作为模板或模型。
为了达到上述目的,连接互补链的氢键必须断裂,两条链还必须解缠和分离。很可能这种单链(或其适当部分)本身可以呈螺旋构型,并作为模板,游离的核苷酸(正确的多核苷酸前体)通过形成氢键自动地结合到它上面。我们认为,只有当新产生的链形成我们所提出的这种结构时,才能发生前体物形成新链的多聚化过程。由于空间排列的原因,除了适合于我们这种结构的单体外,晶格化的单体,不能进到第一条链上去彼此接近,因此就不能彼此连接形成新链。至于实现多聚化过程是需要一种特异的酶呢?还是单螺旋能像一种酶那样有效地起作用呢?现在还不清楚。
复制方式上的异议
当这种方式产生了吸引力后,虽然也出现了许多异议,但我们认为,没有一种能站住脚。第一种异议认为我们的结构没有区分C和5-MeC,因此在复制过程中,含有这些碱基的顺序之特异性是不持久的。虽然5-MeC通常数量很少或没有,但不同种间数量变化很大,最近的实验结果(Wyatt,1952)表明了每个种内都有特定的量。而且还表明了C+5-MeC的量比C的量更接近G的量。很可能这两种胞嘧啶在功能上没有明显差异。如果能够改变一种生物的DNA中5-MeC的量,而不改变DNA的遗传结构,那么就会大大支持上述解释。
5-HMC在T系偶数列噬菌体中的出现,说明不存在上述争议,因为5-HMC这时完全代替了C,它在DNA中的量与G相等。
对我们这种结构提出的第二种异议,就是认为它完全忽视了在大多数活的生物中与DNA结合在一起的碱性精蛋白、组蛋白和蛋白质的作用。我们之所以没有考虑这些蛋白质的作用,有以下两个理由。第一,我们可以系统地讲出DNA自我复制的方式,为了简便起见,最好相信(至少在目前)遗传特性的传递不经过一种蛋白质媒介。第二,关于精蛋白和组蛋白的结构特性,我们几乎一点不了解。我们仅有的线索就是Astbury(1947)及Wilkins和Randall(1953)的发现,即核精蛋白的X射线图像与DNA的X射线图像非常相似。这表明蛋白质——至少是某些蛋白质——也是螺旋构型的,鉴于我们的模型是一种开放性的,我们认为蛋白质可能在两条多核苷酸链之间形成第三条螺旋链(见图4)。至今对于蛋白质在其中的作用一无所知,也许它是控制着螺旋和非螺旋的构型,也许它是协助保持多核苷酸单链呈螺旋构形。
第三种异议就是为了作为一条新链的模板,两条互补链是否必须解缠。当两条链像在我们的模型中那样交织在一起的时候,这确实是一个主要异议。使两条螺旋能够一起旋转的两种主要方式被称为相缠螺旋和平行螺旋。细胞学家曾用这些术语描述过染色体的螺旋[Huskins,1941,可以看Manton(1950)的一篇评论文章]。我们模型中的螺旋叫做相缠螺旋。把两条分开的螺旋并排地放着,然后合在一起,使我们的轴心大致重合,这就是平行螺旋。要把两条有规律的螺旋合在一起,就必定会使它们发生相缠。用两条有规律的简单螺旋围绕同一轴心旋转,不可能得出平行螺旋来。这一点只有通过研究模型才能清楚的理解。
当然,一条展开的DNA单链卷曲成螺旋状这是不难的,因为一条多核苷酸链有很多单键,在单键处发生自转是完全可能的。具有共同轴心的两条单链组成的一条双链,要发生自转就比较困难了。这是一种拓扑学上的困难,而且不是通过简单的处理所能克服的。除了把链切断以外,还有两种方法可以把两条相缠卷曲的链分开。第一种方法是拿住一条链的一端和另一条链的另一端,在轴线方向上把它们拉开,两条链彼此滑过,最后从一端到另一端彼此分开。对我们的结构来说,可能不是这种情况,因此不再进一步去讨论它。第二种方法是两条链必须定向解缠。定向解缠后,两条链就相互分开了,但仍彼此紧靠着,使它们完全解缠的转数与一条链围绕着共同轴心旋转的转数相等。在我们的结构中,每34埃转一圈,每百万分子量的DNA大约转150圈,即相当于我们结构中5000埃的长度。对于解缠过程的疑问分成以下两类:①必须解缠的圈数是多少?解缠时如何避免发生混乱现象?②产生解缠的物理或化学力是什么?
目前,我们主要讨论第一个问题。要确定具有功能活性的DNA之连续长度是比较困难的。经过分离,我们所能得到的DNA分子量的下限,长度为50000埃,大约有1000圈。有证据说明,这仅是DNA纤维在提取过程中断裂的下限。上限在病毒中可能就是全部的DNA,在高等生物中可能就是一个染色体中的全部DNA。在T2噬菌体中这个上限约为800000埃,相当于20000圈,在高等生物中这个上限有时可高达1000倍。
如果一条染色体的连续部分是反向螺旋,那么上述疑问就很容易解决。最明显的方式可能在顺序上既有右手旋转的DNA螺旋,也有左手旋转的DNA螺旋,这与只有以右手旋转的螺旋才能建成我们模型的情况是不同的。另一种可能性是右手旋转的DNA长链被左手旋转的多肽螺旋的互补链连在一起。这种主张的优越性是难以估计的,但是,噬菌体DNA似乎与蛋白质不连在一起,这一事实又使这种主张不那么引人注意了。
如果两条链一分开,复制就在末端上开始,那么解缠过程就没那么复杂了。这种机制将形成一种新的双链结构,任何时候也不需要有一个游离的单链阶段。在这种方式中,像双链结构中发生紊乱的危险就大大地降低了,双链结构比单链结构牢固而坚硬,因此不容易围绕其相邻者形成螺旋。复制过程一旦开始,在两条链的延伸末端上,双链结构的出现,促进了尚未复制部位氢键的断裂,并使复制过程以类似开拉链的形式向前进行。
也有可能在旋转的扭力作用下,偶尔使两条链中某条链断裂,因此,那条完整的多核苷酸链就能松开,由于围绕单键转动而积累起来的缠绕,随后那些断裂了但仍很靠近的末端,又能重新连接起来。
虽然我们已经作了多种假设,但解缠仍然是一个不可忽视的难题,因此,对我们为什么提出是相缠螺旋而不是平行螺旋,进行重新检验还是很有必要的。在平行螺旋中,两条螺旋不是相互缠绕的,而只是彼此并列在一起。我们的回答是,在平行螺旋中,特异性的碱基配对不能使得每条螺旋的连续残基都保持对于螺旋轴线的一致方向。我们坚决反对这种可能性,还因为它暗示着对于糖-磷酸主链可能有许多空间化学的选择,这和我们关于空间化学模型的发现(Crick和Watson,1953)是矛盾的,在我们的模型中,糖-磷酸基团的位置是相当特定的,因此没有在平行螺旋中那样大的变化能力。而且平行螺旋构型不能产生特异性碱基配对,特异性碱基配对只有当配糖键在空间有规律地排列时才能发生。因此,我们认为,如果螺旋结构确实存在,那么两条螺旋之间的关系必定是相缠的。
但是,我们不禁要问,是否还有别的既保持了必需的规律性,但不是螺旋状的互补结构呢?事实上是有的,就是一条带状排列,在这条带状排列中,两条链也是通过特异性碱基配对连在一起,碱基对间相距3.4埃,但糖-磷酸主链不是形成螺旋,而是一条直线,它和碱基对形成的直线成30°角。这种带状结构与B型结构的X射线图像有许多相同之处,我们还不能确定它是怎样集合成显微镜下的纤维,特别是它为什么会在20~24埃处产生强烈的赤道反射?因此,虽然我们不能驳倒它,但我们对这种模型不热心。
有两个几何学问题,虽然与我们的模型关系不大,但这是任何一种DNA模型都会遇到的。这两个问题都与某种形式的超折叠过程的必要性有关,这可以用噬菌体来进行说明。首先,在噬菌体T2中,DNA总长度大约为8×105 埃,因此和其他来源的DNA一样,T2DNA的分子量很大(Siegal和Singer,1953),它必须来回弯曲许多次,才能装在直径仅为800埃的噬菌体头部。其次,DNA必须自我复制而不紊乱。平均大小为104 ×104 ×2×104 埃的一个细菌内,可以合成大约500个噬菌体颗粒。新形成的DNA总长度为4×108 埃,我们认为,所有这些新形成的DNA都以某种间隔与其亲本模板接触。无论复制机制多么精确,我们认为避免产生紊乱的最合理的方法就是把DNA折叠成它所形成的那种致密束。
自发突变的一种可能机制
在我们的复制图式中,复制的特异性是通过嘌呤和嘧啶碱基的特异性配对来实现的,即A与T配对,G与一种胞嘧啶配对;这种特异性是由下述假定引起的,假定碱基通常以一种稳定的互变异构形式出现,这种互变异构形式比其他任何一种可能的互变异构形式都稳定得多。但是,化合物是互变异构的,这一事实本身就说明氢键偶尔也可以改变其位置。以前我们认为自发突变是一种碱基顺序的改变,现在看来,很可能是因为在形成互补链时,一个碱基偶尔以其罕见的一种互变异构形式出现所造成的。例如,正常情况下,A与T配对,如果A的一个氢原子发生了互变异构移动,A就能与C配对(见图7)。下次配对时,这个A(已经恢复了其通常的互变异构型)将与T配对,而这个C将与G配对,这样,碱基的顺序就发生了一次改变。弄清在物理条件下,各种互变异构型之间自由能的确切差异,可能是非常有意义的。
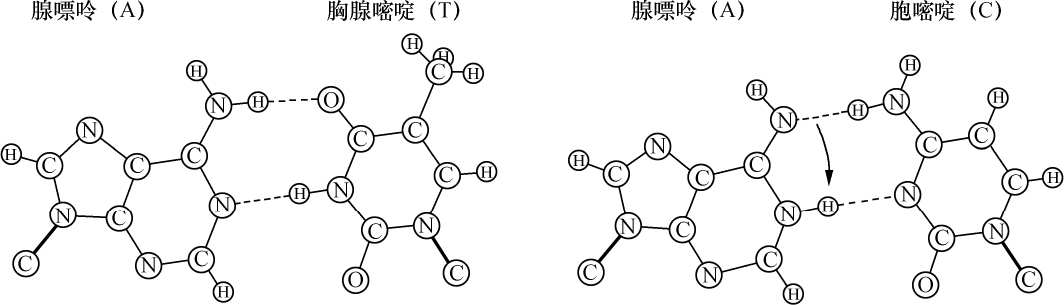
图7 A在发生一次互变异构变化前(左)后(右)的配对排列
一般性结论
通过进一步的结晶学分析,将会对我们的结构给予证明或反驳。我们希望这一任务能尽早完成。但是,如果这种互补链假说也被证明是错误的,那将是出乎意料的。我们原先是考虑了结晶学上的规律性,才提出这种特性来的。因此,如果认为这种特性与自我复制的密切联系是偶然的问题,我们认为这是不可能的。相反,这种相缠螺旋,至少从表面上看生物学吸引力不大,因此需要确切的结晶学证明。如果能确切证明遗传特异性只是携带在DNA上,并且阐明在分子水平上,这种结构如何对细胞行使特异性影响,那么无疑将对这种模型和所提出的复制机制提供更多的证据。
参考文献
[1]Astbury, W. T., 1947. X-Ray Studies of nucleic acids in tissues. Sym. Soc. Exp. Biol., 1 : 66~76.
[2]Chargaff, E., 1951. Structure and function of nucleic acids as cell constituents. Fed. Proc., 10 : 654~659.
[3]Crick, F. H. C., and Watson, J. D., 1953. Manuscript in preparation.
[4]Franklin, R. E., and Gosling, R., 1953a. molecular configuration. in sodium thymonucleate. Nature, Lond, 171 : 740~741.
1953b. Fiber diagrams of sodium thymonucleate, I. The influence of water content. Acta Cryst., Camb., (in press);
1953c. The structure of sodium thymonucleate fibers. II. The cylindrically symmetrical patterson function. Acta Cryst., Camb., (in press).
[5]Fraser. M. S., and Fraser, R. D. B., 1951. Evidence on the structure of desoxyribonucleic acid from measurements with polarized infra-red radiation. Nature, Lond., 167 : 760~761.
[6]Friedrich-freksa, H., 1940. Bei der Chromosomen Konjigation wirksame Krafte und ihre Bedeutung für die identische Verdoppling von Nucleoproteinen. Naturwissenshaften, 28 : 376~379.
[7]Gulland. J. M., and Jordan, D. O., 1946. The macromolecular behavior of nucleic acids. Sym. Soc. Exp. Biol., 1 : 56~65.
[8]Gulland, J. M., Jordan, D. O., and Taylor, H. F. W., 1947. Electrometric titration of the acidic and basic groups of the desoxypentose nucleic acid of calf thymus. J. Chem. Soc., 1131~1141.
[9]Huskins, C. L., 1941. The coiling of chromonemata, Cold Spr. Harb. Quant. Biol., 9 : 13~18.
[10]Jordan, D. O., 1951. Physiochemical properties of the nucleic acids. Prog. Biophys., 2 : 51~89.
[11]Kahler, H., and Lloyd, B. J., 1953. The electron microscopy of sodium desoxyribonucleate. Biochim. Biophys. Acta, 10 : 355~359.
[12]Manton, I., 1950. The spiral structure of chromosomes. Biol. Rev., 25 : 486~508.
[13]Muller, H. J., 1947. The Gene. Proc. Roy. Soc. Lond. Ser. B., 134 : 1~37.
[14]Pauling, L., and Dedbrück, M., 1940. The nature of the intermolecular forces operative in biological processes. Science, 92 : 77~79.
[15]Siegal, A., and Singer, S. J, 1953. The preparation and properties of desoxypentosenucleic acid. Biochim. Biophys. Acta, 10 : 311~319.
[16]Vilbrandt, C. F., and Tennent, H. G., 1943. The effect of pH changes upon some properties of sodium thymonucleate solutions. J. Amer. Chem. Soc., 63 : 1806~1809.
[17]Watson, J. D., and Crick, F. H. C., 1953a. A structure for desoxyribose nucleic acids. Nature, Lond. 171 : 737~738. 1953b. Genetical implications of the structure of desoxyribose nucleic acid. Nature, Lond., (in press).
[18]Wilkins, M. H. F., Gosling, R. G., and Seeds, W. E., 1951. Physical studies of nucleic acids—nucleic acid: an extensiblemolecule. Nature, Lond., 167 : 759~760.
[19]Wilkins, M. H. F., and Randall, J. T., 1953. Crystallinity in spermheads:molecular structure of nucleoprotein in vivo. Biochim. Biophys. Acta, 10 : 192 (1953).
[20]Wilkins, M. H. F., Stokes. A. R., Wilson, H. R. 1953.molecular structure of desoxypentose nucleic acids. Nature, Lond., 171 : 738~740.
[21]Williams, R. C., 1952. Electron microscopy of sodium desoxyribonucleate by use of a new freezedrying method. Biochim. Biophys. Acta, 9 : 237~239.
[22]Wyatt, G. R., 1952. Specificity in the composition of nucleic acids. In The Chemistry and Physiology of the Nucleus, 201~213. N. Y. Academic Press.
[23]Wyatt, G. R., and Cohen, S. S., 1952. A new pyrimidine base from bacteriophage nucleic acid, Nature, Lond., 170 : 1072.
基因 【69】
路易斯·斯塔德尔(L. J.Stadler) 【70】
(1954年)
生物学的普遍问题是生命物质的物理性质。正是这一点,它使基因的研究增添了动力和兴趣,因为研究基因物质的行为现在似乎是我们解决这一问题最直接的入门。
当前关于生命细胞行为的知识,有两点格外引人注目。第一,是生命细胞中发生的化学反应几乎难以置信的精密的平衡,通过这种平衡,能量得到供应,合成得以进行,从而提供了生长所需要的物质。第二,是基因物质的行为,它显然操纵着这些反应。基因被携带在染色体精细的绳索上,它们拼凑在一起只是细胞物质的一个微小的部分。这些染色体绳索沿着它们的长度分成数百个作用不同的节段,因此,这些节段大概有不同的组成,即我们所说的基因。基因物质在每一细胞世代都进行复制。在许多已知的情况下,它的有区别的节段决定是否会发生一种特殊的化学反应,大概,至少在某些情况下它决定一种特异催化剂的产生。
细胞物质的绝大多数显然是由上述所操纵的反应产生的物质组成。关于这些物质的性质和行为,就我们所知,并不需要人们去想到它们具有在本质上同非生命物质不同的特性。
相反,基因物质的性质看来同我们根据对非生命物质的物理学方面知识熟知的物质却是大不相同。现代物理科学未能为我们提供一种模型可用以解释基因线在每一细胞世代的再复制,或解释产生足够量的特异酶或由特殊基因所产生的别的作用物。同源基因线在减数分裂时节段的精确配对和互换也使人设想到这种物质有其新的物理特性。这些事实说明,了解基因物质的本性和特性,可能为阐明生命的不同物理机理提供线索。
研究基因物质的困难是明显的。它不能分离出来作化学分析或纯培养。直接分析一些特殊节段或个别基因的可能性,当然是更加遥远的事了。基因的特性只能从它们作用的结果进行推理。
此外,只有通过比较其基因型,除一个基因不同外,其他总的方面都相似的个体,或可严密地研究一个单基因的效应。这意味着基因突变对这类研究的重要性,因为只有通过基因突变,我们才能鉴定出仅由一个单基因产生差别的个体。为此,确定基因特性的前景依赖于发展出研究基因突变的可靠方法。
这里引用H. J.Muller对这一问题的研究所作出的不朽贡献是合适的。30多年前,他清楚地认识到基因突变在研究生命的物理性质方面有着独一无二的意义[1 ] ,并勇敢地解决了阻碍进行实验研究的给人深刻印象的技术问题。
上述分析方面的困难,在涉及其他问题时也同样如此,例如,关于分子和原子的结构问题,所假设的元素特性,必须从它们产生的效应来进行推理。在这类研究中,研究工作者的工作进展是通过先形成一个与已知事实相符的最简单的模型,然后采用每一个重大的实验来测试根据这个模型可以作出的各种预测。通过一系列连续的近似值,这个模型最后推导出一种形式,它似乎对观察到的行为提供了好像最合理的机理。从纯遗传学证据研究基因的物理性质同这种研究很相似。
通过直接观察染色体,有可能平行地研究突变的某些问题,这使得分析方面的困难有所克服。虽然在显微镜下看不见基因线本身,但可以说,它的行为在染色体上提供了一个可见的影子。根据染色体可见的改变很容易探查出基因线的改变。各别突变的细胞遗传学分析能对统计突变率所得出的假设进行有益的核对。
这方面的一个例子是对X射线处理和温度影响突变率这一证据所作的某些解释。X射线诱发突变的早期研究中,Delbrueck[2 ] 形成了一个纯属推测的基因“原子物理模型”,这是根据在不同物理条件下看到的点突变频率而推导出来的。数年后,著名的理论物理学家薛定谔(Erwin Schrödinger)发表了《生命是什么》一书[3 ] ,在这本引人入胜的小书中,他应用和讨论了基因的这个模型而为人们所熟知。
这个见解是把基因看成一个分子,而看到的突变则代表由于热激发或辐射能的吸收,使分子从一个稳定的状态迁跃到另一个状态。不管时间因素变化如何,线性剂量曲线和突变产量的稳定性,说明X射线诱发的突变是单“打击”造成的结果;不管波长怎样变换,突变产量与电离作用呈稳定的比例,说明“打击”单位是一次电离。对所看到的突变,计算其必定发生“打击”的体积,以这个体积为基础来估测基因分子的平均大小。计算结果是1000个原子左右。根据不同温度条件下自发突变的相对频率能计算出发生一个突变所需要的激活能量,它的计算结果约为1.5电子伏特。据推测,不稳定的基因相应地具有较低的激活能量,而事实上,温度对它们的突变率的影响比通常对稳定基因的影响要低些,这一点同以上所作的预测是一致的。一次电离所耗费的能量大约为30电子伏特,因此可以指望辐射将使任何一个基因发生突变,而不管在正常条件下它们的相对稳定性如何。所以,在正常温度下,明显不稳定的基因较之通常稳定的基因,其突变率有比例的增加要低得多,这些预测也是可以理解的。
这是一幅给人印象深刻的情景,但多年来很明显它同得到这个推测的实验数据无可靠的关系。在X射线诱发突变当中,对各个例子的详细分析清楚地证明,这些突变结果中有许多不是由于一个基因的结构改变,而由于基因外部的一些变动,如基因线一个节段的物理丢失或重排。我们没有根据去估测在看到的突变总数中这类基因外突变所占的比例;也没有理由认为在各种实验处理下所看到的突变当中这个比例是相同的。
这个模型的基础是假设所看到的突变统计,实际上是组成基因线分子发生结构改变的统计。某些突变的研究同这一假设不符,并证明这个模型事实是没有什么根据的。
考虑到以下情况是有兴趣的,即如果碰到遗传决定子更小,并处在能对个别例子进行实验研究的水准之下,我们仍然可以对这种模式形成越来越改进的基因模型。当根据模型作出的预测同实验结果相抵触时,我们就变换各数字值,或引进其他变数,或者如需要的话,甚至可创造其他假设单位。但模型主要地仍然是从为数不多的突变推导出来的一个想象的结构,因为我们不可能对这一个突变同另一个突变,同样有效这一似乎有理的假设进行反驳。
基因是什么?
基因突变的早期研究主要涉及这种极罕见的现象提出的技术性问题。尽管作为突变理论基础的月见草突变已被证明是错觉引起的结果,但不久就弄清楚,突变变化的确是存在的,其遗传情况好像它们是由于个别基因的改变所致。摩尔根和他的同事对果蝇所作的大量的遗传分析,证明有许多这样的情况,即事实上差不多所有在基因图上出现的位点都代表看得见变化的突变发生,这种突变经以后的测验证明它们是以典型的孟德尔方式遗传的。在每一种情况下,都认为这些突变是由于野生型基因改变成另一种形式,它产生一种可以识别的不同的表型效应。然而,这些突变频率,要允许做实验以研究影响它们发生的条件似乎太低了。
Muller于1917年指出[4 ] ,基因突变造成无生活力(“致死”),可能比能存活下来而对基因进行修饰(可见的)的突变要更多些。实验继续到第二个10年[5 ] ,他发展了各种专门的技术,用这些技术有可能测定某个染色体或某区内全部位点的致死突变总数。这些总的频率经证明已高到足以在不同温度下对突变频率作重要的实验比较。产生致死突变的位点分布在染色体上的情况,同根据可见突变的位点分布所预期的情况很接近,因而得出结论,可以合理地把致死突变用作总的基因突变的一个指标。
与此同时,作了许多外部处理的尝试,包括各种化学、放射和血清液处理的研究,并在研究中使用了各种动物、植物,以提高遗传改变的频率。这些实验都没有就任一种实验处理时突变频率的影响作出肯定性的证明,虽然有几项试验其遗传改变或许是由处理诱发的。失败应归结于以下两点困难:①难以证明在受处理个体后代中看到的遗传改变,实际上的确是处理的结果,而并非受处理品系中存在一些遗传的不规则性的结果;②难以表明受处理组的突变频率的提高在统计学上是令人信服的。所需要的是一种适合在一个有机体里能探查出足够数目突变的遗传技术,在这个有机体里,突变性状的基因决定遗传能很容易得到证实。
Muller在果蝇方面设计的C1B技术特别适合于这个目的,而用这个技术进行的X射线实验[6 ,7 ] ,毫无疑问地证明X射线对突变频率影响很大。X染色体上的总的致死频率提高了100倍以上。此外,发现了许多可见突变,包括显性和隐性的突变,也包括从它们发生自发突变为过去已知道的突变体,以及许多以前没看到过的突变体。
紧接着这些实验,其他研究工作者设计了实验更严格地测试诱发突变的基因本性。可以猜想突变体的致死因子是缺失,即使是可见突变,也能想象到它是由于短的缺失或基因的自毁。如能诱发突变成一个不同的等位基因,和再用处理把这个等位基因回复突变成亲本的等位基因,那就太好了。但这是有争议的,因为两次突变不能都归结为基因的丢失。在Patterson和Muller[8 ] 以及Timofeéff-Ressovsky[9 ] 的实验中,已证明果蝇的若干位点在同一个位点上发生诱发突变和诱发回复突变。
继后用多种多样类型的高等动物、植物和微生物所作的实验,证明了电离辐射对突变频率的效应有着巨大的普遍性。以后用紫外线照射和各种化学处理的实验,也证明了它们对突变频率有影响。
然而,诱发突变的分析很快指出,需要对采纳的有关基因、基因突变的定义和准则作重新考虑。
基因突变的实验目的是研究新的基因形式的进化。因此,研究基因突变的技术为用以测定这些基因改变的频率。但基因的改变或许只能通过它的效应来识别,且不久就搞清楚,各种基因外的变动能产生典型基因突变所特有的效应[10 ] 。
这样,突变的工作定义与理想的定义必然不同。工作定义是必须考虑从实验证据概括出一般性结论。经实验鉴定为基因突变的突变,不仅包括基因内改变所引起的变异,也包括由于基因丢失、基因增添和基因彼此间空间关系所引起的变异。能采用某种测验来鉴定这些机械的改变。但没有一种测验能鉴定基因内部改变所引起的突变;简单的推理是那些由于特殊的机械原因而未能鉴定的突变,实际上可归结为含义理想的基因突变[11 ] 。
当我们根据一项实验作出由于X射线的作用,而形成了新基因的结论时,我们不是简单地叙述实验结果。在单独叙述中,我们是把以下两个步骤结合起来:①叙述所观察的实验结果;②阐述由于一种特殊机理所引起的突变。把这两个步骤分隔开这一点很重要,因为第一个步骤代表对已知事实永久性的增添;而第二个步骤只代表一种推理,这种推理以后可能为新的事实所修改或否定。如果把这两个步骤无意识地混同起来,我们就会有把我们所知道的同我们所设想的混淆在一起的冒险。
普遍认为X射线处理能大大提高基因突变的频率,就是这种把两个步骤混淆一起产生的错觉。其根源在于使用具有两种不同含义的基因突变一词。基因突变据设想是遗传物质的一个单位组成发生改变,从而产生出一个基因活动改变了的新基因。基因突变在实验中是通过发生一个遗传的突变性状来鉴定的,这个突变性状的发生,好像是因为一个基因的变化所引起的。
使用具有两种概念的同一名词所带来的危害性十分明显。坚持认为X射线诱发基因突变,是因为所产生的突变体满足基因突变全部被采纳的标准,认为这些突变体代表某些特殊基因的质量上的变化,是因为它正是我们用基因突变所指的东西,坚持这种论点可引用矮胖子在《透过镜子》里的一句名言,矮胖子说:“当我使用一词时”,“这是指我就是挑选它来解释——既不多也不少”。
现在我们关于基因的概念,全部地依赖于基因突变的发生。如果没有基因突变,我们就不能对各个基因进行鉴定,因为一个单个染色体的全部遗传效应能作为一个单位遗传下来。如果被我们解释成基因突变的突变,在实际上是由于大批基因发生改变的结果,那么,我们作为基因来看待的实体,事实上将是相应的一大批基因。这种明显的模棱两可性不在于我们对基因突变所下的定义,而在于我们对基因本身所下的定义,因为任何一种基因突变的定义都是以基因的定义为先决条件的。
对这些困难和用实验概念更精确的定义对它们作出修改的可能性进行讨论的是,只有在生物学中使用那种在现代物理学中占有共同地位的可使用的观点,这种观点在很大程度上是根据P. W.Bridgman[12 ] 的严密的研究结果提出的。Bridgman注意到,如果使方法具有灵活性而足以处理可能发展的任何一种事实,那么在科学见解上,随着相对论的发展,物理学必须作出这种严格的重新考虑是必要的。这种可以使用的观点,其主要特点是不能用超越实验测定的假想的性质,来规定一个处于实验研究条件下的物体或现象,而必须用能处理该物体或现象的实际可行的方法来规定它。这不是一个新的原则;至少在早期的个别科学家的工作中,已含蓄地承认了这一点。William James[13 ] 在他关于实用主义的演讲中主要阐述了这一点,并引用了Wilhelm Ostwald的话加以说明!
化学家长期以来围绕着某些称之为互变异构物体的内部结构问题进行辩论。它们的特点似乎同这样一种想法完全一致,即在它们的里面有一个不稳定的氢原子在振荡,或者它们是两个物体的不稳定的混合物。论点激烈,却一直未得结局。Ostwald说:“如果论战者问问他们自己,有哪些特定的实验事实同这一种或另一种所谓正确的观点有所不同,则从来没有发生过这样的事,因为接着看到不可能追溯到事实上有什么不同;而这种争论就好像在原始时候对产生酵母面团所作的推论那样的不真实,一方祈求于‘神仙’,而另一方则坚持‘妖怪’为其现象的真正起因”。
用可以使用的话来说,什么是基因?换言之,我们怎样才能用一种把确立下来的事实同推论和解释分割开来给基因下定义?这个定义不仅要考虑到发生突变的实验证据,也要考虑到任何一种遗传差别在遗传方面的实验证据,或者同基因本性有关的任一种其他的实验证据。这个定义能详细说明基因的特征,用公认的实验操作能测定出究竟它们是以往实验中早已确立的特征,还是在今后的实验中可予确立的特征。
从实用的角度来看,只能把基因定为基因线的最小的节段,经证明它始终同发生一种特异的遗传效应相联系。不能把它定为一个单分子,因为我们没有能用于实际例子的实验操作,来确定某一个基因是否是一个单分子。我们不能把它定为一个不可分的单位,因为,尽管我们的定义提出,我们将承认任何一个通过交换或易位,实际上分开的决定子是分开的基因,但没有实验操作能保证不能再把它分开。鉴于相似的理由,不能把它定为繁殖的单位或基因线的活动单位,也不能证明通过一定的界限能把它同邻近的基因划分开。
这并不是指有关上述未确定下来的问题是一些毫无意义的问题。反之,它们全部是重要的问题,我们希望通过对实验证据的阐明和发展新的实验操作来最终地对此作出答复。所谓实用的定义,仅仅代表迄今用现有的方法,从实验证据中可以确立的这种现实的基因特性。从这些证据作出的推理对假设的基因提出了一个假设的模型,一个模型对从事这一问题的不同研究者看来会略有差别,并将通过以后的研究进一步作出修改。
现代遗传学文献中所用的基因一词,有时指实用的基因,有时指假设的基因,而有时必然众所周知,是两种定义荒谬的混合。由此引起的混乱可以通过两位如此富有才能和卓有见识的遗传学家,Richard Goldschmidt和A. H.Sturtevant所发表的似乎矛盾的陈述,有力地说明了这一点。Goldschmidt在评述位置效应的证据后,声称基因是不存在的[14 ] ,或不管怎样,必须放弃微粒基因的经典理论[15 ] 。Sturtevant,在引用染色体区域上有差别,特定区域为有机体特定反应所必需,以及特定区域的行为犹如交换单位这方面的证据时,声称“这些情况……证明基因的存在”[16 ] 。
如果说Goldschmidt基本上是正确的话,他所说的基因是指假设的基因,而他记住的是特定的假设基因。他关于不存在基因的肯定性结论容易使人误解,但显然只是指这种假设基因并不存在。他关于把基因特性一般都归结到“经典的微粒基因”缺乏证据这一论点,我想是完全正确的。
如果说Sturtevant是正确的话,我们说他指的是实用定义的基因,因为这种基因定义不包括未经证明的特性,Sturtevant指出,如果确实在基因线上没有连续的单位,那么在实验上确立这一事实的最直接的途径,将仍然是研究这些可以区分的染色体区的特性和相互关系。这些都是实用定义的基因。
什么是基因突变的实用的定义?我们承认,我们在基因突变方面的研究,就其主要问题来说,只是在用它们鉴定和分析代表新的遗传单位进化的突变这一范围是重要的。但显然,在这种含义上,现在尚不能形成基因突变的实用的定义,因为这些遗传单位不是实用定义的基因;它们是我们为阐明实验证据而假定的假设基因。所以说,现在不能定下实用的定义,只是用不同的话来重申前面提到的见解,即我们没有正确的准则来鉴定基因内部变化而产生的突变,而在实验中被解释成基因突变的变化仅只是未经分类的残留物,还不能证明是否由其他原因造成的结果。今后研究的主要问题必须发展出这类准则。
特殊基因的突变研究
本文的主要目的[17 ] 是强调指出以下令人不愉快的事实;要使我们在了解基因突变方面取得重大的进展,需要研究特殊基因的突变。实际情况之所以令人感到难办,是因为当必须用单基因作研究,尤其是研究单基因的自发突变时,由于突变所特有的极低的频率而产生的各种技术性困难,使我们处于不利的境地。说起来使人感到不快的是事实上像我们看到的那样,要在实验中排除伪造的突变或基因外突变,在混杂的非特异性位点上产生的突变率是没有希望的。
集中研究单基因的主要优点,是有可能用某些特殊突变体的直接实验分析,代替在所有的位点上都使用假想的一般化概念。所研究的每一个突变体都可以在诊断同一基因的其他突变体方面,提供的详细资料增添内容。
另一个更重要的优点是被选作研究的特殊位点可能是那样一些位点,它们在识别和分析它们的突变体方面,具有非凡的技术上的优越性。例如,玉米的Rr 和Ab 基因,像其他各种物种已知的基因一样,它们能产生自发突变体,这些突变体的样子是由于在这些位点上可识别出短的缺失而形成,使它们能清楚地区别出来。这并不证明自发突变不能由更小的缺失产生,但它提供了鉴定缺失这一大类的常规筛选,而不需要再作研究。隐性等位基因a是另一个有助于区别基因丢失和基因改变的非常有用的基因。这个等位基因尽管在表现型上只能通过它没有A的作用而予以识别,但也可以从基因缺失,通过它对诱发基因Dt的反应,即当存在Dt基因时它有时会回复到显性等位基因A而得以辨认出来。这种保持Dt的反应,在阐明A基因的自发突变和诱发突变实验时,为排除基因丢失提供了一个准则。R等位基因提供了一种不同的技术上的优点。R基因的表现型效应是它的很多等位基因,借助于植株颜色的深浅和模式方面非常轻微的差异,而可以客观地把它们区分开。一个具有变化相等的等位形式的基因,如只能通过它对某些反应之有反应或无反应来进行鉴别,它看起来就只有两个等位基因,并除非把具有这两种可予区别的活动水平的品系,在它们之间作杂交,否则它的突变查不出来。另一个具有巨大的实际重要性的优点是R和A都是影响胚乳性状的基因,因而适用于在大群体中鉴定突变。两个基因都明显地表现为轻微的生理效应,它们的突变体都能存活下来,没有查出生活力的丧失。
对于可导致产生一种与孟德尔式遗传相反的遗传现象作有效的分析,或许能深入研究经挑选合适的基因的突变,尽管事实上它在研究混杂的非特异性位点的突变,看来好像是没有希望的。
如果单基因的自发突变率实际上太低,而不能作有效的实验研究,那么这些考虑就没有什么价值了。选择不寻常的高突变率的基因作研究,不能安全地躲开这一困难,因为不能保证同不稳定基因行为有关的机理,可以代表有关典型基因突变的机理。微生物可以在实质上无限制的群体里对突变体进行有效的筛选,这样,使用微生物将克服这一困难,但不幸的是,它们没有提供研究所需的严格的遗传背景。
一种测定特异基因自发突变率的技术,对玉米行之有效,因为突变率低到每一百万个配子大约有一次[18 ] 。测试了8个基因,这些基因除了能在胚乳上显示出它们的效应这一技术上的优越性外,其他方面不加选择,它们全都产生了突变,但有一个测试的基因,其突变率约为每一百万个测试配子中有500次左右[19 ] 。这种产生突变数较多的基因,使比较试验足以证明在不同栽培条件下突变频率变动很大。例如,R基因在一些栽培条件下大群里找不到突变,但在另一些栽培条件下它的突变率高到0.2%。以后的研究证明这种差异部分是由于R等位基因固有的差异,部分是由于对R突变率进行修饰的因子所造成的差异[20 ] 。这些因子显然是相当常见的,因为有一项只有强烈效应才能探查出突变的研究,它表明在标记的7个区中,有3个区碰到过这类修饰因子[21 ] 。
所测定的平均突变率,对于就影响突变率的因素作有效的研究或取出适量的突变体样本作个别的研究,都显得太低了。然而,事实上突变率很容易受到形形色色的修饰因子的影响;这就有可能分离出经常发生特异基因突变,从而能直接进行实验研究的品系。
假基因突变的探查
即使选择性质最有利的基因进行研究,发展鉴定具有进化意义的基因突变的准则是困难的。在以往的研究中,用各种设想使这一问题看起来毋庸置疑地简单化。有些设想是未经证实的,有些则通过以后的发现证明它们是无效的。
例如,我们感到在我们实验中探查出来的一些突变,必定是有关基因在质量上的改变,因为无疑地,质量上起变化的基因是在进化过程中得到的。这主要同一种广泛传播的信念有关,这种信念是即使有些鉴定过的明显的基因突变经验证明是错误的,而真正的基因突变必须包括在未经分类的残留物中。
这种信念是荒谬的。姑且承认质量上发生改变的基因必然由突变产生,其突变率高到足可进行实验研究,但不能保证在我们突变实验中,发现的突变体可以代表它们进化的步骤。当我们着手在一项突变实验中鉴定突变体时,我们必须把自己局限在具有效应比较大的突变,大到足以使突变体的表现,超过了由于环境条件和遗传修饰因子所引起的各种各样表现的范围内发生,我们就没有办法去鉴定它们。对于发生这种潜在的突变,尚无良好的证据。支持假设基因高度稳定性的证据,仅仅涉及少数明显的突变。如果明天提出有说服力的证据,说明基因型自身的繁殖,只是群体内每一个世代取样的统计结果,而这个群体的基因在一种不易觉察的范围内有遗传变动,那么在我们现在的见解中再不会有什么地方同这个结论发生矛盾了。
研究起源不同的R等位基因,说明在植株颜色表达的水平上经常碰到微小的差异[22 ] 。如果这个基因变异的唯一来源是我们实验中所研究的那种变异,则不能指望有这类等位的差异,但由于有潜在的突变,能指望它们会发生这种差异。
如果发生潜在突变,它能大大地或整个地说明自然界中新的基因形式的进化。这样,在我们实验中得到鉴定的明显不同的突变,或许正好是基因外现象的结果,这一点是相当可能的。
第二种假设,或有一批假设涉及根据基因丢失来辨别基因突变的可能性。按照全部基因基本上都能存活的假设,最初认为诱发出来的隐性可见的性状,可以放心地把它看做是基因突变。各种细胞学例子否定了这一点,经细胞学证明单倍体组织或半合子个体生活力不足,而二倍体个体的纯合子则能存活。这类例子比较少。但由于随着缺失节段变得更小,缺失的细胞学和遗传学标准都到达它们有效的应用范围的限度,这就有理由怀疑,在缺失不能得到论证的情况下,物理学丢失也可能同观察到的突变有关系。随着我们对各个基因及其功能有更多的了解,那种认为基因各自对生命都是必需的假设照例已失去它的似真性。
有时把一个间性等位基因突变看做突变消失的证据。这涉及另一个关于基因的不可分性的假设,这个假设是在基因理论的早期发展中有意识地和经过仔细考虑而作出的,但通过以后的证明这个假设必然问题很大。这个假设只能是这样一种情况,复等位基因是一个单独单位的不同形式,由于突变的消失,我们可以排除发生复等位基因的可能性。根据复等位基因代表一个基因紧密连锁的复合体的假设,我们就能用基因链上不同节段的丢失来说明不同水平的突变。
以等位性经典的准则为基础的探究,或许能最好地说明选用不可分假说的根据所在。1919年摩尔根[23 ] 是这样说的:
大概有关基因本性最重要的证明是从复等位性得到的。现已有证明,同这些事例相关联的现象不能归之于成串的连锁的基因,我们或许要呼吁把这些事例作为至关重要的事例。……复等位基因是染色体上相同位点的修饰,而不是基因紧密连锁的事例,这种论证只能在其起源已知的情况下得到……
试以图1的5个圆圈来说,A代表一串紧密连锁的基因。如果第一行的头一个基因(B, a行)和另一行的第二个基因(B, b行)发生隐性突变,把a和b这两个突变体杂交,将得返祖类型,因为a带给B(b)正常的等位,而b带给(BA)a正常的等位。……现在的确不会发生这种情况,当一批等位基因间的成员进行杂交时,它们不产生野生型,但产生一个另一种突变类型或一个中间性状。显然,如果新的基因各自直接从一个不同的正常等位基因产生的话,一串连锁的正常基因的独立突变将不能解释其结果。
图1 (部分)
图解说明一串如此紧密连锁基因的突变,不发生交换。
人们将注意到试验否定存在着成串的紧密连锁的基因,这种看法仅只根据这样一种假设,即每次突变必须使这批连锁基因中有一个发生改变。然而,如果每次突变就是使这批成串的连锁基因有一个基因,或更多的相邻基因丢失掉的话,那么它们之间的杂交,一般都会证明它们是等位的,这一事实就不能排除作为复等位基因系列基础的复合基因。这可用以图2解排列予以说明:
图2
复合基因在某种意义上其含义是矛盾的,因为根据定义假设基因是不可分的。但经我们实验鉴定的基因不能得出定义上的不可分性。图2中所列的5个基因成分实际上不是一个基因的几个部分,它们是5个基因。但如果某种复等位系列具有这种主要成分,就有可能唯有在这种最有利于作分析的情况下通过实验确立这种事实。因此,有许多情况可能是在实验上鉴定为一个单基因的基因线的节段,实际上可能是一簇效应相同或相似的基因。
复合基因的概念,或者一些等效的单位,可以证明是有意义的,因为在这些把它们同邻近无关的成分区别开,作为一组的成簇成分中可能有特殊的关系。其中之一或许是成簇成分间基因活动的相互关系,当通过交换或易位把这簇基因的成员分开时,这种相互关系就能导致位置效应的发生。这可能是一般解释位置效应的一个主要因素。另一种可以预料到的相互关系是造成交换机会不等的联合等势。这是我们在后面要谈到的。
Rr 交换突变体提供了一个惊人的例子,即看起来好像基因突变的微小缺失[24 ] 。某些Rr 等位基因至少包括两个独立诱变的基因成分:P是决定某些植物组织或果皮的花色素苷色素;S是决定胚乳和胚的花色素苷色素。因此,由于不相等交换而产生的交换突变体Rg 和rr ,必须在一种情况下丢失P;和在另一种情况下丢失S。它们在细胞学和遗传学方面都没有得到缺失证据,并用非常敏感的试验,在突变体通过雄性生殖细胞传递方面比较花粉管的生长情况,结果也说明它们在单倍体配子体的发育中是完全正常的。交换突变体在外观和遗传行为方面与在相同栽培条件下发生的非交换突变体完全能区别出来。
R复合体内发生不相等交换得到一些有意义的指标,它涉及复等位系列的遗传性质和基因丢失关系到外观上质量突变的可能作用。除P和S外,还有别的表现型上可识别的R复合体的基因成分。某些使色素变淡的Rr 等位基因,其植株和种子颜色都由一个单基因成分D来决定。色素特别深的各种Rr 等位基因,看来有决定植物色素表现为某些样子的其他成分。此外,有各种不同的糊粉色种类,如点状、花纹状、那伐鹤点状以及有些植株有色、有些无色等等。可以把每一个可以区别的复合体看成R基因复等位基因一个长系列中的一段。
让我们花一点时间来澄清一下术语。为避免混淆起见,我把已公认的R等位基因在其上方以常用斜体字标明(Rr ,Rg ,rr ,RNj 等等),虽然分析表明,这些所谓等位基因,其中有一些实际上是两个或更多个基因的复合体。
基因成分这一名词将用于经鉴定为R等位基因之一的一个成分的任何一种相似与基因的组分。在缺乏进一步的证明前,使用这一名词并不意味着这个基因成分是不可分的。基因成分用字母,不用斜体字表示,如P, S, D等等。
除交换突变体外,有许多无交换突变体。一个无交换突变体rr ,不单是P,而是Ps的结构。假定成分s是一个无价值的成分,但它可能以相同的途径与S联合起作用,这些假定成分以s, p, d等标明。
当然,这种复杂情况包括由以往的突变所得到的别的无价值的成分,这些突变的亲本成分是不清楚的。这一类成分称为n。
在一些情况下,碰到种子颜色中等水平的无交换突变体,它包括各种色淡和模式种类。这些突变体以“Sd ”,“Ss ”等表示之。
一旦这些基因成分中有两个在同一个染色体上确定了它们的相邻位置,就对不相等交换提供了一个机会,可最终发展出更复杂的基因簇。例如,由互换而产生的上述交换突变体的情况如下:
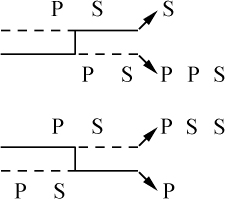
交换产物“S”经识别为一交换突变体Rg ,而交换产物“P”则为交换突变体rr 。交换产物“PPS”和“PSS”不能识别,但它们代表了带有3个而不是两个基因成分的潜在的新等位基因的产生。借助于在原始化合物中辨别S或P的形式,或能认出别的交换,并通过这个方法,有可能得到像R(点状-那伐鹤)那样新的综合的等位基因等等。照此下去,将可预料,通过连续的步骤,将发展出更复杂的基因簇,除非这个基因的作用局限在它复制的生活力上。
可以指望有可能代表等位系列成员的多种多样的基因型或可用下述少数例子予以说明:
①SSpn; ②SPPnS; ③D; ④DSP; ⑤Ss PD。
等位基因②和④是标准的Rr 类型;③是色淡的Rr 型;①是Rg 型,以及⑤是植株有色的点状糊粉型。总之,等位基因间的差别应归结于基因外变化,而不是基因内变化的结果。但④和⑤之间的表现型差异并不一定如此。
有关复合体基因成分间的关系方面,等位性的位点的概念没有多大意义。复合体的所有成员彼此间是同源的;大概它们全都是以几个单祖先基因经过长期的一系列突变产生的。在某种意义上,可以认为它们彼此间全都是等位的。例如Sn (RNj 的种子颜色成分)是否与S等位?这就没有什么意义,因为没有什么方法能证明Sn 与S的关系,比起它同P或该复合体的任何其他成分的关系有什么不同,像D成分对P成分是重要还是次要的这一类问题,其情况也是如此。它或许在这个原种中是重要的,在另一个原种中则是次要的;在D属于重要的原种,一个短系列的不相等交换就足以把它挪到一个次要的地位。
虽然不同等位基因可以具有数目不大相同的基因成分,但实际上没有一个缺失。就基因簇的假设起源来说,可以把那些具有一个单独成分以上的基因簇看成是复制。另一方面,当我们任意取一个带有几个基因成分的等位基因作为标准类型时,其他成分较少的等位基因看起来就像是缺失,产生这种等位基因就好像这种标准类型发生突变,其机理将是基因的丢失。
有一种基因复合体,它的可分离的成分作用相同,在这种情况下,相同的机理只能产生一种表示各颜色变浅等级的复等位基因的直线系列,或者它们根本不产生等位基因的复合系列。
有证明具有效应相同或相似的基因成簇的例子日益增多[ 24 ~ 27 等、 28 和 29 作参考]。这说明不相等交换在产生外表上属于质量方面的等位差异是一个重要的因素。
另一个简单化的设想是突变在基因效应方面的改变,必须代表基因本身的变化,而不是影响到基因表达的一些变动。正是这个假设,它论证了X射线诱发突变和同一基因的回复,这似乎是诱发基因内变化的严格的证明。这个假设同位置效应的证据显然有矛盾。这一证明无可争议地说明了一个突变并不必定代表该有关基因的一种变化或丢失,换句话说,它可能是一种易位,从而影响未改变的基因的表达。
McClintock在玉米中通过引进经历断裂—融合—桥周期的第九染色体,发现对突变行为有影响[30 ,31 ] ,他的这些出众的研究,证明了这种限定在基因突变实验研究中是非常重要的。存在这种结构上不稳定的染色体,就存在许多种基因,包括第九染色体和其他染色体的基因,并说明不稳定的隐性形式方面的突变,其特点为各种染色体的不规则性。研究不稳定突变体及其回复突变,使人毋庸置疑这种现象是由于有关基因的表达有些可逆的抑制。
在有些情况下,当发生突变时,表现不稳定的那个位点或该位点的附近,同时可探查出染色体畸变,但另一些情况则没有同发生突变有联系的细胞学上可探查出的染色体畸变。有许多例子,隐性突变体的不稳定性同发生与此有关的染色体不规则性取决于是否存在一种称之为激活剂(Ac)的互补因素,而当除去这个因素时,突变体的行为看起来就像一个具有正常染色体行为的稳定的突变体了。
McClintock还证明用Dt(点状)来控制隐性a的回复突变,或许是激活剂的一种反应。当有畸形的第九染色体而无Dt时,显然由于突变成A,标准的a等位基因产生了偶尔带点状的胚乳,如果是这样的话,它在Dt影响下的回复突变,必定也是由于影响基因表达的条件发生了一些改变。
不管怎样,有人接受我如下的假说,即不稳定基因的这些表现形式是由于有一点点看不见的异染色体转移到受影响的基因位点而引起的,这项卓越的研究清楚地表明了,表达效应或许是明显的基因突变的真实起因,甚至当所观察的突变没有看到位置的变化或任何一种与此有关的染色体改变也是如此。
在分析所观察到的突变方面产生的困难,进一步强调指出,在进行分析时必须优先在特殊位点对突变作细致的研究。如果我们用整个突变研究所特有的概括出的假设来考虑这些结果,我们或许会倾向于把这些发现大体上应用于基因不稳定的性质,或甚至大体上应用于突变体的等位基因。如果我们以一些经过深入研究的基因的相反突变为背景来考虑这些结果,我们则倾向于把这类突变体同其他种突变体和别的起源模式的突变体作仔细的比较,希望发展出能区别不同种突变体的准则。
与此同时,在研究基因突变方面,我们现在处在一个难办的地位。一个突变体可能在每次测试基因突变体时都会碰上它,然而,如果这个突变体不能回复突变,那就有理由怀疑它可能是基因丢失,而如果它能够回复突变,则有理由怀疑它可能是一种表达效应。要摆脱这种进退两难的困境的唯一办法,是选用最适宜于作详细遗传分析的特殊基因进行更深入的突变研究,以期发展出鉴定基因突变的更灵敏的准则。
参考文献
[1 ]Muller, H. J., 1922. Am. Naturalist. 56 , 32.
[2 ]Delbrueck, M., 1935. Nachr. Ges. Wiss. Gottingen (Math. Physik. kl., Biol.)1 . 223.
[3 ]Schrödinger, E., 1944. What is Life? (Cambridge Univ. Press, New York).
[4 ]Muller, H. J., 1917. Proc. Natl. Acad. Sci., U. S. A. 3 , 619.
[5 ]Muller, H. J., 1928. Genetics. 13 , 279.
[6 ]Muller, H. J., 1927. Science. 66 , 84.
[7 ]Muller, H. J., Z. ind., 1928. Suppl. 1 , 234.
[8 ]Patterson, J. T., and Muller, H. J., 1930. Genetics. 15 , 495.
[9 ]Timoféeff-Ressovsky, N. W., 1930. Naturwiss. 18 , 434.
[10 ]Stadler, L. J., 1931. Sci, Agr. 11 , 645.
[11 ]Stadler, L. J., 1932. Proc. Tth Intern. Congr. Genet. 1 , 274.
[12 ]Bridgman, P. W., 1927. The Logic of Modern Physics (Macmillan New York).
[13 ]James, W., 1907. Pragmatism (Longmans, Green, London).
[14 ]Goldschmidt, R., 1938. Monthly. 46 , 268.
[15 ]Goldschmidt, R., 1946. Experientia. 2 , 1.
[16 ]Sturfevant, A. H., 1950. In Genetics in the 20th Century (Macmillan, New York). 101~110.
[17 ]Given as the presidential address, American Society of Naturalists, annual meeting, Boston, Mass., Dec. 30, 1953. It is a report of the cooperative investigations of the Field Crops Research Branch, Agricultural Research Service, U. S. Department of Field Crops, University of Missouri (Missouri Agri. Expt. Sta. J., Ser. No. 1409). The Work Was aided by a grant from the U. S. Atomic Energy commission.
[18 ]Stadler, L. 1946. Genetics. 31 , 377.
[19 ]Stadler, L., 1942. Spragg Memorial Lectures, 3rd ser. (Michigan State College, East Lansing). 3~15.
[20 ]Stadler, L., 1948. Am. Naturalist. 82 ; 289.
[21 ]Stadler, L., 1949. Ibid. 83 , 5.
[22 ]Stadler, L., 1951. Cold Spring Harbor Symposia. 16 , 49.
[23 ]Morgan, T. H., 1919. The Physical Basis of Heredity (Lippincott, Philadelphia).
[24 ]Lewis, E. B., 1945. Genetics. 30 , 137.
[25]Green, M. M., and Green, K. C., 1949. Proc. Natl. Acad. Sci., USA. 35 , 586.
[26]Laughnan, J., 1949. Ibid. 35 , 167.
[27 ]Silow, R. A., and Yu, C. P., 1942. J. Genet. 43 , 249.
[28 ]Green, M. M., 1954. Proc. Natl. Acad. Sci., USA. 40 , 92.
[29 ]Stephens, S. G., 1951. Advances in Genetics. 4 , 247.
[30 ]McClintock, B., 1950. Proc. Natl. Acad. Sci., USA. 36 , 344.
[31 ]McClintock, B., 1951. Cold Spring Harbor Symposia. 16 , 13.
细菌噬菌体遗传区的精细结构 【71】
西摩尔·本泽(S. Benzer) 【72】
(1955年)
本文将叙述在细菌噬菌体遗传物质中有一个与功能有关的区域,这个区域是通过突变和遗传重组细分出来的。这类突变同许多生物中观察到的相似情况颇为相像,通常把它称为拟等位基因(见Lewis[1 ] 和Pontecorvo[2 ] 的评论)。这类例子特别有意思,因为它们对遗传决定子的结构和功能有影响。
遗传重组的现象为分离突变体和在染色体上识别突变的位置提供了一个有力的工具。当突变发生紧接着毗邻的突变,就造成了困难,因为这两个突变越是彼此紧接着,它们之间发生重组的概率就越小。因此,当在数目有限的后代中看不到重组体类型时,通常不能就此认为下述结论是正确的,即这两个突变是不可分的,而只能认出它们之间连锁距离的一个上限。高水平地解决这个问题需要对许许多多后代进行检验。如果可提供一种选择特点来探查为数不多的重组体,则能最好地达到此目的。
本文所描述的T4噬菌体的rⅡ突变的例子就提供了这样一个特点。野生型噬菌体在两个细菌寄主B或K,都产生噬菌斑。而rⅡ组突变体只对B产生噬菌斑。因此,如在两个不同的rⅡ突变体间作杂交,由此产生的任何一种野生型重组体,即使其比例小到10-8 ,也能够通过把它涂布在K上而检查出来。
这种巨大的灵敏性引起了下面这样一个问题,即这个问题的解决同遗传物质的分子水平是怎样密切相关的。根据Hershey和Chase[3 ] 的试验,看来实际上肯定噬菌体的遗传信息是被携带在它的DNA中。经Hershey、Dixon和Chase[4 ] 测定,一个噬菌体T2颗粒的DNA总量为4×105 核苷酸。T4的量相似[5 ] 。如果我们采纳Watson和Crick[6 ] 提出的DNA结构由两条核苷酸链组成的模型,这就相当于每个T4颗粒DNA的总长度为2×105 的核苷酸对。我们希望把遗传重组试验所得出的连锁距离翻译成分子单位。目前尚不能非常精确地做到这一点。不知道是否一个噬菌体颗粒的全部DNA都是必不可缺的遗传物质。也不知道一个噬菌体染色体(即用遗传方法鉴定的一个连锁群的物理副本)是否由一个单个(双重的)DNA纤维组成,或者遗传重组是否在所有染色体区域很可能是相等的。然而,为了作一粗浅的计算,先假定这些看法是正确的。这样我们就把T4总的连锁图整理成与DNA的2×105 核苷酸对相一致。噬菌体T4 3个连锁群总的已知长度等于约100个单位(一个单位等于一次标准杂交中有1%的重组)[7 ] 。此外,有人证明[8 ] 大体上另有100个单位的长度连接两个连锁群。因此,如果我们假设200个重组单位相当于2×105 个核苷酸对,则每个核苷酸对的重组为10-3 %。这就是说,有两个噬菌体突变体,它们在染色体上发生突变的部位仅仅相距一个核苷酸对,那么这两个突变体之间的杂交必然得到一个后代群体,它在105 中有一粒来自突变之间的重组(当然,假定在相邻核苷酸对之间进行重组是可能的)。这种计算极为粗糙,只打算指出比率因子在数量上的次序而已。这里提出的是一项打算把遗传学研究扩大到分子(核苷酸)水平课题的一些初步的结果。
r突变体
野生型噬菌体T2、T4和T6,当把它们涂布在大肠杆菌B菌株时,产生边缘粗糙的小的噬菌斑。从这些噬菌斑清洗出来的部分,很容易分离出产生噬菌斑大而边缘清晰的突变体(Hershey[9 ] )。这些突变体因其溶菌迅速而称为“r”;它们与野生型不同的是不能对B菌株造成溶菌抑制(Doermann[10 ] )。当野生型与r突变体这两种类型一起在B菌株上生长时,野生型的选择优越性超过r突变体。Hershey和Rotman[11 ] 研究了r突变体的遗传,他们发现,产生r表型的各种突变,其中包括经证明在遗传上同别的突变不同的一种大的成串突变体,位于T2连锁群的3个区域上。Doermann和Hill[7 ] 对T4的遗传研究证明其r区域与T2的那两个区是一致的。T6也至少有两个这样的r区域。
rⅡ组
对所有3种噬菌体T2、T4和T6来说,r突变体根据它们对B以外的菌株的行为能分成不同组。本文将只涉及一个组,称为rⅡ组。rⅡ组突变体同别的组和野生型都不同,它不能对某些带有λ噬菌体的大肠杆菌溶原菌株产生噬菌斑[12 ] 。如表1所示,rⅡ组的一个突变体在B菌株上产生r型噬菌斑,对K12S菌株(对λ敏感的非溶原菌株)产生野生型噬菌斑;对K12S(λ)(用λ对K12S产生溶菌作用而得到)则不产生噬菌斑。野生型噬菌体对这3个菌株都产生相似的噬菌斑。对我们将在本文中谈到的T4来说,当然,除rⅡ对K12S(λ)外,其平皿培养的效率对3种菌株差不多是相等的。这3个细菌菌株在本文中叫做B、S和K。
表1 涂布在各种寄主上的T4野生型和rⅡ突变体的表现型噬菌斑的形态学
在B菌株上分离出来的将近三分之二独立产生的r突变体为rⅡ型。这一组包括Hershey和Rotman叙述过的T2的成串r突变体,以及Doermann和Hill在T4的相应的图区上介绍过的r47和r51,但不包括那个区以外部位的r突变体。同样地,如图1所示,所有表现rⅡ性状的新分离出来的突变体,却原来是在同一个区域里。
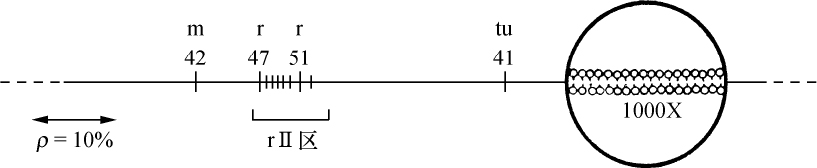
图1 T4(Doermann)的部分连锁图
表明rⅡ区的位置m和tu分别指微小噬菌斑和混浊噬菌斑突变。圆圈插图以图解表示DNA直径放大1000倍的相应的大小。
rⅡ组的特性特别有利于作细致的遗传研究。一个rⅡ突变体对3个寄主菌株有3种不同的表现型(见表1):①对B的噬菌斑形态上有改变;②对S的噬菌斑与野生型无区别;③对K不能产生噬菌斑。这几种特性全都有用。凭借它们在B菌株上改变了的噬菌斑类型,很容易分离出r突变体,再通过对K菌株的测验鉴定rⅡ组的那些突变体。当希望避免与野生型相比较有选择优点时,例如在测定突变率方面,能把S菌株作为一个没有区别的寄主。rⅡ突变体涂布在K上无效,使人们能探查出由于回复或由于不同rⅡ突变体之间的重组而产生的比例极小的野生型颗粒。
rⅡ突变体在K中的命运
野生型和rⅡ突变体都能很好地吸附S和K菌株。然而野生型还诱发溶菌作用,并在两个菌株上都能释放出许多后代,rⅡ突变体只在S上正常地生长。用一个rⅡ突变体侵染K,诱发一点(和/或很晚)溶菌作用裂解,尽管所有被侵染的细胞都死亡。rⅡ突变体生长的抑制同存在被携带的λ噬菌体有联系。这种联系的原因尚不清楚。
表现型的数量差异
尽管所有rⅡ突变体表现在K上增殖很少相同的表现型效应,但它们在这种效应的程度上是各异的。用rⅡ侵染一部分K,确实释放出一些后代,通过把侵染细胞涂布在B上面能够探查出来。把产生后代的侵染细胞的分数定为该突变体所特有的一个传递系数。传递系数对侵染的增殖是不敏感的,却大大地取决于细菌(K)的生理状态和温度。然而,在特定条件下,这个系数可用作表现型效应程度的一个相对指标,渗漏突变体的系数高。从表2可看到各种值。
表2 rⅡ组① T4突变体的特性
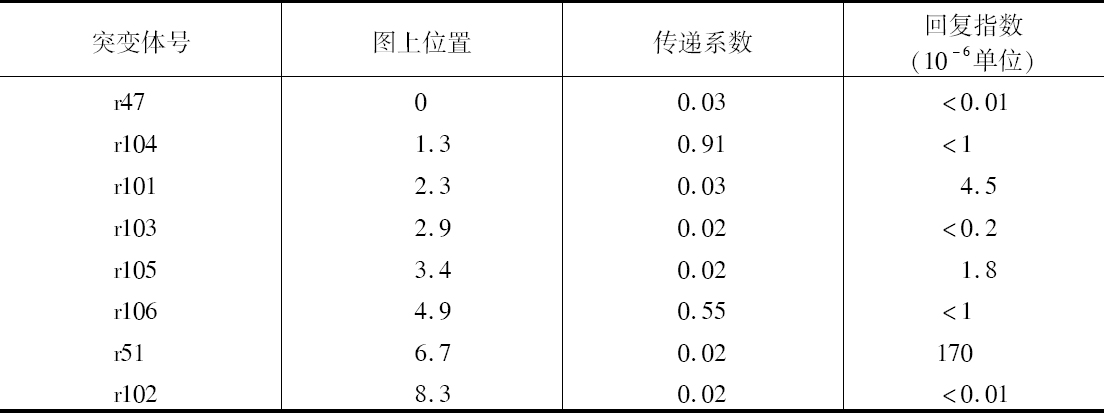
①每个突变体得出3个参数。图上位置是根据图2上所列的最近的间隔总数计算出来,并以r47的位置作为0,而得出重组单位的百分率。传递系数是用供研究的突变体侵染K细菌测出的表现型效应的一种测定,并以这种在B菌株上产生噬菌斑的侵染细胞的分数表示。回复指数是指在一个非选择性寄主上一个小的接种体所长出的突变体,其溶菌产物中出现的野生型颗粒的平均数。
K上的噬菌斑
有些rⅡ突变体在K上不产生噬菌斑,即使涂布接种原种多达108 颗粒(通过在B上计算噬菌斑测定)也如此。但别的rⅡ突变体,在K菌株上产生各种比例的噬菌斑。把在K上出现的噬菌斑挑出来再测验,就可以将它们分成以下3种:①一种类型像原始突变体,在K上产生非常稀少的噬菌斑,而在B上产生r型噬菌斑;②一种类型在K上产生效率良好的噬菌斑(常常次于野生型),但在B上产生r型噬菌斑;③一种同原始的野生型无区别的类型。这3种类型据知为以下原因产生:①泄漏效应,即突变体在K上能长一点点,所以有机会形成少数可见的噬菌斑;②一种突变,它部分地使rⅡ突变的效应失效,因而在K中能够增殖,但没有达到完全的野生表型;③明显的回复突变,它对于原始的野生型或许是真的回复突变,或许不是。
一个原种里面发生的每一种类型的比例,对一特定rⅡ突变体来说是特有的并能重复出现,但这一个rⅡ突变体同另一个rⅡ突变体则大不相同。这3种类型的发生比率无明显的相关。
rⅡ突变体的回复比率
Hershey[9 ] 曾证实r突变体回复到一种与野生型无法区别的类型,他利用野生型在B上的选择优点来提高它连续转移的比例。假设rⅡ突变体不能在K上产生噬菌斑,这种回复易于被探查出来,即使其比例很小。通过从一个小的接种体(例如,约100个颗粒,因而引进在原种中存在的野生型颗粒的机会非常小),制备溶菌产物能得出一特定的rⅡ突变体的回复频率的指数。如用S为寄主,从对照混合物可见,rⅡ突变体和任一种由它产生的回复突变,能复制的机会很少。在一些溶菌产物中出现野生型颗粒的平均数是一个指数。经证明这个指数同rⅡ突变每复制一次所发生的回复概率大致上成比例。在测定条件下,这个指数为每次复制回复概率的10~20倍的级数。在K上出现的噬菌斑,必须把它挑出来再涂布到B上进行测试。这将消除由部分回复突变和渗漏突变所产生的假噬菌斑,经揭露它在B上为r型。在表2可见rⅡ突变体的回复突变指数是多种多样的。发现有一个突变体,它回复的次数常超过r51的10倍以上,因而回复率为已知范围的105 倍以上。
尚未证明这些明显的回复变异是真正地回复到原始的野生型。但通过对原始野生型的回交,排除了远离rⅡ突变位置发生抑制因子突变的可能性。Krieg[13 ] 发现在一些回复变异的回交中,即使有r型重组体也很少,这种重组体使回复变异局限于相距原始rⅡ突变的连锁百分率的十分之几之内。也曾用回交测试过一种部分回复,而没有观察到rⅡ型重组体使部分回复突变集中在rⅡ区内。
rⅡ区的制图
两个rⅡ突变体之间的杂交是用每一种突变体相等的复感染(每个细菌3次)侵染B培养物来进行的。溶菌后的产物具有两个亲本类型,而如果亲本在遗传上是不同的话,则有两个重组体类型,即双突变体和野生型。在许多细胞的平均产物中,各重组体类型数目相等。在迄今所测试的全部例子中,双rⅡ突变体,像单突变体一样,对K不产生噬菌斑。假定这一点是普遍正确的话,那么把K上计算的噬菌斑(它只记载野生重组体)与B上计算的噬菌斑(它记载所有的类型)的比率加倍,就能简单地测出产物中重组体部分。这样,所测出的野生型百分率同B上噬菌斑类型的间接计算是很一致的。
用这个方法,把一批6个T4 rⅡ突变体(分离出来的头6个突变体,没有用任何方法选择过),在它们彼此之间和同r47和r51(由A. H. Doermann好意提供)作杂交,28次可能的配对中杂交了23次。这些杂交结果(见图2)与所指出的突变体的顺序排列是一致的。这种距离只是粗糙的累加;有一些有规律的偏差,即长的距离倾向于比它的短距离组成的总和要小些。这种偏差有一部分可以用Visconti Delbrück对多次交配[14 ] 的校正来说明。在这些杂交中,回复率小到微不足道的程度。这样,当这一批所有的rⅡ突变体都属于这个噬菌体连锁图的一小部分时,把它们明确地进行顺序排列是可能的,而它们的位置很好地分散在区域里面。
图2 8个rⅡ突变体包括Doermann的r47和r51在内的较大的比例图
新分离的突变体从101起计数。通过把后代涂布在K和B上,然后把K上计算的噬菌斑和B上计算的噬菌斑的比率加倍,得到每个杂交的重组值(%)。
对拟等位性的测验
两个产生相似缺陷的紧密连锁突变的功能关系,可通过构成具有结构不同的两个突变体的杂合二倍体进行测验。在一个染色体里面有两个突变的顺位类型,由于第二个染色体提供一完整的功能单位(或几个单位),其行为通常像野生型。然而,每一个染色体中有一个突变的反位类型,或者可以、或者不能产生野生表现型。如的确如此,结论是所说的这两个突变位于不同的功能单位。
对rⅡ突变体采用这种测验时,两种噬菌体的混合侵染看起来像是杂合二倍体。rⅡ表型不能对K溶菌,而野生型则引起溶菌作用,如K为野生型和rⅡ突变体混合侵染,细胞就被溶胞,释放出两种噬菌体。这样,细胞里面存在的野生型就发挥rⅡ类型所缺少的功能作用,而能把rⅡ突变体看做是隐性。尽管还没有进行过测试,也可假定双重rⅡ突变体加野生型的顺位结构在所有情况下都能产生溶菌作用。用所说的这一对rⅡ突变体侵染K得到反位结构。发现它是否产生溶菌作用是由哪些rⅡ突变体组成这一对突变体来决定的。其结果总结为图2中的虚线。虚线表示把rⅡ区分成两段。如若这两个突变体在同一节段,对K的混合侵染可得到突变体表型(非常少的细胞溶菌)。如果这两个突变体为不同节段,则发生大量溶菌,释放出两种侵染类型(和重组体)。这些结果综合于图3。这样,根据这一测验,rⅡ区的两个节段符合于独立的功能单位。
图3 对rⅡ突变体位置效应拟等位性测验的总结
每一个图解代表用具有所述突变体的两种噬菌体混合侵染一个细菌(K),看起来像是杂合二倍体。活性意味着混合侵染细胞的大量溶菌;失活意味着一点点溶菌。虚线代表rⅡ区的分裂点,其位置是由这些结果规定的。
实际上,对于用同一节段的两个(无渗漏)突变体混合侵染K,很小部分的细胞溶菌并释放出野生重组体,其比例随突变间连锁距离递增。对于以1%连锁距离(通过在B上作标准杂交测出)分开的两个rⅡ突变体来说,混合侵染K产生任何一个野生颗粒的比例约为0.2%。
这个值对于在B上两个rⅡ突变体之间杂交而产生的杂合噬菌体颗粒的K/B值有影响。在紧密连锁的rⅡ突变体之间作这样的杂交,其后代应当包括大约2%的具有反位结构杂合片[15 ] 的颗粒。当取这些颗粒中的一个涂布在K上,就有一定机会在侵染的第一个周期形成一个野生重组体,从而产生一个噬菌斑。如认为这些颗粒的侵染不可能超过用两个完全的突变体颗粒混合侵染K,就能得出结论,倘若这两个rⅡ突变体是在同一个节段上,则这些杂合颗粒对K上计数的影响是微不足道的。但是,对于不同节段的突变体来说,杂合颗粒的效率必然大得多,而用K/B方法测定的重组值必将大大地超过纯值。由于这一理由,图2中的重组值,对超越节段划分的杂交,可能要作一些校正。
用点测验进行粗略的绘图
如取两个rⅡ突变体中的这一个或那一个原种涂布在K上,都不产生噬菌斑,但如果把两者一起涂布在K上,则有些细菌会被两个突变体侵染,而如果这导致野生型重组体的发生,就产生噬菌斑。如果这两个突变体在它们之间不产生野生重组体(例如,如它们是相同的突变体),则不会出现噬菌斑。可以在一个单独的平皿培养上,用其他几个突变体来测试某一种rⅡ突变体,其方法是先用K加上所研究的这个突变体接种到平皿上,进行培养(上层常用软琼脂),然后用含有别的rⅡ突变体的水滴点缀在上面。
检查这种平皿培养,未知的突变体很快就占据正常的节段,这因为把节段A的任一个突变体滴在节段B的任一个突变体上,由于细菌被混合侵染产生大量溶菌而得到一个非常清楚的斑点。但是,对属于同一节段的一对突变体,只有相当少数被混合侵染,从而能产生野生重组体的细菌产生了噬菌斑。突变之间的连锁距离愈大,出现斑点的噬菌斑数就愈多。同一节段上的一批突变体就是这样通过把每一种突变体接种到平皿上进行培养,再用所有其他的突变体点缀在其上的方法予以顺序排列。如已有一早已安排好次序的突变组,一个新的突变体就能用这个方法迅速地在这个突变组中给以定位。
这一方法最适用于稳定(即低的回复率)和非渗漏的突变体,从而能对许多噬菌体颗粒在平皿上进行培养。回复变异或显著的渗漏效应明显地造成一含糊不清的背景。
这种测验已被用在大批稳定,非渗漏的rⅡ突变体。根据这些测验推导出来的近似位点见图4。有些突变体表现异常,致使不能把它们作为一个系列中的成员来定位。它们与位于某一距离之内的任一个突变体都很少发生重组,而同位于这段距离之外的突变体则表现正常。它们在图4中用延伸这段距离的水平线表示。
图4 根据点测验对各种rⅡ突变体的初步定位
对许多其他突变的点测验表明,回复率、传递系数和部分回复率各异的突变体,发生在两个节段的分散的位置上。
小串突变体的制图
点测验使我们能挑出小串突变体,即一批邻接很紧的突变。图4指出选作进一步研究的这样4个组,对它们绘图的结果列于图5。虽然有些间距合乎情理地表现出良好的累加特性,但一些突变体产生极其异常的结果。这样,在小串突变体a, r47同其他3个突变中的任何一个都不产生野生重组体(即106 中少于一个),但这3个突变体中有两对的确表现重组。如认为这每一个突变超过染色体的一定长度,而野生型的产生又需要在这些长度之间的范围内进行重组,那么这些结果是能够理解的。按照这种解释,突变将布满由图5中的线条指出的长度。这些异常现象同点测验观察到的现象颇为相像,只是它们更被限制在间距中。
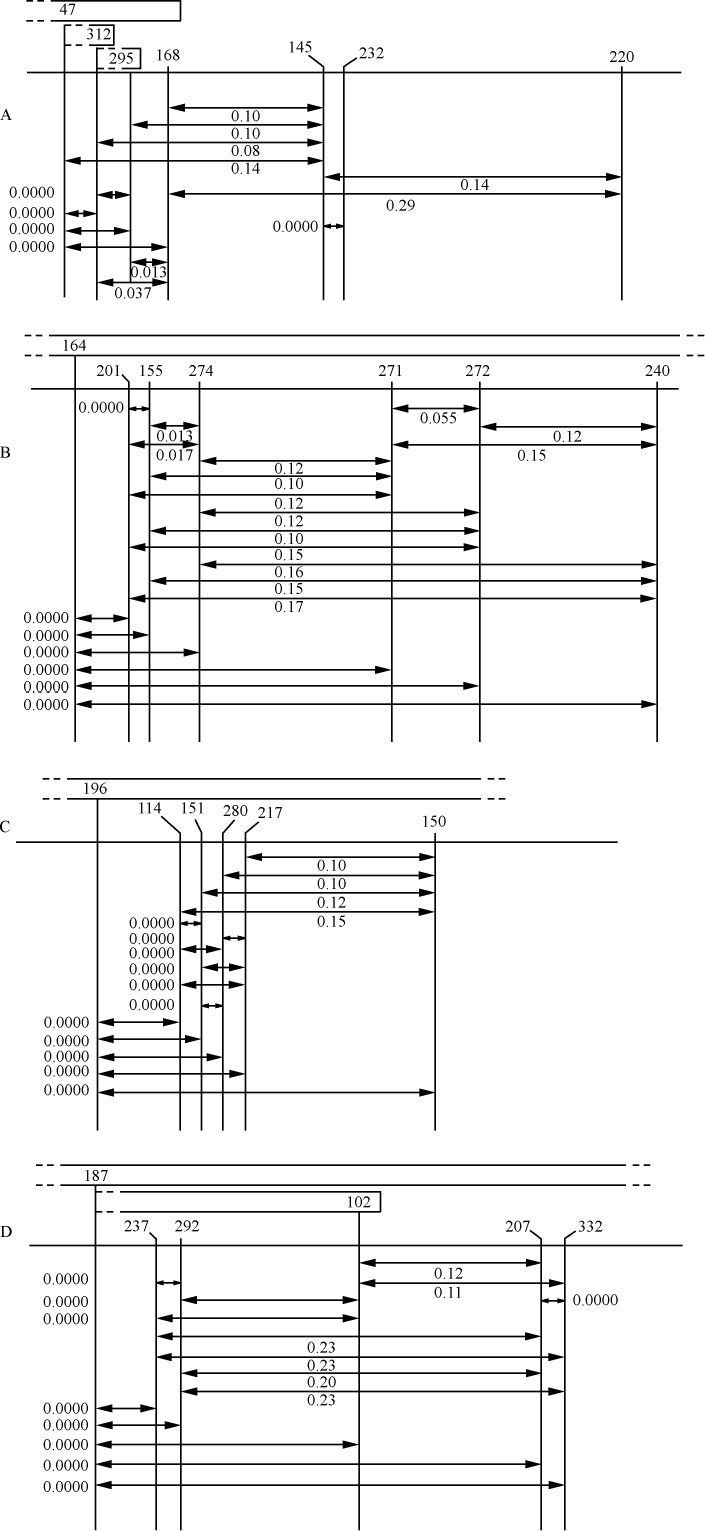
图5 小串突变体图
这项观察提出了一个问题,究竟是否存在着真正的点突变(即只有一个核苷酸对的变换)或是否所有的突变都牵涉到多多少少长的染色体片。必须记住这些试验所用的突变体经选择对抗回复变异方面是极其稳定的。可以预料这一步骤将使显著的染色体改变的突变体部分增多。就目前所知,所看到的异常例子可以同样想象到其原因是双重突变(即两个邻接着的点),倒位或野生型染色体的缺失。在继续进行这些试验时,看来要考虑到只采用那些有些回复变异可观察到的突变体。
讨论
规定一批rⅡ突变体有一个固定的连锁群区,在其中的各个位点上可发生突变,所有的突变都产生在质量上相似的表现型效应。因此,看起来rⅡ区是功能上有联系的,所以在这个区内的不管什么地方引起的突变都对同一种表现型有影响。在B菌株为寄主的情况下,这种效应表现为不能产生溶菌抑制作用;在S菌株为寄主的情况下,表现为无结果,而在K菌株为寄主的情况下,通常表现为不能增殖。rⅡ突变体不能在K菌株中成熟,但当同一细胞中存在野生型噬菌体时,就能克服这一困难。如果野生型染色体区的功能,是控制生产这种噬菌体在K细胞中繁殖所需的一种或几种物质,那么这种情况是可以理解的。
对拟等位性的表现型测验,把这个区分成两个功能上有别的节段。可以想象到这些节段对两个必要的连续发生的事件有影响,或者它们制造出一种单一的物质,这种物质的两个部分在使物质充分活化的次序上是明确无误的。例如,每一个节段可以控制一个特异的多肽链的生产,以后这两个链结合成一个酶。虽然不知道这种描述是否合适,但这种模型能用来说明rⅡ突变体所观察到的特性。一个突变在图上的位置将定位区域中的一个变异(也是酶分子的改变),回复率的特点是变化的类型牵涉到遗传物质,而表现型效应的程度则是酶活性产生变化程度的一种表达。渗漏突变体是一种末效应小的突变体。虽然在rⅡ突变体的这3个参数之间尚未观察到明显的相关性,人们通过更彻底的研究可以很好地予以揭示。
经交换分开的相似突变体的成串化,就Doermann和Hill研究的噬菌体的几个性状进行了观察,并看来可代表这种规律。或许所有的有机体都是这种规律,因为很简单,功能的遗传单位是由更小的重组和突变成分所组成,人们将指望在噬菌体中更容易看到这种效应,这因为噬菌体的每单位遗传物质重组的概率要比高等生物大得多。
把这些试验扩大到甚至更紧密连锁的突变,人们或许希望在分子水平上来描述遗传重组,突变和功能的最终单位大小的特征。我们初步的结果认为,由重组区分的染色体成分不会大于12个核苷酸对的等级(根据最小的不是零的重组值计算所得),而突变则牵涉到染色体长度各不相同,它可以扩及数百对核苷酸。
为了描述一个功能单位的特性,必需规定所指的是什么功能。在区域内不管哪里发生突变都产生rⅡ表型,就这个含义来说,整个rⅡ区是一个整体。根据反位结构杂合子的表型测验,可以把这个区再分成两个功能不同的节段,经估测每一个节段具有4×103 对核苷酸的等级。如果人们认为每个节段有指定一个多肽链上氨基酸顺序的功能,那么可以把每一个各别氨基酸的详细说明看做是一个完整的功能。用这个系统把遗传学研究甚至扩大到后一种功能成分看来是可行的。
小结
已发现T4噬菌体rⅡ区的突变有一共同特点,它同图中的所有其他部分的突变是分开的。此特点是一种寄主有效范围的降低,即它在对λ噬菌体为溶原的寄主(K)上不产生噬菌斑。这种突变体噬菌体颗粒吸附并杀死K,但溶菌正常,而不释放噬菌体。
具有这种特性的所有突变体都定位在噬菌体连锁图的一个明确规定的部分。但是,在这个区域内,它们的位置是大大散开的。除某些异常例子外,用大致累加的距离,能作出突变体的清楚的顺序排列。
K中同时存在着一种野生型噬菌体颗粒,有助于rⅡ突变体进行增殖,这显然由于提供了一种突变体所缺少的功能所致。用两个突变体类型混合侵染K,很像反位结构的一种杂合的二倍体。对rⅡ突变体进行表现型测验,可把这个区域分成两个功能不同的节段。
这些突变的大多数都看到了向野生型的自发回复变异。这些回复变异究竟是不是名副其实的回复尚待研究。每个突变体都以一特定的比率回复,但不同突变体的回复率大不相同。也观察到部分回复成中间类型。
突变体在K上生长的残余能力的程度差别很大。各突变体的回复率和残留活性程度在图位置之间没有看出有什么相关性。
K对野生型重组体的选择特性,为重组研究扩大到对这个区域作详尽细致的分析提供了可能性。
这种类型的初步研究表明,重组的单位不会大于12对核苷酸的级数,而突变可涉及染色体的各种长度。
参考文献
[1 ]Lewis, E. B., 1951. Cold Spring Harbor Symposia Quant. Biol., 16 : 159~174.
[2 ]Pontecorvo, G., 1952. Advances in Enzymol., 13 : 121~149.
[3 ]Hershey, A. D., and Chase, M., 1952. J. Gen. Physiol., 36 : 39~56.
[4 ]Hershey, A. D., Dixon, J., and Chase, M., 1953. J. Gen. Physiol., 36 : 777~789.
[5 ]Volkin, E. K., personal communication.
[6 ]Watson, J. D., and Crick, F. H. C., 1953. Cold Spring Harbor Symposia Quant. Biol., 18 : 123~131.
[7 ]Doermann, A. H., and Hill, M. B., 1953. Genetics, 38 : 79~90.
[8 ]Streisinger, G., and Bruce, V., personal communication.
[9 ]Hershey. A, D., 1946. Genetics, 31 : 620~640.
[10 ]Doermann, A. H., 1948. J. Bacteriol., 55 : 257~276.
[11 ]Hershey, A. D., and Rotman, R., 1949. Genetics, 34 : 44~71.
[12 ]Lederberg, E. M., and Lederberg, J., 1953. Genetics, 38 : 51~64.
[13 ]Krieg, D., personal communication.
[14 ]Visconti, N., and Delbrück, M., 1953. Genetics, 38 : 5~33.
[15 ]Hershey, A. D., and Chase, M., 1951. Cold Spring Harbor Symposia Quant. Biol., 16 : 471~479; Levinthal, C., 1954. Genetics, 39 : 169~184.
和核酸相似的多核苷酸的酶促合成 【73】
玛丽安娜·玛那哥(M. G.-Manago) 【74】 普利斯拉·奥尔蒂斯(P. J. Ortiz)
塞韦罗·奥乔亚(S. Ochoa)
(美国 纽约大学医学院生化系,1955年)
虽然我们对单核苷酸、嘌呤和嘧啶碱基,以及糖的酶促合成机制的了解,已有显著的进展,但是,对核酸的多核苷酸链的合成机制仍不清楚。
本实验室最近发表了一篇简讯[1 ] ,报道了从棕色固氮菌(Azotobacter vindandii)中分离到一种酶,它能催化用5′-核苷二磷酸来合成多核苷酸,并释放出正磷酸盐。此反应需要镁离子,并且是可逆的。有证据表明固氮菌的酶催化下列反应:
这里R代表核糖,P-P代表焦磷酸盐,P代表正磷酸盐,X代表一个或多个腺嘌呤(A)、次黄嘌呤(HX)、鸟嘌呤(G)、尿嘧啶(U)、或胞嘧啶(C)碱基。
用化学法和酶法降解这种生物合成的多核苷酸,已证明多核苷酸是由5′-单核苷酸单位组成的,像在RNA中一样,它们彼此通过3′-磷酸核糖酯键连接起来[2 ] 。因此,与多糖类似,在多核苷酸链的生物断裂和合成中,可逆的磷酸解作用可能是一种主要的机制。根据这个理由,把这种新酶叫做多核苷酸磷酸化酶[1 ] 。本文介绍了这种酶能催化合成和RNA类似的多核苷酸的证据[2 ] 。
多核苷酸磷酸化酶
这种酶是在研究生物磷酸化机制的过程中[3 ] 发现的。实验表明,在有Mg++ 存在的条件下,固氮菌的提取液能催化32 P-标记的正磷酸盐与腺苷、肌苷、鸟苷、尿苷和胞苷的5′-核苷二磷酸的末端磷酸基发生交换。随着对酶的进一步纯化,发现与5′-核苷一磷酸或5′-核苷三磷酸,如AMP、ATP、IMP或ITP都无反应。正如人们从反应式(1)中所预料的那样,交换和正磷酸盐的释放是相伴发生的,带有放射性同位素的磷酸盐就这样掺入到各种5′-核苷二磷酸中去了。
和以前报道的一样[1 ] ,通过硫酸铵的分级分离和磷酸钙凝胶吸附,酶活性提高约40倍;这可用ADP-正磷酸盐交换的速率来检验。在纯化过程中,ADP-正磷酸盐交换的速率与正磷酸盐释放的速率之比保持不变,证明这两种活性是彼此相关的。
单一聚合物
将纯化过的多核苷酸磷酸化酶与核苷二磷酸在有Mg2+ 存在的情况下一起保温,随着正磷酸盐的释放(化学数量),核苷二磷酸就会消失。当大约50%~60%的核苷二磷酸已消失时,反应达到平衡,并趋于停止。以下事实证明消失的二磷酸盐转变成了一种多核苷酸:
(1)新形成的成分带有很强的负电荷,它能留在Dowex-1阴离子交换柱上[4 ] ,随后用甲酸洗脱,所需要的浓度要比洗脱绝大多数酸性单核苷酸时要高些。
(2)产物是不能用蒸馏水或稀盐溶液来透析的,在冷却状态下,可用三氯醋酸或酒精来定量沉淀。因此,可用这种方法来分离它。
(3)它可溶于水,产生稍带黏性的溶液,这种溶液具有典型的核苷酸紫外吸收光谱。
(4)纸层析时,无论用哪一种溶剂系统,它都停留在原来区域,而且用水洗脱不下来。
把多核苷酸磷酸化酶与相应的5′-核苷二磷酸一起保温,可以得到只含有AMP、IMP、GMP、UMP或CMP作为碱基单位的单一聚合物。分离聚合物时,如果不使用加热和(或)酸的话,它们的平均分子量就有可能很高。AMP和IMP聚合物用光散射法测定时,得到的分子量分别是570000和800000道尔顿[5 ] 。
从IDP形成的多核苷酸——IMP聚合物,经弱碱水解[6 ] ,产生一种含有等克分子的2′-IMP和3′-IMP的混合物。这些产物再用Markham和Smith[7 ] 的No.3溶剂系统做纸层析,来进行鉴定。已知RNA经弱碱水解,产生的是2′-和3′-单核苷酸的混合物[8 ] 。IMP聚合物的碱水解产物是2′-和3′-IMP,可用下列实验进一步证实(见表1)。
表1 IMP多核苷酸的碱水解产物
多核苷酸用三氯醋酸沉淀两次,并用0.4mol/L的KOH,于37℃,水解39小时。然后把这种混合物用三氯醋酸处理;离心除去少量沉淀。根据紫外吸收得知(波长为260纳米)中性的上清液含单核苷酸为14.3微克分子。
(1)IMP多核苷酸的碱水解产物,大约有90%能被1.0mol/L盐酸在20分钟内水解掉。在这些条件下,将观察到,当5′-IMP几乎还未遭水解时,2′-IMP和3′-IMP都被水解达90%。
(2)这种碱水解产物,不受特异性的蛇毒5′-核苷酸酶的破坏[9 ] ,而5′-IMP却能被水解达96%的程度。
(3)Schuster和Kaplan[10 ] 的3′-特异性植物核苷酸酶能水解约一半的底物,它能水解3′-IMP,但不能水解2′-或5′-IMP。用层析法已证明,底物与这种酶一起保温,得到的产物是肌苷和2′-IMP。
通过研究特异性的磷酸二酯酶对这种生物合成的多核苷酸的降解作用,进一步阐明了它们的结构。这些实验有一部分是与美国国立健康研究所的Leon A. Heppel协作进行的。如图1所示,从蛇毒中提取的磷酸二酯酶[13 ] 水解RNA产生5′-单核苷酸;而Heppel和Hillmoe[11 ] 从脾脏中提取的磷酸二酯酶,水解RNA产生的却是相应的3′-单核苷酸[8 ] 。在这两种酶中,每一种酶作用于各种生物合成的聚合物,如AMP聚合物、IMP聚合物以及在下一部分将谈到的某些混合聚合物,都产生预期的5′-或3′-单核苷酸。用层析法和前面提到的特异性的单核苷酸酶的作用,可以对这些产物进行鉴定。用结晶的胰核糖核酸酶——它是一种磷酸二酯酶,主要是水解RNA的嘧啶核苷酸,形成3′-衍生物——将UMP聚合物降解,产生3′-UMP,要是用稀释的酶来进行降解,还有一定程度的环化尿苷酸(2′-3′-尿苷单磷酸)可以积累下来[12 ] 。这与观察到的胰核糖核酸酶对RNA的作用是一致的[13 ] 。

图1
AMP聚合物和其他生物合成聚合物纤维[14 ] 的X射线衍射图像与天然RNA纤维产生的图像是很相似的[15 ] 。
混合聚合物
混合聚合物——含有二种或多种不同碱基单位的多核苷酸——是把几种5′-核苷二磷酸的混合物与多核苷酸磷酸化酶一起保温来制备的。至今,已制得了两种这样的聚合物,一是从ADP和UDP等克分子 【75】 的混合物(A-U聚合物)制得的;另一种是从ADP、GDP、UDP和CDP分别以1∶0.5∶1∶1的克分子比例的混合物(A-G-U-C聚合物)来制备的。用纯化过的蛇毒磷酸二酯酶来降解A-U聚合物时,产生5′-AMP和5′-UMP,两种产物的克分子比例接近1∶1。用胰核糖核酸酶来降解时,能产生3′-UMP[12 ] ,以及含有一个尿苷酸单位和一个或多个腺苷酸单位的2′-、3′-、4′-和5′-核苷酸,同时还产生一个由腺苷酸基团组成的核心。
混合聚合物比相应的单一聚合物的混合物较易受核糖核酸酶的作用 【76】 ,这可从表2的结果得到明确说明。表2列出了A-U聚合物和相应的混合物的降解结果。表3说明A-G-U-C聚合物和单一聚合物的对照混合物也有同样的结果。和预期的一样,核糖核酸酶对AMP聚合物无作用。用Krebs和Hems[17 ] 的溶剂系统,对核糖核酸酶水解的混合物进行纸层析,证实了这些结果。看来好像是在混合物中,嘌呤核苷酸的聚合物妨碍核糖核酸酶作用于嘧啶核苷酸聚合物。从表3进一步看出,A-G-U-C聚合物与核糖核酸酶的反应和酵母RNA与该酶的反应是相似的。下述事实进一步证实了这种相似性:酵母RNA和A-G-U-C聚合物被核糖核酸酶彻底水解,只分别保留有20%和22%的核心部分[18 ] ,这种核心部分不能被蒸馏水透析,而AMP、GMP、UMP和CMP聚合物(用于表3的实验4中)的混合物,经同样处理后,保留有43%的不能被透析的核心部分。
表2 生物合成的多核苷酸用核糖核酸酶作短暂的降解
详细情况是在0.5毫升、浓度为50微克分子、pH7.35的Tris缓冲液中,含有1.48微克结晶的胰核糖核酸酶和指定的聚合物,把这种反应混合物在30℃、保温20分钟。根据能和醋酸双氧铀-高氯酸试剂产生沉淀的紫外吸收物质(波长为260纳米)的消失情况,用改进的Anfinsen等人的方法[16 ] 测定水解作用的大小。
①表示的是单核苷酸。
②单一的AMP(1.06 μM)聚合物和UMP(1.20 μM)聚合物混合,于30℃保温过夜,再用核糖核酸酶水解。
③假定混合物中,核糖核酸酶只作用于UMP聚合物时,UMP聚合物水解的百分数表示于括号内。
用高氯酸水解A-G-U-C聚合物,释放出A、G、U、C 4种碱基。
用32 P标记了两个磷酸盐基团[19 ] 的ADP和未标记的各种5′-核苷二磷酸[20 ] 的混合物合成各种多聚物,从这些实验中得到一些证据说明,在混合多聚物中,不同的单核苷酸单位之间形成了5′-3′-磷酸核糖二脂键[19 ] 。通过多核苷酸磷酸化酶的作用,标记的核苷酸以5′-(32 P)磷酸腺苷的形式掺入一条多核苷酸链,并释放出32 P-正磷酸盐。如果标记的AMP和另一些核苷酸之间连接起来的话,后者在这种聚合物受到脾脏磷酸二酯酶水解时,将以一种标记的3′-单核苷酸的形式释放出来,这样得到的3′-AMP的放射比活性将比掺入的5′-AMP的要低。相反,用蛇毒磷酸二酯酶水解时,释放出来的唯一标记核苷酸就是5′-AMP,而且和掺入的5′-AMP具有相同的放射比活性,释放出的其他5′-核苷酸是未标记过的。把用磷酸二酯酶水解释放出来的核苷酸再用纸层析和放射性自显影进行鉴定,确实也得到了这样的结果。在这些实验(详细报道正在整理)中,所用的生物合成聚合物是用下述材料制备的:①只有标记的ADP;②只有标记的UDP;③标记的ADP及未标记的UDP;④标记的ADP及未标记的GDP,UDP和CDP。
表3 生物合成的多核苷酸和酵母RNA经核糖核酸酶长时间的降解
详细情况是1.6毫升浓度为20微克分子,pH7.0的Tris缓冲液中,含有29.6微克的核糖核酸酶和指定的聚合物,把这种反应混合物在37℃保温。其他情况见表2。
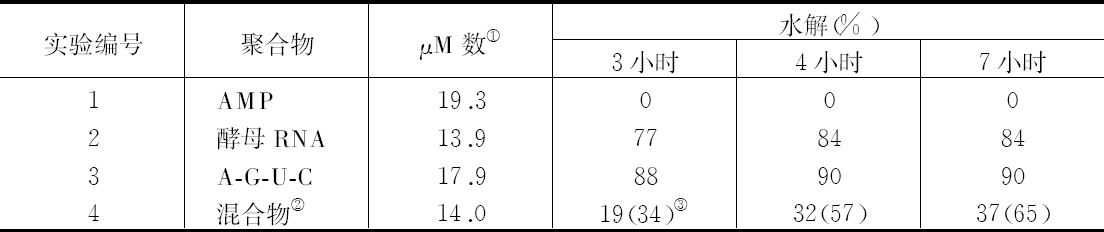
①表示的是单核苷酸。
②单一聚合物的混合物含有如下成分:AMP, 4 μM;GMP, 2 μM;UMP, 4 μM;CMP, 4 μM。
③假定在反应混合物中,核糖核酸酶只作用于嘧啶核苷酸聚合物时,这种成分水解的百分数表示于括号内(可以达到总数的57%)。
对照是①标记的AMP单一聚合物和未标记的UMP聚合物的混合物;或者②标记的AMP单一聚合物和未标记的GMP、UMP和CMP聚合物的混合物。在任何情况下,对照只产生标记的AMP(可能是3′-的,也可能是5′-的),其他的单核苷酸都是未标记的。
可逆性
前面已提到,用多核苷酸磷酸化酶催化的反应是可逆的。在这种酶、正磷酸盐和Mg++ 存在时,生物合成的多核苷酸经过磷酸解作用,产生相应的5′-核苷二磷酸。在正磷酸盐不存在时,不能发生反应。有关IDP反应向两个方向进行的化学计算的定量资料,以前已有过报道[1 ] 。迄今为止,关于这个平衡的情况,我们还没有确切资料。在多核苷酸合成方向上,在通常的实验条件下(pH8.1、30℃),当正磷酸盐的浓度与核苷二磷酸的比例从1.5到2.0时,反应趋于停止。核苷二磷酸的焦磷酸键能转变成多核苷酸的磷酸二酯键,从这个事实预料,在某种程度上,上述反应有利于多核苷酸的合成。
单一聚合物的磷酸解作用要比混合聚合物的磷酸解作用容易得多,道理现在还不很清楚。已经报道过[1 ] 从固氮菌分离出来的RNA的磷酸解作用,是通过放射性磷酸盐的掺入和放射性ADP、GDP、UDP和CDP的层析鉴定来证明的。这样形成的标记GDP和UDP的一致性,通过用肌苷二磷酸酶进行特异性酶促水解,得到了进一步证实[21 ] 。用同样的技术还发现,固氮菌的酶可以催化其他来源(酵母、牛肝、大肠杆菌和酿脓链球菌)的核糖核酸的磷酸解作用;这说明在这方面是没有特异性的。然而,小牛胸腺的DNA和酵母RNA中对核糖核酸酶有抗性的核心部分,是不受这种酶作用的。
结论
目前,有关结构、大小、X射线衍射图像和对不同酶的行为等方面的资料说明:在多核苷酸磷酸化酶催化下,由5′-核苷二磷酸合成的多核苷酸与RNA是非常相似的。这种A-G-U-C聚合物,即一种含有腺苷酸、鸟苷酸、尿苷酸和胞苷酸残基的聚合物,看来与天然RNA确实是一样的。当然,最理想的应当是在把它们当做核酸前,能确定这种生物合成的化合物是否具有影响蛋白质合成之类的生物学性质。
由多核苷酸磷酸化酶催化的这种反应,是否代表RNA合成的普遍性生物学机制呢?到目前为止,尚不能确定,但这种可能性是存在的。这种酶除了存在于固氮菌中以外,在其他微生物中也是存在的 【77】 ,但我们至今尚未得到确切证据说明它在酵母和动物组织中存在。
来源于不同细胞或组织的RNA,在组成上是不同的[18 ] ,这个问题得到证明以后,又出现了另一个具有重要意义的问题,即是否不同来源的多核苷酸磷酸化酶,将合成具有不同特异性的多核苷酸呢?因为DNA具有特异性的遗传效应[22 ] ,因此想到,和DNA类似的多核苷酸是否也能通过一种相似的机制来合成呢?这些问题都是很紧迫的问题,希望对其中某些问题能尽快得到回答。
参考文献
[1 ]Grunberg-Manago, M., and Ochoa, S., 1955. J. Am. Chem. Soc., 77 , 3165.
[2 ]See the third International Congress of Biochemistry. Brussels, Belgium, 3 Aug. 1955.
[3 ]Grunberg-Manago, M., and Ochoa, S., 1955. Federation Proc. 14 , 221.
[4 ]Cohn, W. E., 1950. J. Am. Chem. Soc., 72 , 1471.
[5 ]Cavallieri, L. F., and Rosoff, M., unpublished experiments.
[6 ]Vischer, E., and Chargaff, E., 1948. J. Biol. Chem., 176 , 715; Carter, C. E., 1950. J. Am. Chem. Soc., 72 , 1466.
[7 ]Markham R., and Smith, J. D., 1951. Biochem, J. 49 , 401.
[8 ]Cohn, W. E., Doherty, D. G., Volkin, E., 1953. in A Symposium on phosphorus metabolism Johns Hopkins press, Baltimore, 2 , 339.
[9 ]Guliand, J. M., and Jackson, E. M., 1938. Biochem. J., 32 , 590, 597; Hurst, R. O. and Butler. G. C., 1951. J. Biol. Chem., 193 , 91.
[10 ]Schuster, L., and Kaplan, N. O., 1953. J. Biol. Chem., 201 , 535.
[11 ]Heppel, L. A., Markham, R., and Hillmoe, R. J., 1953. Nature, 171 , 1152; Herpel, L. A., and Whitfeld. P. R., 1955. Biochem. J., 60 , 1.
[12 ]Heppel, L. A., unpublished experiments.
[13 ]Markham, R., and Smith, J. D., 1952. Bioehem. J., 52 , 552, 558.
[14 ]Rich, A., and Watson, J. D., 1954. Proc. Natl. Acad. Sci. USA, 40 , 759.
[15 ]Rich, A., unpublished experiments.
[16 ]Anfinsen, C. B. et al., 1954. J. Biol. Chem., 207 , 201.
[17 ]Krebs, H. A., and Hems, R., 1953. Bioehim. et Biophys. Acta, 12 , 172.
[18 ]Zamenhof, S., 1952. in A Symposium on Phosphorus metabolism( Johns Hopkins Press, Baltimore), 2 , 301.
[19 ]Prepared by the Sehwarz Laboratories, Mount Vernon, N. Y.
[20 ]These experiments were suggested by Plaut. G. W. E.
[21 ]Plaut, G. W. E., 1955. Federation Proc., 14 , 263.
[22 ]Hotchkiss, R. D., 1955. The Harvey Lectures (Academic Press, New York), 49 , 124.
病毒重建 【78】
Ⅱ.来自不同生理小种的蛋白质和核酸的重建
海因茨·康拉特(H. Fraenkel-Conrat) 【79】 B. 辛格(B. Singer)
(美国 加利福尼亚大学病毒实验室,1957年)
用病毒蛋白质和病毒核酸的小分子组分离体合成典型的烟草花叶病毒(TMV,下同)颗粒,以前已有报道[1 ] ,在这个过程中,同时也恢复了与原来的病毒性质相似的侵染性。这类研究现已扩大到多种TMV小种。来源于不同病毒小种的蛋白质和核酸能掺入到一个病毒颗粒中去,这在理论上和实践上都有重要意义。用4个不同小种的核酸和3个小种的蛋白质所进行的各种重建实验,都成功了。这种混合病毒制品的生物学和免疫学特性,以无可争议的证据支持了重建病毒的侵染性实际上是新形成的病毒颗粒性质的这一观点。对重建(从一个或两个小种)病毒后代的生物学和化学特性也进行了研究。其中某些结论在以前的文章中已有叙述[2 ] 。
材料和方法
病毒的制备和分离
实验中所用的各种TMV小种和本实验室以前的研究中所用的一样[3 ,4 ] 。
各种病毒制品用差速离心来分离。制备这些小种核酸的方法是以前用过的去垢剂法[1 ] ,但稍有改动[5 ] 。大约有90%的实验可以得到具有生物活性的核酸,这种核酸在-60℃保存数月仍然很稳定。
为了制备天然蛋白质,将病毒在3℃,pH 10.1~10.5进行降解[1 ] 。用2-氨基-2-甲基乙醇和乙醇胺作缓冲液,有利于这种降解。1%的病毒液(20~50毫升)在1000毫升的0.1%胺溶液中(用HCl调至pH10.5)透析16小时,然后进行超速离心(40000转,1小时,冷冻),如果出现的沉淀很少,则说明降解已十分完全。上清液用(NH4 )2 SO4 调到0.28饱和度,离心,把沉淀下来的蛋白质再溶于水。去掉用低浓度(NH4 )2 SO4 可以沉淀下来的少量物质,蛋白质在盐饱和度为0.15~0.25之间重新沉淀下来。对于每一部分中所含物质的性质和数量用分光光度法确定。最终得到的蛋白质沉淀,一般在280纳米的吸光率有一个突增的最大值,R值(最大值/最小值)为2.2~2.4,经透析后达2.4~2.5。污染0.1%的核酸,会使R值降低0.1。低温条件下彻底透析后,将蛋白质溶液调到pH8.0,再作超离心(40000rpm,2小时,并冷冻)。蛋白质溶液在冷冻状态下贮存,有防止损伤的明显趋势。冰冻干燥可能引起变性并降低它们重建的适应性。
制备M小种(masked strain)的蛋白质可以采用同样的方法。HR小种(Holmes ribgrass strain)的蛋白质也可以这样制备,只是困难较大,而且产量也低,将蛋白质分离开,需要较低的pH(9.8~10),因为从大的趋势来看,碱能使这些蛋白质变性。从YA小种(yellow aucuba strain)分离不到天然的蛋白质,可能原因就在这里。
抗血清
兔抗血清和γ-球蛋白是由R. C. Backus和G. Perez-Mendez夫人制备的。血清是按常规方法制备的。每逢双周肌肉注射1毫克的TMV或HR(用矿物油和水作辅助剂)。三周后,从兔子抽血,然后一周注射,一周抽血,交替进行。
在分析电泳仪中分离γ-球蛋白的微量成分。
抗血清的效价可用沉淀素试验及中和试验来测定。以后一种测定为例,把1毫升0.01%的TMV或HR病毒溶液(在0.075mol/L NaCl溶液中),与不同量的同源或异源抗血清混合,在3℃,处理16小时,为了降低病毒(100微克)的侵染性需加同源抗血清0.01~0.025毫升,系数为5或10。按通常的分析方法,经适当稀释后,用产生病变的数量来表示侵染性。然而,未经分离的血清和γ-球蛋白都存在时,有明显的交叉反应,特别是在TMV抗血清和HR病毒之间,交叉反应更为明显。为了降低这种异源特异性,把抗血清先用不同量的异源病毒(0.16~4.0毫克/毫升)处理几小时,然后进行超速离心。这样得到的HR抗血清和γ-球蛋白,对TMV即使有影响也是很小的,而降低HR的侵染性达95%。
从抗TMV血清中,不能分离到类似的选择性抗体。用大量的HR病毒反复处理,能从溶液中除去所有的抗体活性。能与少数HR产生交叉吸附的血清,可以使TMV的侵染性降低97%,对HR的侵染性也有不同程度的降低,平均为44%。庆幸的是,后一种抗体可以使两个病毒小种的血清学鉴定获得明确结果(见表1)。
表1 病毒小种和抗血清的中和及交叉反应
①2~8个实验的平均值,每个测定大约有8枚半片的叶子,每半片叶子上大约有20个病斑。
②与异源病毒产生交叉吸附。
分析方法
为了分析氨基酸,把病毒制品(在0.2~0.4毫升中约含6毫克)与2毫升双蒸过的、恒沸点的盐酸混合,真空密封,在108℃加热16小时。在一个干燥器中将酸反复蒸发后,把水解产物(每毫克病毒约50微克)放在50倍的水中,取一份样品,按最近报道的技术[6 ] ,在纸上作单向层析,测定并分析组氨酸、蛋氨酸、酩氨酸和精氨酸。再取一份样品(1毫克),根据Levy[7 ] 原理使之二硝基苯基化,并进行全氨基酸分析。我们对作为DNP衍生物的氨基酸复原过程中的校正因子作了再次研究,发现了[8 ] 在相似条件下,得到的结果与两年前得到的结果不同。因为只需要比较资料,所以我们未作不同时间的水解实验,而且水解过程中对酸敏感的氨基酸所受到的破坏也未作校正。双DNP胱氨酸是几乎没有的,也没用有关的DNP半胱氨酸的量来说明它。从DNP分析结果来看,对组氨酸、蛋氨酸、酪氨酸和精氨酸的结果重复性不好,在层析分离[6 ] 后用比色分析法被认为是较可靠的。
用Beavan和Holiday[9 ] 提供的分光光度法,对未水解蛋白质制品中的色氨酸和酪氨酸进行了测定。一般来说,酪氨酸的值与比色法[6 ] 得到的值是一致的。TMV和HR的氨基酸分析结果以及从文献中引用的结果,都列于表2。
表2 TMV和HR的氨基酸组分与文献中报道数值的比较①
①所有数值都是以每100克病毒中氨基酸的克数来表示的,对水解期间的破坏未作校正。必须提请注意的是Fraser和Newmark的值是从离子交换柱中回收的物质的百分数。半胱氨酸(约0.6%)未作测定,不列入表中。
②列出的TMV的两排分析数字及平均值引自Knight[3 ] ,Black和Knight[11 ] 发表的资料。而在HR中,第二行数字代表期望值。1953年对HR进行了再次分析,看到的变化与在TMV中见到的变化是相似的。
③Fraser和Newmark[10 ] 。
④不同的蛋白质部分,可能色氨酸含量也不同(2.6%~3.2%),对这种可能性仍在研究中。(见此处 脚注。)
结果
用普通的TMV蛋白质和核酸进行病毒重建
通常所用的重建技术如下:1~10毫克的蛋白质(0.5%~1.0%溶液、pH8)加入十分之一量的核酸和3克分子,pH6的醋酸盐(每毫升反应混合物为10微克)。也常用磷酸盐缓冲液(pH6,8克分子,每毫升50微克),有时还能得到较高产量的活性病毒。低于pH5.0和高于pH8.5,就几乎不能形成有活性的病毒。至少起初几小时,溶液是放在室温下的。取出几小份反应混合物,它很快成为乳白色的,隔不同时间后(例如15分钟、1小时、20小时)经过稀释,再进行分析。根据分析可以看出,有时最大的活性是在很短的反应时间之后得到,但一般来说,大多数情况下,最大活性是在20小时之后得到。偶尔也能见到反应混合物活性下降或消失。这类实验大多数是在核酸真正侵染之前来做的,并认识到在分析培养基中,核酸是不稳定的;这些实验还在重复,并适当考虑了这两类侵染因素的性质。在不同的时间间隔加进RNase,使游离的核酸失活,这样就能用侵染性来测定重建的真实程度。看来,在磷酸盐中,在室温下,反应进行得很迅速,但在醋酸盐中进行,24小时中也能达到同样的程度(见图1)。
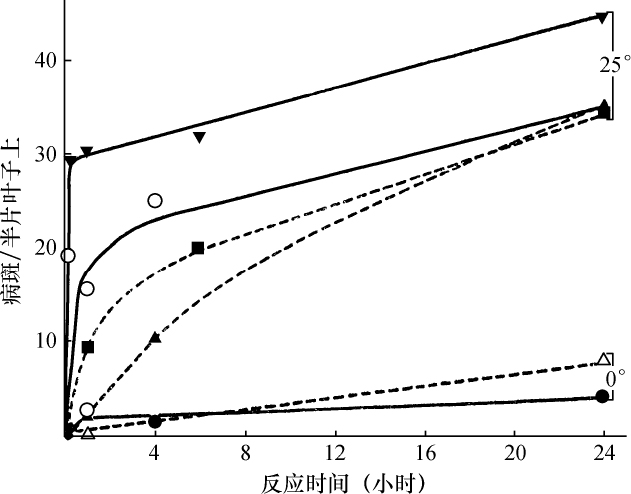
图1 在含有TMV蛋白质(1%)、核酸(0.1%)和缓冲液的反应混合物中,有活性的病毒重建速率分析之前,全部样品都用RNase(每毫升用浓度为0.1%的RNase10微升)处理16小时,使未结合的核酸失活
实线表示用pH7的磷酸盐(0.05克分子)得到的结果;虚线表示用pH6的醋酸盐(0.03克分子)得到的结果。实验Ⅰ(▼■)得到的活性较高,分析时蛋白质浓度为5微克/毫升。最终达到的活性表明了产量达到5%。实验Ⅱ(○、●、▲、△)是在蛋白质浓度为25微克/毫升时分析的。
为了测定在蛋白质和核酸中所含未降解病毒的量,曾经花费了很大的努力。蛋白质制品在通常的测定浓度1~5毫克/毫升时,不产生病斑,即使在浓度为重建制品的100~500倍的水平,在每半片叶子上也只有20~50个病斑。当TMV(0.1~0.5微克/毫升)加入这种蛋白质(1~5毫克/毫升),会得到预期病斑的10%~50%,这表明了在这些条件下,污染的TMV是能致病的。W. Takahashi用一种特殊敏感的分析技术来分析典型的蛋白质制品,他提出在这种蛋白质制品中含有的病毒不到0.0001%。
对含有病毒的核酸制品进行了类似的研究,发现核酸本身就是有侵染性的[2 ,5 ] 。将核酸制品超速离心能十分有效的除去污染的病毒。经过反复的超速离心,一般来说,小于300纳米的大多数棒状颗粒,可以从这种溶液中沉降下来。盐的存在有利于它们的出现,这可能是由于污染了微量蛋白质引起病毒重建所造成的。这些发现为核酸中不存在极微量污染病毒的说法提供了确切证据,这些极微量的污染病毒既不重要,又很难得到。鉴于在最近重建实验中得到了具有很高活性的产物,这种假说的极微量逐渐变得更不重要了。而且,用下面谈到的小种混合物来做的实验,已肯定地表明了重建实验中的活性,在新形成的病毒颗粒中是可遗传的,它不是由于未经降解的病毒颗粒造成的。
混合小种
使用的病毒是通常的TMV以及M、HR、YA小种[3 ] ,从下列组合中能成功地重建出有活性的棒状病毒:①TMV核酸和M蛋白质;②M核酸和TMV蛋白质;③YA核酸和TMV蛋白质;④YA核酸和M蛋白质;⑤HR核酸和TMV蛋白质;⑥TMV核酸和HR蛋白质;⑦HR核酸和M蛋白质。这些混合病毒制品的侵染性和用TMV核酸加上TMV蛋白质得到的病毒的侵染性是一样的。令人惊讶的是,在HR小种中以重量为标准,原来的病毒只有TMV活性的5%。(此重量标准以后称为侵染比活性。)由于HR核酸和TMV核酸的致病性是相同的,因此,HR侵染性较小可能是由于蛋白质成分引起的。这样就有可能得到一种重建的病毒,其侵染比活性比起原来提供核酸的HR小种要高4倍(上面讲的第5种类型的混合病毒,称之为M. V. HR/TMV)。
这些混合病毒实验中,重建病毒与两个亲本小种相比,最有意义的是其生物学性质。当反应产物在两种烟草(N. tabacum和N. sylvestris)上分别测定时,每种情况下都得到与提供核酸的原始小种相同的症状。因此在Turkish烟草中,M. V. TMV/M和TMV/HR都引起绿色花叶病,M. V. M/TMV产生的病毒不引起明显症状,M. V. YA/TMV和YA/M都引起黄色花叶病,而M. V. HR/TMV和HR/M引起典型的环斑。 【80】 这些发现有力地揭示了在TMV和有关的小种中,核酸是遗传决定因素,就像在噬菌体中DNA起的决定作用一样。
混合病毒制品的侵染性是由核酸决定的,而血清学特性好像是由蛋白质成分决定的。当把抗TMV血清或它的γ-球蛋白(见材料和方法)加到这两种制品中去,它中和M. V. HR/TMV与中和TMV的程度一样,但对M. V. TMV/HR无效。相反,抗HR血清,中和M. V. TMV/HR要比中和M. V. HR/TMV有效得多(见表3)。在同样的时间,用同样的方法,检验病斑的性质,清楚地表明M. V. HR/TMV是HR性质的,而M. V. TMV/HR是TMV性质的。对照实验中,在过量同源或异源蛋白质存在的情况下,测定了抗血清对这两种病毒的中和能力,进一步证实了实验程序的可靠性。由此看来,在HR核酸和TMV蛋白质的反应混合物中出现的活性,无疑是由于一种病毒颗粒引起的,这种病毒颗粒含有一种决定遗传性的HR核酸核心和一个决定免疫性的TMV蛋白质外壳。看来没有其他解释可以说明观察到的这个事实。这就明确证实了,从两种化学组分重建起了有侵染性的病毒颗粒。
表3 对TMV、HR和由它们得到的两个混合病毒制品侵染性的中和作用①
①表中全部数字都是表示在粘毛烟草植株上,每半片叶片上的平均病斑数。多数测定是每个样品在8或10枚半片叶片上进行2~3次测定。
从30个实验中选出两个实验列于表中,各次实验都说明了同种现象(尽管其中有些不太完全,另一些表现出较多的交叉反应)。使用的TMV和HR都是5微克,混合病毒制品为20~30微克,γ-球蛋白部分先用异源病毒预处理过(见实验),使用10或20微升,反应混合物的最终体积是0.3毫升。加入到反应混合物中去的蛋白质为25微克。有特异性中和作用的结合,在数字底下划横道表示。
②HR/TMV表示混合病毒是用HR核酸和TMV蛋白质来制备的;TMV/HR表示病毒是用TMV核酸和HR蛋白质制备的。
混合病毒后代的性质
发现TMV核酸可以与HR蛋白质结合,也可以反过来结合,鉴于这两个小种的蛋白质之间有很大差异,因此这是非常令人惊奇的。从Knight[3 ] 的分析看出了在HR和TMV中,只有2~4种氨基酸的数量相同,我们进一步证实了这一事实。Knight所研究过的所有小种,除了HR之外,都不含有组氨酸和蛋氨酸。病毒蛋白质有一个重要的功能特性,即它们能特异性地结合在一个超螺旋排列的核酸链周围(假定它们存在的话)。从本实验看出,不同的病毒蛋白质可以互相交换,因此我们推断病毒蛋白质和核酸结合的活性只取决于在适当位置上的几个关键位点,在这些关键位点周围是非特异性区域。
TMV和HR的蛋白质组成有明显的不同,因此,从这两种病毒得到的混合病毒制品,在研究重建病毒后代的性质方面具有特殊的意义和价值。对TMV、HR以及它们的两种混合病毒制品后代的水解产物,用纸层析进行比较,清楚地说明了在HR和M. V. HR/TMV子代中,约有0.7%的组氨酸和约2%的蛋氨酸,这些氨基酸在TMV和M. V. TMV/HR的后代中是没有的。然后对这4种类型的制品进行了全部的氨基酸分析(见表4)。最初的印象,根据组氨酸和蛋氨酸的存在与否从这些分析中得到了广泛的证实。每种后代的蛋白质与为混合病毒提供核酸的病毒的蛋白质是很相似的。发现它们只有很小的差异(不管哪种氨基酸的含量差异都小于10%)。M. V. HR/TMV 5个不同后代的8种水解产物与HR相比,甘氨酸含量差异很小,而且它们的赖氨酸含量差异也很小,几乎是不变的。对于认为蛋白质对核酸传递的遗传信息也有影响的几种假说而言,这些差异是太小了。必须进一步指出,观察到的这些差异对每个亚基来说,差异还不到一个氨基酸残基,要承认这一点需要假设亚基产生的差异是不一样的。要说明TMV中色氨酸的含量,看来也需要这个假设,因为TMV中每个18000分子量的亚基大约有2.5个色氨酸残基。用化学方法[8 ] 和分光光度法(在0.1mol/L的NaOH中,和在中性pH下[8 ,9 ] 【81】 )两个途径都得到了这个数值,这个数值看来确实要比用微生物学方法得出的数值更可靠些,用微生物学方法得出的结果偏低。
HR和M. V. HR/TMV后代之间的另一个差异是表现在侵染比活性上,多数M. V. HR/TMV的侵染比活性要比任何HR样品所得到的最高值还要高(20%~40%比12%)。
表4 HR、TMV与实验制品后代的氨基酸组分的比较①
①所有数值都表示每100克病毒中氨基酸的克数。此值对水解期间的破坏未做过校正。前2行数值是8个水解产物的平均值,后3行数值是3个水解产物的平均值。
看到有明显差异的,则在数字下面画横道表示。
原来的病毒小种和混合病毒制品之间这些差异的意义,将依赖于这些差异在后代制品中是否会连续出现,这好像是侵染比活性问题,但至今尚未完成对连续的单个病斑后代制品的足够数目的氨基酸分析,这一工作的成功将使有关这项工作的明确结论获得公认。目前,从这些工作中人们唯一可得出的结论是每个小种的核酸,在寄主细胞内能促进新病毒蛋白质的合成,这种新的病毒蛋白质与它本身同源的蛋白质尽管不是完全一致,但也是十分相似的。即使在离体条件下把核酸裹入另一种病毒小种的蛋白质中,它仍保留着这种能力。
可遗传的饰变
从混合病毒或用来分离核酸的病毒的单个病斑上分离它们的子代病毒,然后观察这些子代病毒的蛋白质组成及其对植物的致死性,曾不止一次见到有显著的变化。对这种表现型突变体,就其后代病毒在病症学和氨基酸组成两个方面的差异进行了鉴定(见表4),认为它的出现表明了遗传物质通过化学处理和人工操作容易发生变化,因为从原来病毒小种的单个病斑繁殖病毒时,没有发现类似的变化。与其他遗传学上较容易变化的例子不同,这个突变小种通过几次传代和分离,包括分离它的核酸和蛋白质,在Turkish烟草中还能继续产生明显的坏死病症。
摘要
(1)叙述了从TMV和其他小种制备天然蛋白质的方法。
(2)用不同小种的蛋白质和核酸来进行病毒颗粒的重建,已得到了活性很高的制品,其中有一种制品表现出比其亲本小种之一更高的侵染性。
(3)由混合病毒制品引起的病症的性质,在任何情况下都与提供核酸的病毒特征相似。
(4)混合病毒制品后代的化学性质也与提供核酸的病毒很接近(尽管对氨基酸组分微小差异的意义何在,尚未弄清)。
(5)混合病毒制品的血清学特性与上述的性质相反,它是取决于提供蛋白质的那些病毒。
(6)明确证实了重建病毒颗粒活性上的这种双重性。
(7)在这个工作过程中,具有不同的生物学和化学特性的变种或突变体会随机地产生,这表明遗传物质通过化学操作后变得不稳定了。
参考文献
[1 ]Fraenkel-Conrat, H. and Williams, R. C., 1955. Proc. Natal. Acad. Sci. USA, 41 , 690.
[2 ]Fraenkel-Conrat, H., 1956. J. Am. Chem. Soc., 78 , 882.
[3 ]Knight, C. A., 1947. J. Biol. Chem., 171 , 297.
[4 ]Niu, C. I., and Fraenkel-Conrat, H., 1955. Arch, Biochem. Biophys., 59 , 538.
[5 ]Fraenkel-Conrat, H., Singer B., and Wiliams, R. C., 1957. Biochim. Biophys. Acta, 25 , (in press); see also: Symposium on the Chemical Basis of Heredity, McCollum Pratt Institute, Johns Hopkins University, Baltimore, June 1956.
[6 ]Fraenkel-Conrat, H., and Singer, B., Arch. Biochem. Biophys., (in press).
[7 ]Levy, A. L., 1954. Nature, 174 , 126.
[8 ]Fraenkel-Conrat, H., and Singer, B., 1956. Arch. Biochem. Biophys., 60 , 64.
[9 ]Beavan, G. H., and Holiday, E. R., 1952. Advances in Prot. Chem., 7 , 320.
[10 ]Fraser, D., and Newmark, P., 1956. J. Am. Chem. Soc., 78 , 1588.
[11 ]Black. F. L., and Knight, C. A., 1953. J. Biol. Chem., 202 , 51.
脱氧核糖核酸的生物合成 【82】
阿瑟·科恩伯格(A. Kornberg) 【83】
(美国 斯坦福大学医学院生化系,1960年)
近年来,通过对细菌转化[1 ] 和病毒侵染细菌[2 ] 的了解,再结合其他方面的证据[3 ] ,几乎使我们大部分人都确信脱氧核糖核酸(DNA)是遗传物质。我们认为,DNA不仅指导蛋白质的合成和细胞的发育,而且还能够复制,通过复制使细胞世世代代都保持相似的发育。DNA就像一条录音带一样,携带着一种信息,里面有进行工作的特异性指令,而且,也像录音带可进行拷贝一样,DNA也可以进行精确的复制,以便这种信息能在另外的时间和空间被再次使用。
这两种功能——密码的表达(蛋白质合成)和密码的复制(物种的保存)是紧密相关的呢?还是各自独立的呢?在过去5年多的时间里,我们从研究中得知,尽管在细胞中DNA如何指导蛋白质合成的奥秘仍不清楚,但DNA的复制是可以检验出来的,至少可在酶促水平上部分地了解。
结构
首先我想简短地评述一下DNA结构的某些方面,这对本讨论是很必要的。很多研究工作者对多种来源的DNA样品进行了组成分析[4 ] ,揭示了一个明显的事实,即DNA中嘌呤含量和嘧啶含量总是相等的。嘌呤中,腺嘌呤(A)和鸟嘌呤(G)含量可以有明显差异;在嘧啶中,胸腺嘧啶(T)和胞嘧啶(C)含量可以明显不同。然而,在环的6-位上带有一个氨基基团的碱基与在6-位上带有一个酮基基团的碱基是相等的。沃森(Watson)和克里克(Crick)[5 ] 在DNA结构的巧妙假说中说明了这些事实。如图1所示,在他们的DNA双螺旋模型(下面还将讨论)中,腺嘌呤的6位-氨基基团是通过氢键与胸腺嘧啶的6位-酮基基团连接的,鸟嘌呤与胞嘧啶也是通过氢键以同样方式连接的,由此说明嘌呤与嘧啶是相等的。
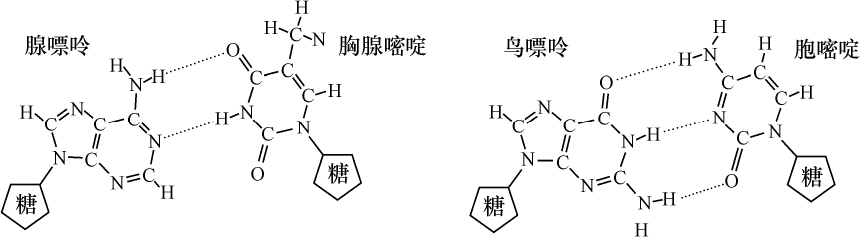
图1 碱基间的氢键结合
考虑到这些情况,以及Wilkins与他的合作者[6 ] 所做的X射线结晶学测定结果,沃森和克里克提出了一种DNA结构,在这种结构中,两条长链以一种螺旋方式互相缠绕着。图2是长度约为10个核苷酸单位的一段DNA链的图示。根据物理学测定来看,DNA链的平均长度约有10000个核苷酸单位。这里我们可以看到,脱氧戊糖环通过磷酸残基连接起来,形成主链,嘌呤和嘧啶环在与链的主轴垂直方向上呈平面结构。图3是较详细的分子模型[7 ] ,并表示在这种结构中,原子的一种理想布局。一个链的嘌呤和嘧啶碱基按图1所示的方式通过氢键与互补链的嘧啶和嘌呤结合。
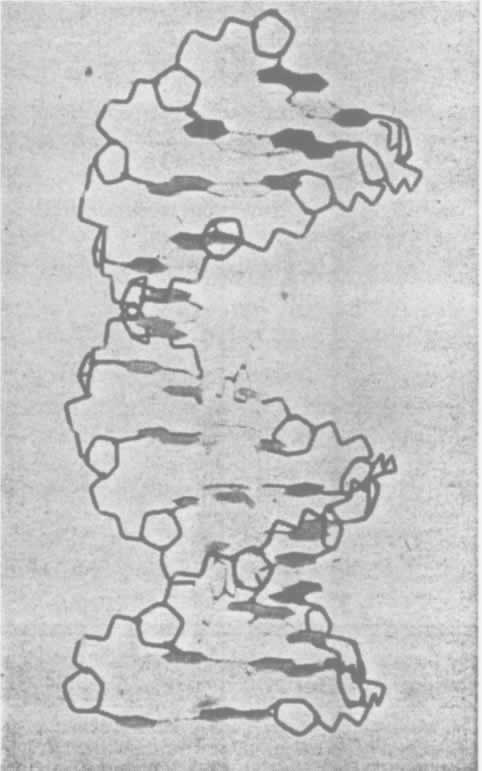
图2 DNA的双螺旋结构
(Watson和Crick模型。)
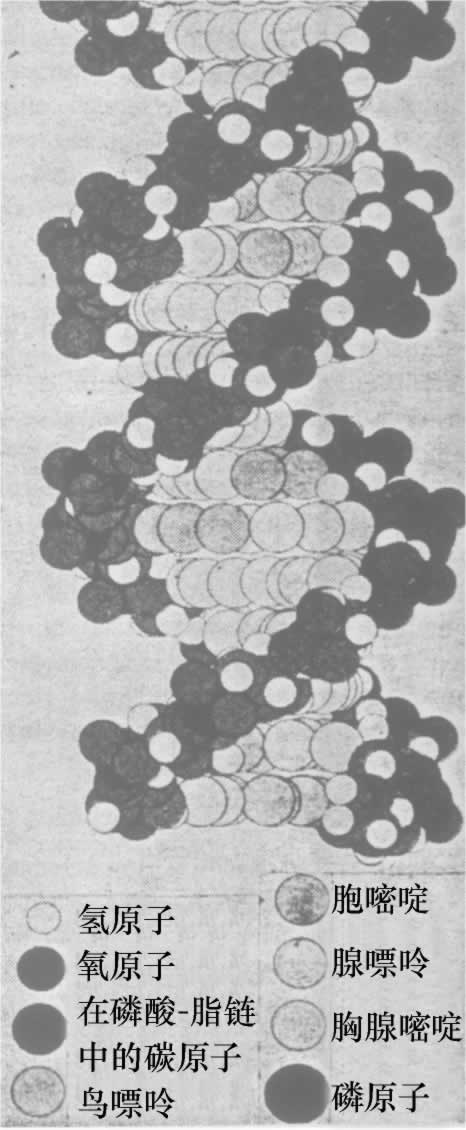
图3 DNA的分子模型
(M. Feughelman等[7 ] 。)
X射线测定已证明,在这个模型中,两条相对的链之间的空间与通过氢键连接的一个嘌呤和一个嘧啶间的计算值是一致的;这个值对两个嘌呤来说是太小了,对两个嘧啶来说是太大了。从生物学的观点来说,这种结构的重要性就是为说明DNA在细胞内是如何复制的,提供了一种有用的模型。我们推测,在复制时两条链分开,其中每一条链都能形成一条互补的新链,结果就形成了两条双链,每一条双链与原来的亲本双链是相同的,而且这两条双链的成分是完全相同的。
复制的酶促途径
虽然我们知道沃森和克里克提出了一种DNA复制机制的模型,但我们在这一点上还有一个疑问,即在细胞中,建成这种高分子的化学机制是什么?大约60年前,用酵母细胞进行蔗糖酒精发酵,被认为这是一种与活细胞不可分离的有生命过程,但是通过Buchner发现了提取液有发酵作用,以及20世纪上半叶酶学的进展,我们了解到酵母的发酵作用是一系列(现已熟悉)完整的化学反应的结果。
5年前,DNA的合成也被认为是一种有生命的过程。有些人认为,它对于生化学家测定细胞的氧化穴位(combustion chambers)是有用的,但是,遗传结构受损的本身除了引起失调外,绝对不产生其他影响。这些渺茫的预言没被证实,现在摆在我们面前的关于细胞结构和特异性功能问题,不是这样的悲观情况。在酶学方面,前面有着艰巨的任务,在碳水化合物、脂肪、氨基酸和核苷酸酶学方面受过训练的许多人员将从事开拓这项工作。
现在我感到,(正如我们已经做的)解决核酸生物合成问题的有效途径,最根本的是要弄清单个核苷酸和辅酶的生物合成,并且还要很好地掌握这些概念和方法。从这些研究中,我们确信,一种被激活的5′-磷酸核苷是核酸的基本生物合成单位[8 ] 。应再重复一次:嘌呤和嘧啶生物合成的主要途径都是形成这种5′-磷酸核苷[8 ] ;除了补救机制之外,他们一般不含有游离的碱基或核苷酸。核苷酸的2′和3′异构体可能主要是由于核酸经某些种类的酶促降解而引起的。从最简单的核苷酸缩合产物——辅酶的生物合成[9 ] ,可以联想到腺苷三磷酸(ATP)与烟酰胺单核苷酸缩合形成二磷酸吡啶核苷酸(辅酶I)、与核黄素磷酸缩合形成黄素腺嘌呤二核苷酸(FAD)、与泛酰巯基乙胺磷酸缩合形成辅酶A的前体物等等。由于发现脂肪酸和氨基酸的激活作用机制是一样的,这个模型得到了详细描述,并且进一步证明了尿苷、胞苷和鸟苷的辅酶都是由这些核苷各自的三磷酸盐形成的。
如图4所示的方式,核苷-磷酸在活化的焦磷酸腺苷基团上发生亲核反应[10 ] ,导致辅酶的形成,现已采用这种机制作为研究一条DNA链合成的工作模型。按图5所示可以推断:DNA的基本单位是脱氧核苷-5′-三磷酸,它与一条多聚脱氧核苷酸链的延伸末端上的3′-羟基起反应;除去无机的焦磷酸盐,这样,核苷酸就加长了一个单位。我们对DNA合成的研究结果(见下面的叙述)是与这种反应类型一致的。
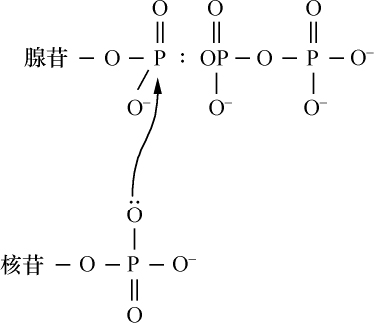
图4 核苷-磷酸在ATP上发生的亲核反应
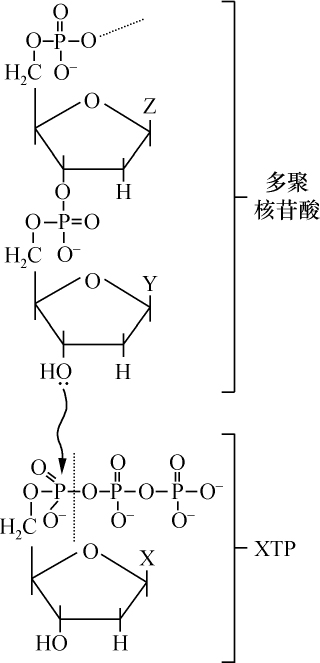
图5 一条DNA链延伸的假设机制
酶的性质
首先让我们讨论酶和评论发现这种酶的方法[8 ,11 ] 。将通常在DNA中出现的4种三磷酸脱氧核苷与胸腺、或与骨髓、或与大肠杆菌的提取液一起混合,都不能引起DNA的纯合成。相反,正如所预料的一样,在这个过程中,这些细胞和组织的提取液对DNA的降解却是占绝对优势的。人们只有凭借较灵敏的设计,才能测定这样一种生物合成反应。我们采用了一种14 C标记的放射比活性很高的底物,将它与腺苷三磷酸和大肠杆菌(一种每20分钟便可繁殖一代的细菌)提取液一起保温。最初得到的正结果表明,酸溶性物质只有很小部分(占加入量的十万分之五左右)变成酸不溶的。这说明起反应的只有几微微克分子,但这是重要的。我们就试图通过这个很小的缝隙,打进一个楔子,所用的锤子便是酶的纯化[12 ] 。
酶的纯化是当务之急。我们最好的酶制品,就蛋白质而言已比粗提液浓缩了几千倍,但仍污染有一定数量的、在大肠杆菌中存在的一种或多种核酸酶或二酯酶。在动物细胞和其他种类的细菌中,也观察到了一种类似的DNA合成系统[13 ] 。要与大肠杆菌系统进行可靠的比较,还必须对这些来源的酶进行纯化[12 ] 。
在图6的反应式中,表示出了用纯化过的大肠杆菌的酶来进行DNA纯合成的要求[14 ] 。形成A—T和G—C碱基对的4种脱氧核苷酸都必须存在。底物必须是三磷酸盐,而不能是二磷酸盐,而且只有脱氧糖化合物才是有活性的。脱氧核糖核酸必须存在,它可以从动物、植物、细菌或病毒中得到,所有这些DNA样品在DNA合成中都同样有效,而且合成的DNA分子量都是很大的。这种产物(下面我还将详细讨论)不断积累,直到其中一种底物用完为止,在数量上可能会达到加入DNA的20倍或更多。因此使加入到反应混合物中的底物的95%或更多,都形成了DNA。释放出的无机焦磷酸盐,数量上与转变成DNA的脱氧核苷酸是等克分子的。
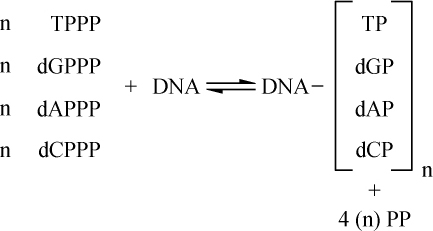
图6 酶促合成DNA的反应式
要是这些底物中的某一种被省去,反应的进行要降低104 倍以上,那么就需要用特异的方法才能将它检出。其原因是当一种脱氧核苷酸底物缺少时,极少的但仍有显著数量的核苷酸与DNA引物连接。我和我的合作者对这种所谓的受限反应[15 ] 已有叙述,并证明在这种情况下,只有少数脱氧核苷酸被加到其中某些DNA链的核苷末端上去,但是终因缺少这种省去的核苷酸而使进一步的合成停止了。最近的研究揭示了,这种受限反应相当于在长短不等的两条单链组成的一条双螺旋中,较短的那条单链上发生的修复作用;还说明这个反应受到A—T或G—C碱基对氢键的控制。
当4种三磷酸盐都存在、但无DNA时,反应根本不发生。为什么需要DNA呢?DNA的功能是以糖原的方式作为一种引物呢?还是作为模板指导它本身的精确复制物的合成呢?我们有充分理由相信后者是正确的,这也是本文的中心议题。应当强调一下,反应中原有的DNA和作为底物加入的核苷酸之间能通过氢键进行碱基配对,这就是反应需要DNA的原因。
根据我们的经验,迄今为止,我们正在研究的这种酶是唯一能从模板接受指令的,这种酶的作用是把能与模板上的一个碱基形成碱基对(通过氢键结合)的那种特殊的嘌呤或嘧啶底物加到模板上去(见图7)。主要有5个方面的证据支持这篇论文。
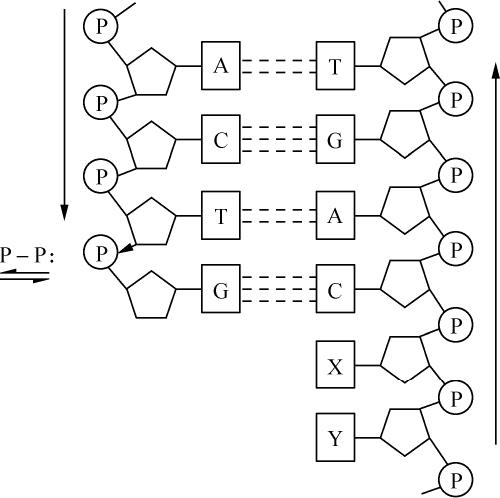
图7 DNA的酶促复制机制
酶促合成的DNA的物理性质
第一方面的证据是从研究由这种酶催化产生的DNA的物理性质中得到的。这些叙述以及对DNA化学性质(下面将讨论)的叙述中,有90%到95%的DNA样品来自这个反应所用的底物。在与H. K. Schachman的合作研究中,我们从他那儿得到很大帮助。据说,这种酶促产物与从天然来源分离到的高分子量的双链DNA是没有差别的[16 ] 。它的沉降系数约为25,黏度为每克40分升,根据这些测定,我们认为它是很长的直棒状的,分子量约为600万道尔顿。当这种DNA受热时,棒状散开,分子变成一种致密的、随机卷曲的结构;推断这时把两条链连在一起的氢键已消失,这可以从这些分子黏度和光学性质上的典型变化得到证明。用胰脱氧核糖核酸酶分解这种分子,也得到类似的结果。在所有这些方面,酶促合成的DNA与从天然来源分离到的DNA是没有差别的,因此可以推断它有一种与天然DNA相同的氢键结构。
DNA经热处理后解离出来的这种混乱的DNA链,是否可以作为DNA合成的一种引物呢?按平常的经验来看,以一种混乱的缠绕链来指导复制,人们很可能认为它不可能作为模板。现在搞清楚了,散开的DNA是一种极好的引物,这种不黏滞的、随机缠绕的单链DNA可导致合成高度黏滞的、双链DNA[17 ] 。Sinsheimer已从极小的 X174病毒中分离到一种似乎是单链的DNA[18 ] 。已证明它像热处理过的DNA一样,是一种极好的引物[17 ] ,而且是目前一种很有用的研究材料[19 ] ,用密度梯度沉降证明,在酶促合成过程中,它能逐步转变成一种双链状态。
在这篇文章中,要对复制的物理学方面做详细的讨论是不可能的,但应当提及,这种DNA在单链状态下,不仅是一种很合适的引物,而且当使用很纯的酶制品时,只有单链形式才有活性。使用大肠杆菌的这种纯酶制品时,天然的、双链DNA是没有活性的,除非这种DNA经过热处理,或用脱氧核糖核酸酶稍微预处理一下才行。Bollum用他自己从小牛胸腺提纯出来的这种酶,也观察到了同样的结果[20 ] 。
类似物的替代
第二方面的证据是当嘌呤和嘧啶碱基被替代时,从研究底物的活性得到的。从有关溴尿嘧啶[21 ] 、氮鸟嘌呤[22 ] 和其他的类似物参入到细菌和病毒DNA中去的许多有趣报告可以推断出碱基结构中可以容许有些自由,并证明这种自由对它们的氢键没有妨碍。发现用脱氧尿苷三磷酸或5-溴尿苷三磷酸代替胸苷三磷酸时,DNA合成反应能够完成,但当代替脱氧腺苷、脱氧鸟苷、或脱氧胞苷三磷酸时,DNA的合成反应就不能完成。如前所述[23 ] ,5-甲基-胞嘧啶和5-溴胞嘧啶能特异地代替胞嘧啶;次黄嘌呤只能代替鸟嘌呤;还有刚才提到的,尿嘧啶和5-溴尿嘧啶能特异地代替胸腺嘧啶。这些发现从A—T、G—C形成氢键的方式,可以得到很好的解释。
和这些证据有关的一件事还应当提一下,有一种天然存在的胞嘧啶类似物——羟甲基胞嘧啶(HMC),发现在大肠杆菌的T系偶数列噬菌体DNA中,这种HMC代替了胞嘧啶[24 ] 。这种情况下,DNA含有等量的HMC和鸟嘌呤,并与通常一样,腺嘌呤和胸腺嘧啶也是等量的。有趣的是,虽然在噬菌体T2和T6的DNA中,有些HMC基团是不含葡萄糖的[26 ] ,但噬菌体T2、T4和T6的DNA却含有一定比例的与HMC的羟甲基基团相连的葡萄糖[25 ,26 ] 。
这些特性在有关DNA合成方面提出了两个似乎是与单个碱基配对的假说互相矛盾的问题。首先,在正常条件下,细胞含有脱氧胸苷三磷酸,并能参入细胞的DNA,有一种什么机制,能阻止一个细胞中含有胞嘧啶呢?第二,假如参入作用是经过葡萄糖基化或非葡萄糖基化的HMC核苷酸发生的,那么在DNA中葡萄糖和HMC的恒定比例又是怎么产生的呢?我们最近的实验已经证明,在病毒侵染过的细胞中,多聚酶反应仍是通过正常的氢键限制来控制的,但是还有几种新酶的辅助作用,这些新酶是随着一种病毒的侵染,特异性地产生的[27 ,28 ] 。在这些新酶中,有一种酶能把脱氧胞苷三磷酸切开,这样就能把它从多聚酶作用的部位上除去[28 ] 。另外,还有一类葡萄糖基化酶,它能从尿苷二磷酸葡萄糖上直接而特异地将葡萄糖转移到DNA的某些HMC基团上[28 ] 。
化学组成
第三方面的证据是在分析酶促合成的DNA的嘌呤和嘧啶碱基组成的过程中得到的。我们可以提出两个问题:第一,这种产物含有的腺嘌呤与胸腺嘧啶,鸟嘌呤和胞嘧啶相等吗?因为这是天然DNA的典型特性。第二,用作引物的天然DNA的组成,能影响和决定这种产物的组成吗?表1的结果回答了这两个问题[29 ] 。实验中,分别使用草分枝杆菌(Mycobacterium phlei)、大肠杆菌、小牛胸腺和T2噬菌体的DNA做引物,各种情况下除了所用的这些DNA引物不同外,其他方面都是一样的。
对于第一个问题的回答是很清楚,在酶促合成的DNA中,腺嘌呤等于胸腺嘧啶,鸟嘌呤等于胞嘧啶,所以在每种情况下,嘌呤与嘧啶的含量都是一样的。对第二个问题的回答是很明显,一种给出的DNA引物,其A—T对与G—C对的特异性比值,对合成的产物有绝对的影响。用同位素示踪方法来测定时,纯DNA的增加,不论是在仅仅为1%时,还是达到1000%,结果都是这样的。
采用克分子浓度大不相同的底物,或用任何其他手段,都不能破坏这些碱基的比例。表1的最后一行是一种很新奇的DNA,对它的合成条件,这里我们就不叙述了[17 ,30 ] 。只说一下,经过很长的延迟期以后,产生了一种脱氧腺苷酸和胸苷酸(A—T)的共聚物,它的大小和物理学性质都和天然DNA一样,而且其中的腺嘌呤和胸腺嘧啶是以一种分毫不差的交替顺序排列的。当用这种类似DNA的多聚体的稀有形式作为引物时,新的A—T多聚体的合成可立即开始,而且即使4种三磷酸核苷都存在,在产物中也测不出有微量的鸟嘌呤和胞嘧啶存在。由此看来,只能得出这样的结论:在酶促合成过程中,碱基组成被复制了,而且腺嘌呤和胸腺嘧啶,鸟嘌呤和胞嘧啶之间形成氢键是复制的指导机制。
表1 采用不同的引物时,酶促合成的DNA化学组成
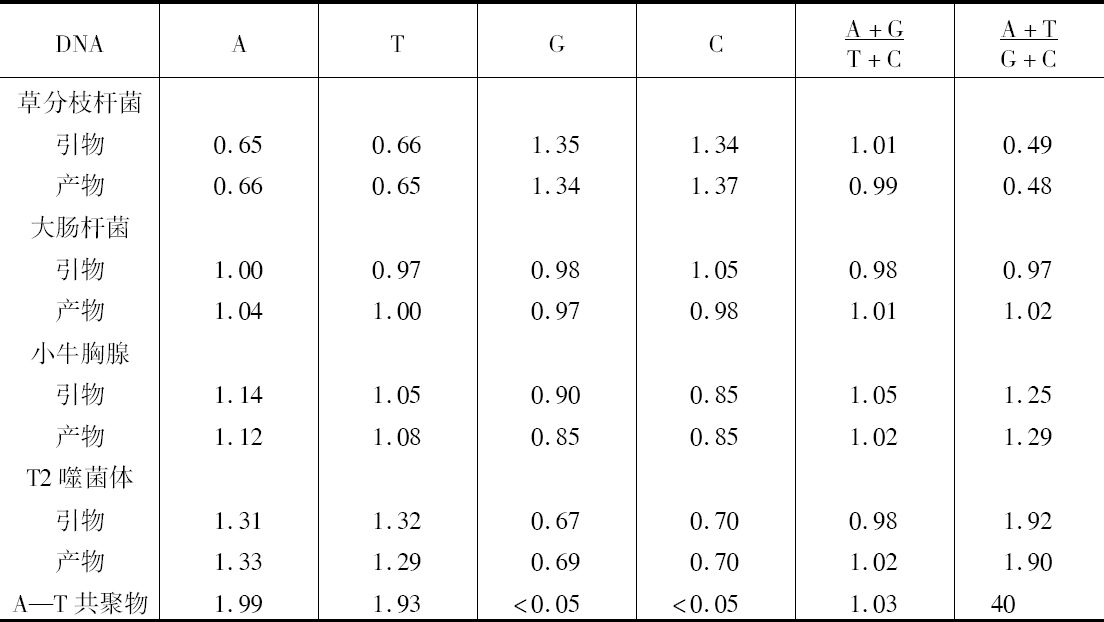
核苷酸顺序的酶促复制
我想引证的第四方面的证据是从最近对于DNA的碱基顺序及其复制的研究中得到的。我已经谈到,我们相信DNA是遗传密码;4种核苷酸组成一种4个字母的文字,它们的顺序拼成信息。现在,我们还不了解这种顺序;Sanger已经做出了蛋白质中肽的顺序,正在做核酸顺序方面的工作。这个问题是比较困难的,但并不是不可解决的。
有关我们最近测定核酸顺序的工作情况,将在别的文章中详细叙述[31 ] ,这里只概述一下这些工作。脱氧核糖核苷酸是酶促合成的,其中的一种脱氧核苷三磷酸是用32 P来标记的;其他3种底物未经标记。这种放射性磷原子附着在脱氧核糖5位-碳原子上,成为底物分子和链的延伸末端的核苷酸分子之间起反应的桥梁(见图8)。在合成反应结束时(大约形成1016 个二酯键),分离出DNA,并把它用酶降解,就可得到大量的3′-脱氧核苷酸。很明显(见图8),原来吸附在脱氧核苷三磷酸底物5位-碳原子上的磷原子,现在吸附到了在DNA链合成过程中与它起反应的那个核苷酸的3位-碳原子上。由纸电泳分离出来的每种3′-脱氧核苷酸的32 P含量,就是衡量在DNA合成过程中,4种有效核苷酸中的每一种与特异性底物起反应的相对频率的尺度。在这个过程中,每种情况下,用不同的标记底物进行4次实验,就能得到16种可能的二核苷酸(紧挨着的)顺序的相对频率。
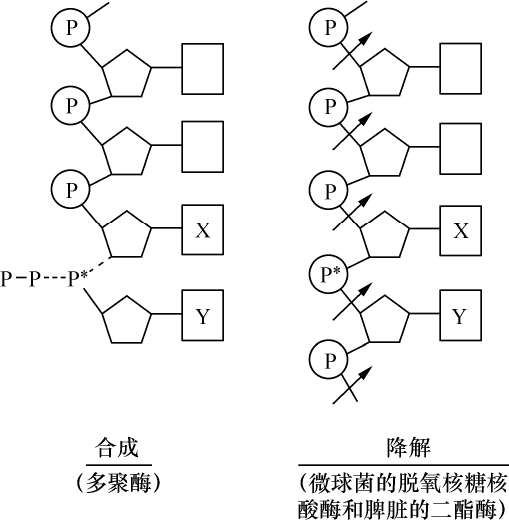
图8 测定DNA顺序的方法
迄今为止,已经用6种来源不同的天然DNA样品作引物,进行了研究。结论如下:
(1)每种情况下都有16种可能的二核苷酸顺序。
(2)每种情况下,这些顺序的相对频率的图形都是一致的、可以重复的,而且不能从DNA的碱基组成来预言。
(3)酶促复制过程与A—T和G—C的碱基配对有关。
(4)最重要的是,这些频率清楚地表明了酶促复制过程产生具有相反方向的两条链,这两条链完全符合Watson和Crick的模型。
从这些研究和预先讨论过的范围来看,在酶促复制过程中,能作为有效引物的任何一种DNA样品的二核苷酸频率,都应当能得到,这就为译解DNA密码提供了线索。不幸的是,这种方法不能提供有关三核苷酸频率的线索,但我们确信,随着分析工作中酶学工具和分离工作中层析技术的发展,在这方面可以迈出第一步。
DNA合成需要4种三磷酸盐和DNA
DNA合成需要有4种脱氧核苷三磷酸和DNA,这在前面已经谈到,现在我们可以认为这种需要是形成氢键第五方面的证据。不加DNA就没有形成氢键的模板;4种三磷酸盐都不加,就会因为在模板中一个碱基没有通过氢键配对,而使合成反应立即停止。
摘要
概述了DNA复制的酶促途径以及从大肠杆菌中纯化出的DNA合成酶的性质。关于这种酶作用的特异性和普遍性指的是它能在一条DNA模板的指导下,催化合成一条新的DNA链;DNA模板的指导作用是由A—T和G—C通过氢键形成碱基对的规律来支配的。这个结论是从下列实验观察中得出的:
(1)酶促合成的DNA具有双链特性,它来源于一条单链分子。
(2)碱基类似物替换天然碱基的情况。
(3)化学组分的复制。
(4)核苷酸(紧挨着的)顺序的复制,以及两条链方向的相反平行。
(5)DNA合成必须要4种脱氧核苷三磷酸(A、T、G、C)和DNA都存在。
参考文献
[1 ]Avery, O. T., MacLeod, C. M., McCarty, M., 1944. J. Exptl Med., 79 , 137; Hotchkiss, R. D., 1957 in the Chemical Basis of Heredity, McElroy, W. D., and Glass, B., Eds. (Johns Hopkins Press, Baltimore), 321.
[2 ]Hershey, A. D. 1953. Cold Spring Harbor Symposia Quant. Biol., 18 , 135.
[3 ]Beadle, G. W. 1957. In The Chemical Basis of Heredity, McElroy, W. D., and Glass, B. Eds. (Johns Hopkins press, Baltimore), 3.
[4 ]Chargaff, E., 1955. In Nucleic Acids, Chargaff, E., and Davidson, J. N. Eds., (Academic press, New York), 1 , 307~371.
[5 ]Watson, J. D., and Crick, F. H. C., 1953. Nature, 171 , 737; 1953. Cold Spring Harbor Symposia Quant. Biol., 18 , 123.
[6 ]Wilkins, M. H. F., 1957. Biochem. Soc. Symposia (Cambridge, Engl.), 14 , 13.
[7 ]Feughelmen, M., Langridge, R., Seeds, W. E., Stokes, A. R, Wilson, H. R., Hooper, C. W., Wilkins, M. H. F., Barclay, R. K., and Hamilton, L. D., 1955. Nature, 175 , 834.
[8 ]Kornberg, A., 1957. in The Chemical Basis of Heredity, McElroy, W. D., and Glass, B. Eds., (Johns Hopkins Press, Baltimore), 579; 1959. Revs. Modern Phys., 31 , 200.
[9 ]Kornberg, A., 1951. in Phosphorus Metabolism, McElroy, W. D., and Glass, B. Eds. (Johns Hopkins Press, Baltimore), 392; 1957. Advances in Enzymol., 18 , 191.
[10 ]Koshland, D. E, Jr., 1954. in The Mechanism of Enzyme Action; McElroy W. D. and Glass, B. Eds., (Johns Hopkins Press, Baltimore), 608.
[11 ]Kornberg, A., Lehman, I. R., and Simms, E. S., 1956. Federation Proc., 15 , 291; Kornberg, A., 1957—1958. Harvey Lectur, 53 , 83.
[12 ]Lehman, I. R., Bessman, M. J., Simms, E. S., and Kornberg, A., 1958. J. Biol. Chem., 233 , 163.
[13 ]Bollum, F. J., and Potter, V. R., 1957. J. Am. Chem. Soc., 79 , 3603; Harford, C. G., and Kornberg, A., 1958. Federation Proc.,17 , 515; Bollum, F. J., 1958. Ibid., 17 , 193; 1959 Ibid., 18 , 194.
[14 ]Bessman, M. J., Lehman, I. R., Simms, E. S., and Kornberg, A., 1958. J. Biol. Chem., 233 , 171.
[15 ]Adler, J., Lehman, I. R., Bessman, M. J., Simms, E. S., and Kornberg, A., 1958. Proc. Natl. Acad. Sci. USA, 44 , 641.
[16 ]Schachman, H. K., Lehman, I. R., Bessmen, M. J., Adler, J., Simms, E. S., and Kornberg, A., 1958. Federation Proc., 17 , 304.
[17 ]Lehman, I. R., 1959. Ann. N. Y. Acad, Sci., 81 , 745.
[18 ]Sinsheimer, R. L., 1959. J. mol. Biol., 1 , 43.
[19 ]Lehman, I. R., Sinsheimer, R. L., and Kornberg, A., unpublished observations.
[20 ]Bollum, F. J., 1959. J. Biol. Chem., 234 , 2733.
[21 ]Weygand, F., Wacker, A., Dellweg, H., 1952. Z. Naturforsch.,7b , 19; Dunn, D. B., and Smith, J. D., 1954. Nature, 174 , 305; Zamenhof, S. and Griboff, G., 1954. Ibid., 174 , 306.
[22 ]Heinrich, M. R., Dewey, V. C., Parks, R. E. Jr., and Kidder, G. W., 1952. J. Biol. Chem., 197 , 199.
[23 ]Bessman, M. J. Lehman, I. R. Adler, J., Zimmerman, S. B., Simms, E. S., and Kornberg, A., 1958. Proc. Natl. Acad. Sci. USA, 44 , 633.
[24 ]Wyatt, G. R., and Cohen, S. S., 1953. Biochem. J., 55 , 774.
[25 ]Sinsheimer, R. L., 1954. Science, 120 , 551; Volkin, E., 1954. J. Am. Chem. Soc., 76 , 5892; Streisinger G., and Weigle, J., 1956. Proc. Natl. Acad. Sci. USA, 42 , 504.
[26 ]Sinsheimer, R. L., 1956. Proc. Natl. Acad. Sci. USA, 42 , 502; Jesaitis, M. A., 1957. J. Exptl. Med, 106 , 233; 1958. Federation Proc., 17 , 250.
[27 ]Flaks, J. G., and Cohen, S. S., 1959. J. Biol. Chem., 234 , 1501; Flaks, J. G., Lichtenstein, J., and Cohen, S. S., 1959. Ibid., 234 , 1507.
[28 ]Kornberg. A., Zimmerman, S. B., Kornberg, S. R., and Josse, J., 1959. Proc. Natl. Acad Sci. USA, 45 , 772.
[29 ]Lehman, I. R., Zimmerman, S. B., Adler, J., Bessman, M. J., Simms, E. S., and Kornberg, A., 1958. Ibid., 44 , 1191.
[30 ]Radding, C. M., Adler, J., and Schachman, H. K., 1960. Federation Proc., 19 , 307.
[31 ]Josse, J., and Kornberg, A., 1960. Ibid., 19 , 305.
蛋白质遗传密码的一般性质 【84】
弗朗西斯·克里克(F. H. C. Crick) 【85】 莱斯利·巴奈特(F. R. S. L. Barnett)
S. 布里纳(S. Brenner) 瓦特·托宾(R. J. Watts-Tobin)
(英国 剑桥Cavendish实验室,1961年)
现在有许多间接证据说明,蛋白质多肽链上的氨基酸顺序,是由遗传物质核酸的某个特殊部分中碱基的顺序决定的。因为自然界中常见的氨基酸有20种,但常用的碱基只有4种,因此推测,4种碱基的顺序,以某种方式为氨基酸的顺序编码。从本文报道的遗传实验,再结合以前别人的工作可以看出,遗传物质具有以下几种普遍性:
(1)3个碱基一组(不太可能是3个碱基的倍数)编码一个氨基酸。
(2)密码是不重叠的(见图1)。
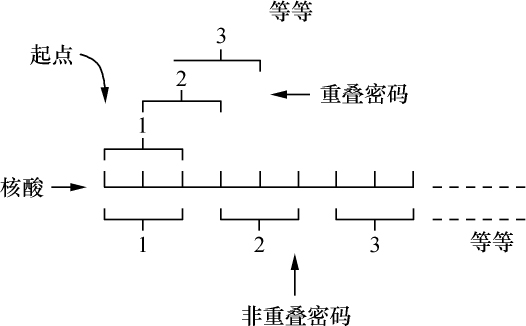
图1 重叠密码和非重叠密码差异的图
垂直短线表示核酸的碱基。图中的情况是以三联体密码来说明的。
(3)碱基的顺序是从固定起点解读的,这就决定了,一段很长的碱基顺序是如何以三联体的形式被正确解读的。没有特异性逗号来说明如何选择正确的三联体。如果起点被一个碱基替换了,那么解读出来的三联体也就被替换了,这样就产生了错误。
(4)密码是简并的,一般说来,即某个特定的氨基酸可以由几个碱基三联体中的一个来编码。
密码的解读
说明遗传密码是不重叠的有关证据(见图1),不是来自我们的工作,而是从Wittmann[1 ] 、Tsugita和Fraenkel-conrat[2 ] 用亚硝酸诱发TMV产生的突变体的研究中得到的。在一个重叠的三联体密码中,改变一个碱基,一般将使多肽链上3个相邻的氨基酸发生改变。他们还研究了病毒蛋白质中产生的改变,结果说明用亚硝酸处理病毒的RNA,通常处理一次,只有一个氨基酸发生改变,在极少情况下,有两个氨基酸发生改变(可能是由于亚硝酸在一段RNA上分别两次脱氨基作用造成的),发生改变的这两个氨基酸,在多肽链上的位置是不相邻的。
Brenner[3 ] 以前曾指出,如果密码是通用的(即整个自然界都相同),那么重叠的三联体密码是不可能有的。而且,已经详细研究过的、不正常的人类血红蛋白,只有一种氨基酸发生了改变[4 ] 。新近的实验结果基本上排除了重叠类型的所有简单密码。
如果密码是不重叠的,那么必然有某种排列方式可以表示出,沿着连续的碱基顺序,如何选择正确的三联体(或者四联体,或者任何一种可能的几联体)。有一种意见认为,每4个碱基一组。另一种意见认为,就像在Crick、Griffith和Orgel[5 ] 的不分组密码子中的情况那样,某些三联体表达一个意思,而另一些三联体则没有意义。从一个固定点开始,一次按3个(或4个、或任何几个)碱基的顺序进行工作,这样就可能作出正确的选择,得知那种排列方式是正确的。这是可能做到的事情,我们也赞成这种做法。
实验结果
噬菌体T4能侵染大肠杆菌的许多菌株,我们的遗传实验结果,是在噬菌体T4rⅡ 区域的顺反子B上做出来的。这个著名的系统是由Benzer[6 ,7 ] 开拓的。rⅡ 区域由相邻的两组基因组成,这两组基因也可以叫做两个顺反子,即顺反子A和顺反子B。野生型噬菌体在大肠杆菌B(这里叫它B)和大肠杆菌K12(λ)(这里叫它K)上都能生长,如果一个噬菌体丧失了其中任何一组基因功能,它就不能在K上生长,这样的一个噬菌体,在B上形成一个r噬菌斑。已知这两组基因的许多点突变,其行为就是这样的。在这个区域中,还发现有一些缺失突变。还有一些突变叫做渗漏突变,这种类型的突变体能表现出部分功能,也就是说它们能在K上生长,但在B上形成的噬菌斑不是真正的野生型。我们在本文中报道的是关于顺反子B的B1片段中的突变体P13(现在我们叫它FC0)的研究工作。突变体P13最初是用吖啶黄处理产生的[8 ] 。
以前我们曾主张[9 ] ,吖啶(如吖啶黄)是通过插入或缺失一个或几个碱基起到诱变作用的。支持这种看法最有力的证据就是吖啶诱发产生的突变体很少是渗漏突变体;这些突变体在这个基因功能上几乎完全丢失。自从我们的短讯发表以来,对我们以前的证据又补充了两方面的实验资料:①我们检查了用吖啶黄诱发产生的126个rⅡ 突变体,其中只有6个是渗漏突变体(用碱基类似物诱发产生的突变体,大约有1/2是渗漏突变体);②Streisinger发现[10 ] ,用碱基类似物诱发产生的、噬菌体T4的溶菌酶突变体,通常是渗漏突变体,而用吖啶黄诱发产生的所有溶菌酶突变体,这种功能都完全丢失了。
如果用吖啶诱发的一个突变体,是通过插入一个碱基形成的,那么,通过缺失一个碱基将使它回复成野生型。我们研究了FC0的回复突变体,发现了回复通常不是原来的突变发生回复,而是在遗传图上与它邻近的一个点上发生了第二个突变。也就是说,是通过同一个基因的抑制基因发生突变而产生回复的。在一种情况(也可能两种情况)下,它可能回复成真正的野生型,但至少在另外18种情况下,所产生的野生型,实际上是具有野生表现型的一种双重突变体。另外一些研究者[11 ] ,用rⅡ 突变体也发现了类似的现象,Jinks[12 ] 还对hⅢ 基因的一些抑制基因进行了详细分析。
在图2中的a行,表示出了FC0的18种抑制基因的遗传图。可以看出,这些抑制基因,虽不是都非常靠近FC0,但都是在这个基因的B1片段上,它们分布在顺反子B大约1/10的区域中。它们不全是在不同的位点上,我们发现它们总共分布在8个位点上,其中大部分都坐落在这些位点的两个连锁丛中,或在其附近。
抑制基因总是一种非渗漏r型。即它在B上形成r噬菌斑,而在K上则不能生长。这是该基因完全缺失的表现型,说明这种功能丢失了。如果抑制基因很快就发生回复突变,以致无法对它进行研究,这种情况除外。
我们曾经谈到,每一个抑制基因突变体在K上都不能生长。因此,我们可以用研究FC0回复突变的方法来研究每个抑制基因的回复突变。发现在少数情况下,这些突变体确实是回复成了原来的野生型,但通常都是通过形成双重突变体来回复的。图2中的b~g行,说明诱发产生的突变体是这些抑制基因的抑制基因突变体。所有这些新形成的抑制基因突变体也都是非渗漏的r突变体,进行遗传作图时,除了一个位点在B2片段上,其他都在B1片段上。
我们用两个新的抑制基因突变体再次重复了这个过程,得到了基本相同的结果,示于图2的i行和j行。
除了原来的FC0之外的所有突变体都能自发产生。但是,我们利用吖啶黄诱发产生了一批突变体(如FC7的抑制基因突变体)。我们得到的这些抑制基因突变体与自发产生的突变体,抑制基因的谱系大致相似(见图2中的h行),而且所有的这些突变体都是非渗漏的r型。我们还用少部分突变体进行了试验,发现用吖啶黄处理,可以使回复突变率增加。
图2 顺反子B左端的一个推测图
这个图只是说明大致情况,图中示出了FC系统突变体的位置。在图的上方、用括号括起来的区域中的位点次序,尚不清楚。用斜体字标出的突变体,只是确定了其大体位置。每一条线表示出从一个突变体(在线上用粗线标出)探察出来的那些抑制基因。
我们总共有80种独立的r突变体,如FC0的所有抑制基因突变体、抑制基因的抑制基因突变体,以及抑制基因的两次抑制基因突变体。这些突变都在这个基因的一个局部区域中,而且都是非渗漏的r突变体。
双重突变体(含有一个突变及这个突变的抑制基因突变)在K上能形成在B上所形成的多种类型的噬菌斑。其中有些噬菌斑与野生型噬菌体产生的噬菌斑并无区别;有些则能区分开,但比较困难;另一些噬菌斑则非常相似于r型,因此,与野生型产生的噬菌斑很容易区分。
在少数的几个例子中,我们检察到,通过互补试验完全可以区分表现型,因为这两个突变体各自的表现型都是r型,当把它们放在一起则是野生型或假野生型的,但必须是在遗传物质的相同部分放在一起才行。如果用不同病毒的两个这样的突变体同时侵染K,则不可能出现这种情况。
概要说明
我们对所有这些事实的解释,都是以本文开头提出的理论为根据的。虽然我们尚无直接证据说明顺反子B形成一条多肽链(可能是通过一种RNA中间物),但我们假定是这样的。为了证实这种看法,我们设想一串核苷酸碱基是从顺反子B左端上的起点开始,每3个一组,依次解读的。例如,我们推断FC0突变体是通过在野生型顺序中,插入一个多余的碱基造成的。那么,在FC0右边所有三联体的解读沿着一个碱基发生移格,因此产生错读。这样,所假设的由顺反子B产生的这种蛋白质的氨基酸顺序,从这个点开始全部改变。这就说明了为什么这个基因的功能丧失了。为了使这种解释更简便,我们提出FC0的一个抑制基因突变(例如FC1)是由于缺失一个碱基造成的。因此,当噬菌体中只有一个FC1突变时,那么从FC1向右的所有三联体都将错误解读,导致这种功能的丧失。然而,当这两种突变出现在同一段DNA中时,如在假野生型的双重突变体FC(0+1)中那样,虽然在FC0和FC1之间三联体的解读将发生改变,但余下的基因将回复原来的正确解读。这样就可以解释,为什么这类双重突变体大都没有真正的野生型表型,而常常呈假野生型。因为根据我们的理论可以看出,它们与野生型的氨基酸顺序有很小的一部分是不同的。
为了方便起见,我们把最初的FC0突变体用+表示(这种选择纯粹是为了方便),我们认为它是由于插入了一个碱基形成的突变体。FC0的抑制基因突变体则用-表示。这些抑制基因的抑制基因突变体同样也用+表示,最后这类突变体的抑制基因突变体,也用-表示(见图2)。
双重突变体
现在我们不禁要问,从我们的80种突变体中,把任何两种突变放在同一个基因中,所形成的双重突变体具有什么特性呢?显然在某些情况下,我们已经知道了答案,因为在分离这些突变体的过程中,已经得到了具有一个+和一个-的一些双重突变体。但是通过鉴定知道,用这种方法没有得到由一个+与另一个+组成的双重突变体,由一个+与一个-组成的许多双重突变体,至今尚未进行鉴定。
现在,我们的理论明确的预言:由+和+(或-和-)组成的所有双重突变体都是r表现型,而且在K上不能生长。表1列出了用这种方法得到的14个双重突变体,进一步证实了这种预见。
乍一看,具有+和-类型的所有双重突变体都应该是野生型或假野生型的,但实际情况还要复杂一些,必须考虑得更周密些,原因就在于下述事实,如果密码是由三联体组成,那么任何一条长的碱基顺序都能以一种方式正确解读,但是,也可以以两种不同的方式错读(在错误点上起读),错读的方式取决于解读骨架是向右转移一个位置,还是向左转移一个位置。
表1 具有r表现型的双重突变体
如果我们以一个位置为准,把解读骨架向一个方向转移用→标记,向相反的方向转移用←标记,那么我们可以确定,按照惯例,我们的+总是在箭头记号的头部,而-总是在尾部,并示于图3。
图3 图示我们对箭头记号的规定是不变的
字母ABC各表示核酸中一个不同的碱基,为了方便起见,用ABC碱基重复顺序来表示(这样编码出来的一条多肽链,各个氨基酸都是相同的)。假定密码是以三联体形式编码的。虚线表示假想的解读骨架,它表示碱基顺序从左边开始,每3个一组,进行解读。
现在的问题是我们的抑制基因突变为什么不会扩大到这个基因全部区域中?最简单的推测是解读骨架的这种转移所产生的某些三联体密码子是不能解读的,例如,可能是无意义密码子,或链终止密码子,或者由于蛋白质结构的复杂性,以某些别的方式不能解读。这意味着,所说的FC0抑制基因必须在一个一定的区域内,所以,由FC0和它的抑制基因之间解读骨架发生转移所产生的三联体密码子都是能解读的。但是,由于任何顺序都存在两种可能的错误,因此,我们可以期望,由一个→转移所产生的不能解读的三联体密码子,在一个←转移所产生的突变中,也可能在遗传图的不同位置上出现。
检查这些抑制基因的突变谱系(在每种情况下分别用箭头←或→表示)看出,在我们的区域中(而不是在它的外面),当→转移在任何地方都是可以解读时,而←转移(从FC0附近的位点开始),只在一个比较小的范围内是可以解读的,这种情况示于图4。在我们的区域左部,FC0和FC1之间,或FC9和FC1之间,如果发生一个←转移,那么必定会有一个或一个以上的三联体密码子是不能解读的,在FC21基因丛右边的区域也有类似的情况。
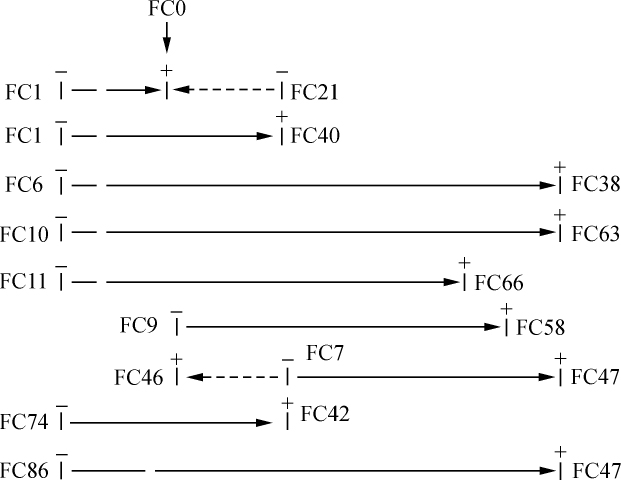
图4 这是图2的一种简化表示形式
每一条线相当于从一个突变体(在其名字底下划一横线标出)得到的抑制基因突变体。箭头记号表示到目前为止已经发现的抑制基因突变的范围。最末端的突变体,在图上标出了它们的名字。
向右的箭头记号用实线表示,向左的箭头记号用虚线表示。
因此,我们预料,含有一个+和-的双重突变体,如果含有一个→转移,那么它将是一个野生型或假野生型;但如果这样的一个双重突变体,含有一个“←”转移,箭头记号穿过一个或两个禁忌位置,由于这样会产生一个不能解读的三联体密码,因此,它将是一个r表现型。
我们用28个例子检验了这种预见,结果示于表2。我们预料,这28个例子中,将有19个是野生型或假野生型,有9个是r表现型。在所有情况下,我们的预料都是正确的。我们认为这有力地证实了我们的理论。在获得这些特异的实验结果之前,首先创立理论,这是很重要的。
表2 含有“+和-”类型的双重突变体
①从FC58(或FC34)产生的双重突变体在K上形成清晰的噬菌斑。
W表现型为野生型或假野生型;
W这种野生型或假野生型双重突变体是用来分离抑制基因突变体的;
r r表现型。
对这个理论的确切陈述
我们曾经说过,到目前为止的证据好像都支持三联体密码学说,但这样解释太简单了。如果用5个碱基一组进行编码,确实也可以获得同样的结果。况且,我们的记号+和-也不一定只表示一个碱基的插入或缺失。
不难看出,我们符号的意义如下所示更为确切:
+表示+m,系数n
-表示-m,系数n
这里n(正整数)是编码比例,即编码一个氨基酸的碱基数目,m是碱基的任何一个整数,可以是正的,也可以是负的。
还可以看出,我们选择的解读方向是任意的,而且在任何一个方向上,也就是说正如通常所画的,不论起点是在基因的右边,还是在左边,遗传物质的解读都将得到大致相同的结果。
三重突变体和编码比例
上面给出的抽象叙述,对于了解密码的普遍性来说是很必要的,但幸运的是,我们有可靠证据说明编码比例实际上是3或3的倍数。
这些证据是在创建具有+、+、+或-、-、-类型的三重突变体时获得的。必须注意的是不能让箭头记号穿过←转移不能解读的区域,这可以通过选择适当的突变体来避免。
到目前为止,我们已检查了6个例子,全列于表3,在所有情况下,这些三重突变体都是野生型或假野生型的。
表3 具有野生型或假野生型的三重突变体

通过了解其中一个三重突变体,例如FC(0+40+38),就可以了解这一结果的显著特性。这3个突变本身都具有相似的类型(+)。我们之所以能这样说,不单单是从分离它们的方法来看的,而且还因为当其中的每一个突变体与突变体FC9(-)发生组合时,所形成的双重突变体都产生野生型或假野生型的表现型。但是当其中任何两种突变结合在一起时(不管是同种突变还是两种不同的突变),所形成的双重突变体都具有r表现型,而且在K上不能生长,也就是说这个基因功能丧失了。即使3个突变都出现在同一个基因中的三重突变体,也只能部分地回复功能,并形成一个可以在K上生长的假野生型噬菌体。
如果编码比例是3,或是3的倍数,我们预期的情况就应该是这样的。
我们之所以能发现这种编码比例,这依赖于下述事实:在我们的呈野生型的混合突变体中,至少其中有一个,在其一条多肽链上,最低限度插入或缺失了一个氨基酸,但对这种基因产物的功能并没有太大影响。
这种情况是很幸运的。我们之所以能使之发生这些变化,并对这样大的一个区域进行研究,可能是因为这部分蛋白质对于其功能的表达不是必须的。Champe和Benzer[13 ] 在他们研究rⅡ 区域互补作用的工作中,曾经提到过这种情况。用一种特异的试验(先在K上进行复合侵染,然后涂在B上)就可以分别测定出顺反子A和顺反子B的功能。有一种特异的缺失,称为1589(见图5),横跨在顺反子A的右端和顺反子B的左端。虽然1589除去了A的功能,但使B的功能还有相当程度的表达。顺反子B由1589所缺失掉的区域,就是产生我们所有FC突变体的区域。
图5 1589缺失结果的梗概
上面两条线分别表示没有1589缺失、但在顺反子A中有一个突变或一个缺失,并不妨碍顺反子B的功能表达。第三条线表示,虽有1589缺失,顺反子B的功能还能表达。其他情况在文章中进行讨论,其中某些情况下,顺反子A中的一种改变,妨碍了顺反子B的功能表达(当1589缺失也存在的情况下)。按照引证文献的习惯,这些情况分别以ⓐⓑ等表示。对于ⓐⓑ两种情况,在本文中没有讨论。√表示有功能;×表示没有功能。
把两个基因连接在一起
我们利用1589缺失进一步证实了碱基顺序是从一个固定起点,一组一组地解读的。通常,顺反子A中的一种改变(可能是吖啶诱发的一种缺失突变体,或别的突变体)不妨碍顺反子B的功能表达。反之,顺反子B中的改变也都不抑制顺反子A的功能。这暗示在这两个顺反子之间有一个区域,这个区域把这两个顺反子隔离开,使它们的功能可单独表达。
我们认为1589缺失导致这个隔离区的丢失,因此这两个局部受损的顺反子就被连接在一起了。实验表明,情况确实是这样的,因为顺反子A左端的一个改变,如果和1589缺失同时发生,那么就可以抑制顺反子B的功能表达。这些结果示于图5。突变体P43或X142(这两个突变体经吖啶处理,都能强烈地发生回复)虽然都在顺反子A中,但当两个顺反子连接在一起的情况下,这两个突变体必有一个将抑制B的功能。X142 S1的情况也是这样的(见图5ⓑ),X142 S1是X142的一个抑制基因突变体。它们(X142和X142 S1)的双重突变体,具有+和-的类型,这个双重突变体本身是假野生型的,当与1589缺失同时发生时,顺反子B的功能仍能表达(见图5ⓒ)。我们曾用这种方法检验了Benzer[7 ] 所列出的10个缺失突变,这些缺失全都发生在1589缺失的左边。发现其中有3个(386、168、221)能抑制顺反子B的功能表达(见图5ⓕ),而其他的7个则不妨碍顺反子B的功能表达(见图5ⓔ)。我们猜想,这7个缺失突变中的每一个都已经丢失了一些碱基,这些碱基的数目正好是3的倍数。缺失的长度不是随机的,缺失的碱基数目常常是编码比例的一个整数倍,这是有理论根据的。
如果最终搞清楚1589缺失产生的蛋白质,一部分是顺反子A的蛋白质,另一部分是顺反子B的蛋白质,这两部分蛋白质以某种多肽链连在一起,而且在某种程度上具有完整的顺反子B蛋白质的功能,这将是不足为奇的。
编码比例是3还是6?
还要说明编码比例是3,而不是3的倍数。以前对编码比例相当粗略的估计[10,14 ] (显然是不可靠的)表明,编码比例接近6。根据我们的理论来看,这暗示了在FC0中的变化不是一个碱基,而是两个碱基(或更确切地说,碱基的数目是偶数)。
我们还有另外一些证据说明,上述估计是不可靠的。第一,在我们用吖啶黄诱发产生的126个突变体中(见前文),有4个独立的突变体是在FC9位点上,或在FC9位点附近。通过选择一种适当的配对方式,我们了解到,其中有两个是+的,两个是-的。第二,我们用肼诱发产生的两个突变体[15 ] (X146和X225),是在FC30位点上,或在FC30位点附近。我们了解到这两个突变体都是一样的。
这样看来,除非吖啶和肼都是引起偶数个碱基的缺失或插入才行,这些证据支持编码比例是3。但是,由于对这些诱变剂的作用,还没有详细弄清,因此,我们尚不能肯定编码比例不是6,尽管编码是3的可能性更大。
我们有一些初步的结果说明,另一些由吖啶诱发产生的突变体,常常由于紧挨着的抑制基因突变而发生回复,但还太粗糙,因此不能在这儿报道。在顺反子B另一端B9a片段上,有一个突变体,叫做P83。把P83的一些抑制基因突变体的放大图示于图6,它们出现的范围比FC0的抑制基因突变体出现的范围要短,大约占B顺反子距离的5%。正如所预料的一样,双重突变体WT(2+5)是r表现型。

图6 P83及其抑制基因突变体WT1等的遗传图
这个区域在顺反子B右端附近的B9a片段中。至于这个图与其周围别的图形关系如何,到目前为止尚不清楚。
密码是简并的吗?
如果密码是三联体密码,那么共有64种(4×4×4)可能的三联体。如若这64种三联体中,只有20种表示20氨基酸,其余的44种都是无意义的,从我们的结果来看,这似乎是不太可能的。如果确实是这样的话,那么出现FC0系统突变体的那个区域(约为顺反子B的1/4),应当比我们观察到的要小得多,因为那样在很近的距离上,偶尔一次移码突变,就会形成一次无意义的解读。这种争论取决于我们前面所假定的、由顺反子B产生的那种蛋白质的大小。这种蛋白质的大小我们尚不清楚,但从顺反子B的长度可以看出,这种蛋白质可能大约含有200个氨基酸。这样看来,密码可能是简并的,即一般说来,由一个以上的三联体编码一个氨基酸。如果确实是这样的,那么关于编码问题的一个最主要的疑难问题也就可以解释了。这个疑难问题就是在不同的微生物中,DNA的碱基组成,可能差异很大,但它们蛋白质的氨基酸组成,差异并不十分明显[16 ] 。但是我们还不能确切地说出,有多少三联体是编码氨基酸的,有多少三联体还有别的功能。
将来的发展
我们的理论引导出一种明确的预见,预料人们将能测定具有“+和-”类型的双重突变体产生的假野生型的氨基酸顺序。从传统的理论来看,由于一个基因只在两个地方发生变化,因此应当只有两个氨基酸发生改变。相反,我们的理论则预言:和两个突变之间的基因区域相对应的多肽链上的一串氨基酸,都将发生改变。噬菌体的溶菌酶就是在这个区域中,它是可以用来检验这种假说的一种很好的蛋白质,目前Dreyer[17 ] 正在对它进行化学研究,Streisinger[10 ] 正在对它进行遗传学研究。
最近在莫斯科举行的生化会议上,我听了Nirenberg的报告,不禁大吃一惊,Nirenberg和Matthaei[18 ] 把多聚尿苷酸(这是一种碱基全为尿嘧啶的RNA)加到一种能合成蛋白质的无细胞系统中,已经合成了多聚苯丙氨酸(这是一种所有残基全为苯丙氨酸的多肽)。这暗示了一段尿嘧啶顺序能为苯丙氨酸编码,我们的工作说明,它可能是一种尿嘧啶的三联体。
通过各种化学或酶学方法合成具有限定或部分限定顺序的多聚核糖核苷酸,这是可能的。如果是这样的话,产生特异的多肽链也是可能的,那么编码问题将由于实验的推动而得到广泛的说明。实际上,现在已有很多实验室,包括我们的实验室在内,已经在做这方面的工作了。假定如我们的结果所揭示的那样,编码比例确实是3,而且如果整个自然界密码都是相同的话,那么遗传密码问题将在一年之内得到解决。
参考文献
[1 ]Wittman, H. G., Symp. 1, Fifth Intern. Cong. Biochem., 1961. for refs. (in press).
[2 ]Tsugita, A., and Fraenkel-Conrat, H., 1960. Proc. Nat. Acad. Sci., USA, 46 , 636; J. mol. Biol., (in press).
[3 ]Brenner S., 1957. Proc. Nat. Acad. Sci., USA, 43 , 687.
[4 ]For refs. see Watson, H. C., and Kendrew, J. C., 1960. Nature, 190 , 670.
[5 ]Crick, F. H. C., Griffith, J. S., and Orgel, L. E., 1957. Proc. Nat. Acad. Sci., USA, 43 , 416.
[6 ]Benzer, S., 1959. Proc. Nat. Acad. Sci., USA, 45 , 1607. for refs. to earlier papers.
[7 ]Benzer, S., 1961. Proc. Nat Acad. Sci., USA, 47 , 403; see his fig. 3.
[8 ]Brenner, S., Benzer. S., and Barnett, L., 1958. Nature, 182 , 983.
[9 ]Brenner, S., Barnett, L., Crick, F. H. C., and Orgel, A., 1961. J. mol. Biol., 3 , 121.
[10 ]Streisinger, G. (personal comnmnication and in press).
[11 ]Feynman. R. P., Benzer, S., Freese, E. (all personal communications).
[12 ]Jinks, J. L., 1961. Heredity, 16 , 153, 241.
[13 ]Champe, S., and Benzer, S. (personal communication and in preparation).
[14 ]Jacob, F., and Wollman, E. L., 1961. Sexuality and the Genetics of Bacteria. Academic Press, New York, Levinthal, C. (personal communication).
[15 ]Orgel. A., and Brenner, S. (in preparation).
[16 ]Sueoka, N., Cold Spring Harb. Symp. Quant. Biol. (in press).
[17 ]Dreyer, W. J., 1961. Symp. 1, Fifth Intern. Cong. Biochem., (in press).
[18 ]Nirenberg, M. W. and Matthaei, J. H., 1961. Proc. Nat. Acad. Sci., USA, 47 , 1588.
大肠杆菌无细胞系统蛋白质合成对天然或合成多聚核糖核苷酸的依赖作用 【86】
马歇尔·尼伦伯格(M. W.Nirenberg) 【87】 海因里斯·马太(J. H. Matthaei)
(美国 国立健康研究所,1961年)
从大肠杆菌得到了一个稳定的无细胞系统,它使14 C-缬氨酸快速地掺入进蛋白质中去,这样来合成蛋白质是需要能量的。L-氨基酸混合物能促进合成,而RNA酶(RNAase)、嘌呤霉素和氯霉素则显著地抑制合成[1 ] 。本文叙述了这个系统的一个新特性,即:即使有sRNA和核糖体存在,氨基酸的掺入还需要模板RNA。而且还说明,加入模板RNA促进氨基酸掺入,其很多特性与从头开始蛋白质合成的特性是相似的。在这个系统中,天然的RNA和合成的多聚核苷酸一样有活性。这种合成的多聚核苷酸可能为由一种氨基酸组成的“蛋白质”编码。在这些资料中,有一部分已在以前的报道中发表过[2 ,3 ] 。
材料和方法
酶提取液是按以前介绍的方法制备的[1 ] ,并在某些方面有所改进。对数期早期收集的大肠杆菌W3100菌体,经过洗涤,再按以前报道的做法[1 ] ,在5℃,用铝粉(湿细胞重量的两倍)研磨5分钟将细胞破碎,加入和铝粉等重量的缓冲液进行抽提,缓冲液(标准缓冲液)成分如下:
| Tris | pH7.8 | 0.01mol/L; | Mg(Ac)2 | 0.01mol/L; |
| KCl | 0.06mol/L; | 巯基乙醇 | 0.006mol/L。 |
然后经20000×g离心20分钟,去掉铝粉和未破碎的菌体。轻轻取出上清液,加入DNA酶(DNAase)(3微克/毫升,华盛顿生化制品公司出品),使悬液的黏度迅速降低,再经20000×g离心20分钟。吸出上清液,用30000×g离心30分钟,除去抽提液中的细胞碎片。吸出液体层(S-30),在105000×g离心2小时,使核糖体沉降。再吸出上清液(S-100),而靠近沉淀的部分溶液要小心地倒出,弃去。将核糖体重悬于标准缓冲液中,经过洗涤,再于105000×g离心2小时。弃去上清液,将此核糖体悬浮在标准缓冲液中(W-Rib)。将S-30、S-100和W-Rib等各部分分别在60倍体积的标准缓冲液中(5℃)透析过夜。分成小份贮存于-15℃。
有时,新鲜的S-30在35℃保温40分钟。每毫升反应混合物所含成分的微克分子数如下:
| Tris | pH7.8 | 80; | 20种氨基酸每种为 | 0.075; |
| Mg(Ac)2 | 8; | ATP(钾盐) | 2.5; | |
| KCl | 50; | PEP(钾盐) | 2.5; | |
| 巯基乙醇 | 9; | PEP激酶 | 15微克/毫升。 |
反应混合物经过保温后,在5℃、60倍体积的标准缓冲液中透析10小时,透析期间换一次缓冲液。将保温过的S-30部分分成小份贮存在-15℃待用(称之为保温过的S-30)。
用酚提取法制备RNA时,所用的酚必须是刚蒸馏出来的。rRNA是用按上述方法新鲜制得的、洗过的核糖体来制备的。制备rRNA时,进行酚处理前,在核糖体的悬液中加入0.2%SDS溶液(SDS用Crestfield等人的方法[4 ] 重结晶过)。悬液在室温下振荡5分钟。通过SDS处理,可能得到较多的RNA;但省去这一步也能得到很好的RNA制品。用SDS处理后,将悬浮于标准缓冲液中的核糖体,加入等体积的水饱和酚,悬液在室温下用力振荡8~10分钟。1450×g离心15分钟后,吸出水相。按同样方法,将水相再用酚提取两次,每次用1/2体积的水饱和酚。最后水相冷却到5℃,加入NaCl,最终浓度达0.1%。在-20℃边搅拌边加入两倍体积的乙醇,使RNA沉淀。悬液用20000×g离心15分钟,倒掉上清液,RNA沉淀溶于最低浓度的标准缓冲液(不加巯基乙醇),并在一个玻璃Potter-Elvehjem匀浆器中缓缓搅匀(通常所用的缓冲液体积约为原来核糖体悬液体积的1/3)。乳白色的RNA溶液在5℃用100倍体积的标准缓冲液(不加巯基乙醇)透析18小时。中间换透析液一次。透析后,RNA溶液在20000×g离心15分钟,弃去沉淀。此RNA溶液中,蛋白质含量不到1%,可将它分成小份,贮存在-15℃备用。
sRNA是用上述的酚提取法,从105000×g上清液中制备的。sRNA也贮存于-15℃。将RNA样品与0.3mol/L的KOH在35℃一起保温18小时,然后中和,并在标准缓冲液中透析(不加巯基乙醇)来制备碱降解的RNA。将RNA与结晶RNAase(2微克/毫升,华盛顿生化制品公司出品)在35℃保温60分钟,来制备酶降解的RNA样品。RNAase是用上述的4次酚提取来破坏的。最后一次酚提取后,样品在不加巯基乙醇的标准缓冲液中透析。RNA样品与经两次重结晶的胰蛋白酶(20微克/毫升,华盛顿生化制品公司出品)一起在35℃保温60分钟。溶液用酚处理4次,并按同样方法进行透析。
所用的同位素标记氨基酸、它们的来源和各自的比活性如下:
本实验中所用的其他材料及方法,可参阅[1],全部实验都有一个重复。
结果
核糖体RNA(rRNA)的促进作用
前篇文章[1 ] 中已证明,在这个系统中加入DNAase 20分钟后,氨基酸的掺入显著降低。从本研究的目的出发,把预先和DNAase及反应混合物的其他各种成分经过保温处理的30000×g上清液部分(保温过的S-30部分)用于很多实验中。
图1表示,用保温过的S-30部分作实验时,加入纯化过的大肠杆菌sRNA对14 C-L-缬氨酸掺入蛋白质的促进作用。加入大约1毫克sRNA可起到最大的促进作用,在有些实验中,浓度增加5倍,对该系统的促进作用并不再增加。除非另有特指外,在所有的反应混合物中都要加入sRNA。
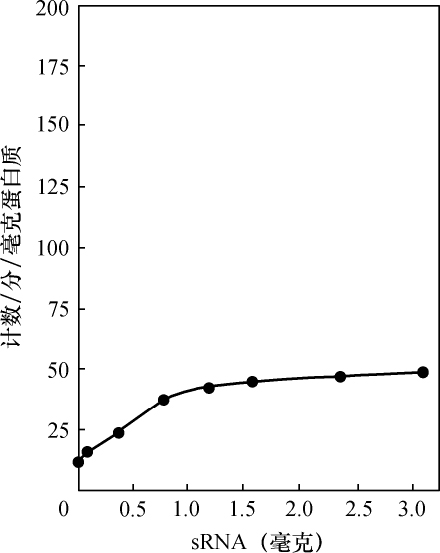
图1 大肠杆菌sRNA对氨基酸掺入蛋白质的促进作用
表1详细记载了反应混合物的组成。样品在35℃保温20分钟,反应混合物含有4.4毫克保温过的S-30蛋白质。
图2证明了即使在反应混合物中sRNA处于最大的促进浓度时,加入大肠杆菌rRNA制品也能显著地促进14 C-缬氨酸掺入蛋白质。当使用低浓度的rRNA时,rRNA浓度和14 C-缬氨酸掺入蛋白质之间是一种直线关系。将sRNA浓度提高3倍以上,也不能代替加入rRNA的效果。

图2 在有sRNA存在时,大肠杆菌rRNA对氨基酸掺入蛋白质的促进作用
反应混合物的详细组成见表1。样品在35℃保温20分钟,反应混合物含有4.4毫克保温过的S-30蛋白质和1.0毫克大肠杆菌sRNA。
图3详细说明了rRNA在促进14 C-缬氨酸掺入蛋白质中的作用。在无rRNA存在的情况下,用保温过的S-30部分作实验时,14 C-缬氨酸掺入蛋白质的数量比用S-30(在贮存于-15℃前,未经保温处理)时低得多,而且30分钟后几乎全部停止。在rRNA浓度低时,掺入蛋白质的氨基酸数量最大值与加入的rRNA数量成正比,这说明rRNA的作用是化学作用,而不是催化作用。实验中,即使当sRNA处于最大促进浓度时,加入rRNA,也可以使掺入蛋白质的氨基酸总量增加3倍以上。在反应过程的任何时间加入rRNA,经过进一步保温后;都可以使掺入蛋白质的14 C-缬氨酸增加。
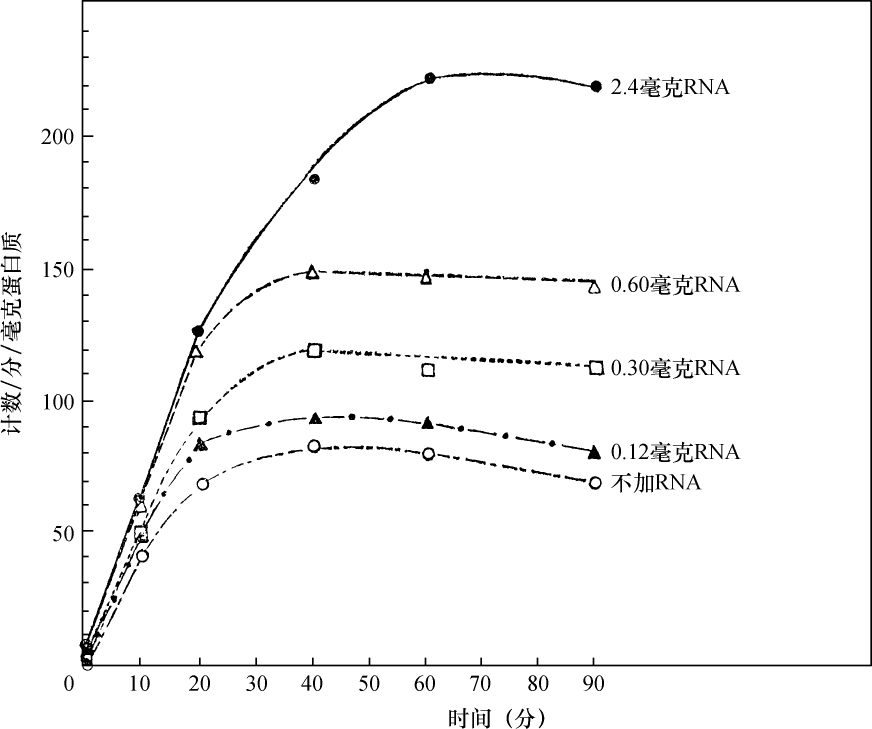
图3 14 C-L-缬氨酸掺入蛋白质对rRNA的依赖
反应混合物的组成和保温条件见表1。反应混合物含有大肠杆菌sRNA 0.98毫克和保温过的S-30蛋白质4.4毫克。
rRNA促进氨基酸掺入的特性
表1列出了加入rRNA促进14 C-L-缬氨酸掺入蛋白质的一些特性。在每毫升反应混合物中加入0.15μmol的氯霉素或0.20μmol嘌呤霉素,氨基酸的掺入就受到强烈抑制。而且,掺入作用还需要ATP和一个产生ATP的系统,每毫升中加入10微克RNAase,掺入作用就受到强烈的抑制。加入等量的DNAase,并不影响由于加rRNA促进的掺入作用。把rRNA制品放在沸水浴中10分钟,不会破坏它的14 C-缬氨酸掺入活性,相反,常常见到还能稍微增加它的活性。但是把这些RNA制品放在沸水浴中更长一些时间,会产生白色沉淀。在冰浴中将悬液冷却,沉淀会立刻溶解。
表1的资料还说明:在有rRNA存在的条件下,加入20种L-氨基酸的混合物,可以进一步促进氨基酸掺入蛋白质,这表明发生的是无细胞的蛋白质合成作用。
表1 14 C-L-缬氨酸掺入蛋白质的特性①
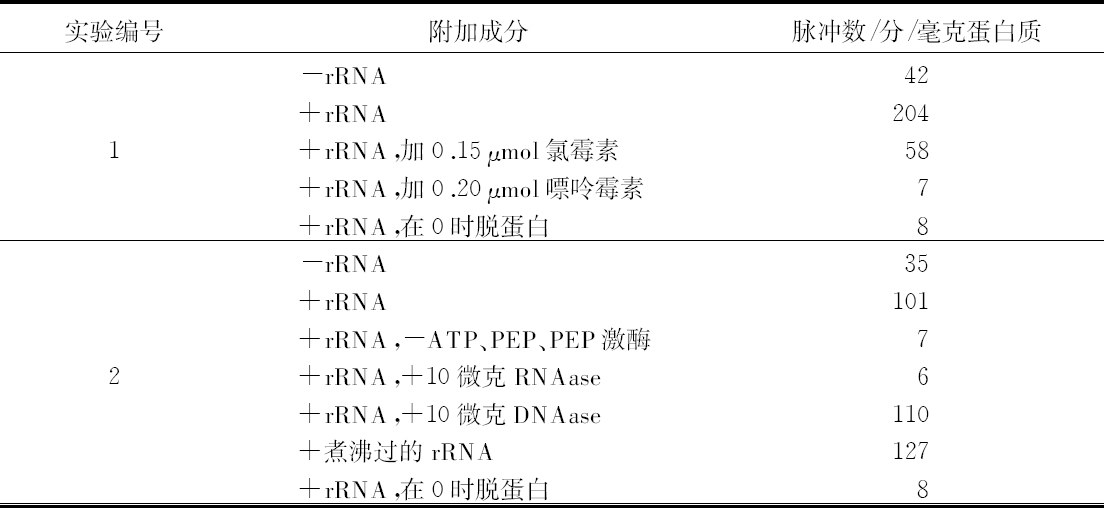

①反应混合物含有下列成分(μmol/mL):
| Tris | pH7.8 | 100; | ATP | 1.0 |
| Mg(Ac)2 | 10; | PEP钾盐 | 5.0 | |
| KCl | 50; | PEP激酶(晶体) | 20微克。 | |
| 巯基乙醇 | 6; |
20种L-氨基酸(缬氨酸除外)每种各为0.05;
GTP、CTP和UTP各为0.03;
14 C-L-缬氨酸 0.015(约为70000脉冲数);
表中的+rRNA表示加3.1毫克大肠杆菌rRNA和1.0毫克大肠杆菌sRNA:“-”号表示不加;
在实验1、2、3中分别加入保温过的S-30蛋白质3.2、3.2和1.4毫克;
在实验3中加入4.4毫克洗过的核糖体蛋白质;
总量为1.0毫升。
样品于35℃保温20分钟,用10%TCA脱蛋白和洗沉淀,用Siekevitz[22 ] 的方法计数。
对反应中依赖于rRNA的产物,分别用羧肽酶和1-氟代-2、4-二硝基苯进行了羧基-和氨基-末端分析。(这些分析由Frank Tietze博士承担。)发现有4%的放射性从羧基末端释放出来,有1%放射性结合在氨基末端上,其余的14 C标记在中间。当用S-30部分(未经DNAase处理)进行反应时,得到了同样的结果。保温后,把从反应混合物中分离到的蛋白质沉淀用HCl彻底水解,通过纸层析证明,掺入到蛋白质中的14 C标记氨基酸是缬氨酸。
本文的很多实验中,用加入DNAase减低黏度来制备酶部分。当使用非DNAase制备的酶提取液时,rRNA也能促进14 C-缬氨酸的掺入作用。
在有洗涤过的核糖体存在的情况下,rRNA能更有效地促进氨基酸掺入蛋白质。表2的资料表明:依赖rRNA的氨基酸掺入作用必须要有核糖体和105000×g上清液。只把105000×g上清液加在rRNA制品中,氨基酸掺入蛋白质的作用不能发生,这表明rRNA制品中不含有完整的核糖体。加热煮沸虽然可以破坏核糖体的活性,却不能破坏rRNA制品的活性,这也证实了上述结论。
表2 在只有核糖体或只有105000×g上清液时,rRNA不能促进14 C-L-缬氨酸掺入蛋白质①

①反应混合物成分和保温条件见表1;
核糖体(W-Rib)为0.86毫克蛋白质;
105000×g上清液(S-100)部分为3.3毫克蛋白质。
表3列出了rRNA对7种不同氨基酸的掺入作用的影响,加入rRNA能增加每种被测氨基酸的掺入作用。
表3 rRNA促进氨基酸掺入的特异性①
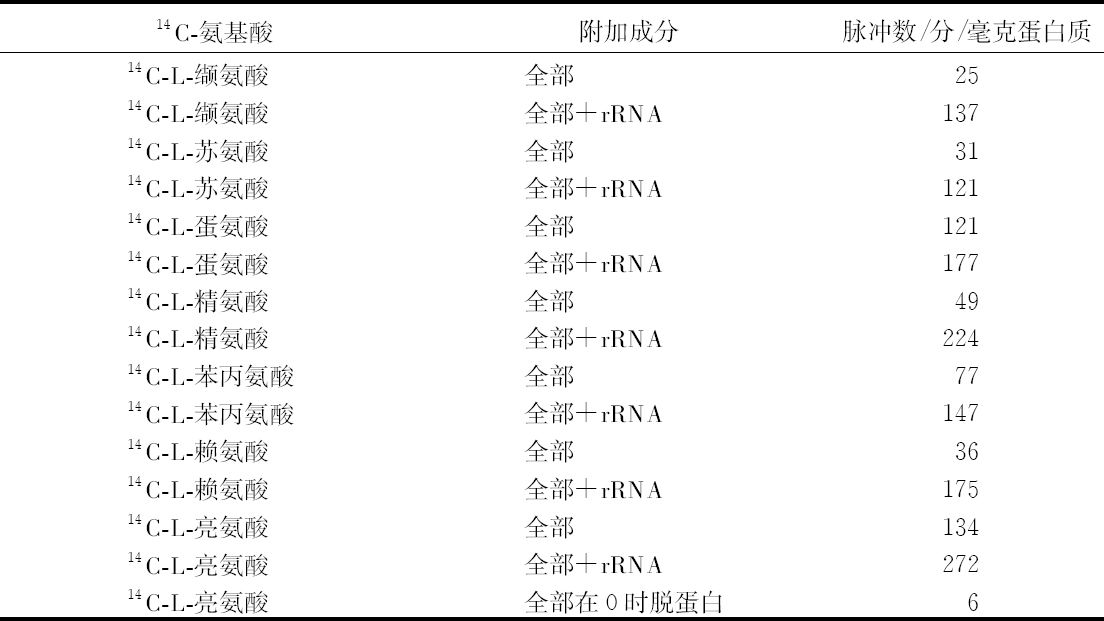
①反应混合物的组成和保温条件见表1。20种L-氨基酸的混合物,包括除了加入反应混合物中的14 C-氨基酸外的所有(另外19种)氨基酸。反应混合物含有4.4毫克保温过的S-30蛋白质。样品在35℃,保温60分钟,加入的rRNA为2.1毫克。
当用其他聚阴离子,如poly(A),高聚鲑精子DNA,或高分子量的葡萄糖羧酸多聚体时,就见不到像rRNA出现的那种效应(表4)。用胰蛋白酶预处理rRNA,并不影响它的生物活性。但用RNAase或碱处理rRNA,就会使其促进活性完全丧失。由此看来,活性因素可能是RNA。
表4 rRNA的对照实验①


①反应混合物的组分和保温条件见表1。在实验1、2和3中,分别加入保温过的“S-30”蛋白质4.4、3.2和4.4毫克。分别加入大肠杆菌sRNA 2.4、0.98和0毫克。
②rRNA是按材料和方法中规定的方法,在酶降解后,用酚抽提脱蛋白制备得到的。
③rRNA的毫克数指的是降解前RNA的浓度。
用Spinco E型超速离心机测定了rRNA制品的沉降特性(见图4A),具有S-30、S-50或S-70核糖体性质的颗粒,在这些制品中未曾发现。第一个峰的 是23、第二个是16、第三个小峰是4。用胰蛋白酶预处理,对 的峰值没有显著的影响(见图4C),若用RNAase处理则会完全破坏这些峰(见图4B),这也附带证实了主要成分是高分子量的RNA这一看法。
是23、第二个是16、第三个小峰是4。用胰蛋白酶预处理,对 的峰值没有显著的影响(见图4C),若用RNAase处理则会完全破坏这些峰(见图4B),这也附带证实了主要成分是高分子量的RNA这一看法。
图4 大肠杆菌rRNA制品的沉降特性
A.未经处理的;B.用RNAase降解了的;C.用胰蛋白酶降解了的。样品的制备和降解见材料和方法。水解前,A中RNA浓度为9.8毫克/毫升;B中为11.5毫克/毫升;C中为10.5毫克/毫升。照片是在一个装有暗线光学照相设备的Spinco E型超速离心机中拍摄的。
最初,rRNA的分步提取是用蔗糖线性梯度的密度梯度离心来做的。其中一个试验的结果示于图5。RNA指导的氨基酸掺入活性不取决于在260纳米的吸光率。相反,活性似乎是集中在No.5(分步收集的编号)部分,它在离管底约1/3处。这些结果进一步说明了活性与sRNA部分无关,因为sRNA主要集中在接近管顶的No.11部分。另外,在sRNA存在的条件下,分析了所有的氨基酸掺入情况,发现即使加入更多的sRNA,也不能促进14 C-L-缬氨酸掺入蛋白质。
图5 rRNA的蔗糖密度梯度离心
从管底到管顶蔗糖线性梯度的浓度从20%到5%[23 ] 。蔗糖溶液(总量4.4毫升)含有:Tris pH7.8, 0.01mol; Mg(Ac)2 0.01mol; KCl 0.06mol。每管顶部铺0.4毫升rRNA(4.6毫克),用Spinco L型离心机,Spinco SW-39水平转子,在3℃,38000×g,离心4.5小时,刺穿离心管底,以每管0.30毫升分步收集[24 ] 。
每部分取样0.025毫升,用水稀释到0.3毫升,测定260纳米的吸光率,取0.25毫升来分析氨基酸的掺入。反应混合物所含的成分见表1。加入0.7毫克大肠杆菌sRNA和2.2毫克保温过的S-30蛋白质。对照加0.25毫升12.5%的蔗糖来代替其他各部分,脉冲计数为79次/分。从每个值中减去这个数。总体积为0.7毫升。样品在35℃,保温20分钟。
从不同菌种得到的RNA的效应
表5表示不同来源的RNA对14 C-缬氨酸掺入蛋白质的促进作用。用Crestfield等人[4 ] 的方法制备的酵母rRNA在促进掺入作用上,要比等量的大肠杆菌rRNA有效得多。用这个方法制得的酵母rRNA分子量约为29000道尔顿,它几乎没有氨基酸受体活性[7 ] 。通过酚提取制备的烟草花叶病毒(TMV)RNA,分子量约为1700000道尔顿,能很有效地促进氨基酸掺入作用。用未经DNAase处理过大肠杆菌的酶提取液作实验时,也观察到加入TMV RNA对氨基酸掺入有显著的促进作用。这个工作的详细情况将在以后发表。
表5 从不同菌种制备的RNA对氨基酸掺入的促进作用①

①反应混合物组分和保温条件见表1。反应样品含有1.9毫克保温过的S-30蛋白质。
合成的多聚核苷酸对氨基酸掺入的促进作用
图6的资料表明:每毫升反应混合物中加入10微克的多聚尿苷酸Poly(U)能显著地促进14 C-L-苯丙氨酸的掺入。苯丙氨酸的掺入几乎完全依赖于加进Poly(U)。加入Poly(U)后,经过很短的延迟期,在大约30分钟以内,掺入作用以直线速度上升。
图6 Poly(U)对U-14 C-L-苯丙氨酸掺入的促进作用
反应混合物组成和保温条件见表1。每毫升反应混合物加0.024μmol的U-14 C-L-苯丙氨酸(约为500000次脉冲计数/分)和2.3毫克保温过的S-30蛋白质。
表6的资料还表明:测定过的其他多聚核苷酸都不能代替Poly(U)。随机混合的腺苷酸和尿苷酸的多聚体(Poly A-U,比例为2/1和4/1)在这个系统中是无活性的,这就更肯定了Poly(U)有绝对的特异性。一种Poly(U)和Poly(A)的溶液(形成三链螺旋)完全没有活性,这说明要有活性必须是单链的。表6的实验还说明了UMP、UDP或UTP不能促进苯丙氨酸掺入。
表6 多聚核苷酸促进苯丙氨酸掺入的特异性①


①反应混合物组分见表1。反应混合物含有2.3毫克保温过的S-30蛋白质。每份反应混合物加0.02微克分子U-14 C-L-苯丙氨酸(约125000脉冲数/分)。样品在35℃,保温60分钟。
表7的资料表明了核糖体、100000×g上清液、ATP以及一个产生ATP的系统,都是依赖Poly(U)的苯丙氨酸的掺入所必需的。嘌呤霉素、氯霉素和RNAase能抑制掺入;但加入DNAase不会抑制掺入。不加其他19种L-氨基酸的混合物,也不会抑制苯丙氨酸的掺入,这说明Poly(U)只促进L-苯丙氨酸的掺入。这个结论被表8的资料所证实。Poly(U)对其他17种放射性氨基酸的掺入几乎无促进作用。我们对每种标记的氨基酸分别进行了测定,这些资料证实了表8的结果,并将在以后的文章中发表。
表7 依赖于Poly(U)的苯丙氨酸掺入的特性①

①反应混合物的组分见表1。除了特别指出的一个实验外,其他的样品中都加10微克的Poly(U)。只测定核糖体和100000×g上清液的两个处理,分别加入0.7毫克W-rib蛋白质和1.3毫克S-100蛋白质,其他的每份反应混合物加入2.3毫克保温过的S-30蛋白质。每个反应混合物加入0.02μM U-14 C-L-苯丙氨酸(比活性=10.3mCi/mmol,约125000脉冲数/分)。样品在35℃,保温60分钟。
表8 Poly(U)促进氨基酸掺入的特异性①

①反应混合物的组分见表1。不加未标记的氨酸混合物。每种标记的氨基酸用量为0.015μM。标记氨基酸的比活性见材料和方法部分。每份反应混合物加2.3毫克预先保温过的S-30部分。所有样品在35℃,保温30分钟。
我们对反应产物做了部分鉴定,结果见表9。反应产物与真正的多聚L-苯丙氨酸的物理特性很相似,与很多其他的多肽和蛋白质不同,在100℃用6mol/L HCl处理时,反应产物和多聚体都不被水解,而在120~130℃,用12mol/L HCl处理48小时,则完全水解。
表9 反应产物和多聚L-苯丙氨酸的特性比较

①不溶,表示产物在100毫升溶剂中(24℃),溶解小于0.002克。提取是在5毫升溶剂中加真正的多聚L-苯丙氨酸0.5毫克和反应混合物的14 C-产物(1800次脉冲/分)。悬液在24℃,用力振荡30分钟,离心。将沉淀涂布在滤膜上并测定放射性。
多聚L-苯丙氨酸在很多溶剂中是不溶的[25 ] ,但能溶于33%的HBr(在冰醋酸中)。反应产物和真正的多聚L-苯丙氨酸表现出一样的溶解性。根据反应产物的特殊溶解性对它进行了纯化。具体做法是:保温后,将反应混合物脱蛋白,沉淀下来的蛋白质按通常的Siekevitz方法洗涤[22 ] 。把含有加入的多聚L-苯丙氨酸载体的干燥蛋白质沉淀,再用33% HBr(在冰醋酸中)来提取,弃去大量的不溶性物质。向溶液中加水,使多聚苯丙氨酸沉淀出来,并用水洗涤数次。用这个程序能回收因加入Poly(U)而掺入进蛋白质中去的14 C-L-苯丙氨酸总量的70%。用12mol HCl把纯化过的反应产物全部水解,然后进行纸电泳,证明反应产物含有14 C-苯丙氨酸。除此而外无其他放射性的点。
讨论
本研究证明,无细胞系统氨基酸的掺入需要模板RNA。在这个系统中,加人sRNA不能代替模板RNA。另外,密度梯度离心实验表明了在rRNA制品中,有活性的部分要比sRNA沉降得快得多。必须注意rRNA与sRNA在性质上是不同的,因为在sRNA中含有假尿嘧啶、甲基化鸟嘌呤等碱基,而rRNA中却没有[5 ] 。
在我们分离的rRNA中,大部分RNA可能没有模板活性,在促进氨基酸掺入作用上,TMV RNA的活性是等量的大肠杆菌rRNA的20倍。此外,对rRNA的初步分级分离表明,总的RNA中只有一部分是有活性的。
必须强调,rRNA不能代替核糖体。这说明,核糖体全然不是由加入的RNA装配起来的。虽然已经知道总的RNA中有一部分是作为蛋白质合成的模板,并称之为信使RNA[12 ~14 ] ,至于rRNA的功能仍然是个谜。也许有一部分RNA,对于从核糖体小亚基来合成有活性的核糖体是不可缺少的[15 ~21 ] 。
rRNA可能是一个亚基的集合体,经过适当的处理后可以解离[6 ~8 ] 。大肠杆菌核糖体经酚提取,可以得到两种类型的RNA分子,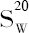 分别为23和16(见图4),分子量分别相当于1000000和560000[9 ,10 ] 。经过煮沸,这些RNA降解成为沉降系数为13.1、8.8和4.4的产物,相应的分子量分别为288000、144000和29000。虽然从这些降解产物的沉降分布来看,各类分子是高度均一的,但这些观察并不排除各亚基通过共价键互相连接的可能性[8 ] 。初步证据表明了在我们的系统中,亚基可能是有活性的,因为把大肠杆菌rRNA煮沸10分钟,再经105000×g离心60分钟,得到的上清液是有活性的。用Spinco E型超速离心机分析煮沸过的rRNA,看到有一个分散的峰,沉降系数为4~8,这个峰与在蔗糖密度梯度离心实验中(用未煮过的RNA制品)常看到的那个比sRNA稍重的、有活性的小峰,是同种物质(见图5)。
分别为23和16(见图4),分子量分别相当于1000000和560000[9 ,10 ] 。经过煮沸,这些RNA降解成为沉降系数为13.1、8.8和4.4的产物,相应的分子量分别为288000、144000和29000。虽然从这些降解产物的沉降分布来看,各类分子是高度均一的,但这些观察并不排除各亚基通过共价键互相连接的可能性[8 ] 。初步证据表明了在我们的系统中,亚基可能是有活性的,因为把大肠杆菌rRNA煮沸10分钟,再经105000×g离心60分钟,得到的上清液是有活性的。用Spinco E型超速离心机分析煮沸过的rRNA,看到有一个分散的峰,沉降系数为4~8,这个峰与在蔗糖密度梯度离心实验中(用未煮过的RNA制品)常看到的那个比sRNA稍重的、有活性的小峰,是同种物质(见图5)。
在我们的系统中,rRNA浓度低时,掺入到蛋白质中去的氨基酸与加入的rRNA量成正比,这说明rRNA是起化学作用而不是催化作用。相反,sRNA是以催化形式起作用[11 ] 。
结果表明Poly(U)含有合成某种蛋白质的遗传信息,这种蛋白质具有多聚L-苯丙氨酸的许多特性。这种合成与加入天然模板RNA所得到的无细胞蛋白质合成是非常相似的,即既需要核糖体,也需要100000×g的上清液,并受嘌呤霉素和氯霉素的抑制。由此看来,苯丙氨酸的密码可能是一个或多个尿苷酸基团。至于编码是单个的、还是三联的或其他类型,至今尚未确定。多聚尿苷酸看来是起合成模板或信使RNA的作用。这种稳定的、无细胞大肠杆菌系统能很好地合成与加入的RNA带有的有意义信息相对应的任何一种蛋白质。
摘要
从大肠杆菌得到一个稳定的、无细胞系统。在这个系统中,掺入到蛋白质中去的氨基酸的数量取决于所加入的热稳定的模板RNA。sRNA不能代替模板RNA。另外,氨基酸掺入既需要核糖体,也需要有105000×g上清液。掺入的氨基酸数量和加入RNA的量之间的关系,说明了模板RNA是起化学作用而不是起催化作用。依赖于模板RNA的氨基酸掺入还需要ATP和一个产生ATP的系统,它受L-氨基酸完全混合物的促进,受嘌呤霉素、氯霉素和RNAase的显著抑制。加入一种合成的多聚核苷酸——多聚尿苷酸,能特异性地使L-苯丙氨酸掺入进蛋白质,形成多聚L-苯丙氨酸。多聚尿苷酸可能起到合成模板,或者叫mRNA的功能。本文还简单讨论了这些发现的含义。
附注
从最近的实验看出,所需要的多聚尿苷酸和掺入的L-苯丙氨酸分子数之比,接近1∶1。关于为苯丙氨酸编码所需要的尿苷酸残基数目以及模板所起的化学数量作用,至今尚未得到直接证据。正如Poly(U)为L-苯丙氨酸的掺入编码一样,Poly(C)特异地支配着L-脯氨酸掺入到能被TCA沉淀的产物中去。有关这些发现的全部资料将在以后发表。
参考文献
[1 ]Matthaei, J. H., and Nirenberg, M. W., 1961. Proc. Nat. Acd. Sci. USA, 47 , 1580.
[2 ]Matthaei, J. H., and Nirenberg, M. W., 1961. Biochem. & Biophys. Res. Comm.,4 , 404.
[3 ]Matthaei, J. H., and Nirenberg, M. W., 1961. Fed. Proc.,20 , 391.
[4 ]Crestfield, A. M., Smith, K. C., and Allen, F. W., 1955. J. Biol. Chem.,216 , 185.
[5 ]Davis, F. F., Carluccl, A. F., and Roubein. I. F., 1959. ibid.,234 , 1525.
[6 ]Hall, B. D., and Doty, P., 1959. J. mol. Biol., 1 , 111.
[7 ]Osawa, S., 1960. Biochim. Biophys. Acta,43 , 110.
[8 ]Aronson, A. I., and McCarthy, B. J., 1961. Biophys, J., 1 , 215.
[9 ]Kurland, C. G., 1960. J. mol. Biol., 2 , 83.
[10 ]Littauer, U. Z., and Eisenberg, H., 1959. Biochim. Biophys. Acta,32 , 320.
[11 ]Hoagland, M. B., and Comly, L. T., 1960. Proc. Nat. Acd. Sci., USA, 46 , 1554.
[12 ]Volkin, E., Astrachan, L., and Countryman, J. L., 1958. Virology, 6 , 545.
[13]Nomura, M., Hall, B. D., and Spiegelman, S., 1960. J. mol. Biol.,2 , 306.
[14 ]Hall, B. D., and Spiegelman, S., 1961. Proc. Nat. Acd. Sci., USA, 47 , 137.
[15 ]Bolton, E. T., Hoyen, B. H., and Ritter, D. B., 1958. in Microsomal Particles and Protein Synthesis, ed. Roberts, R. B., (New York: Pergamon Press), p.18.
[16]Tissieres, A., Watson, J. D., Schlessinger, D., and Hollingworth, B. R., 1959. J. mol. Biol.,1 , 221.
[17]Tissieres, A., Schlessinger, D., and Gros, F., 1960. Proc. Nat. Acd. Sci. USA, 46 , 1450.
[18]McCarthy, B. J., and Aronson, A. I., 1961. Biophys. J.,1 , 227.
[19]Hershey, A. D., 1954. J. Gen. Physiol.,38 , 145.
[20]Siminovitch, L., and Graham, A. F., 1956. Canad. J. Microbiol.,2 , 585.
[21 ]Davern, C. I., and Meselson, M., 1960. J. mol. Biol.,2 , 153.
[22 ]Siekevitz, P., 1952. J. Biol. Chem.,195 , 549.
[23 ]Britten, R. J., and Roberts, R. B., 1960. Science,131 , 32.
[24 ]Martin, R., and Ames, B., 1961. J. Biol. Chem.,236 , 1372.
[25 ]Bamford, C. H., Elliott, A., and Hanby, W. E., 1956. Synthetic Polypeptides (New York: Academic Press), p.322.
蛋白质合成的遗传调节机制 【88】
弗朗索瓦·雅各布(F. Jacob) 【89】 雅克·莫诺(J. Monod)
(法国 巴斯德研究所,1961年)
在细菌中,酶的合成受到一种双重的遗传控制。一种所谓的结构基因决定着蛋白质的分子组成;另外一些有特殊功能的遗传决定子被称为调节基因和操纵基因,它们通过细胞质成分的中间产物或阻遏物,来控制蛋白质的合成速度。某些特定的代谢物可使阻遏物失活(诱导)或激活(抑制)。这种调节系统是通过一种短命的中间产物(或称为信使)的基因直接在合成水平上起作用的,这种信使的出现与蛋白质合成场所——核糖体的出现是偶联的。
引言
据现代最普遍接受的含义,基因这个词指的是一段DNA分子,它自我复制出来的特异性结构能通过尚不清楚的机制,翻译成一条特定结构的多肽链。
结构基因决定着蛋白质结构的多重性、特异性和遗传稳定性,顾名思义这种结构是不受环境条件或作用物控制的。然而,很早就知道,在一个细胞中,各种蛋白质的合成,在特定的外界因素影响下,可受到刺激或抑制,最常见的是:合成不同的蛋白质,依外界条件不同,其合成速率会有很大的变化。而且,从对这类影响的研究中,清楚地了解到它们的作用对于细胞的生存来说是很重要的。
过去曾经提出过,也证明了这些影响一方面可能是基因的互补作用造成的,另一方面也可能是由于决定着蛋白质最终结构的一些化学因子引起的。这个观点至少有一部分是与结构基因的观点相矛盾的,而且至今尚无实验证据。本文中,我们将简要地讨论这些相反的证据。至少暂时还要严格的遵循结构基因假说。我们假设:一个基因中,所含的DNA信息对决定一种蛋白质的结构既是必须的、也是足够的。除了结构基因外,其他作用物在促进或抑制蛋白质合成方面的选择性影响必定是控制结构信息从基因到蛋白质的传递速率。由于蛋白质是在细胞质中合成的,而不是在遗传水平上直接合成,因此这种结构信息的传递必然与基因合成的一种中间产物有关。这种假说中的中间产物我们称之为结构信使。那么,信息传递的速率,即蛋白质合成的速率,可能依赖于基因合成信使的活性,或依赖于信使在合成蛋白质中的活性。这个简图有助于说明与本文有关的两个问题。假定一种作用物能正向或反向特异地改变蛋白质合成的速率,我们不禁要问:
(1)这种作用物是通过控制信使的活性,在细胞质水平上起作用,还是通过控制信使的合成,在遗传水平上起作用呢?
(2)这种影响的特异性是取决于从结构基因传递到蛋白质的信息的性质呢?还是取决于某种特定的控制因素呢?(这种因素并不说明蛋白质、基因或信使的结构。)
从难易程度来看第一个问题还是较容易的。第二个问题则不好直接说明。这个问题概括起来说就是:基因组只带有结构基因呢?还是还带有其他决定子呢?这些决定子根据所给的条件来控制蛋白质合成的速率,但不决定任何一种蛋白质的结构。这两种看法是否能统一起来还得不到证明,我们考虑实验例子时,希望使这两种看法含义明确化,希望证明它们确实是统一的。
蛋白质合成受特异性作用物控制的最好系统是酶适应的例子。适应在这里的含义包括酶的诱导(即酶的形成受底物的选择性的刺激)和酶的抑制(即由一种代谢物引起酶形成的特异性抑制)。只有少数几个诱导和抑制系统在生化和遗传学上已被证明可用于讨论我们这里感兴趣的问题。我们将从这少数系统中推而广之。各溶源系统中,噬菌体的蛋白质合成虽然遵循着不同的规律,但可以用相似的方法进行分析,这一事实又给了我们鼓舞。因此,我们将连接地讨论某些诱导、抑制和各溶源系统。
最好在开头把我们要获得的一些主要的结论讲一下,这些结论是:
(1)从抑制蛋白质合成而不是激活蛋白质合成的意义上看,这些系统的控制机制都是负控制。
(2)除了经典的结构基因外,这些系统中,还包括有另两类在控制机制中完成特异性功能的遗传决定子(调节基因和操纵基因)。
(3)控制机制是在遗传水平上起作用,即通过调节结构基因的活性起作用。
酶的诱导和阻遏系统
酶的诱导现象概述
60多年前就知道,某些微生物的酶只有在它们的特异性底物存在时才能形成(Duclaux,1899;Dienert,1990;Went,1901)。后来Karstrom(1938)把这种效应叫做酶促适应,它已成为很多实验和推测的主题。很长时间以来,酶促适应与生长群体中自发变异的选择不能明确区分。也有人认为,获得一种新的酶促特性的机制,或者是酶促适应,或者是选择作用。直到1946年,才证明在细菌中适应酶系统是受分散的、特异的、稳定的(即遗传的)决定子所控制的(Monod和Audureau,1946)。在细菌中,已经发现和研究了很多种诱导系统。事实上,按一般规律,在这些细菌中,作用于胞外底物的酶是可诱导的。要在较高等生物组织或细胞中研究这种现象是很困难的,但在很多例子中,已经十分清楚地得知这种现象也是存在的。尽管不是一条规律,但也经常发现一种底物不止诱导形成一种酶,而是诱发形成相继卷入到底物代谢中去的几种酶(Stanier,1951)。
从对大肠杆菌乳糖系统的研究中,弄清了诱导效应最基本的特性(Monod和Cohn,1952;Cohn,1957;Monod,1959),并且已从生物化学和生理学的观点将这些基本特性总结于对这个系统的简略讨论中。后面我们将转向对这个系统的遗传学分析。
大肠杆菌的乳糖系统
在大肠杆菌(和某些别的肠道细菌)中,乳糖和其他β-半乳糖苷的代谢过程是通过水解转葡糖基酶β-半乳糖苷酶的作用来完成的。这种酶可从大肠杆菌分离到,随后结晶出来。对这种酶的特异性、被离子的激活作用以及转葡糖基酶对水解酶活力,都进行了详细研究(参看Cohn,1957)。我们只需提一下对本讨论意义重大的那些性质。此酶专一地作用于半乳糖环上未被取代的β-半乳糖苷。当这个配基基团是一个相当大的疏水基团时,酶活性和亲合力受糖苷配基部分性质的影响都是最大的。底物的半乳糖苷键中用硫代替氧,就完全失去水解活性,但是硫代半乳糖苷和同源的氧代化合物,对酶的位点仍能保持着大致相同的亲和力。
按现有的方法分离到的β-半乳糖苷酶会形成各种聚合体(多数是六聚物),聚合体的每个基本单位的分子量是135000。每个单位有一个末端基团(苏氨酸),也有一个酶的位置(用对硫代半乳糖苷的平衡透析来测定的)。至于单体是否也有这样的活性,还是只在体内存在,还不能确定。以邻位-硝基苯-β-D-半乳糖苷作底物,用Na+ (0.01M)作激活物质,在28℃,pH7.0,六聚物分子的周转率为240000克分子/分。在大肠杆菌中,失去β-半乳糖苷酶活性的突变体,在仅有乳糖做碳源时不能生长,这个事实表明了大肠杆菌中只有一种同源的β-半乳糖苷酶,而不能形成别的能代谢乳糖的酶。
然而,具有β-半乳糖苷酶的活性,对于完整的大肠杆菌细胞利用乳糖来说还是不够的。底物进入细胞还需要与β-半乳糖苷酶不同的另一成分(Monod,1956;Rickenberg等,1956;Cohen和Monod,1957;Pardee,1957;Képès,1960)。通过测定同位素标记的硫代半乳糖苷进入完整细胞的速率和/或在细菌细胞中累积的水平,可以测出这种成分的存在和活性。对这种主动的渗透过程的分析看出,它是遵从经典的酶动力学原则的,能测定出其Km 和Vmax 。由于这个系统只对半乳糖苷(β或α),或硫代半乳糖苷是有活性的,因此是高度特异性的。它表现出的亲和性谱系(I/Km )与β-半乳糖苷酶很不相同。因为这个渗透系统,像β-半乳糖苷酶一样是可以诱导的(见下面),所以可以在体内研究它的形成,而且它总是与蛋白质合成偶联的。根据这些标准,这个特异性渗透系统无疑与在诱导过程中形成的一种特异性蛋白质(或几种蛋白质)有关,人们把这种特异性蛋白质叫做半乳糖苷透性酶。不能生成β-半乳糖苷酶的突变体仍能富集半乳糖苷,而失去半乳糖苷透性酶的突变体仍能合成半乳糖苷酶,这些事实说明这种透性酶与β-半乳糖苷酶是不同的,而且功能上是各自独立的。后一类突变体(叫做隐蔽突变体)也不能利用乳糖,因为胞内的半乳糖苷酶要通过特异的渗透系统才能全部到达底物上。
诱导生成的这种蛋白质(或这些蛋白质),据推测可能与半乳糖苷透性酶的活性有关,但直至今日,还不能对它进行离体的鉴定。去年,对能完成
乙酰·辅酶A+硫代半乳糖苷→6-乙酰硫代半乳糖苷+辅酶A
这个反应的一种蛋白质进行了鉴定,并从有半乳糖苷存在条件下培养的大肠杆菌的提取液中纯化了这种蛋白质(Zabin等,1959)。在这个系统中,这种酶的作用还很不清楚,因为,在体内一种游离的共价乙酰化合物的形成根本不涉及渗透过程。另一方面:①不能形成β-半乳糖苷酶、但仍保留有半乳糖苷透性酶的突变体,还保留有半乳糖苷乙酰基转移酶;②不能生成透性酶的突变体也不能形成乙酰基转移酶;③透性酶、乙酰基转移酶都缺陷的突变体回复了生成透性酶的条件,同时就恢复了形成乙酰基转移酶的能力。
这些相关性有力地提出了半乳糖苷乙酰基转移酶是与渗透过程有关的,尽管它在体内的作用尚不清楚,看来它总是与其他几种蛋白质(对于这个系统是特异性的或非特异性的)有关。不管哪一种情况,这里我们感兴趣的不是渗透机制,而是β-半乳糖苷酶、半乳糖苷透性酶和半乳糖苷乙酰基转移酶作用的控制机制。因此,正如我们将见到的,最重要的一点是半乳糖苷乙酰基转移酶和半乳糖苷酶都遵循着同种控制 【90】 。
酶的诱导和蛋白质的合成
在没有半乳糖苷的培养基中培养出来的野生型大肠杆菌细胞,每毫克干重约含有1~10个单位的半乳糖苷酶,也就是说每个细胞平均含有0.5~5个活性分子,或者说每个类核体含有0.15~1.5个分子。在一种含有适当诱导物的培养基中培养出来的细胞,每毫克干重平均有10000个单位的半乳糖苷酶。这就是诱导效应。
这种比活性的显著增加,是与全“新”酶分子的合成有关,还是与原来就存在的蛋白质前体物的活化或转变有关,这是一个很重要的问题,在这方面已经做了大量的实验工作。免疫学和同位素方法相结合,已证实诱导形成的这种酶:(1)它与在非诱导细胞中存在的所有蛋白质都不相同(Cohn和Torriani,1952);(2)它的大部分硫(Monod和Cohn,1953;Hogness等,1955)或碳(Rotman和Spiegelman,1954)不是从原有的蛋白质中得到的。
因此,诱导物引起产生的酶分子,完全是从头合成的,这些酶分子的特异性结构,以及他们的成分来源都是新的。对另外几种诱导系统的研究,完全证实了这个结论,现在可以认为这个结论只是这种效应的定义的一部分。在这里我们将用诱导这一术语来表示由合成酶蛋白的诱导物所引起的激活作用。
诱导的动力学
接受(还是临时的)这种结构基因的假说,我们就可以考虑到诱导物是怎么促进从基因到蛋白质的信息传递速率的。这可以通过促进信使的合成或活化信使来做到。如果信使是一种稳定的结构,在蛋白质合成中起着催化模板的功能,根据诱导物是在遗传水平上起作用或在细胞质水平上起作用,我们预料就会有不同的诱导动力学。
在适当的实验条件下进行测定时,半乳糖苷酶的诱导动力学已弄清楚是十分简单的(Monod等,1952;Herzenberg,1959)。在一种生长培养物中添加适当的诱导物,酶活性增加的速率与该培养物中总蛋白质增加成正比;也就是说,用总的酶活性对培养物群体作图时,得到的是线性关系(见图1)。这条直线的斜率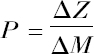 是合成反应的差示速率,按定义它是用来衡量这个效应的。把直线延长可以看出,在加入诱导物后约3分钟(在37℃)开始形成酶(Pardel和Prestidge,1961)。去掉诱导物(或加入一种特异性的抗诱导物,见下面叙述)在同样短的时间内会造成酶合成的停止。合成反应的差示速率,随着诱导物浓度而变化,对不同种的诱导物达到不同的饱和值。因此诱导物的作用方式与在一个酶系统中一种可以解离的活化剂的作用方式是(动力学上)一样的。由于活化剂的加入或除去随之而来的活化和失活是很快的。
是合成反应的差示速率,按定义它是用来衡量这个效应的。把直线延长可以看出,在加入诱导物后约3分钟(在37℃)开始形成酶(Pardel和Prestidge,1961)。去掉诱导物(或加入一种特异性的抗诱导物,见下面叙述)在同样短的时间内会造成酶合成的停止。合成反应的差示速率,随着诱导物浓度而变化,对不同种的诱导物达到不同的饱和值。因此诱导物的作用方式与在一个酶系统中一种可以解离的活化剂的作用方式是(动力学上)一样的。由于活化剂的加入或除去随之而来的活化和失活是很快的。
图1 诱导酶合成的动力学
差示作图表示在大肠杆菌生长培养物中β-半乳糖苷酶的累积是细胞群体增加的函数。因为横坐标和纵坐标都是用相同的单位(蛋白质的微克数)表示的,所以直线的斜率给出的是在诱导物存在时,半乳糖苷酶占合成总蛋白质的部分数(P)(Cohn,1957)。
从这些动力学研究恰好得出了相反的结论:诱导物不能活化一种稳定的、能够在细胞内累积的中间产物的合成(Monod,1956)。除了蜡状芽孢杆菌(Bacillus cereus)的青霉素酶之外,已经做过适当研究的大多数的或全部的其他系统(Halvorson,1960)都已观察到类似的诱导动力学。Pollock的著名工作表明,在培养基中去掉诱导物(青霉素)之后,酶的合成会以递减的速率持续一段较长的时间。这种效应显然与“细胞和青霉素经过短暂接触后即不可逆地保留有微量青霉素”这个事实是有关的(Pollock,1950)。因此,这个系统的独特行为与“从细胞群体中去掉诱导物,受诱导的合成反应就停止”这一规律是不矛盾的。一个细胞可以合成比它带有的诱导物分子多得多的酶分子,从这个意义上,Pollock和Perret(1951)利用这个系统,说明诱导物是起催化作用的。
诱导的特异性
诱导效应的最显著特性之一就是它的极端特异性。一般来说,只有酶的底物,或与正常底物十分相似的物质才能对这种酶有诱导活性。这一点明确地提出了诱导物的分子结构和酶催化中心的结构之间的相关性与诱导机制有着特异关系。为了说明这种相关性,并且进而说明诱导物的作用机制,所提出的假说主要有两类:
(1)在酶合成中,诱导物作为“局部模板”,推测它是作为催化中心的模板;
(2)诱导物通过与以前形成的酶(或称前酶)特异性的结合而起作用,因此以某种方式促进酶分子的进一步合成。
这些经典的假说不需要详细地讨论,因为现在似乎已证实,所讨论的这种相关性,事实上并不是诱导机制所特有的。
表1列出了经测定认为可作为半乳糖苷酶的诱导物和这种酶的底物(或特异性抑制物)的一些化合物。请注意以下几点:(1)每种化合物都具有一个完整未被取代的半乳糖苷基团;(2)很多化合物,它们不是底物(如,硫代半乳糖苷类),却是很好的诱导物(例如,异丙基-硫代半乳糖苷);(3)酶的亲和性和诱导能力之间没有相关性(参看硫代苯基半乳糖苷和蜜二糖)。
随着诱导物不同,所形成的酶是否可能有不同的特异性,这一点也应予以考虑,对此进行了相当周密的测定,得到的都是负结果(Monod和Cohn,1952)。
那么是否测定过的各种半乳糖苷在诱导能力和底物活性或亲和性参数之间根本没有定量的相关性呢?然而,事实是只有半乳糖苷能诱导半乳糖苷酶,这种酶的结合位点是与半乳糖环结构互补的。因此,前面的结果不能完全排除“这种相关性是诱导机制的一个必要因素或结果”这一可能性。
正如我们以后将谈到的,已发现某些半乳糖苷酶结构基因(Z)突变体,不能合成正常的酶,而合成一种新的蛋白质,它的免疫学性质与正常酶相同,但它完全没有任何酶促活性。当用平衡透析进行测定时,这种无活性的蛋白质表现出对半乳糖苷没有亲和性,换句话说,它已失去了特异性的结合位点。在一个携带正常和突变基因的二倍体中,在不同浓度的诱导物存在时,正常的半乳糖苷酶和这种无活性的蛋白质都能形成,而且数量很接近(Perrin, Jacob和Monod,1960)。
从这些发现以及前面的发现可以说明:诱导机制不包含“诱导物的分子和酶的结合位点在结构上存在着特异相关性”的意思,对这种看法不必怀疑。
另一方面,半乳糖苷酶和乙酰基转移酶的诱导是完全相关的,这可以从表1得到说明。从表1不仅可以看出,同种化合物同时作为两种酶的诱导物时,可能有诱导活性,也可能没有诱导活性;而且还可以看出:在不同的诱导物存在时,或者在同种诱导物的不同浓度下,合成的半乳糖苷酶和乙酰基转移酶绝对数量虽然变化很大,但相对数量不变。在这两种极不相同的酶蛋白的诱导过程中,质量和数量上的明显相关性有力地说明了这两种酶的合成受一种共同的控制因素的直接支配,诱导物与这种控制因素是相互作用的。在某种意义上,这种相互作用必定与诱导物的立体特异结合有关,因为诱导在空间上是有特异性的,还因为没有任何诱导活性的某些半乳糖苷,在活性诱导物存在时可以充当诱导作用的竞争性抑制物(Monod,1956;Herzenberg,1959)。这说明了与半乳糖苷酶或乙酰基转移酶都不同的一种酶,或一种其他蛋白质,它可作为诱导物的受体。以后我们将转向验证这种诱导受体的难题。
表1 各种半乳糖苷对半乳糖苷酶和半乳糖苷乙酰基转移酶的诱导作用①

①纵行诱导值指的是野生型大肠杆菌K12培养在以甘油做碳源、并加入了指定克分子浓度的各种半乳糖苷的培养中,所得到的培养物中酶的比活性。给出的值是以与异丙基-硫代半乳糖苷为10-4 M时(这时,实际上每毫克细菌中β-半乳糖苷酶约为7500单位,半乳糖苷乙酰基转移酶约为300单位)所得的值的百分比来表示的。V行指的是对半乳糖苷酶而言,每种化合物的最大底物活性,给出的值是以与苯基半乳糖苷所得活性的百分比来表示的。I/Km 表示对半乳糖苷酶而言,每种化合物的亲和性。给出的值是用与苯基半乳糖苷所得到的值的百分比来表示的。在半乳糖苷乙酰基转移酶的情况中,只给出了V/Km 相对值,因为这种酶的亲和性低,阻碍了常数(I)的单独测定(从Monod和Cohn,1952;Monod等,1952;Buttin,1956;Zabin等,1959;Képès等未发表的结果计算出来的)。
酶的阻遏
正向酶促适应(即诱导)已经知道60多年了,而反向适应(即酶合成的特异性抑制)是1953年在发现色氨酸合成酶的形成受到色氨酸和某些色氨酸类似物的选择性抑制时才被发现的(Monod和Cohen-Bazire,1953)。其后不久,还观察到这种效应的另一些例子(Cohn等,1953;Adelberg和Umbarger,1953;Wijesundera和Woods,1953),在随后的几年中,对几个系统进行了详细研究(Gorini和Maas,1957;Vogel,1957a、b;Yates和Pardee,1957;Magasanik等,1959)。这些研究揭示了,这种后来被Vogel(1957a、b)叫做阻遏的效应,虽然与诱导效应恰好相反对称,但二者是十分类似的。
酶的阻遏与诱导一样,在连续的代谢过程中,一般不是与一种酶而是与一系列的酶有关。诱导能力是对负责降解外来底物的分解代谢酶的衡量标准,而阻遏能力是对与主要代谢产物,诸如氨基酸或核苷酸的合成有关的组成酶的衡量标准 【91】 。阻遏与诱导一样,是有高度特异性的,但是诱导物一般是代谢过程的底物(或底物的类似物),而阻遏代谢物一般是这个代谢过程的产物(或产物的类似物)。
这种效应抑制酶的合成,但不抑制酶的活性(直接地或间接地),这在第一个例子的研究中已经搞清楚了(Monod和Cohen-Bazire,1953),并用同位素参入实验最后证实了(Yates和Pardee,1957)。强调这一点是很重要的,因为酶的阻遏不能与另一种叫做“反馈抑制”或“返回抑制”的不同效应相混淆,这种反馈抑制也同样经常出现,而且可以和阻遏发生在同一个系统中。Novick和Szilard所发现的这种反馈抑制效应(Novick,1955),是通过代谢过程的最终产物,来抑制反应组成序列中早期的一个酶活性(Yates和Pardee,1959;Umbarger,1956)。我们将用阻遏单独命名酶合成的特异性抑制 【92】 。
阻遏的动力学和特异性
通过去阻遏而激活的酶合成的动力学和诱导的动力学是一样的(见图2)。当野生型大肠杆菌被培养在含有精氨酸的培养基中时,只形成微量的鸟氨酸-氨甲酰基转移酶。一旦从生长培养基中去掉精氨酸,酶合成的差示速率会增加约1千倍,而且在重新加入精氨酸之前保持不变;加入精氨酸,酶的合成会立即降回到受阻遏的水平。这种阻遏代谢物这里起的(动力学上)应该是酶系统中一种可以解离的抑制物的作用。
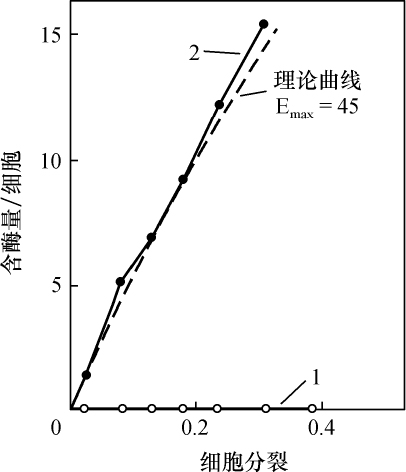
图2 精氨酸对鸟氨酸-氨甲酰基转移酶的阻遏
需要组氨酸和精氨酸的大肠杆菌培养在一个加组氨酸1微克/毫升及精氨酸6微克/毫升(曲线1)或加组氨酸10微克/毫升及精氨酸5微克/毫升(曲线2)的恒化器中。菌体先在过量的精氨酸中培养到对数生长期,洗涤菌体后,再接种于上述培养基。理论曲线是根据细胞分裂4次后,每个细胞所含酶量的恒定数值计算出来的(Gorini和Maas,1958)。
阻遏作用的特异性提出了一些很有意义的问题。一般来说,一个组成系统的阻遏代谢物是这个系统的终产物。例如,除了其他种类的氨基酸外,L-精氨酸阻遏有关精氨酸生物合成系统中的酶。精氨酸本身对这个系统早期的酶,例如特别是鸟氨酸-氨甲酰基转移酶,是没有特异性亲和力的。这时,精氨酸是这种酶蛋白的一种“安慰”阻遏代谢物,就像半乳糖苷是突变了的(无活性的)半乳糖苷酶的“安慰”诱导物一样。但是,通过这个系统本身,精氨酸可能被转化,成为这种酶的中间产物或底物,这也是一种应该考虑的可能性。Gorini和Maas(1957)证明了这个系统晚期的一种酶有缺陷的突变体中,它的鸟氨酸-氨甲酰基转移酶受精氨酸阻遏的程度和野生型是一样的,因而排除了上述的可能性。而且,在不能把中间产物转化成精氨酸的突变体中,这个系统中的鸟氨酸和其他任何一种中间产物,都无阻遏活性。因此,阻遏代谢物作用的特异性并不依赖于酶位点的特异性构型,这是十分清楚的。
同样的结论也适用于组氨酸合成途径中的各种酶,不管是野生型还是缺少其中一种酶的各种突变体,这个合成途径都受组氨酸存在的阻遏。Ames和Garry(1959)的工作证明了在这个系统中,在一定的培养条件下,不同的酶合成的速率在数量上随着恒定的比例而变化,但在缺少其中一种酶的各种突变体和野生型中,不同酶合成的比例是相同的。这里还是与乳糖系统的情况一样,几种完全不同的酶(虽然功能上是有关的)受到一种共同机制的控制,通过这种机制,阻遏代谢物与酶能特异性地互相影响。
总而言之,阻遏和诱导虽然结果相反,但效应似乎很相似,都是控制酶蛋白合成的速率,都是高度特异性的。但是,这两种情况下,这种特异性与所控制的酶的作用(或结合)特异性都没有关系。诱导和阻遏的动力学是一样的。功能上有关的各种酶常常可由同一种底物或代谢物来同时诱导或同时阻遏,而且在数量上能达到同样的程度。
诱导和阻遏明显的相似性说明了这两种效应代表着基本相似的机制的两种不同表现形式(Cohn和Monod,1953;Monod,1955;Vogel,1957a、b;Pardee, Jacob和Monod,1959;Szilard,1960)。这也暗示了,或者在诱导系统中,诱导物是作为一种内在阻遏物的拮抗物,或者在阻遏系统中,阻遏代谢物是作为一种内在诱导物的拮抗物。这并不是一个神秘不可测的问题,而是向我们提出一个十分恰当的问题,即当诱导物和阻遏物都被除去时,在这两种类型的适应系统中,会发生什么情况呢?事实上,这正是我们在下面将试图回答的主要问题。
调节基因
因为诱导作用或阻遏作用的特异性与受控制的酶的结构特异性无关,又因为不同的酶合成的速率似乎是受到一种共同因素的控制,所以推测这种因素不受结构基因本身的控制或阻遏。正如我们现在将看到的,这一结论从对能把诱导或阻遏的系统转变成组成系统的某些突变的研究中,得到了证实。
乳糖系统中的表现型和基因型
假定这种结论是正确的,那么影响控制系统的突变应该没有结构基因的等位基因的行为。为了检验这种预见,必须证明它们的结构基因是一样的。研究得最透彻的例子是大肠杆菌的乳糖系统,现在我们就来谈谈这个系统。在这个系统中,已经得到六类表现型不同的突变体。目前,我们将只考虑其中的3种,并用记号表示和规定如下:
1.半乳糖苷酶突变
Z+  Z- 表示合成有活性的半乳糖苷酶能力的丧失(有诱导或无诱导)。
Z- 表示合成有活性的半乳糖苷酶能力的丧失(有诱导或无诱导)。
2.透性酶突变
Y+ Y- 表示形成半乳糖苷透性酶能力的丧失。大多数(但不是全部)这类突变体同时丧失了合成有活性的乙酰基转移酶的能力。我们的讨论仅限于这种乙酰基转移酶缺陷的亚类。
3.组成型突变
i+ i- 表示在不加诱导物时能大量合成半乳糖苷酶和乙酰基转移酶(Monod,1956;Rickenberg等,1956;Pardee等,1959)。
前两类突变对半乳糖苷酶或乙酰基转移酶是特异性的:半乳糖苷酶缺陷突变体形成的乙酰基转移酶的量是正常的;反过来,乙酰基转移酶缺陷突变体形成的半乳糖苷酶的量也是正常的。相反,已经观察过的上百次组成型突变,都对半乳糖苷酶和透性酶(乙酰基转移酶)有影响 【93】 。这些表现型有8种可能的组合,而且在大肠杆菌ML和K12中都已观察到了。
在大肠杆菌K12中,通过重组对这3种类型的每种突变体多次反复出现的相应位点,已经进行了遗传作图。图3也标出了以后将讨论到的某些其他突变(0突变)的位点。
图3 大肠杆菌K12乳糖区域的遗传图
上方的那条线表示Lac区域和其他已知记号的相对位置。中间的那条线表示带有y、z、o和i四个位点的Lac区域的放大图。下面的那条线表示z和o位点的放大图,线下给出的重组频率是用Hfr ad+ Ss ×F-
ad+ Ss ×F-  ad- Sr 双因子杂交,从Lac+ ad+ Sr 重组体/ad+ Sr 重组体的比例计算出来的。估计z基因的总长度为0.7作图单位,也就是说,对于约有1000个氨基酸的β-半乳糖苷酶单体来说,它约有3500个核苷酸对。
ad- Sr 双因子杂交,从Lac+ ad+ Sr 重组体/ad+ Sr 重组体的比例计算出来的。估计z基因的总长度为0.7作图单位,也就是说,对于约有1000个氨基酸的β-半乳糖苷酶单体来说,它约有3500个核苷酸对。
正如所见到的那样,所有这些位点都局限于染色体的很小的片断——Lac区域中。所有这些突变都是非常接近的,这就提出了一个问题:它们是属于一个功能单位呢?还是属于几个独立的功能单位呢?要进行这种功能分析,必须研究杂合二倍体中各种遗传结构的生化表达。直到最近,在大肠杆菌中才使用了过渡二倍体;在这种细菌中,最近发现了一种新的基因转移类型(性导),因而开辟了得到稳定的、带有染色体不同小片断的二倍体(或多倍体)菌株的可能性。在这个过程中,细菌染色体的一些小片断参入到性因子F中去了。这种新的复制单位可以通过接合作用进行传递,然后加到受体细菌的正常基因组中去,因而使受体细菌变成了二倍体。把这些单位分离出来,其中有一个携带了完整的Lac区域(Jacob和Adelberg,1959;Jacob, perrin, Sanchez和Monod,1960)。为了命名这些二倍体的遗传结构,染色体的等位基因用正常方式书写,而附着在性因子上的那些等位基因,要在前面写上字母F。
把我们的注意力转向Z和Y突变体类型的行为,首先可以注意到:具有Z+ Y- / 或Z- Y+ /
或Z- Y+ / 结构的二倍体都是野生型的,都能够发酵乳糖,形成的半乳糖苷酶和乙酰基转移酶的量都正常,Z- 和Y- 突变体之间这种完全的互补作用,说明它们属于独立的顺反子。相反,各种Y- 突变体之间没有互补作用,这说明它们都属于同一个顺反子。大多数Z- 突变体之间也没有互补作用。某些具有
结构的二倍体都是野生型的,都能够发酵乳糖,形成的半乳糖苷酶和乙酰基转移酶的量都正常,Z- 和Y- 突变体之间这种完全的互补作用,说明它们属于独立的顺反子。相反,各种Y- 突变体之间没有互补作用,这说明它们都属于同一个顺反子。大多数Z- 突变体之间也没有互补作用。某些具有 结构的二倍体,合成半乳糖苷酶的量减少了,但是,互相不互补的突变体配对与互相互补的突变体是重复的,再次说明,这只涉及一个顺反子,这和人们预料的是一致的,因为半乳糖苷酶单体只有一个单一的氨基末端基团。应该重新提到的是顺反子内的部分互补作用现已观察到好几个例子(Giles,1958),并被(推测)解释为与蛋白质的一种聚合状态有关。
结构的二倍体,合成半乳糖苷酶的量减少了,但是,互相不互补的突变体配对与互相互补的突变体是重复的,再次说明,这只涉及一个顺反子,这和人们预料的是一致的,因为半乳糖苷酶单体只有一个单一的氨基末端基团。应该重新提到的是顺反子内的部分互补作用现已观察到好几个例子(Giles,1958),并被(推测)解释为与蛋白质的一种聚合状态有关。
Z基因突变影响半乳糖苷酶的结构。这可从下面的事实看出,大多数Z- 突变体不能合成有活性的酶,却能合成另一种蛋白质,这种蛋白质能把与特异性抗体结合的真正半乳糖苷酶(野生型的)置换出来(Perrin, Bussard和Monod,1959)。在不同的Z- 突变体(命名为Cz1 ,Cz2 等)合成的蛋白质之中,有些能与所用的血清起完全的交叉反应(即能使100%的特异性抗半乳糖苷酶的抗血清沉淀下来),而另一些只有不完全的反应。因此,不同的Cz蛋白质,不仅与野生型的半乳糖苷酶不同,而且彼此也不同。正如我们已经提到的,Z+ / 组成型二倍体能合成野生型的半乳糖苷酶,同时也能以同样的速率合成修饰过的蛋白质(Perrin等,1960)。这些观察证实了“Z区域(或顺反子)含有β-半乳糖苷酶的结构信息”这种结论。目前,在Y区域还没发现既能抑制又能在某种程度上修饰乙酰基转移酶结构的突变,但是,根据Y突变体的性质来看,认为“Y区域并不代表(至少其中一部分)乙酰基转移酶蛋白质结构基因"的假说,似乎是正确的。
组成型二倍体能合成野生型的半乳糖苷酶,同时也能以同样的速率合成修饰过的蛋白质(Perrin等,1960)。这些观察证实了“Z区域(或顺反子)含有β-半乳糖苷酶的结构信息”这种结论。目前,在Y区域还没发现既能抑制又能在某种程度上修饰乙酰基转移酶结构的突变,但是,根据Y突变体的性质来看,认为“Y区域并不代表(至少其中一部分)乙酰基转移酶蛋白质结构基因"的假说,似乎是正确的。
i+ 基因和它的细胞质产物
现在我们注重来讨论组成型(i- )突变。这些突变最显著的特征是:它们总是同时影响两种不同的酶蛋白,正如我们刚才所见到的,这两种酶蛋白是由不同的结构基因各自决定的。事实上,大多数的i- 突变体要比受诱导的野生型细胞合成更多的半乳糖苷酶和乙酰基转移酶,但是十分引人注目的是:在组成型突变细胞中,与在受诱导的野生型中,半乳糖苷酶与乙酰基转移酶的比例是一样的,这有力地说明i基因控制的机制和那种通过诱导物和酶相互作用的控制机制是一样的。
对i+ Z- /Fi- Z+ 或i- y+ /Fi+ y- 这种结构双杂合体的研究表明(见表2,第四行和第五行):可诱导的i+ 等位基因对组成型是显性的;还表明,相对于y+ 和Z- 来说,可诱导的i+ 等位基因在反位中是有活性的。
表2 调节基因突变体的单倍体和杂合二倍体合成半乳糖苷酶和半乳糖苷乙酰基转移酶的情况①
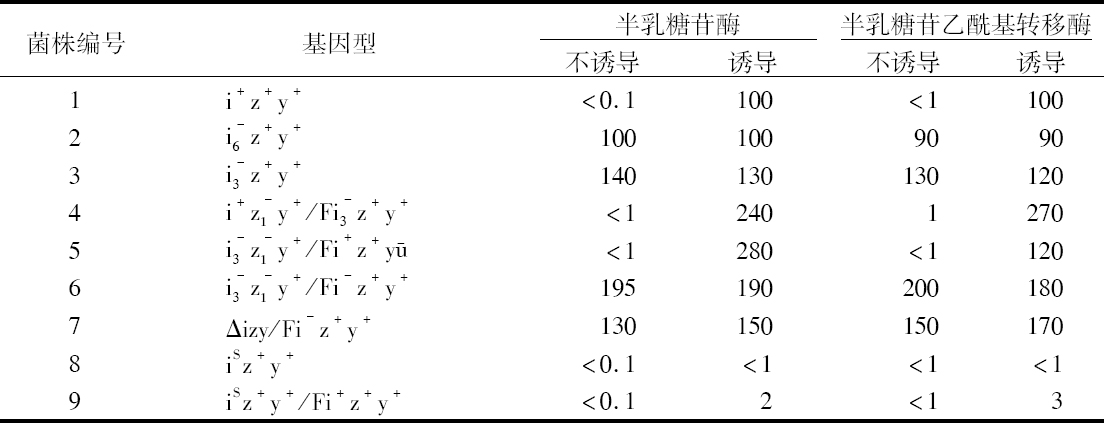
①细菌培养在以甘油做碳源的培养基中,诱导时,加入10-4 M的异丙基-硫代半乳糖苷。表中数值是用这些菌株与经诱导的野生型菌株的酶活百分比来表示的(绝对值,见表1注)。Δizy指的是整个Lac区域都缺失。可以看出在F因子上带有其中一个结构基因(z或y)的野生型等位基因的那些菌株,要比单倍体形成相应的酶多。推测这是由于在每个染色体上存在F-Lac单位的几个拷贝。在i+ /i- 杂合体中,从未诱导细胞所得的值,有时比单倍体对照还高,这是由于在群体中存在大量i- /i- 的同源重组体造成的。
因此,i突变属于一个独立的顺反子,这个顺反子通过一种细胞质成分,控制y和Z的表达。可诱导的等位基因相对于组成型的等位基因是显性的,这意味着前者相当于i基因的活性形式。那些携带izy区域缺失的二倍体菌株的行为与i- 相似(见表2,第七行),这一事实证实了上述看法。但是,必须考虑到对i+ 基因功能有以下两种不同的解释:
(1)i+ 基因决定着一种阻遏物的合成,这种阻遏物在i- 等位基因中是没有的,或者是没有活性的。
(2)i+ 基因决定着一种能破坏诱导物的酶的合成,这种诱导物是通过一条独立的途径产生的。
第一种解释是最直接的,它的最大优点是提出了控制作用的基本机制可能与诱导和阻遏系统的机制是一样的。有几种证据都说明这种解释是正确的。
首先,我们提出这样一种事实,在i- z+ y+ 类型突变体中,组成型的β-半乳糖苷酶的合成不受硫代苯基半乳糖苷的抑制,但已证明(Cohn和Monod,1953)这种硫代苯基半乳糖苷是外源半乳糖苷诱导作用的一种竞争性抑制物(见前面的叙述)。
直接的特异性证据来自对乳糖系统一个特殊突变体的研究,这个突变体(iS )失去了合成半乳糖苷酶和透性酶的能力,但它与所有的z- 和y- 突变体重组时,得到的都是Lac+ 类型,因此它不是缺失。在与z- i- 菌株杂交时,后代都是i- 的;而在与z- i+ 菌株杂交时,后代都是i+ 的,这说明此突变与i区域是紧密连锁的。最后弄清楚了,在组成型iS /i+ 二倍体中,iS 是显性的:这种二倍体既不合成半乳糖苷酶也不合成乙酰基转移酶(见表2,第八行和第九行)。
这种特殊的性质看来是极难说明的,除非明确采用十分特异的假说,即假设突变体iS 是i的一个等位基因,在这个位置上阻遏物的结构再也不能被诱导物所拮抗。如果这种假设是正确的,那么预料通过回复到野生型(iS →i+ )和使i基因失活(可能更经常),也就是说获得组成型条件(iS →i- ),都可以使iS 突变体回复代谢乳糖的能力。事实上,在突变体iS 的群体中,Lac+ 回复突变体的50%确实是i- (隐性的)类型的组成型(另一些回复突变体也是组成型的,但属于以后将谈到的OC 类型)。显然,按照i基因控制着一种破坏诱导物的酶的合成这一假说,是不可能了解这种特殊突变体的性质的(Willson, Perrin和Monod,1961)。
如果接受i+ 基因控制一种胞内阻遏物的合成这一结论,现在我们可以考虑:这种底物在细胞质中的出现问题,以及它的化学性质问题。
把i+ 和z+ 基因引入只有失活的(z- 和i- )等位基因的细胞的细胞质中,研究i+ 和z+ 基因表达的动力学和条件,业已获得了有关上述问题的重要说明。Lac片断从雄性细胞到雌性细胞的有性转移,为这种研究提供了一个适合的实验系统。应该想到,大肠杆菌的接合主要包括雄性染色体(或染色体片段)转移到雌性细胞中去这样一个过程。这种转移是有方向的,总是从这个染色体的一端开始,而且这是一种渐进的过程,在一对指定的交配细胞中,接合开始后,每个染色体片断都在一个十分精确的时间内进入受体细胞(Wollman和Jacob,1959)。这种接合似乎不涉及任何细胞质的混合,因此,事实上,合子继承了雌细胞的全部细胞质,而从雄性细胞接受的只是染色体或染色体片断。为了用合子来研究半乳糖苷酶的合成,因此必须提供使未交配的亲本就不能形成这种酶的条件。这种情况就是:在链霉素(Sm)存在的情况下,诱导型的、能形成半乳糖苷酶、但对链霉素敏感的雄性细胞(♂z+ i+ SmS )和组成型的、不产半乳糖苷酶但抗链霉素的雌细胞(♀z- i- Smr )之间发生的交配。因为:(1)对Sm敏感的雄性细胞在有Sm存在的条件下不能合成这种酶;(2)雌性细胞在遗传上是无产酶能力的;(3)接受了z+ 基因的大多数合子,并不变成对Sm敏感(因为直至很晚期,只有一小部分SmS 基因转移到合子中)。在无诱导物条件下,做的这样一个实验的结果示于图4。可见随着z基因的真正进入,半乳糖苷酶的合成几乎立刻就开始了。后面我们将更精确地分析z基因的表达。这里要强调的重点是:在开始的一段时间,合子的行为和一个组成型细胞相似,在没有诱导物的情况下,也能合成酶。然而,大约60分钟以后,半乳糖苷酶合成的速率降到零。假如在这时加入诱导物,能使酶的合成重新达到最高速率。换句话说,我们亲眼看到合子细胞从原来的i- 表现型回复成了i+ 表现型。这个实验清楚地说明:可诱导的状态与i+ 基因控制合成的一种细胞质物质的存在(并达到足够水平)是有关的。[还要指出:用一个携带Lac区域缺失的雌性菌株,代替这种i- z- 的等位基因,也可以得到同样的结果(Pardee等,1959)]。

图4 可诱导的产半乳糖苷酶的雄性细胞和组成型的、不产半乳糖苷酶的雌性细胞之间发生接合作用,形成的部分合子合成半乳糖苷酶的情况
雄性(Hfri+ z+ T6S SS )和雌性(F- i- z- T6r Sr )细菌分别培养在含有甘油作碳源的合成培养基中,然后在不含诱导物的同种培养基中混合(时间为0)。在这种杂交中,接受了雄性细胞的Lac区域的第一批合子,在第20分钟形成。根据整个群体酶活性的数量,可以决定酶合成的速率,在箭头表示出的时间加入链霉素和噬菌体T6,用以防止进一步形成重组体,并防止对雄性亲本的诱导。可以看到,在没有诱导物情况下,在第一批z+ i+ 片断进入后的60~80分钟,酶的合成停止,但是加入诱导物就能重新恢复(Pardee等,1959)。
假如在z+ 基因进入前几分钟,将5-甲基色氨酸加入到交配过的细胞中去,就不能形成半乳糖苷酶,因为这种化合物通过反馈抑制作用,抑制色氨酸的合成,因此阻断了蛋白质的合成。假定阻遏物是一种蛋白质,或假定它是由一种特异的酶形成的,并受到i+ 基因的控制,那么它的积累在这种情况下也应该受到阻断。反过来,假定这种阻遏物不是一种蛋白质,它的合成不需要i+ 基因所控制的一种特异性酶的预先合成,那么在5-甲基色氨酸存在时,它也可以积累,已知(Gros,未发表的结果)5-甲基色氨酸不能抑制能量的转移或核苷酸的合成。
Pardee和Prestidge(1959)的结果表明:在这些条件下,阻遏物不能积累,因为在加入5-甲基色氨酸后60分钟,再加入色氨酸会立即和完全重新恢复酶的合成,但只有在诱导物存在时才行;换句话说,当蛋白质的合成受到阻断时,合子细胞的细胞质已经从组成型转变成了可诱导状态。用氯霉素作为阻断蛋白质合成的作用物,也能得到了同样的结果,用其他的基因转移系统也重复了这个结果(Luria等,未发表的结果)。
从这个实验得出结论,阻遏物不是蛋白质。这又一次反驳了i+ 基因控制着一种能破坏诱导物的酶的假说。这里我们要强调一下:这个结论并不是说阻遏物的合成与酶无关,而只是说有关的酶不受i+ 基因控制。对了解阻遏物本身的化学性质而言,这些实验尚不充分,因为它们仅能排除蛋白质的候选资格。然而,从这些实验可以推测,阻遏物可能是i+ 基因的初步产物,再进一步推测,这样一种初步产物可能是一种多核苷酸。
结束这一部分之前,还必须指出:在几个诱导系统中都发现了组成型突变体;事实上,不管在那里都是由于采用了适当的选择技术才寻找到它们[大肠杆菌的淀粉麦芽糖酶(Cohen-Bazire和Jolit,1953);蜡状芽孢杆菌的青霉素酶(Kogut, Pollock和Tridgell,1956);大肠杆菌的葡糖苷酸酶(F.Stoeber,未发表的结果);半乳糖激酶和半乳糖转移酶(Buttin,未发表的结果)]。任何一个诱导系统都应该可能产生组成型突变体,这有力地表明,某种功能的丢失就会发生(或至少经常发生)这种突变。在大肠杆菌半乳糖系统中,已经发现这种组成型突变是多效的,能影响连续的3种不同的酶(半乳糖激酶、半乳糖转移酶、UDP-半乳糖差向异构酶),而且这种突变体是发生在与相应的结构基因不同的一个位点上(Buttin,未发表的结果)。
从所评论的这些观察得出的主要结论可以总结为:规定了一类新的基因,我们将把它叫做调节基因(Jacob和Monod,1959)。调节基因不为它所控制的蛋白质提供结构信息。调节基因的特异性产物是一种细胞质物质,这种细胞质物质抑制从一种(或多种)结构基因到蛋白质的信息传递。与经典的结构基因不同,一个调节基因可以控制几种不同的蛋白质的合成,一个基因一种蛋白质的规律对调节基因并不适用。
诱导和阻遏之间的极度相似性,说明这两种效应代表同种机制的不同表现形式。如果这是真实的,而且假定上述的结论也是有确实证据的,预期将发现阻遏系统的遗传控制也与调节基因有关。
阻遏系统中的调节基因
从几种阻遏系统的组成或去阻遏突变体得到的证据,应验了上述预见。为了分离这种突变体,可把某些正常阻遏代谢物的类似物用作特异性的选择因素,因为除了作为阻遏代谢物之外,它们不能代替代谢物。例如,在蛋白质合成中,5-甲基色氨酸不能代替色氨酸(Munier,未发表的结果),但是,它阻遏色氨酸合成系统的酶(Monod和Cohen-Bazire,1953)。正常的野生型大肠杆菌,在有5-甲基色氨酸存在时不能生长。对5-甲基色氨酸完全有抗性的稳定突变体,看来其中大部分原来是色氨酸系统组成型的 【94】 。这些菌株的性质表明:它们是由于一个调节基因RT 的突变引起的(Cohen和Jacob,1959)。在这些突变体中,色氨酸合成酶以及至少还有与这个系统前面步骤有关的另外两种酶,合成速率是相同的,色氨酸的存在对它们并无影响;而在野生型中,所有这些酶都受到色氨酸的强烈阻遏。事实上,在色氨酸存在时突变体形成的酶,要比无色氨酸存在时野生型形成的酶还要多(这和下述情况完全一样:i- z+ 突变体在无诱导物时形成的半乳糖苷酶,要比野生型在饱和浓度的诱导物存在条件下形成的酶还要多)。突变体从培养基中聚集色氨酸的能力未受到损害,它们的色氨酸酶的活性也没增加。因此阻遏代谢物对色氨酸敏感性的丢失,不能归结为是细胞把色氨酸破坏了,或是排泄出去了。只能是反映了控制系统本身的破坏。对RT 突变的几个回复突变进行了遗传作图。发现它们都位于染色体的同一小区域内,与Yanofsky和Lennox(1959)证实的、合成这个系统各种酶的基因丛,相距很远。Yanofsky(1960)的工作又证明:这些基因(包括两个顺反子)中,有一个是色氨酸合成酶的结构基因,据可靠推测,这个基因丛中的另一些基因,决定着这个系统中前面的各种酶的结构。由此看来,RT 基因是控制几种不同蛋白质合成的速率,而不是决定它们的结构。因为它的位置离结构基因很远,因此只有通过一种细胞质中间产物才能起到这种作用。为了证实它是一个调节基因,必须证实组成型( )等位基因就是相当于这个基因(或基因产物)的失活状态,即是隐性的。这种情况下,不能用稳定的杂合子。但是♂ ×♀
)等位基因就是相当于这个基因(或基因产物)的失活状态,即是隐性的。这种情况下,不能用稳定的杂合子。但是♂ ×♀ 杂交得到的一些过渡(有性)杂合体对5-甲基色氨酸是敏感的,这表明可阻遏的等位基因是显性的(Cohen和Jacob,1959)。
杂交得到的一些过渡(有性)杂合体对5-甲基色氨酸是敏感的,这表明可阻遏的等位基因是显性的(Cohen和Jacob,1959)。
在精氨酸合成系统中,大约有7种酶同时受精氨酸阻遏(Vogel,1957a, b;Gorini和Maas,1958)。控制这些酶的特异性基因(即可能是结构基因)分散在染色体的不同位点上。已经得到一些抗刀豆氨酸的突变体,在这种突变体中,其中有几种(也可能是全部)酶同时被去抑制。这些突变发生在与这些结构基因位点距离很远的一个位点上(靠近Smr )。对这种显性的相关性,还未做分析(Gorini,未发表的结果;Maas等,未发表的结果)。
碱性磷酸酶的情况更有趣,因为与这种蛋白质相对应的结构基因已经得到了很好的证实,证据就是在这个位点上的各种突变都会造成所合成的磷酸酶发生改变(Levinthal,1959)。这种酶的合成受正磷酸盐的阻遏(Torriani,1960)。已分离到了一些在正磷酸盐存在时能合成大量酶的组成型突变体。这些突变发生在与结构基因不等位的两个位点上,所有的测定都证明这种组成酶与野生型(可阻遏的)酶是一样的。这两个位点的组成型等位基因都被证明相对于野生型是隐性的。相反,结构基因(P基因)的突变不影响调节机制,因为在正磷酸盐存在时,由P基因突变体形成的改变了的(无活性的)酶,和野生型酶受到相同程度的阻遏(Echols等,1961)。
阻遏物、诱导物和辅阻遏物的相互作用
综合上述观察,可以看出,毫无疑问,阻遏和诱导一样也是受特异性的调节基因控制的,在这两类系统中,调节基因都是通过一种基本相似的机制进行操纵的,换句话说,就是通过控制一种胞内物质的合成来进行操纵的。这种胞内物质能抑制从结构基因到蛋白质的信息转移。
因此,很明显,在阻遏系统中能抑制酶的合成的那些代谢物(如色氨酸、精氨酸、正磷酸盐),本身是没有阻遏活性的,只有通过与在调节基因控制下合成的一种阻遏物互相作用才有阻遏活性。它们的作用最好说成是对受遗传控制的阻遏系统的一种激活作用。为了避免用词上的混淆,我们将把阻遏代谢物称为辅阻遏物,仍旧把调节基因的细胞质产物叫做阻遏物(或主阻遏物)。
阻遏物和辅阻遏物(在阻遏系统中)或阻遏物和诱导物(在诱导系统中)之间相互作用的性质确实是个难题。单纯作为一种形式上的叙述,人们可以把诱导物认为是阻遏物的拮抗物,把辅阻遏物认为是阻遏物的激活物。为了说明这种拮抗作用或激活作用,可以提出各种各样的化学模型。我们不准备讨论这些假说,因为目前还没有证据可以支持或否定任何一种特异的假说。但必须指出,不管哪一种模型,必须说明诱导物或辅阻遏物的结构特异性,要说明这一点,只有假设这种相互作用和一种立体特异受体有关。阻遏物不是一种蛋白质,这一事实引起一个极大的困难,因为与小分子形成立体特异复合物的能力,好像是蛋白质的一种特性。如果一种蛋白质(可能是一种酶)和这种特异性有关,而这种蛋白质的结构假定又是由一个结构基因决定的,那么这个基因的突变将造成失去被诱导(或被阻遏)的能力。这类突变体(具有多效的、隐性的、并且能被其他结构基因突变体互补的这些明确可预见特性)在乳糖系统中尚未见到;但受控制的酶本身(半乳糖苷酶或乙酰基转移酶)起诱导酶作用的可能性已被排除。
可以想象,在合成氨基酸的阻遏系统中,这种作用是由担负主要功能(这些主要功能丧失就会致死)的酶,例如活化酶,同时承担的。但这在大多数诱导系统的情况中似乎是难以想象的。通过这些观察仍不能排除的一种可能性是阻遏物本身合成诱导蛋白质,然后阻遏物再与诱导蛋白质结合,这样,诱导酶的遗传失活将与阻遏物本身结构改变是偶联的,而且一般表现为是调节基因的组成型突变 【95】 。在这里谈到这种可能性,只是用来作为我们已简短分析过的这个进退两难的问题的一种解释,问题的解决还取决于对阻遏物化学结构的鉴定。
在温和噬菌体系统中的调节基因和免疫性
某些基因的潜在功能,或者可以表达出来,或者受到特异性的抑制,最突出的一个例子就是温和噬菌体系统中的免疫现象(Lwoff,1953;Jacob,1954;Jacob和Wollman,1957;Bertani,1958;Jacob,1960)。
这种所谓的温和噬菌体,它的遗传物质可以下述两种状态中的一种在寄主细胞内存在。
1.营养体状态
噬菌体基因组自主地增殖。在这个过程中,全部的噬菌体组分都合成了,最终的结果导致寄主细胞溶菌,并释放出有侵染性的噬菌体颗粒。
2.原噬菌体状态
通过两种遗传成分作为一个单位进行共同复制的方式,使噬菌体的遗传物质附着到细菌染色体的一个特异位置上去。这种寄主细胞就叫做是溶源的。只要噬菌体基因组处于原噬菌体状态,就不产生噬菌体颗粒。为了使溶源细菌产生噬菌体,噬菌体的遗传物质必须经过从原噬菌体到营养体状态的转变。在溶源细菌正常生长期间,这种转变极少发生。但是,含有某几种原噬菌体的溶源菌培养物,经过紫外线、X射线或能改变DNA代谢的各种化合物的处理,可以诱发这种转变(Lwoff, Siminovitch和Kjeldgaard,1950;Lwoff,1953;Jacob,1954)。
通过对因突变而改变了产生噬菌体颗粒所需要的步骤之一的、某些缺陷噬菌体基因组的研究可以看出:至少存在两组不同的病毒功能,它们都与合成特异性蛋白质的能力有关(Jacob, Fuerst和Wollman,1957)。某些早期功能看来是噬菌体基因组的营养增殖所预先需要的(至少在T系偶数列的烈性噬菌体中是这样),现在已经知道,这些早期功能与一系列新酶的合成有关(Flaks和Cohen,1959;Kornberg, Zimmerman, Kornberg和Josse,1959)。一组晚期功能与组成噬菌体外壳的结构蛋白质的合成有关。这些不同的病毒功能的表达,仿佛在某种意义上是与一连串的过程同等的,因为某些早期功能的缺陷突变,也可能导致不能完成噬菌体增殖的几个后期步骤(Jacob等,1957)。
相反,在原噬菌体状态下,病毒的功能是不表达的,而且在溶源细菌中噬菌体外壳的蛋白质组分是测不出来的。另外,如果一种噬菌体颗粒的基因组已经以原噬菌体状态存在于溶源细菌中,那么这种溶源细菌就会对这种噬菌体颗粒表现出显著的特异的免疫特性。用同源噬菌体颗粒侵染溶源细胞时,这些颗粒吸附到细胞上,并把它的遗传物质注入细胞,但细胞仍能存活。注入的这种遗传物质不表现它的病毒功能,它不能起始外壳蛋白质成分的合成,也不能进行营养体的增殖。在细菌的细胞增殖过程中,它仍是不起作用的,并不断被稀释(Bertani,1953;Jacob,1954)。
因此,在溶源细菌中,噬菌体基因功能的抑制不仅适用于原噬菌体,也适用于其他的同源噬菌体基因组。它只取决于原噬菌体的存在(而和原噬菌体刺激造成的、细菌基因的永久性变化无关),因为要使细菌重新变得对噬菌体敏感,必须(也只需)去掉原噬菌体。
有两种解释可考虑用来说明这些免疫性的相互关系:
(1)原噬菌体占据并封阻了寄主染色体上的一个位点,在某种意义上说,这个位点是同源噬菌体营养增殖所特异需要的。
(2)这种原噬菌体产生一种细胞质抑制物,妨碍了起始营养增殖所需要的一些反应(推测是一种特异性蛋白质的合成)的完成。
要在这两种可选择的假说中作出抉择,可以通过研究宿主的二倍体(溶源特性是杂合的)来达到。分离到一个性因子,它攫取了一小段细菌染色体,这个染色体小片段带有控制乳糖发酵的基因(Gal)和原噬菌体λ的附着位点。带有Gal- λ- /FGal+ λ+ 或Gal- λ+ /FGal+ λ- 结构的二倍杂合体,对于λ噬菌体超感染是免疫的,结果表明免疫性对非免疫性是显性的,而且是通过细胞质进行表达的(Jacob, Schaeffer和Wollman,1960)。
对溶源(λ+ )和非溶源(λ- )细胞发生接合的过程中所形成的过渡合子的研究,也得到了同样的结果。在♂λ+ ×♀λ- 的杂交中,由雄性染色体携带的原噬菌体转移进非免疫的受体,从而引起向噬菌体营养状态的转移,使噬菌体在合子中增殖,并发生溶菌,释放出噬菌体颗粒。这种现象被称为合子的诱导(Jacob和Wollman,1956)。但是,在反向杂交♂λ- ×♀λ+ 中,不会发生合子的诱导。这种由雄性染色体携带的非溶源特性转移进免疫受体,不能引起原噬菌体的发育,产生的这种合子对λ噬菌体的超感染是免疫的。
用溶源和非溶源的雄性和雌性细胞进行正反交,所得的相反结果与乳糖系统中用可诱导和不能诱导的细胞进行正反交得到的结果,是完全类似的。在这两种情况中,决定因子显然都是合子细胞质的来源,而且必然得出如下结论:溶源细菌的免疫性是起因于一种细胞质成分,在这种成分存在时,病毒的基因就不能表达(Jacob,1960)。
在乳糖系统中,我们曾经用来解释调节基因产物的两种假说,同样也适用于在溶源细菌中保证得到免疫性的细胞质抑制物。这两种假说是:
(1)这种抑制物是一种特异性的阻遏物,它阻止起始噬菌体营养增殖所需要的一种(或多种)早期蛋白质的合成。
(2)这种抑制物是一种酶,它能破坏由非溶源细胞正常合成的、为噬菌体的营养增殖所特异需要的一种代谢物。
对第二个假说提出争议的有几方面的论据(Jacob和Campbell,1959;Jacob,1960)。首先,对于所给的一个细菌菌株,已知有多种温和噬菌体,其免疫图像各不相同。根据第二种假说,这些噬菌体中的每一种,在营养增殖时都特异地需要由非溶源细胞正常产生的一种代谢物,推测各种代谢物互不相同。第二个争论是由下列事实引起的:像乳糖系统的阻遏物一样,在有氯霉素存在时,(即在没有蛋白质合成时)能合成与免疫性有关的抑制物,在有氯霉素存在的情况下进行♂λ+ ×♀λ- 杂交,不发生合子的诱导,并发现在重组体中原噬菌体能正常传递。
为了说明溶源细菌中的免疫性,我们采用了和在适应酶系统中同样类型的解释。根据这种解释,原噬菌体控制着一种细胞质阻遏物,这种细胞质阻遏物能特异地抑制一种(或几种)对起始噬菌体营养增殖所必需的蛋白质的合成。在这个模型中,不论是通过侵染还是通过接合,将噬菌体的遗传物质引入非溶源的细胞,都将引起特异性阻遏物的合成和噬菌体营养增殖所需要的早期蛋白质合成之间发生一场竞争。寄主细胞的命运如何?是通过溶源化存活下来呢?还是因噬菌体增殖导致溶菌呢?这要看条件是有利于阻遏物的合成,还是有利于那种蛋白质的合成。如果培养条件的改变(如在低温下侵染,或有氯霉素存在)有利于阻遏物的合成,就会导致溶源化,若有利于那种蛋白质的合成,就会导致溶菌。以紫外线诱发现象为例,可通过下列途径来理解:可诱导的溶源细菌暴露于紫外线或X射线,将短暂地破坏调节系统,例如,阻碍阻遏物的进一步合成。假定阻遏物是不稳定的,那么它在细胞内的浓度将降低,并达到很低的水平,足以使早期蛋白质能够合成。于是噬菌体的营养增殖便被不可逆转地起始了。
通过对免疫性的遗传分析,进一步增进了对溶源系统和适应系统之间相似性的认识。从遗传图上看,λ噬菌体的基因组可能包括两部分(见图5):一是中心片断——C区域,它含有一些决定子,这些决定子控制着与溶源化有关的各种功能(Kaiser,1957);这个连锁群的另一部分含有一些控制病毒功能的决定子,即与噬菌体的各种蛋白质有关的结构基因。某些温和噬菌体菌株虽然免疫图像不同,但仍然能够进行遗传重组。在这样的杂交中,特异性免疫图像的差异是受C区域的一个小片段im控制的(Kaiser和Jacob,1957)。换句话说,一个原噬菌体,在它的C区域含有一个小片段im,这个im片段控制着对带有同源im片段的噬菌体基因组有特异活性的阻遏物的合成。
图5 λ温和噬菌体连锁群的图示
图中表示各个标志的直线排列。图中符号指的是各种噬菌斑的大小、类型和寄主范围的标志。符号d指的是各种缺陷突变体。用粗线表示的是C区域,在下方是它的放大图。数字表示相对应的各种C突变。这个C区域可以再分成3个功能单位,C1 、C2 和C3 ;控制免疫性的那个片段定名为im。
在im区域,有两类突变,它们的性质与影响适应酶系统调节基因的各种突变是极相似的。
(1)某些突变( →C1 )引起在单一侵染中溶源化能力的完全丧失。所有的C1 突变都位于im片段的一小段上的一个基因丛中,在互补试验中,它们的行为属于同一个顺反子。
→C1 )引起在单一侵染中溶源化能力的完全丧失。所有的C1 突变都位于im片段的一小段上的一个基因丛中,在互补试验中,它们的行为属于同一个顺反子。
用C1 和 噬菌体进行混合感染,可得到带有C1 和 两种原噬菌体的双溶源无性系。这些双溶源无性系发生分离时,会产生只带有C+ 类型的单溶源细胞,但不会产生只带有C1 类型的单溶源细胞。这些发现表明了野生型等位基因对突变体C1 等位基因是显性的,而且通过细胞质表达出阻止突变体基因组进入原噬菌体状态。因此,C1 突变的性质与适应系统隐性组成突变体的性质是相似的。这些证据说明,C1 位点控制着与免疫性有关的阻遏物的合成,还说明C1 突变相当于这个位点(或它的产物)的失活。
(2)发现有一种突变(ind+ →ind- ),引起原噬菌体丧失可诱导性,即溶源菌经过紫外线、X射线或化学诱导物处理引起噬菌体营养增殖的这种能力丧失了。这种突变是位于C1 片段上。突变体等位基因ind- 对野生型等位基因ind+ 是显性的,因为双溶源性的λind+ /λind- 或结构为:Gal- λind+ /FGal+ λind- 或Gal- λind- /FGal+ λ+ ind+ 的二倍杂合子,都是不可诱导的。另外,λind- 突变体表现出一种独特的性质。如果带有一个野生型原噬菌体的溶源细菌K12(λ+ ),经紫外线照射整个群体便会溶菌,并释放出噬菌体。在照射前或照射后立即用λind- 突变体侵染这种细胞,将完全抑制噬菌体的产生和溶菌作用。
ind- 突变体的性质好像在各个方面,都与以前叙述过的乳糖系统中iS 突变体的性质很相似。对ind- 突变体的独特性质只能用同一种假说来解释,即ind- 突变通过产生更多的阻遏物或产生更有效的阻遏物的方式,在数量上或质量上影响阻遏物的合成。如果这种推测和C1 突变能使一种活性阻遏物的产生能力丢失的假说都是正确的,那么,C1 ind-双重突变体就会丢失在野生型噬菌体侵染溶源菌时,抑制噬菌体增殖的能力。这是已经观察到的事实。很明显,认为C1 位点控制着一种破坏代谢物的酶合成的这种假说,是不能解释ind- 突变体的性质的(Jacob和Campbell,1959)。
总之,对溶源系统的分析揭示了在这些系统中,病毒基因的表达是受一种细胞质阻遏物控制的,而这些阻遏物的合成又受病毒基因组一个特殊的调节基因控制。这种阻遏物能阻遏哪些蛋白质的合成,这一点尚未证实,但这些蛋白质很可能就是起始噬菌休整个营养增殖过程早期的酶。鉴于这些蛋白质对不同类型的诱导条件十分敏感,看来噬菌体的阻遏系统完全可以与酶适应系统相匹敌。
操纵基因和操纵子
操纵基因是阻遏物作用的部位
在前面的章节中,我们已讨论过的证据表明了从结构基因到蛋白质的信息传递是受特异的阻遏物控制的,阻遏物由调节基因合成。我们现在应当考虑下一个问题,即阻遏物作用的部位和模型。
关于这个问题,阻遏物最重要的性质是它作用所特有的多效性和特异性。在大肠杆菌乳糖系统中,i基因的突变不影响任何其他系统,所以,阻遏物是具有高度特异性的;这些突变又同时影响着半乳糖苷酶和乙酰基转移酶,而且在数量上达到相同的程度,所以阻遏物又是多效的。
阻遏物操纵作用的特异性指的是,它通过与该系统的一个具有适当(互补的)分子构型的部分,形成立体特异结合而起作用。而且必须假定,当这一部分与阻遏物结合时,从基因到蛋白质的信息流动就会被中断。这个控制部分,我们将称之为操纵基因(Jacob和Monod,1959)。我们应当引起对下述事实的注意,一旦能够证实一种特异性阻遏物确实存在,那么,上面规定的那种操纵基因成分的存在,随之也就是必然的了。因此,我们的问题不是研究操纵基因是否存在,而是研究在信息传递系统中,它在哪里以及如何介入的问题。
从上面的考虑立即得到一个重要的预示——任何一种有关操纵基因性质的假说,它的特异的互补构型必定受遗传决定;因此,有些突变能改变或取消操纵基因对阻遏物的特异性亲和力,但不会降低它起始信息传递的活性。这些突变也必然影响到操纵基因本身,将引起一种(或多种)蛋白质的组成型合成。这些突变将用来定义一个操纵基因位点,它与调节基因在遗传学上是不同的(即它的突变应该没有调节基因的等位基因的行为);这种突变体特有的、可预期的性质是组成型的等位基因对野生型应该是显性的,因为(实际上还是一种假说)在一个二倍体细胞中,对阻遏物敏感的操纵基因的存在,并不妨碍对阻遏物不敏感的操纵基因的操纵作用。
组成型操纵基因突变
至今,具有上面预料那些性质的组成型突变体,已在两个受阻遏物控制的系统,即在大肠杆菌的λ噬菌体和Lac系统中发现了。
在λ噬菌体系统中,由于这些突变体在免疫细菌中能进行营养发育,而在野生型细菌中则是溶源的,因此很容易挑选,而且已对这类突变体进行了鉴定。这一特性意味着这些突变体(ν)对溶源细胞中的阻遏物是不敏感的。事实上,用这些突变体颗粒侵染溶源细胞时,也诱发了野生型的原噬菌体的发育,产生的噬菌体群体是一种ν和ν+ 颗粒的混合物。这和预料是一致的,据推测原噬菌体发育的起始只依赖于由烈性噬菌体颗粒所提供的一种或少数几种早期酶蛋白的形成(Jacob和Wollman,1953)。
在Lac系统中,已经分离到一些显性的组成型(OC )突变体,分离方法是在Lac区域二倍体的细胞中选择组成型,因而,实际上是除去了隐性的(i- )组成突变体(Jacob等,1960a)。通过重组试验,把这种OC 突变在Lac区域的位置进行了遗传作图,它位于i和z位点之间,顺序是(Pro)yzoi(Ad)(见图3)。这类突变体的某些特性列于表3。首先,我们只讨论这种突变对半乳糖苷酶合成的影响。值得注意的是无诱导物时,这些菌株合成的半乳糖苷酶的量是i- 突变体合成量的10%~20%,即是未经诱导的野生型细胞的100~200倍(见表3的第三行和第七行)。在有诱导物存在时,它们合成的酶量最高。因此,它们只是部分组成型突变[当饥饿条件下,它们在无诱导物存在时也形成最大量的半乳糖苷酶,这一情况除外。(Brown,未发表的结果)]。这里很重要的一点是这种酶具有OC /O+ 结构的二倍体细胞组成型酶的特性(见表3)。因此OC 等位基因是显性的。
表3 用单倍体和杂合二倍体的操纵基因突变体合成半乳糖苷酶、交叉反应物质(CRM)以及半乳糖苷乙酰基转移酶①
①培养细菌用甘油做碳源。注明诱导的就加10-4 M的异丙基硫代半乳糖苷。表中半乳糖苷酶和乙酰基转移酶的数值是用与诱导的野生型酶活百分比表示的。CRM值是用半乳糖苷酶抗原的当量表示的。注意:与性因子携带的等位基因相对应的蛋白质产生的量,常常比受诱导的单倍体野生型产生的量要多得多。推测这是由于每个染色体上存在几个F-Lac因子拷贝造成的。单倍体或双倍体的各种OC 突变体,特别在非诱导培养物中,产酶绝对值随着培养条件的不同,会有很大变化。
②原文此处有误。——译者注
如果OC 突变体的组成性是由于操纵基因丧失了对阻遏物的敏感性造成的,那么OC 菌株对i+ 基因的iS (显性的)等位基因合成的、发生了变化的阻遏物的存在也应该是不敏感的(见前面的有关叙述)。具有iS O+ /Fi+ OC 结构的二倍体的组成型行为(见表3的第十二行)表明,上述情况是确实的,这也是对这两种突变(iS 和OC )的影响所做解释的有力证明。另外,根据这种解释,可以预料到在iS 细胞的群体中,OC 突变体如同Lac+ 的回复突变体一样(见前面的有关叙述)也经常出现。
因此,我们推断这种O+ →OC 突变相当于操纵基因的一种特异的、接受阻遏物结构的修饰作用。我们认为这种突变是在操纵基因的位点上,即在与操纵基因的结构有关的遗传片断上,而不是在操纵者本身上。
操纵子
现在转到另一个问题上来,我们注意到OC 突变(和i- 突变类似)是多效的,它同时而且在数量上以同样程度影响着半乳糖苷酶和乙酰基转移酶的合成(见表3的第七行和第八行)。由此看来,控制着这两种蛋白质合成的这个(或这些)操纵基因是受同一个决定因子控制的 【96】 。
对这个情况有两种可供选择的解释必须考虑:
(1)一个操纵基因控制着z—y遗传片段或其细胞质产物的全部特性;
(2)这个操纵基因位点的特异性产物能够在细胞质中与z和y顺反子的产物结合,从而控制着这两个结构基因的表达。
第二种解释暗示了这个操纵基因位点的突变在行为上应当就像z和y顺反子中的一个独立的顺反子的行为一样。相反,第一种解释则需要这些突变在功能上的行为,就像是它们同属于这两个顺反子一样。因此,对这两种选择性的解释,通过测定O等位基因的转移效应是可以区别的,而与操纵基因作用的任何一种特殊物理模型都是无关的。所谓测定O等位基因的转移效应,就是测定在O+ /OC 二倍体中,这两种结构基因的组成型对诱导型的表达,杂合基因对这两种结构基因中的一种或两种的表达。
从各种结构二倍体得到的结果示于图3。首先,我们注意到了具有O+ z+ /FOC  或O+ /FOC z+ (见表3的第四行和第五行)结构的二倍体,在有诱导物存在时,由z+ 等位基因产生的正常的半乳糖苷酶和由z- 等位基因产生的改变了的蛋白质(CRM)都能形成,而在无诱导物存在时,只能产生与OC 顺位的z等位基因相对应的蛋白质。因此,OC 对反位的z等位基因没有影响。或者反过来说,即使在反位上有OC 存在,与O+ 相连的z等位基因的表达还是对阻遏物十分敏感的。可以说O位点行为和z记号属于同一顺反子。但正如我们已经知道的,OC 突变对乙酰基转移酶同样有影响,乙酰基转移酶属于和z无关的并与操纵基因位点不相邻的另一个顺反子。表3中,第八行和第九行的结果证实了:O→y关系和O→z关系一样,这就是,OC 等位基因的影响可以向外延伸到顺位上的y等位基因。例如,在O+ z- y+ /FOC z+ yū二倍体中,半乳糖苷酶是组成型的,乙酰基转移酶是诱导型的,而在O+ z+ yū/FOC z+ y+ 二倍体中,两种酶都是组成型的。
或O+ /FOC z+ (见表3的第四行和第五行)结构的二倍体,在有诱导物存在时,由z+ 等位基因产生的正常的半乳糖苷酶和由z- 等位基因产生的改变了的蛋白质(CRM)都能形成,而在无诱导物存在时,只能产生与OC 顺位的z等位基因相对应的蛋白质。因此,OC 对反位的z等位基因没有影响。或者反过来说,即使在反位上有OC 存在,与O+ 相连的z等位基因的表达还是对阻遏物十分敏感的。可以说O位点行为和z记号属于同一顺反子。但正如我们已经知道的,OC 突变对乙酰基转移酶同样有影响,乙酰基转移酶属于和z无关的并与操纵基因位点不相邻的另一个顺反子。表3中,第八行和第九行的结果证实了:O→y关系和O→z关系一样,这就是,OC 等位基因的影响可以向外延伸到顺位上的y等位基因。例如,在O+ z- y+ /FOC z+ yū二倍体中,半乳糖苷酶是组成型的,乙酰基转移酶是诱导型的,而在O+ z+ yū/FOC z+ y+ 二倍体中,两种酶都是组成型的。
对这些观察,由第一种解释和第二种解释所得到的推断是矛盾的,因此,产生了一种新的推断,即操纵基因控制着整个ozy遗传片段或它的细胞质产物的全部特性(Jacob等,1960a;Képès, Monod和Jacob,1961)。
由此可以得到另一个推论:某些O片段上的突变,可以通过使整个ozy片段失活的方式来改变操纵基因,造成合成半乳糖苷酶和透性酶的能力都丢失。
这些Oo 突变体对O+ 或OC 应是隐性的,而且它们不能被o+ z+ y- 或o+ z- y+ 突变体所互补。已分离到恰好具有这些性质的几个点突变体(Jacob等,1960a)。正如预料的那样,它们在遗传图上都紧挨着OC (见图3)。有趣的是:发现在这些突变体中,i+ 基因是有功能的(见表3的第十一行),这清楚地表明,不仅i和O突变体不是等位基因,而且控制着z和y基因表达的O片段并不影响调节基因的表达。
总之,ozy遗传片段的整体或协调表达暗示了控制这种表达的操纵基因(见图6)。
图6 蛋白质合成的调节模型
(1)或者与这些基因本身相连(见图6,Ⅰ);
(2)或者与连锁的z和y基因的细胞质信使相连。
在第二种情况下,必需假定这些基因形成一种和ozy片段整体结构相对应的单一的完整颗粒,而且作用上也和一个整体一样(见图6,Ⅱ)。
在第一种情况下,事实上操纵基因应当与O位点一致,它直接控制着这些基因的活性,即结构信使的合成。
这两种模型与到目前为止我们所讨论过的观察都是不矛盾的。在下一部分中,我们将讨论操纵基因(即与阻遏物相互作用的特异部位)是在遗传上,还是通过细胞质起作用的。这两种情况中,尽管ozy片段起码含有两个独立的结构基因,这两个结构基因分别控制着两种独立的蛋白质,但ozy片段在信息传递中都是以一个单位起作用的。这种协调表达的遗传单位,我们把它叫做操纵子(Jacob等,1960a)。
迄今为止,这样一种遗传表达单位的存在,只在Lac片段中得到了证实。正如我们已经见到的,在λ噬菌体系统中,当说明有一个操纵基因存在时,λ噬菌体的ν突变体不能明确定义一个操纵子(因为由这个操纵基因所控制的结构基因的数目和功能是未知的)。然而,有很多迄今为止用经典遗传学理论不能解释、甚至有冲突的观察,用操纵子理论立即便可解释。在细菌中,已经很清楚地知道,控制着和代谢途径有关的各种酶的合成的一些基因,常常发现是紧密连锁的,形成一个基因丛(Demerec,1956)。为了说明遗传结构和生化功能的这种明显相关性,已提出了各种不十分令人信服的假说(Pontecorvo,1958)。因为现在已确定,在这些代谢系统中,同时发生的诱导或同时发生的阻遏一般都是成功的。这样看来,很可能这种基因丛代表着协调表达的一些单位,即操纵子。
我们已经提到过,相继卷入大肠杆菌半乳糖代谢的两种诱导酶,即半乳糖激酶和UDP-半乳糖转移酶,能同时被半乳糖诱导,或被安慰诱导物D-岩藻糖所诱导(Buttin,1961)。控制着这些酶合成的基因(即可能是结构基因)是紧密连锁的,它们在大肠杆菌染色体上形成一个基因丛(Kalckar, Kurahashi和Jordan,1959;Lederberg,1960;Yarmolinsky和Wiesmeyer,1960;Adler,未发表的结果)。发生在这个染色体片段上的某些点突变,使细菌失去了合成这两种酶的能力。这些多效性的丧失突变是不能由任何一种专一性(结构的)丧失突变来补足的,这种现象显然与一个基因一种酶的假说是直接矛盾的。假设这些连锁的结构基因组成受同一个操纵基因控制的一个操纵子,而且这些多效的突变就是这个操纵基因位点的突变,那么上面所说的相关性就可以得到解释,所说的那种矛盾也就解决了。
我们还讨论了沙门氏杆菌中,控制着和组氨酸合成有关的酶合成的共阻遏系统。这个系统包括8个或9个反应步骤。催化其中5个反应的酶已被鉴定出来了。分别决定着这些酶的基因在沙门氏杆菌的染色体上形成一个紧密的连锁群。这些基因中,每一个基因发生突变都会造成合成一种酶的能力的丧失;然而,发生在这个基因丛一端的某些突变,会使全部酶的合成能力都同时丧失,而且这些突变是不能由任何一种专一性的突变来补足的(Ames, Garry和Herzenberg,1960;Hartman, Loper和Serman,1960)。这会使人想起,在这个反应系统中,在任何一种条件下,各种酶合成的相对速率是恒定的(参看前面的有关叙述)。假定这个基因丛组成一个操纵子,受一个与g顺反子相连的操纵基因的控制,那么上述所有的这些值得注意的发现都可以得到解释。
控制着代谢反应中几个连续反应的酶的各个基因组成基因丛的规律,一般来说不适用于细菌以外的生物(Pontecorvo,1958)。也不是对所有的细菌系统都适用,即使已知发生共阻遏、并受同一调节基因控制的系统,也不一定都适用,控制精氨酸生物合成的各种酶就是一个明显的例子。在这样的情况下,就要假设有几个相同或相似的操纵基因位点,分别对一种独立的信息传递系统的阻遏物是敏感的。
很清楚,当一个操纵基因只控制一个结构顺反子表达时,操纵子的概念是不适用的,事实上,遗传学-生物化学测定也难以证实这种控制着操纵基因的遗传片段与结构顺反子本身有什么不同 【97】 。因此,如果在实验上可能把这个概念扩大到分散的(与丛集是相反的)遗传系统上,那将是不可思议的。应该注意很多酶蛋白显然是由两种(或两种以上)不同的多肽链组成的。预期这样的蛋白质将常被发现是由两个(或多个)相邻而且协调表达的结构顺反子(形成一个操纵子)控制着。
结构基因表达的动力学和结构信息的性质
在这一部分中我们要讨论的问题是阻遏物-操纵基因系统,是通过控制结构信息的合成,在遗传水平上起作用呢?还是通过控制信使的蛋白质合成活性,在细胞质水平上起作用呢(见图6)?我们将分别把这两种想到的模型,定名为遗传操纵基因模型和细胞质操纵者模型。
只从细胞质单位所应该达到的大小来看,包括几个结构基因的协调表达单位的存在,在事实上看来与细胞质操纵者模型难于统一。假定信息是一种多核苷酸,取编码率为3,如果一个操纵子控制着平均分子量(单体)为60000道尔顿的3种蛋白质的合成,那么与它相对应的单位信息分子量应当约为1.8×106 ;然而我们已经知道,包括8个以上结构顺反子的操纵子,实际上也是存在的。另一方面,从大肠杆菌和其他细胞的RNA看来,多核苷酸分子的分子量都不超过106 。
这些异议看来是可以澄清的;从我们所了解的现有情况来看,这方面的证据太少,不能排除细胞质操纵者模型,也不能肯定遗传操纵基因模型的可靠性。然而,看来采用遗传操纵基因模型似乎更有利,而且它暗示的某些特异性的预料已被实验所证实。
这些含义中最直接、可能也是最明显的就是结构信息必须由一种十分短命的中间产物来携带,这种中间产物在信息传递过程中快速形成、快速消失。这是诱导的动力学所要求的。正如我们已经见到的,加入诱导物,或除去辅阻遏物,在短短的几分钟之内便可刺激酶合成达到最高速率,而除去诱导物,或加入辅阻遏物,在同样短的时间内,就会阻断酶的合成。这样的动力学与以前的假说是矛盾的,以前认为阻遏物通过和操纵基因相互作用,控制着稳定的合成酶模板的合成速率(Monod,1956;1958)。因此,假说遗传操纵基因模型是可靠的,应该可以期望结构基因表达的动力学与诱导的动力学是基本相同的:一个新基因注入另一个感受态细胞中去,事实上应该导致以最大速率立即合成相应的蛋白质;随着这个基因的去除,蛋白质的合成也将随之而停止。
半乳糖苷酶结构基因表达的动力学
在细胞中加入基因,或从细胞中除去基因,要比诱导物的加入或除去更难完成。但还是可以做到的。在大肠杆菌的雄性Hfr和雌性F- 的接合中,可以只注入基因,而不发生细胞质的混合。在一个雌、雄混合群体中,各对细胞不是在同一时间接合的,但是用适当的遗传学方法,可以相当精确地测定出一个指定基因在何时注入。z+ (半乳糖苷酶)基因从雄性细胞进入不产半乳糖苷酶(z- )的雌性细胞,继之,在合子中酶的合成很快就发生了(参考前面有关叙述)。当群体中,酶合成的速率成为时间的函数时,考虑到随着时间的增加,含有z+ 的合子数目也增加了,进而发现(见图7):①酶的合成在z+ 基因进入2分钟内开始;②每个合子的酶合成速率是恒定的,而且至少在基因进入40分钟后,才达到最大值(Riley, Pardee, Jacob和Monod,1960)。
图7 诱导型、能产生半乳糖苷酶的雄性细胞和组成型和不产半乳糖苷酶的雌性细胞之间,通过接合形成的部分合子的酶形成动力学
在实验条件下,只有合子能形成酶。在适当的选择性培养基上,对重组体计数,测定带有z+ 记号的合子数的增加。在整个群体中,酶形成后随即就测定酶活性。可以看出,酶随着时间的平方成直线增加。因为合子数目随着时间的增加成直线增加,所以从z+ 基因进去的时间起,每个合子酶合成的速率显然是不变的(Riley等,1960)。
这些观察表明了z+ 基因能快速形成结构信使,而且不累积下来。这可用下述两种方式中某一种来解释:(1)结构信使是一种短命的中间产物;(2)结构信使是稳定的,但是这个基因快速形成少数信使分子,此后就停止作用了。
假定第二个假设是正确的,那么在酶开始合成后,去掉这个基因,应该不妨碍酶的继续合成。可以用去除基因的实验来验证这种可能性,其作法是在注入前把雄性染色体用32 P标记;随后注入未标记的雌性细胞中去,并给充足的时间(25分钟)让Z+ 基因表达,然后将合子冷冻使32 P衰变不同的时间。在溶化后立刻测定群体中半乳糖苷酶合成的速率。发现酶合成的快速下降是衰变32 P原子的函数。假如在冰冻前z+ 基因有较长的表现时间(110分钟),则观察到形成酶的能力或Z+ 记号的生存能力都不下降。这是我们所期望的,因为到那时,大多数Z+ 基因已完成了复制,而且这个观察还提供了一种与32 P蜕变的间接效应无关的内在控制。
因此,这个实验表明,即使Z+ 基因已经开始表达后,其完整性仍是酶的继续合成所需要的,正如所预料的那样——可能信使分子是一个短命的中间产物(Riley等,1960)。
对注入和去除实验的解释都建立在一个假设的基础上,假设认为观察到的这些效应不是由于在接合过程中,和遗传物质一起导入的(稳定的)细胞质信使分子所造成的。正如我们已经注意到的,有充分证据说明,在接合过程中不发生细胞质的转移,连小分子的转移都没有。再进一步,假设在合子中,酶的合成是由于以前形成的信使分子,而不是基因的活性引起的,那么对下面的两点就很难解释:①酶合成的开始和基因的进入(在接合实验中)之间在时间上的完全一致性;②酶形成的能力和Z+ 基因的遗传生存能力(在去除实验中)的平行行为。
因此,这些实验看来似乎表明,一个结构基因表达的动力学与诱导-阻遏的动力学是完全相似的。正如所预料的,可能在合成蛋白质过程中,操纵基因是控制着合成一种短命信使的基因的活性,而不是控制着一种已经合成了的(稳定的)信使分子的活性。
在这一点上,回顾一下用烈性噬菌体( 11 、T2、T4)侵染大肠杆菌,在2~4分钟内,就能使细菌蛋白质合成(特别是包括β-半乳糖苷酶)随之发生抑制,这是很有意义的(Cohen,1949;Monod和Wollman,1947;Benzer,1953)。另一方面,已知噬菌体的侵染,引起细菌核体迅速而显著的溶解,但是对以前形成的细菌RNA似乎不发生大的破坏(Luria和Human,1950)。从这些情况来看,烈性噬菌体对特异的细菌蛋白质合成的抑制作用可能主要是由于细菌DNA的解聚作用引起的。这个结论也暗示,细菌基因的完整性是细菌蛋白质的继续合成所需要的。在证明这种解释的过程中,我们注意到了用λ噬菌体侵染大肠杆菌,并不引起细菌核体的破坏,β-半乳糖苷酶的合成几乎能一直持续到溶菌的时间(Siminovitch和Jacob,1952)。
碱基类似物的结构效应
从另一个完全不同类型的实验得出的结论,也认为结构信使是一种短命的中间产物,并进一步提示,这种中间产物是一种核糖核苷酸。已知细菌细胞能把某些嘌呤和嘧啶类似物掺入到核糖核酸和脱氧核糖核酸中去,还发现某种(或某些)蛋白质的合成可以被某些这样的类似物所抑制。解释这些效应的机制之一,认为可能是因为某些类似物掺入到结构信使中去了。如果确实是这样,就可能看到,在一种类似物存在时,形成的特异性蛋白质的分子结构发生了变化。事实上已经发现,在5-氟尿嘧啶(5FU)存在的情况下,大肠杆菌合成的β-半乳糖苷酶和碱性磷酸酶的分子性质发生了明显的改变。就以β-半乳糖苷酶的情况来说,酶活性与抗原的效价之比下降了80%。在碱性磷酸酶的情况下,热失活的速率(这种蛋白质通常是高度抗热的)大大增加(Naono和Gros,1960a, b;Bussard, Naono, Gros和Monod,1960)。
完全可以认为这种影响不可能仅仅是细胞中5FU的存在造成的,必定反映了这种类似物以某种方式掺入到和这个信息转移系统有关的一种组分中去了。不管这个组分本身是什么,这种效应的动力学本身也必定反映了5FU掺入这个组分的动力学。加入5FU就开始合成不正常的酶,从这种意义上看,5FU效应最显著的特点是即时性,而且此后合成的酶分子群体中,异常程度并不随着时间的增长而增加。例如,在半乳糖苷酶的情况下,加入这种类似物在5分钟内就合成异常的酶,此后酶活性与抗原效价的比保持不变。在碱性磷酸酶的情况下,在5FU存在时,合成的异常蛋白质的热失活曲线是单一的,这表明这种分子群体是均匀异常的,而不是由正常和异常分子组成的一种混合物。很清楚,如果与这种效应有关的这种组分是稳定的,就可以预期,在5FU存在时形成的分子群体是异种的,而异常的分子部分会逐渐增加。由此可见,这种有关的组分必定迅速形成,又迅速被破坏。
现在还应该注意,除了结构基因合成的信使外,这种信息传递系统可能还涉及与信息的正确翻译有关的其他组分,例如与氨基酸转移有关的RNA。5FU的这种影响可能是由于它掺入某种转移RNA引起的,而不是由于掺入信使本身引起的。综观上面讨论的各种实验结果,有力地说明了5FU的影响反映了信使本身周转率很高。
信使RNA
先暂时接受上述结论,然后让我们考虑,一种细胞成分需要具备什么样的性质,才能使它与结构信使一样呢?根据一般的推测和上面讨论的结果,应具有下列条件:
(1)它必须是一种多核苷酸。
(2)就分子量来说,它们可能是十分不同的,但假定编码比率是3,那么平均分子量将不低于5×105 道尔顿。
(3)它的碱基组成应该反映出DNA的碱基组成。
(4)至少短暂地、或在某些条件下,应该发现它与核糖体是偶联的,因为已有理由证实核糖体是蛋白质合成的场所。
(5)它应该有很高的周转率,特别是它在不到5分钟内就应该被5FU饱和。
很快就搞清楚了,用经典方法已经辨认出来的细胞RNA都不符合这些特定的条件。人们常常推测核糖体RNA(rRNA)可能代表蛋白质合成的模板,它们分子量非常接近。但在不同种类中,rRNA的碱基组成是相似的,而且不能反映出DNA中碱基比例的变化,此外,在生长细胞中,它似乎是十分稳定的(Davern和Meselson,1960)。它掺入5FU只与纯增加成正比。
转移RNA,或(sRNA)不能反映DNA的碱基组成。它的平均分子量比信使所要求的分子量(5×105 道尔顿)要低得多。可能除了末端的腺嘌呤和胞嘧啶外,它的碱基掺入速率(包括特异性的5FU)不比核糖体RNA的高。
然而,有一种RNA,看来各种性质都符合上面提出的条件。这种RNA首先由Volkin和Astrachan(1957)在受噬菌体侵染的大肠杆菌中发现,最近发现在正常的酵母(Yčas和Vincent,1960)和大肠杆菌(Gros等,1961)中也存在。
这种RNA(我们把它叫做信使RNA或mRNA)仅约占总RNA量的3%;通过柱分离或沉降可以把它和其他RNA分离开(见图8)。它的平均沉降速度系数是13,相当于最小分子量是3×105 道尔顿,但是,据推测这种分子根本不是球形的,所以分子量可能要高得多。32 P尿嘧啶或5FU掺入这种RNA的速度是极快的,发现不到30秒就可达到半饱和状态,这表明它的合成速度比任何其他RNA要快几百倍。从预先标记过的细胞中这种RNA放射性的消失情况可以得知,它的半衰期是很短的。较高浓度的Mg2+ (0.005M),有利于这种RNA和70S核糖体颗粒的结合,当Mg2+ 浓度较低时,它发生沉降,但与核糖体颗粒无关(Gros等,1961)。
图8 在信使RNA中,尿嘧啶的掺入和周转情况
把大肠杆菌在肉汤中培养到对数生长期,再与[14 C]-尿嘧啶一起保温5秒钟。经离心、洗涤、再悬浮于同体积、但含有100倍以上[12 C]-尿嘧啶的同种培养基中。把一半细菌收集、冰冻(Ⅰ图);剩下的一半在37℃,保温15分钟,再收集、冰冻(Ⅱ图)。冰冻样品用铝粉研磨,用含10-4 克分子Mg的Tris缓冲液(2-氨基-2-羟甲基丙烷-1∶3-二醇)抽提,再用DNase处理,用于蔗糖梯度。3小时后,连续取样测定放射性和在260纳米的吸光率。可以看出(Ⅰ图),5秒钟以后,只有mRNA被标记了;还可看出(Ⅱ图)以后尿嘧啶掺入mRNA完全是重新开始的(见Gros等,1961)。
Volkin和Astrachan发现的一个最著名的事实,是在T2噬菌体侵染过的细胞中,这种RNA的碱基组成反映了噬菌体(而不是细菌的)DNA的碱基组成。根据这个事实推断,这种RNA的出现是要开始合成噬菌体DNA的预兆。这种RNA性质和一种短命的结构信使性质之间的一致性证实了,在受噬菌体侵染的细胞以及正常细胞中,事实上这种RNA的作用是把遗传信息从噬菌体DNA传递到蛋白质合成中心上来。这个推论暗示了同一种蛋白质合成中心,在细胞未受侵染时,合成细菌的蛋白质,在细胞受到侵染后,它可以根据噬菌体DNA提供的新结构信息,经过mRNA,来合成噬菌体蛋白质。最近对T4侵染过的大肠杆菌所作的观察,有力地支持了这种解释(Brenner等,1961)。
未受侵染的大肠杆菌细胞,在含有15 N的培养基中进行培养,然后用噬菌体侵染,再悬于14 N培养基中。侵染后,它们经短时间的32 P或35 S脉冲处理,并在密度梯度中分析核糖体,有以下发现:
(1)侵染后测不出有rRNA被合成。
(2)侵染后形成的标记mRNA与侵染前形成的未标记的核糖体颗粒结合了。
(3)发现新形成的(即由噬菌体决定的)蛋白质(根据它的35 S含量来鉴定),在可溶性蛋白质中出现之前,它是与70S核糖体颗粒结合在一起的。
这些观察证明噬菌体蛋白质是由细菌受侵染前形成的细菌核糖体来合成的,并与噬菌体决定的mRNA有关。因为在细菌的核糖体中不可能具备噬菌体蛋白质的结构信息,所以它必定是噬菌体mRNA所提供的。
Lamfrom(1961)的实验最近由Kruh、Rosa、Dreyfus和Schapira(1961)重复出来了。这些实验直接表明在血红蛋白的合成中,种的特异性是由一种含可溶性RNA的部分决定的,而不是由核糖体决定的。Lamfrom所用的重建系统,含有一个种(兔子)的核糖体和另一个种(羊)的可溶性部分,发现通过这个系统离体形成的血红蛋白,部分地属于用来制备可溶性部分的那个种的类型特性。当然,这还不能确切地证明“在血红蛋白结构上的种间差别是由基因决定的,而不是由细胞质决定的”,但是,这个推测看来相当可靠。总之,Lamfrom的实验无疑地证明:不能认为是核糖体完全地决定着(假如果真能决定的话)蛋白质的特异性结构。
对遗传操纵基因模型和细胞质操纵者模型的选择问题,在这一部分中我们已进行了讨论。正如我们已经见到的,遗传操纵基因模型和一种中间产物的行为有关,这种中间产物是十分特殊、而又特异的,并担负着从基因到蛋白质的信息传递任务。这些预见是通过大量的证据得到的,从这些证据推测,所讨论的那种中间产物与一种特殊的RNA是一致的。即使这种一致性被直接实验证实了,还需要通过直接的实验来证实这种“mRNA”的合成是通过阻遏物-操纵基因的相互作用,在遗传水平上进行控制的。
结论
总结本文前面各部分所得结论的一个简便方法就是把它们组织到一个设计的模型中去,把在蛋白质合成的控制中起到特异性作用的那些主要成分具体表现出来;这些主要成分分别叫做结构基因、调节基因、操纵基因、操纵子以及细胞质阻遏物。这样的一种模型可能如下所述:
蛋白质的分子结构是由特异的成分——结构基因决定的。它们通过形成一种它们本身的细胞质转录抄本,即结构信使而起作用,从这种结构信使再合成蛋白质。由结构基因合成信使是一个连续的复制过程,复制只能在DNA链的某些位点上起始,几个连锁的结构基因的细胞质转录可能是依赖于同一个起始点或叫操纵基因。这些协调表达其活性的基因形成一个操纵子。
操纵基因能和具有适当(互补的)顺序的某个特定(RNA)部分进行可逆结合(由于具有一种特殊的碱基顺序)。通过这种结合阻断细胞质转录的起始,因此阻断整个操纵子中由结构基因形成信使的过程。和一个特定的操纵基因起作用的这种特异性阻遏物(RNA?)是由一个调节基因合成的。
在某些系统中(诱导酶系统),阻遏物能与某些小分子特异性地结合。这种被结合的阻遏物对操纵基因是没有亲和力的,因此这种结合引起操纵子的活化。
在另一些系统中(阻遏酶系统),阻遏物本身是无活性的(即它对操纵基因无亲和力),它只有通过和某些特异性的小分子结合才能被活化。因此这种结合导致操纵子的抑制。
结构信使是一种不稳定的分子,它在信息传递过程中受到破坏。因此,信使合成的速率本身又控制着蛋白质合成的速率。
这个模型意在能简便地概括和表达在蛋白质合成的控制中,起特异性作用的各种因子的性质。为了具体地说明这些不同因子的作用,我们不得不引用一些纯属推理的假设。让我们把实验证实的结论和这些推论明确区分一下。
(1)这些结论最肯定的地方是确有调节基因存在,它控制着从结构基因到蛋白质的信息传递速率,但它们本身不给予蛋白质任何信息。这方面的证据是结构基因上的突变能反映在蛋白质上有所改变,却不改变调节机制。能改变调节机制的突变并不改变蛋白质,突变位点作图也不在结构基因上。结构基因服从一个基因一种蛋白质的基本原则,而调节基因可以影响几种不同的蛋白质的合成。
(2)调节基因通过一种特异的细胞质物质起作用,这种细胞质物质的效应是抑制结构基因的表达,这一点从下述几个方面已得到明确证实:基因的转移效应;按细胞质来源看,在遗传学上一致的一些合子表现出不同的性质;无调节基因(或无调节基因产物)会导致以最大速率无控制地合成蛋白质。
(3)在Lac系统(以及λ噬菌体溶源系统)的情况下,通过调节基因显性突变体的特性已证实,调节基因的产物是直接作为一种阻遏物(不是间接地作为一种胞内诱导物或其他活性物的拮抗物)起作用的。
(4)阻遏物化学上与RNA的一致性,这只是根据证明它不是一种蛋白质而做出的一种逻辑推论。
(5)操纵基因是阻遏物作用的部位,它的存在是从阻遏物的存在和作用特异性推测出来的。发现操纵基因控制着几个相邻结构基因的表达。换句话说,证明操纵子是遗传表达的一个协调单位,这有力地揭示了操纵基因与控制着对阻遏物敏感性的遗传片段是一致的。
操纵基因代表几个结构基因的细胞质转录起始点,这只是一个纯粹的推测,意在说明操纵基因控制着形成一个操纵子的一组连锁基因的整体性。目前,关于这个问题还没有证据说明任何一种推测是建立在操纵基因的分子机制基础上的。
(6)在这个模型中,有关阻遏物与诱导物或辅阻遏物的相互作用的推测是根据最不充分、而且最含糊的。诱导物与阻遏物的特异性结合可以造成阻遏物的失活,这一想法看来是很有道理的,但是它也引起了一种我们已经指出过的困难。因为在阻遏物和诱导物之间发生的这种反应必定是立体特异反应(二者都是这样),所以,推测反应应该需要一种特异的酶;至今,在遗传学上或生物化学上都没有发现这种酶存在的证据。
(7)认为结构信使是一种不稳定的中间产物,这是这个模型最新最特殊的含义之一;让我们回顾一下,只要认为控制系统是在遗传水平上起操纵作用,那么诱导的动力学就需要具有这种特性的结构信使。这就产生了信息传递机制的一个新概念,在信息传递中,蛋白质合成中心(核糖体)根据它们通过mRNA从基因接受的特异性指令,合成不同的蛋白质,核糖体起非特异性组分的作用。
从动力学研究和分析研究都有证据支持对信息传递的这一新解释,即使这个模型中所包括的一些其他推测,将来也可能弄清楚是错误的,但这种解释本身也还是具有重要意义的。
这些结论完全适用于得出这些结论的细菌系统;而且,两种类型(诱导和阻遏)的适应酶系统和噬菌体系统都遵循着和同种必需成分有关的同种控制,这一事实有力地证明了蛋白质合成的阻遏遗传调节的普遍性。
人们不知道是不是全部的、或大多数的结构基因(即大多数蛋白质的合成)都服从于阻遏调节。在细菌中,事实上已作过一些研究的所有酶系统,业已证实对诱导或阻遏效应都是敏感的。老的观念认为这些效应只是非主要酶的特性,这肯定是错误的[当然,这些效应只有在天然的或人工的条件下才能被检查出来,这样以至于使研究中的系统至少一部分是非主要的(安慰的)]。能去除这种控制的突变(如组成型突变)结果,说明了控制的生理学重要性。乳糖系统的组成型突变体,合成的β-半乳糖苷酶可以达到其总蛋白质的6%~7%。磷酸酶系统的组成型突变体中,总蛋白质的5%~6%是磷酸酶。用其他组成型突变体也得到了相似的数值。很清楚,如果细胞中控制着酶蛋白合成速度的2个或3个以上的控制系统受到破坏,那么这种细胞很快就会死亡。
高等生物组织中诱导和阻遏效应的发生,已经观察到了很多例子,但到目前为止,对这些系统中的任何一个都还不能进行详细分析(主要困难是安慰性控制条件不好确定)。已经反复指出(和在微生物中研究的情况相似),酶的适应性为解释高等生物组织内和器官间的生化协调性,提供了一个有价值的模型。在微生物中,适应效应主要是负的(阻遏),而且由功能上特异的基因控制着,在遗传水平上起操纵作用,这些证据看来大大地扩大了这种解释的可能性。化学生理学和胚胎学的根本问题就是要弄清:为什么在组织细胞的基因组中不是在所有的时间、所有的潜在遗传性都表达呢?有机体的存活需要很多(在一些组织中则是大多数)潜在遗传性不表达出来,也就是说受到抑制。对恶性肿瘤最合适的描述是其一种或几种控制生长的系统受到了破坏,这种破坏无疑是在遗传上引起的。
按照严格的结构概念来看,基因组被认为是用于建立各种细胞组分的各个独立分子行动计划的一个嵌合体。然而,在这些计划的实施中,协调显然是生存所不可缺少的。调节基因、操纵基因以及结构基因活性的阻遏调节等发现,揭示了基因组不仅含有一系列的行动计划,而且还含有蛋白质合成的一个协调程序和控制其实施的方式。
参考文献
[1]Adelberg, E. A., and Umbarger, H. E., 1953. J. Biol. Chem.,205 , 475.
[2]Ames, B. N., and Garry, B., 1959. Proc. Nat. Acad. Sci., Wash., 45 , 1453.
[3]Ames, B. N., Garry, B, and Herzenberg, L. A., 1960. J. Gen. Microbiol.,22 , 369.
[4]Benzer, S., 1953. Biochim. Biophys. Acta.,11 , 383.
[5]Bertani, G., 1953. Cold. Spr. Harb. Symp. Quant. Biol.,18 , 65.
[6]Bertani, G., 1958. Adv. Virus Res.,5 , 151.
[7]Brenner. S., Jacob, F. and Meselson, M., 1961. Nature, 190 , 576.
[8]Bussard, A., Naono, S., Gros, F., and Monod, J., 1960. C. R. Acad. Sci., Paris, 250 , 4049.
[9]Buttin, G., 1956. Dipiôme Et. sup., Paris.
[10]Buttin, G., 1961. C. R. Acad. Sci., Paris, (in press).
[11]Cohen, G. N. and Jacob, F., 1959. C. R. Acad. Sci., Paris, 248 , 3490.
[12]Cohen, S. N. and Monod, J., 1957. Bact. Rev.,21 , 169.
[13]Cohen, S. S., 1949. Bact. Rev.,13 , 1.
[14]Cohen-Bazire, G. and Jolit, M., 1953. Ann. Inst. Pasteur,84 , 1.
[15]Cohn, M., 1957. Bact. Rev.,21 , 140.
[16]Cohn, M., Cohen, G. N. and Monod, J., 1953. C. R. Acad. Sci., Paris, 236 , 746.
[17]Cohn, M. and Horibata, K., 1959. J. Bact.,78 , 624.
[18]Cohn. M. and Monod, J., 1953. In Adaptation in Micro-organisms, 132. Cambridge University Press.
[19]Cohn, M., and Torriani, A. M., 1952. J. Immunol.,69 , 471.
[20]Davern, C. I., and Meselson, M., 1960. J. mol, Biol.,2 , 153.
[21]Demerec, M., 1956. Cold Spr. Harb. Symp. Quant. Biol.,21 , 113.
[22]Dienert, F., 1900. Ann. Inst. Pasteur,14 , 139.
[23]Duclaux, E., 1899. Traité de Microbiologie. Paris: Masson et Cie.
[24]Echols, H., Garen, A., Garen, S., and Torriani, A. M., 1961. J. mol. Biol., (in press).
[25]Fiaks, J. G., and Cohen, S. S., 1959. J. Biol. Chem.,234 , 1501.
[26]Gale, E. F., 1943. Bact. Rev., 7 , 139.
[27]Giles, N. H., 1958. Proc. Xth Intern. Cong. Genetics, Montreal, 1 , 261.
[28]Gorini, L., and Maas, W. K., 1957. Biochim. biophys. Acta,25 , 208.
[29]Gorini, L., and Maas, W. K., 1958. In The Chemical Basis of Development, Baltimore. Johns Hopkins Press. p.469.
[30]Gros, F., Hiatt, H., Gilbert, W., Kurland, C. G., Risebrough, R. W. and Watson, J. D., 1961. Nature,190 , 581.
[31]Halvorson, H. O., 1960. Advanc. Enzymol., (in press).
[32]Hartman, P. E., Loper, J. C., and Serman, D., 1960 J. Gen. Microbiol.,22 , 323.
[33]Herzenberg, L., 1959. Biochim. biophys. Acta.,31 , 525.
[34]Hogness, D. S., Cohn, M., and Monod, J., 1955. Biochim. biophys. Acta.,16 , 99.
[35]Jacob, F., 1954. Les Bactéries Lysogènes et La Notion de provirus. Paris: Masson et Cie.
[36]Jacob, F., 1960. Harvey Lectures, 1958—1959, 54 . 1.
[37]Jacob, F., and Adelberg, E. A., 1959. C. R. Acad. Sci., Paris, 249 , 189.
[38]Jacob, F., and Campbell, A., 1959. C. R. Acad. Sci., Paris,248 , 3219.
[39]Jacob, F., Fuerst, C. R., and Wollman, E L., 1957. Ann. Inst. Pasteur,93 , 724.
[40]Jacob, F., and Monod, J., 1959. C. R. Acad. Sci., Paris,249 , 1282.
[41]Jacob, F., Porrin, D., Sanchez, C., and Monod, J., 1960a. C. R. Acad. Sci. Paris, 250 , 1727.
[42]Jacob, F., Schaeffor, P., and Wollman, E L., 1960b. In Microbial Genetics, Xth Symposium of the Society for General Microbiology, 67.
[43]Jacob, F., and Wollman, E. L., 1953. Cold Spr. Harb. Symp. Quant. Biol.,18 , 101.
[44]Jacob, F., and Wollman, E. L., 1956. Ann. Inst. Pasteur,91 , 486.
[45]Jacob, F., and Wollman, E. L., 1957. In The Chemical Basis of Heredity, Baltimore: Johns Hopkins press, 468.
[46]Kaiser, A. D., 1957. Virology, 3 , 42.
[47]Kaiser, A. D., and Jacob, F., 1957. Virology, 4 , 509.
[48]Kalckar, H. M., Kurahashi, K., and Jordan, E., 1959. Proc. Nat. Aoad. Sci., Wash., 45 , 1776.
[49]Karstrom, H., 1938. Ergebn. Enzymforsch., 7 , 350.
[50]Képès, A., 1960. Biochim. biophys. Acta, 40 , 70.
[51]Képès, A., Monod, J., and Jacob, F., 1961. In preparation.
[52]Kogut, M., Pollock, M., and Tridgell, E. J., 1956. Biochem. J., 62 , 391.
[53]Kornberg, A., Zimmerman, S. B., Kornberg, S. R., and Josse, J., 1959. Proc. Nat. Acad, Sci., Wash., 45 , 772.
[54]Kruh, J., Rosa, J., Dreyfus, J. C., and Sehapira, G., 1961. Biochim. biophys. Acta, (in press).
[55]Lamfrom, H., 1961. J. mol. Biol.,3 , 241.
[56]Lederberg, E., 1960. In Microbial Genetics, The Xth Symposium of the Society of General Microbiology. p.115.
[57]Levinthal, C., 1959. In Structure and Function of Genetic Elements, Brookhaven Symposia in Biology, p. 76.
[58]Luria, S. E., and Human, M. L., 1950. J. Bact.,59 . 551.
[59]Lwoff, A., 1953. Bact. Rev.,17 , 269.
[60]Lwoff, A., Siminovitch, L., and Kjeldgaard, N., 1950. Ann. Inst. Pasteur,79 , 815.
[61]Magasanik, B., Magasanik, A. K., and Neidhardt, F. C., 1959. In A Ciba Symposium on the Regulation of Cell Metabolism. London: Churchill, p. 334.
[62]Monod, J., 1942. Recherches sur la Croissance des Cultures Bactériennes Paris: Hermmann; 1955 Exp. Ann. Biochim. Méd,,  . Paris: Masson et Cie, 195.
. Paris: Masson et Cie, 195.
[63]Monod, J., 1956. In Units of Biological Structure and Function, 7. New York: Academic Press.
[64]Monod, J., 1958. Rec. Trav. Chim. des Pays-Bas,77 , 569.
[65]Monod, J., 1959. Angew. Chem.,71 , 685.
[66]Monod, J., and Audureau, A., 1946. Ann. Inst. Pasteur,72 , 868.
[67]Monod, J., and Cohen-Bazire, G., 1953. C. R. Acad. Sci., Paris, 236 , 530.
[68]Monod, J., and Cohn. M., 1952. Adv. Enzymol.13 . 67.
[69]Monod, J., and Cohn, M., 1953. In Symposium on Microbial Metabolism. VIth Intern. Cong. of Microbiol., Rome, 42.
[70]Monod, J., Pappenheimer, A. M., and Cohen-Bazire G., 1952. Biochim. biophys. Acta,9 , 648.
[71]Monod, J., and Wollman. E. L., 1947. Ann. Inst. Pasteur,73 , 937.
[72]Naono, S., and Gros, F., 1960a. C. R. Acad. Sci., Paris, 250 , 3527.
[73]Naono, S., and Gros, F., 1960b. C, R. Acad. Sci., Paris, 250 , 3889.
[74]Neidhardt, F. C., and Magasanik, B., 1956a. Nature,178 , 801.
[75]Neidhardt, F. C., and Magasanik, B., 1956b. Biochim. biophys. Acta,21 , 324.
[76]Novick, A., and Szilard, L., in Novick, A., 1955. Ann. Rev. Microbiol.,9 , 97.
[77]Pardee, A. B., 1957. J. Bact.,73 , 376.
[78]Pardee, A. B., Jacob, F., and Monod, J., 1959. J. mol. Biol.,1 , 165.
[79]Pardee, A. B., and Prestidge, L. S., 1961. In preparation.
[80]Pardee, A. B., and Prestidge, L. S., 1959. Biochim. biophys. Acta,36 , 545.
[81]Perrin, D., Bussard, A., and Monod, J., 1959. C. R. Acad. Sci., Paris, 249 , 778.
[82]Perrin, D., Jacob, F., and Monod, J., 1960. C. R. Acad. Sci. Paris, 250 , 155.
[83]Pollock, M., 1950. Brit. J. Exp. Pathol.,4 , 739.
[84]Pollock, M., and Perret, J. C., 1951. Brit. J. Exp. Pathol.,5 , 387.
[85]Pontecorvo, G., 1958. Trends in Genetic Analysis, New York: Columbia University Press.
[86]Rickenberg, H. V., Cohen, G. N., Buttin, G., and Monod, J., 1956. Ann. Inst. Pasteur,91 , 829.
[87]Riley, M., Pardee, A. B., Jacob, F., and Monod, J., 1960. J. mol. Biol,2 , 216.
[88]Rotman, B., and Spiegelman, S., 1954. J. Bact.,68 , 419.
[89]Siminovitch, L., and Jacob, F., 1952. Ann. Inst. Pasteur,83 , 745.
[90]Stanier, R. Y., 1951. Ann. Rev. Microbiol.,5 , 35.
[91]Szilard, L., 1960. Proc. Nat. Acad. Sci. Wash., 46 . 277.
[92]Torriani, A. M., 1960. Biochim. biophys. Acta,38 , 460.
[93]Umbarger, H. E., 1956. Science,123 , 848.
[94]Vogel, H. J., 1957a. Proc. Nat. Acad Sci Wash., 43 , 491.
[95]Vogel, H. J., 1957b. In The Chemical Basis of Heredity, Baltimore: Johns Hopkins Press, 276.
[96]Volkin, E., and Astrachan, L., 1957. In The Chemical Basis of Heredity, Baltimore: Johns Hopkins Press, 686.
[97]Went, F. C., 1901. J. Wiss. Bot.,36 , 611.
[98]Wijesundera, S., and Woods, D. D., 1953. Biochem. J.,55 , viii.
[99]Willson, C., Perrin, D., Jacob, F., and Monod, J., 1961. In preparation.
[100]Wolhnan, E. L., and Jacob, F., 1959. La Sexualite dés Baetéries: Prais: Masson et Cie.
[101]Yanofsky, C., 1960. Bact. Rev.,24 , 221.
[102]Yanofsky, C., and Lennox, E. S., 1959. Virology,8 , 425.
[103]Yarmolinsky, M. B., and Wiesmeyer, H., 1960. Proc. Nat. Acad. Sci. Wash., (in press).
[104]Yates, R. A., and Pardee, A. B., 1956. J. Biol. Chem.,221 , 757.
[105]Yates, R, A., and Pardee, A. B., 1957. J. Biol. Chem.,227 , 677.
[106]Yčas, M., and Vincent, W. S., 1960. Proc. Nat. Acad, Sci. Wash., 46 , 804.
[107]Zabin, I., Képès, A., and Monod, J., 1959. Biochem. Biophys, Res. Gomm.,1 , 289.
核酸的分子构型 【98】
莫里斯·威尔金斯(M. H.F.Wilkins) 【99】
(1962年)
核酸基本上是简单的,它是非常基本的生物学过程——生长和遗传的基础。核酸分子结构及其功能关系的单纯性,说明了基本生物学现象的单纯性,搞清楚了生物学现象的本质,从而能用大分子结构首次广泛地阐明生命过程。只有通过从遗传学到氢键立体化学,把生物学、化学和物理学的研究进行史无前例的结合,才能弄清楚这些物质。在这里我将不讨论它的全部内容,但将集中谈谈我所工作的领域,并说明X射线衍射分析是怎样作出贡献的。我将介绍本人研究的一些背景,因为我想在找到这类通常比总的评论更有意义的解说方面我并不是孤立的。
早期的背景
1938年我在剑桥经受过X射线结晶学方面的一些训练,取得了物理学学位。以后在Cavendish还受到J.D. Bernal关于X射线衍射的影响。我开始在伯明翰由J. T.Randall领导从事研究工作,研究发光和电子如何移入晶体。我在剑桥的同时代人感兴趣的主要是元素粒子,但是我更感兴趣的是固态组织以及依赖于该组织的特殊性质。这可能是我把兴趣放在生物大分子和大分子结构,同其如此重要地决定生命过程的高度特异性质怎样发生联系的一个先兆。
战争期间,我从事于制造原子弹的工作。当战争结束时,同许多其他人一样,我寻找一个新的研究领域。部分地由于原子弹,我对物理学失去了一些兴趣。所以当我拜读Schrödinger的《生命是什么》一书时,就感到十分有趣,并为生命过程是由一种高度复杂的分子结构所控制的概念所打动。感到对这些问题进行研究较之固态物理学前程更为远大。当时,许多杰出的物理学家,诸如Massy、Oliphant和Randall(以后我得悉Bohr的观点同他们一样)相信,物理学能对生物学作出重要的贡献,他们的忠告鼓励我转向生物学。
我到苏格兰St. Andrews的物理系工作,在那里Randall邀我参加他已经开始的一项生物物理研究课题。在Muller用X射线照射实验改变遗传物质的鼓舞下,我想研究超声的效应或许是有意思的,但结果并不十分令人鼓舞。
然后,把生物物理工作转到伦敦皇家学院,Randall在那里取得了惠斯登物理学讲座的职位,并在医学研究会的协助下,为物理系建立了一个不寻常的实验室,在这个实验室里,生物学家,生物化学家和其他科学家同物理学家并肩工作。他建议我用紫外线显微镜研究细胞核酸的量。该工作取法Caspersson的工作,但用反射显微镜来消色差。当时,Caspersson和Brachet的工作已使科学界一致公认核酸具有同蛋白质合成相关联的重要的生物学作用。然而,DNA本身可能就是遗传物质这一思想仅只暗示而已。DNA在染色体里的功能被假定为同蛋白质染色体丝的复制有联系。Avery、MacLeod和McCarty证明细菌通过DNA能在遗传上转化的工作发表于1944年,但即使在1946年尚几乎为人们所不知,或者纵然知道,其重要性也往往不为人们所重视。
通过显微镜观察细胞里面的染色体,确实令人神往,但我开始感到,作为一个物理学家,研究从细胞中分离出来的大分子,或许对生物学贡献更大些。在这一点上我得到了来自Stanley病毒实验室的Gerald Oster的鼓励,并使我对烟草花叶病毒颗粒感兴趣。正如Caspersson曾经指出,能用紫外线显微镜寻找分子中吸收紫外线的基团的位置,和测定细胞内的核酸量。Bill Seeds和我研究了DNA、蛋白质、烟草花叶病毒、维生素B12 等。当我检验准备作紫外二色性研究的DNA分子束的时候,在偏光显微镜下看到极其均匀的纤维在交叉尼古尔之间有明显光吸收。我发现当我操作DNA凝胶时,这种纤维并非人为地产生。我每次用玻璃棒接触凝胶和移去玻璃棒时,就带出细细的几乎看不清的DNA纤维,就像一根蜘蛛网的细丝。纤维的完整和均匀一致,说明在这些纤维中的分子是有规律地排列的,我立刻想到这种纤维或许是X射线衍射分析研究的绝妙对象。我把它们送交Raymond Gosling,他那里有我们唯一的X射线装备(从战争留下的放射照相部分制造),并且他正用此仪器从公羊精子头部得到衍射照片。在W.L.Bragger领导下受过训练,并从事于X射线衍射工作的Randall指导了这项研究。Gosling很快得到了非常鼓舞人心的衍射图像(见图1)。取得这次成功的原因之一是我们使纤维保持在湿润状态。我们记得,为了从蛋白质得到精细的X射线图像,Bernall把蛋白质晶体保存在它的母液中,看来有可能所有水溶性生物大分子的构型都决定于它们的水的环境。我们用Signer和Schwander制备的DNA得到很好的衍射图像。Singer还把DNA带到伦敦一次有关核酸的Faraday学会讨论会上,慷慨地分赠给大家,使所有的工作人员使用他们的不同技术,都可以研究它。
图1 本实验得到的第一批DNA X射线衍射的照片之一
此照片可与以后图10照片相比较(R. Gosling摄影,signer提供DNA)。
遗传物质是一种纯化学物质,其分子结构特别简单
从1946年到1950年,有许多证据说明遗传物质是DNA,而不是蛋白质或核蛋白。例如,发现一组染色体的DNA含量是恒定的,尽管DNA分子的核苷酸顺序是复杂的,而某一物种的DNA具有恒定的组成。曾设想遗传信息存在于4种核苷酸组成的、顺序复杂的多核苷酸分子中。细菌转化的巨大意义现已得到普遍公认:Hershey和Chase证明细菌噬菌体DNA把病毒遗传信息从亲本带给后代,有助于在概念上完成一种十分重大的革命。
已知遗传物质是具有非常特异的化学结构的DNA,而不是特异性很差的核蛋白,用分子结构来阐明遗传信息的前景变得更为明朗。有许多证据说明DNA是一种磷酸脱氧核糖在一个带有3′—5′键的多核苷酸链上,有规则地重复的多聚物。Chargaff发现了一个重要的规律:尽管沿着多核苷酸链上的碱基顺序是复杂的,不同的DNA,其碱基组成显著不同,但腺嘌呤和胞嘧啶的数目总是相等。鸟嘌呤和胞嘧啶的数目也总是相等。DNA在电子显微镜里看起来像一根均匀一致的不分枝的细线,其直径为20埃。Signer、Caspersson和Hammursten用流动双折射测定表明,DNA中碱基的平面同线状分子的长度大致成直角。它们用紫外二色性测定得到相同的结果,并看出精子头部DNA碱基明显的平行性。早些时候,Schmidt和Pattri在光学方面研究了精子头部遗传物质,发现遗传物质非常有次序。Astbury作了先驱性的DNA纤维X射线衍射的研究,并找到DNA非常规律的证据;他把强烈的3.4埃,反射正确地解释为由于平面的碱基彼此连在一起的结果。Gulland和Jordan在电位滴定分析方面的研究指出,碱基是由氢把它连接起来,而Gulland的确设想到,多核苷酸链或可由这些氢链联结成多链胶束。
这样,对一个纯粹的化学物质作出具有深刻含义的生物活性的结论,与物质本性的许多边缘知识的显著增长是相符的。与此同时,我们开始从DNA获得详尽的X射线衍射资料。这是能提供恰当地说明分子立体构型的唯一的资料。
把DNA X射线衍射同建立分子模型结合起来
一旦从DNA纤维得到了良好的衍射图像,它就引起人们巨大的兴趣。在我们的实验室里,Alex Stokes提出了一个从螺旋DNA衍射的理论。Rosalind Franklin(她经历了一生的顶峰以后,于数年前死去)对X射线分析作出了非常有价值的贡献。在剑桥,研究生物大分子结构的医学研究会实验室,我的朋友Francis Crick和Jim Watson对DNA结构深感兴趣。Watson是一位到剑桥来研究分子结构的生物学家。他从事于细菌噬菌体繁殖的工作,并敏锐地领悟到有巨大可能性通过发现DNA的分子结构,或可揭示这一问题。Crick正从事于螺旋蛋白质结构的工作,并对什么东西控制蛋白质的合成感兴趣。Pauling和Covey通过他们发现蛋白质α-螺旋指出,在它自己的活动中建立精确的分子模型,是一个有力的分析工具。来自DNA的X射线资料不是那么完整,以致只能在没有立体化学的有力帮助下推导DNA结构的详细情景。显然,DNA的X射线的研究需要建立确切的分子模型来作为补充。在我们的实验室里,我们集中精力在扩大X射线资料,Watson和Crick则在剑桥建立分子模型。
DNA分子规则性的反论
DNA X射线衍射图像是如此清晰,表明DNA分子是极有规则的,它是如此有规则,以致可以结晶。图像的形状清楚地表明分子是螺旋的,分子线里面的多核苷酸链有规则地缠绕。但已经知道,不同大小的嘌呤和嘧啶沿着多核苷酸链排列成不规则的顺序。为什么这样一种不规则的排列能得到一种高度规则的结构?这一反论提供了解决DNA结构问题的线索,而Watson和Crick的结构假说解决了问题。
DNA分子的螺旋结构
Watson和Crick发现了DNA分子结构的要害,即:如果DNA里面的碱基由氢键成对地联合,腺嘌呤和胸腺嘧啶配对以及鸟嘌呤和胞嘧啶配对的总的大小是相同的。就是说,尽管碱基对的顺序是不规则的,但含有这些碱基对的DNA分子却是高度规则的。这些碱基对见图2。连接碱基同脱氧核糖的键之间的距离对两种碱基对来说,的确是相同的(大致在0.1埃以内),这些键同连接脱氧核糖的C1 原子的线所呈的角度也的确是相同的(大约在1°以内)(见图2)。其结果是,如果两个多核苷酸链由碱基对相连,对两种碱基对来说,两个链之间的距离是相同的,并由于键同C1 —C1 线之间的角度对所有碱基都是相同的,分子的脱氧核糖和磷酸部分的几何学的确能成为规则的。

图2 Watson-Crick碱基对(由S. Arnott修订)
上面:氢键连接鸟嘌呤与胞嘧啶。下面:氢键连接腺嘌呤与胸腺嘧啶。两种碱基对C1 —N3 和C1 —N9 键之间的距离为10.7埃,所有这些键同C1 —C1 线成52°角。(图中Å即埃)
Watson和Crick建立这种二链分子模型,链呈螺旋,其主体如X射线资料所示。在模型中,一个多核苷酸链同另一个多核苷酸链相互盘绕,一个链上的原子排列同另一个链上的原子排列方向相反。由于这一结果,如果从上向下转动,这两个链是相同的,且分子的每一个核苷酸都具有相同的结构,其所处环境也相同。唯一不规则的是碱基的排列。一个链上的排列能毫无限制地变化,但碱基配对要求一个链上的腺嘌呤同另一个链上的胸腺嘧啶相连接,鸟嘌呤同胞嘧啶相连接。因此,一个链上的排列由另一个链上的排列决定,就是说,彼此是互补的。
B型DNA分子的结构见图3。碱基彼此堆积,其间相距3.4埃,其平面几乎同螺旋轴垂直.碱基扁平的一边不能同水分子结合,因此,把DNA放在水中,其碱基相互吸引。这种疏水结合,以及碱基的氢键结合,使DNA结构保持稳定。
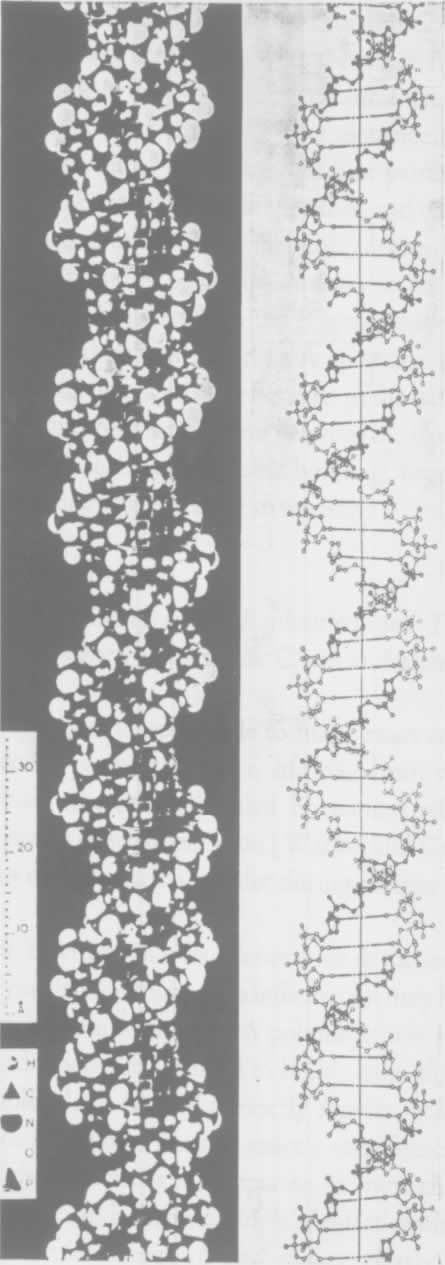
图3
左边:B构型DNA的分子模型原子大小相当于Von der Waals直径。
右边:相当于模型的图解可清楚地看到由氢键连接碱基的两个多核苷酸链。
Watson-Crick关于DNA复制和信息从一个多核苷酸转移到另一个多核苷酸的假说
对遗传物质来说,重要的是能够精确无误地复制自身;否则生长就会失调,生命不能发生,有益的类型不能通过自然选择保存。碱基配对提供了自我复制的手段(Watson和Crick)。看来它也是蛋白质合成不同时期信息转移的基础。
遗传信息用4个字母编码在沿着多核苷酸链上的4个碱基的顺序之中。这个信息可以从一个多核苷酸链转移到另一个多核苷酸链。一个多核苷酸链起一个模板作用,核苷酸在模板上排列,从而建成一条新链。倘若所形成的两链分子完全有规则,碱基配对就保证新链的顺序对亲本链的顺序是精确无误地互补的。如果这两个链接着分开,新链能起模板作用,又形成一个新链;它同原始的链是相同的。大多数DNA分子有两个链,显然复制过程能重新制造这样一个分子。它也能把信息从一个DNA链转移到另一个RNA链(信使RNA的合成就是如此)。
碱基配对也使一条多核苷酸链同另一条多核苷酸链的互补顺序之间进行特异性的连接成为可能。这种特异的相互作用或许是氨基酸连接一个多核苷酸链的必不可缺部分的一种方法,一个蛋白质的氨基酸顺序就在这上面编码。在这种情况下,氨基酸连接一个转移RNA分子,而这个RNA某部分多核苷酸链同编码链配对。
自从1953年Watson和Crick首次介绍了碱基对以后,已提供了许多关于嘌呤和嘧啶大小和氢键长度的新资料。碱基配对最新图解(归功于S.Arnott)见图2。现在我们对C1 原子间的距离采用10.7埃,以代替原先采用的11.0埃值,主要原因是有关N—H…N的新资料,证明这段距离在环状氮原子之间比不呈环状的原子间要短些。碱基对的氢键直线性极好,而键的长度同晶体中发现的一样(它们的长度变动约0.04埃)。
碱基对惊人的精确性反映了DNA复制的精确无误。然而,人们怀疑这种精确性为什么如此之大,因为破坏碱基对致使其完整性明显降低,所需要的能量大概不会大于一个量子的热能。或许可以解释其精确性。复制是一种包括许多碱基对的协作的现象,在任何情况下都必须强调,碱基配对的特异性取决于碱基同脱氧核糖连接起来的键彼此间的关系得到正确安排。这种安排可能是由DNA多聚酶决定的。不管这过程的机理如何,双螺旋的每一个核苷酸的几何学和周围环境的正确安排,必然导致精确的复制。如果质子的互变异构变化牵涉到氢的结合或碱基的化学改变,就会产生复制过程的错误。这些错误相当于突变。
DNA螺旋结构的普遍性和稳定性
当我们进行了初步的X射线研究以后,我的朋友Leonard Hamilton给我送来了人的DNA,这是他同Ralph Barclay从一个患有慢性骨髓性白血病病人的白细胞中提取出来的。他正在研究人的核酸代谢同癌的关系,并制备DNA以便比较正常人白细胞和白血病患者白细胞的DNA。此DNA的X射线图像很好。这样就开始了一项经过多年才告结束的协作,我们在协作中使用了Hamilton的DNA,以许多种盐的形式来确立双螺旋结构的可靠性。Hamilton从各种各样的生物和不同的组织制备DNA。这就得以证明精子和细菌噬菌体的惰性遗传物质和正缓慢或快速分裂、或正分泌蛋白质的细胞都有双螺旋的DNA(Hamilton等)。正常组织和癌组织的DNA之间,或由我的同事Geoffery Brown把小牛胸腺DNA分离成不同碱基成分的碎片,都没有发现DNA结构的差异。
同Harriet Ephrussi-Taylor协作,我们还进行肺炎球菌有转化活性的转化因素的研究,并观察转化DNA的结构。至今发现对双螺旋DNA唯一的例外是一些很小的细菌噬菌体,它们的DNA是单股。但是,我们早就发现那些腺嘌呤含量特别高的DNA、或有葡萄糖和羟甲基胞嘧啶连接的DNA,其结晶不同。
DNA结构决非假象
看来单从DNA作X射线衍射研究尚嫌不足。显然,人们将试图探讨完整细胞的遗传物质。分离出来的DNA结构同活体条件下多数与蛋白质结合在一起的DNA结构或许有所不同,这一点是可能的。光学研究表明精子头部分子有着明显的次序,因而它们是X射线研究的良好对象,而绝大多数细胞的染色体却是复杂的对象,很少看到有秩序的结构。Randall几年来一直对这一问题感兴趣并开始了Gosling关于公羊精子的研究。看来由Schmidt发现的、在光学上具有高度各向异性的棒状头足纲精子是X射线研究的极好材料。Rinne在研究自然界许多分支的液体结晶时,早就对这种精子作过衍射照相;但大概他的技术不大恰当,致使他得出核蛋白是液体结晶的错误结论。我们的X射线照片(Wilkins和Randall)清楚地证明精子头部的物质是立体的,即它是结晶体而非液体结晶。衍射图像(见图4)同DNA的情况(见图5)很相像,这说明提纯的DNA的纤维结构基本上不是假象。我在那不勒斯动物站工作时,发现有可能给精子头部的纤维定位。一束束自然定向的精子,其完整、湿润的精球产生了良好的衍射图像。Watson给我的T2细菌噬菌体也得到了类似的DNA的图像。

图4 头足纲精子X射线衍射图像
精子头部的DNA分子有其纵轴。多核苷酸间相距3.4埃,符合图像顶部和底部的强烈衍射。图像中央部分的明显反射说明分子是晶体排列。

图5 DNA纤维(B构型)在高湿件下的X射线衍射照片
纤维是垂直的。3.4埃反射在顶部和底部。中央区域反射产生明显的X型的角度,相等于螺旋分子多核苷酸链斜度的恒定角度(H.R.Wilson摄影,Hamilton提供DNA)。
DNA的X射线衍射图像及分子的各种构型
X射线衍射是能得到有关DNA分子构型非常详尽知识唯一的一项技术。光学技术尽管对补充X射线分析有价值,但它提供的资料,主要在键和基团定位方面,其局限性大得多。X射线资料分两个阶段对推导出DNA结构作出了贡献。第一个阶段,提供有助于确立Watson-Crick模式的资料;第二个阶段,证明Watson-Crick的设想在其主要方面是正确的,设想包括模式的重新调整和改进。
X射线研究(如Langridge等、Wilkins)证明DNA分子的突出之处,在于它们有大量的不同构象,其中绝大多数能有几种晶体形状。决定分子构象和晶体形状的主要因素是纤维的水含量和盐含量以及用于中和磷酸基的阳离子(见表1)。
表1 各种DNA纤维的小结
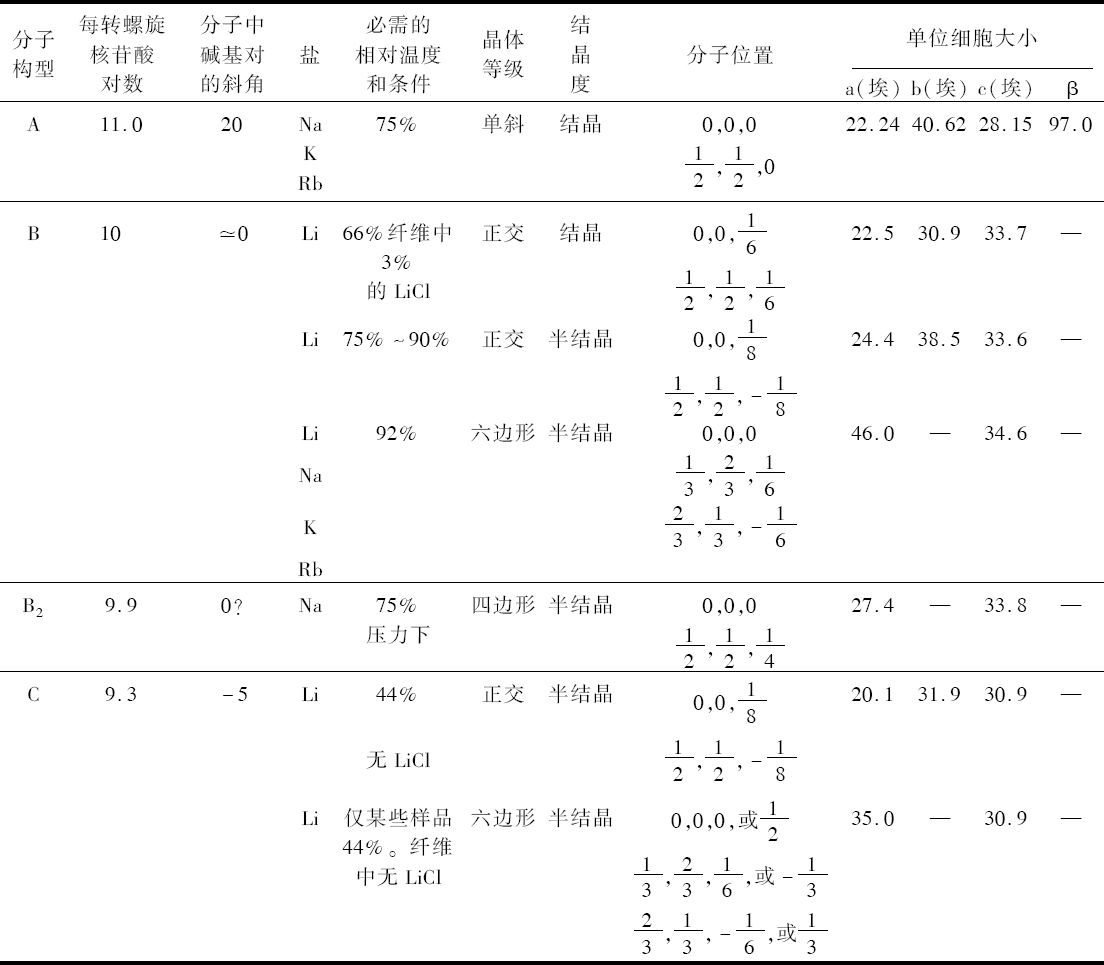
我将扼要地谈一下DNA的主要构型。在所有情况下,都能用同样基本的Watson-Crick结构来令人满意地说明衍射资料。这比单研究一种构型更为令人信服地证实这种结构的正确性。基本做法是调整分子模型,直至从模型计算的衍射强度同所观察的结果相符(Langridge等)。
就大多数X射线资料而言,它只提供DNA衍射束的强度,而不提供衍射束的相。因此不能直接得出它的结构。如若X射线资料的解答足以把一个结构中的绝大多数原子分开,则除非假定这个结构是由已知其平均大小的原子组成,否则就只能在不用立体化学设想的情况下推导其结构。然而,单用X射线不能对DNA的大多数原子一一定位(见图7)。所以,作了更广泛的立体化学设想,从这些设想得以建立分子模型。这些设想多数都无可选择,但只有一种选择,即:氢键在一对碱基上的排列,必须用X射线资料来确立这种设想的可靠性。换言之,必须确立所提出的这个结构是独一无二的。近年来我们的多数工作是这种性质。可以有理由肯定DNA结构是正确的,X射线资料必须尽可能广泛地搜集。
B构型
图5说明当分子被水分开,并在很大程度上彼此独立行为时,DNA的一根纤维在高湿度下的衍射图像。我们没有在这些条件下深入研究DNA。这种图像可以改进,但它们有理由很好地规定下来,而它们的许多特征非常清楚地说明分子有一规则的结构。已知构型为B(见图3),这是在活体看到的,并且证明,当DNA溶于水的时候,也是如此。每一圈螺旋有10对核苷酸。为什么这个数字必须是整数,这一点无明显的结构上原因,如确实如此,它的重要性至今尚不清楚。
当DNA结晶时,结晶过程使分子受到限制,并能使它特别有规则。还有,纤维微晶体中分子的周期性排列使衍射图像分裂成相当于各晶体面的强烈反射(见图6)。仔细测定反射位置和推导晶体的晶格使得在立体上鉴定反射的方向成为可能。大多数纤维物质的衍射图像同图5相似,其衍射数据是平面的。相反,DNA结晶纤维得到相当完全的立体数据。这些数据得出从所有角度观察这个分子样子的资料,并可同从单晶体得到的资料相比较。可以使用诸如立体傅立叶合成(见图7)这些技术,并使结构测定有理由予以信赖。
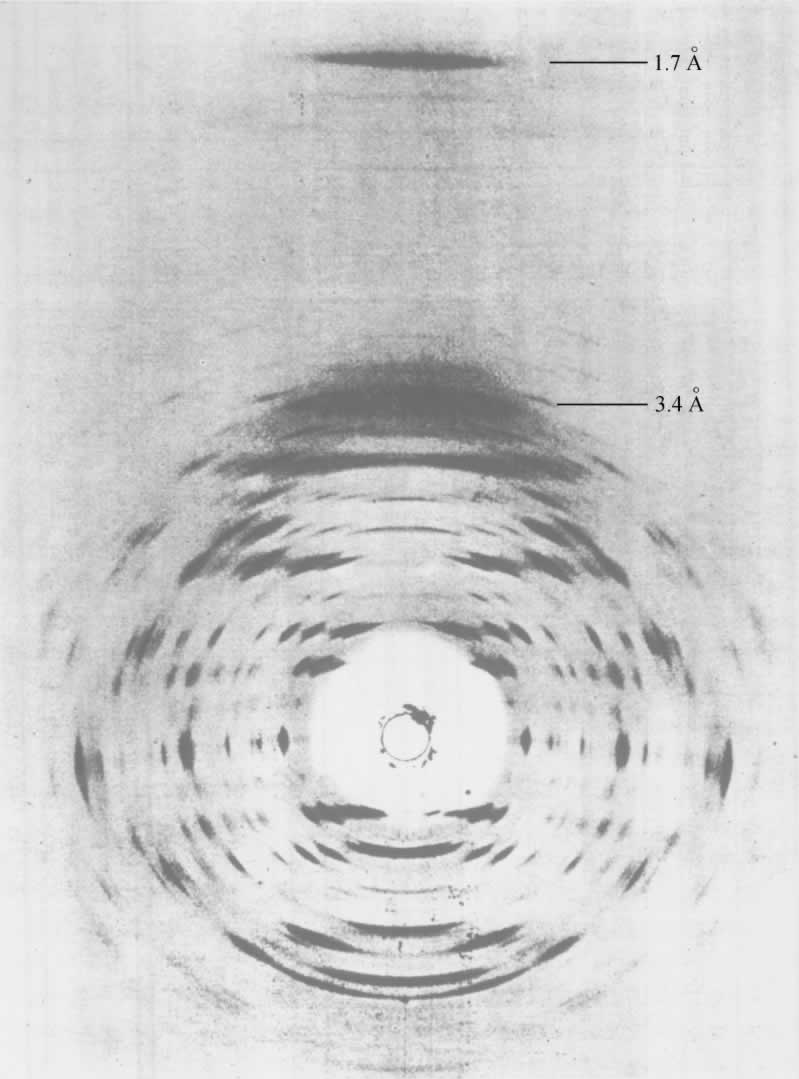
图6 DNA微结晶纤维的X射线图像
总强度的分布与图4相似,但由于分子在晶体中的规则排列,衍射分裂成明显的反射。明显反射扩及小至1.7埃的位置(N.Chard摄影,L. D. Hamilton提供DNA)(图中Å即埃)。
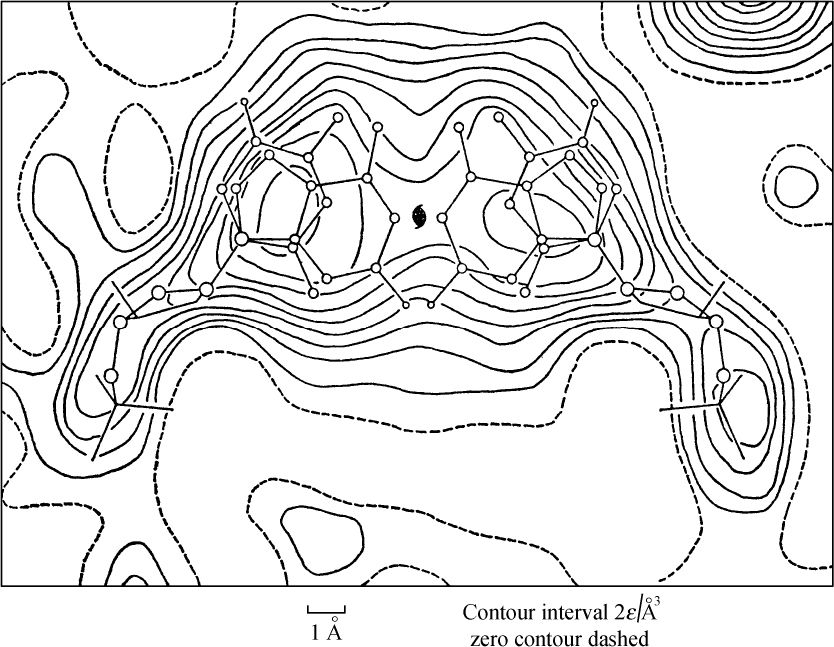
图7 傅立叶合成图
表示DNA B构型里面一个碱基对平面上电子密度的分布。分布相当于一个平均碱基对。图中看出碱基对的形状,但没有解决一个碱基对里面的各个原子(正在修订傅立叶合成,因而此图尚待改进)(由S. Arnott提供)。(图中Å即埃)。
A构型
在这个构象中分子的每圈螺旋有11对核苷酸,螺旋间距为28埃。碱基和核苷酸的脱氧核糖及磷酸部分的相对位置和定位同B构型的情况显然不同,特别是碱基对要从对螺旋轴垂直的角度倾斜20°(见图8)。
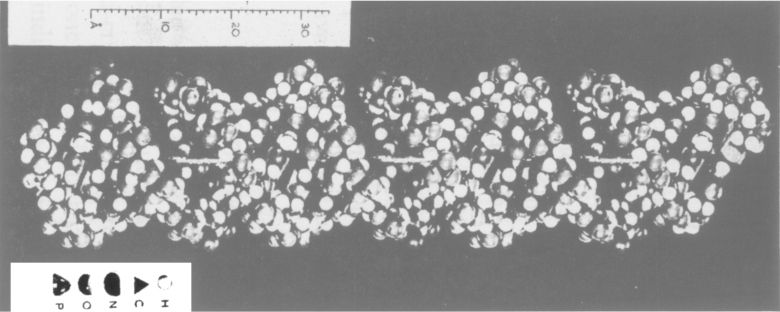
图8 A构型的DNA分子模型
(可以看到碱基对同水平面倾斜20°。)
A构型DNA(见图1)是观察到的第一个结晶类型。虽然它不是在活体中看到的,但因为螺旋RNA的构型同它非常相似,而显得特别有意义。不久将提供A构型DNA的充分报道,图9为A构型DNA图像的一张好照片。

图9 A构型DNA微结晶纤维的X射线衍射图像
(H. R. Wilson摄影,L. D. Hamilton提供DNA。)
C构型
可以把这种构型看做是由于部分干燥而形成的一个假象。螺旋为非整数,每一圈约有 对核苷酸。螺旋叠在一起形成一个半结晶结构,一个分子里面这一个核苷酸同另一个核苷酸的位置之间没有什么特别的关系。个别核苷酸的构象则同B型极相似。B和C衍射图像间的差别可用核苷酸在螺旋上的不同位置来说明。类型间比较更肯定了结构的正确无误。在某种程度上,存在的问题好像是正试图通过观察其影子来推导折叠位置的结构:如果位置的构象有一点改变,它的结构就更清楚。
对核苷酸。螺旋叠在一起形成一个半结晶结构,一个分子里面这一个核苷酸同另一个核苷酸的位置之间没有什么特别的关系。个别核苷酸的构象则同B型极相似。B和C衍射图像间的差别可用核苷酸在螺旋上的不同位置来说明。类型间比较更肯定了结构的正确无误。在某种程度上,存在的问题好像是正试图通过观察其影子来推导折叠位置的结构:如果位置的构象有一点改变,它的结构就更清楚。
RNA分子的螺旋结构
与DNA相反,RNA得到的衍射图像不好,尽管包括我们自己在内的各研究工作者为此付出了很大的努力。有许多例子表明RNA具有螺旋区,即RNA溶液的光学特性使人强烈地想到(如Doty),RNA分子的各部分像DNA那样,碱基彼此连接,结构是螺旋的;以及合成的多核糖核苷酸的X射线研究使人设想RNA像DNA(Rich)。RNA衍射图像(Rich和Watson)总的方面同DNA相像,但因为定位紊乱和漫射而不能清楚地辨别其图像的性质。重大的困难是在3.3埃和4埃处出现强烈的子午线反射。用一个螺旋结构不可能对此进行解释。
在早期的工作中,许多RNA制剂都很不纯。我们想更纯的植物病毒RNA可能得到较好的图像,但结果并非如此。然而,当制备核糖体RNA和可溶性RNA成为可行时,我们感到结构分析的前景就好得多了。我们决定集中研究可溶性RNA,这在很大程度上是由于我们实验室里的Geoffrey Brown,正在制备大量可溶性RNA中一种高度纯净的转移RNA,用于他在物理和化学方面的研究,还因为他正在把它分级分离成专门用于把特殊氨基酸掺入蛋白质的各种转移RNA。有另一些理由使这种RNA引人注目:对一个核酸来说,这个分子小得出众,有证据表明它或许有规则的结构,它的生化作用是重要的,并在许多方面它的功能是了解的。
我们发现要在纤维中找到转移RNA十分困难,然而借助于一台解剖显微镜,在大气干燥的情况下,小心地层开RNA凝胶,我发现能制出同DNA一样高的具双折射的纤维。但这种纤维得到的图像并不比用其他种RNA得到的图像好些,以及当提高纤维的含水量时,分子就迷失方向。Watson, Fuller, Michael, Spencer和我本人成年累月地工作,想制出更好的标本供X射线研究用。我们的进展不大,直到Spencer发现了一种标本,除通常漫射的RNA图像外,还得到一些微弱但明显的衍射环。这种标本是一种被密封在一个小室内作X射线研究的RNA凝胶,并且他发现凝胶由于漏气而慢慢变干。衍射环是如此明显,使我们几乎可以肯定,它们是由于结晶不纯造成的非真实的衍射,这在生化制剂X射线研究中是常见的。一个RNA样品由于混有DNA而得到了和DNA非常相似的环。因此我们对环就不抱很大希望了。但数周后Spencer排除了所有其他的可能性,看来很明显,环是RNA本身形成的。他通过控制慢慢地干燥得到了更加明显的环,以及Fuller用我们改良过的方法展开RNA,并采用Brown使凝胶慢慢浓缩的方法,使RNA定位而不破坏它的结晶性。这些纤维得到清清楚楚的衍射图像,并且当纤维成水合物时仍然排列整齐。看来我在早些时候使用过的把纤维尽最大可能拉长的办法,破坏了结晶性。如果不是这样做,就可以先使材料慢慢结晶,再把它们拉长以得到排列整齐的微晶体和RNA分子。单分子要得到良好的排列显得太小了,除非通过结晶把它们聚集起来。很出乎意料的是在我们试验过的所有不同类型的RNA中,分子量最小的转移RNA排列得最整齐。
转移RNA的衍射图像十分清楚,排列整齐良好(Spencer、Fuller、Wilkens和Brown)。这些改进揭示了RNA和A构型DNA图像惊人地相像(见图10)。于3.3埃和4埃处的两个反射难题得到了解决(见图11),在RNA图像中反射到三层线上的位置同DNA的情况不同;其结果是,如图像定向不好,3个反射就重叠,从而给人产生2个反射的印象。无疑地RNA有一与A构型DNA几乎相同的规则的螺旋结构。RNA与DNA图像间的差别只是这两个结构间的小异而已。
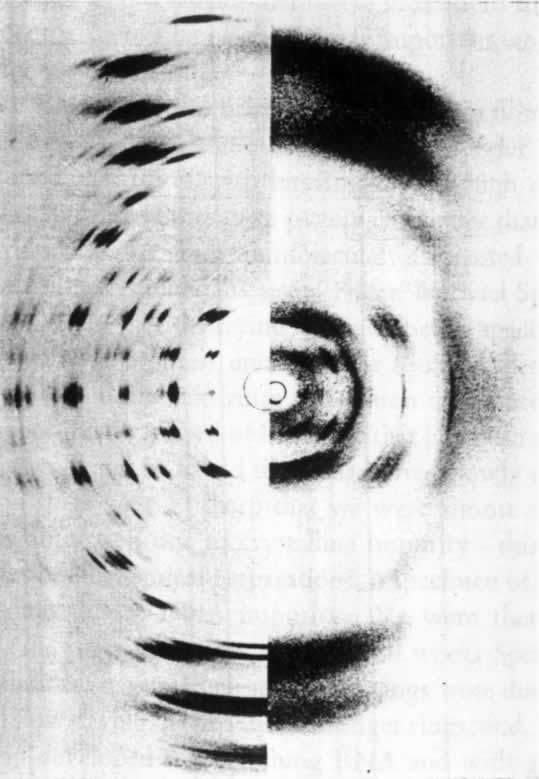
图10 A构型DNA纤维(左)和转移RNA纤维(右)X射线衍射图像的比较
这两种图像的强度总分布很相似,但因为在这两种情况下晶体里的分子装配不同,其明显的结晶反射的位置就不同(W. Fuller和M.Spencer摄影,G. L. Brown提供RNA)。
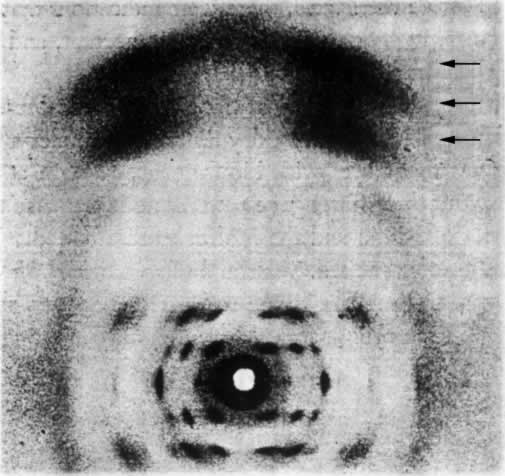
图11 转移RNA的衍射图像
表明在3.3埃和4埃区衍射分解成用箭头表示的三层线,并与A构型DNA图像相符(Fuller和M. Spencer摄影,G. L. Brown提供DNA)。
RNA结构与DNA结构相像的一个重要原因,是RNA必须具有在很大程度上或全部互补的碱基顺序。分子的核苷酸数目大约为80个。与X射线结果相符的最简单的结构是一个单个的多核苷酸链自己反过来折叠,通过碱基配对,链的一半同另一半相连。这种结构见图12。必须强调指出,当我们肯定螺旋结构为正确时,却并不知道链的两端是否在分子的末端。这个链可以在分子的两端同有些地方沿着螺旋的链端折叠。已知氨基酸连接到以胞嘧啶-胞嘧啶-腺嘌呤结尾的链末端上。
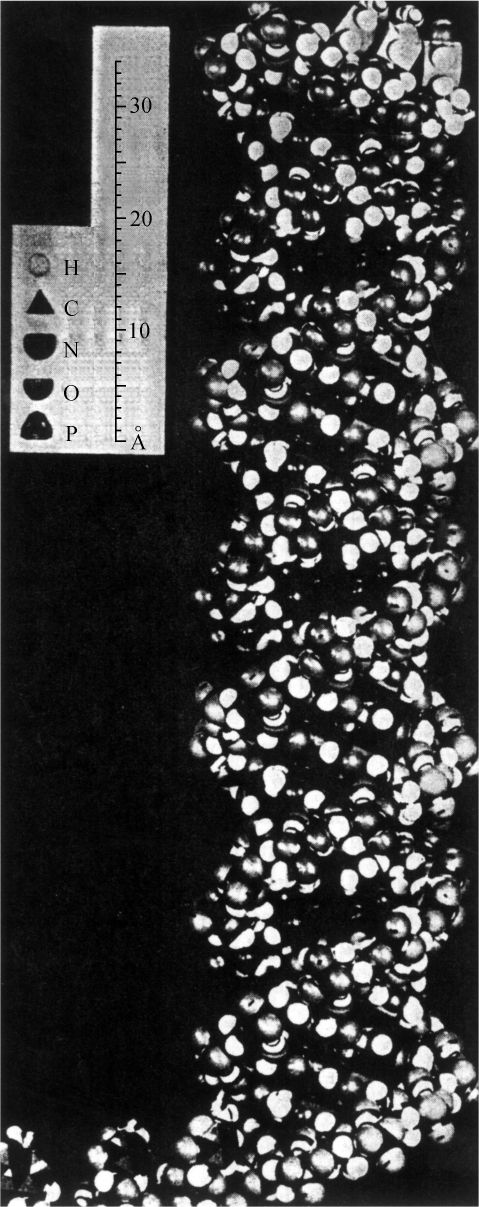
图12 转移RNA分子的分子模型和图解
RNA分子结构与功能的关系
分子模型的建立说明在一个转移RNA分子末端形成折叠的核苷酸数必须是3个或3个以上。在我们的模型中,折叠由3个核苷酸组成,这3个核苷酸各具有1个不配对的碱基。或许是这样一种情况:碱基三联体是分子同密码RNA多核苷酸链必要部分相连接的部分。而所连接的那个密码多核苷酸链的一定部分,又决定氨基酸在一个蛋白质多肽链上的顺序。相信一个密码RNA的三联体相当于一个氨基酸,转移RNA的三联体则通过氢键和碱基对的形成特异地把它自己同密码三联体相连接。但必须强调指出,这些资料是推测出来的。
我们假设,转移RNA分子的那一部分同连接氨基酸与RNA的酶起特异的相互作用;但我们不清楚它是怎样发生的。同样,对于与DNA复制有关的酶同DNA相互作用的途径或DNA复制的情景,我们都所知甚微。在转移RNA分子中存在互补碱基顺序,使人设想它或许像DNA那样自我复制;但至今支持这种意见的证据不足。病毒和核糖体RNA的衍射图像证明这些分子也具有螺旋区;其功能也不肯定。
就DNA来说,发现其分子结构立刻得出复制的假说。这要归功于DNA结构的简单性。在多数情况下,分子结构和功能的直接关系似乎不多。得出RNA分子的螺旋构型对阐明RNA功能向前迈了一步;但是在形成有关RNA功能的充分描述前,为了确定碱基顺序,以及更多地了解各种RNA如何在核糖体中相互作用,可能需要更完整的结构资料。
用X射线衍射分析确定转移RNA碱基顺序的可能性
核酸的生物特异性看来完全是由它们的碱基顺序决定的,对这些顺序的测定很可能是当代核酸研究的最基本的问题。要使X射线衍射进行碱基顺序测定行之有效,一个DNA分子的碱基数目就太多了,然而,转移RNA的碱基数却不是那么多。有两次观察说明,用X射线对转移RNA进行完全的结构分析是可能的。我们在转移RNA X射线图像中看到分开的点(见图13),每个点相当于一个RNA单晶体。我们估计其大小约为10微米,通过用偏光显微镜观察看到很可能是晶体的双折射区,证明这是对的。要使晶体增大几倍而足以作单晶体X射线分析无多大困难。
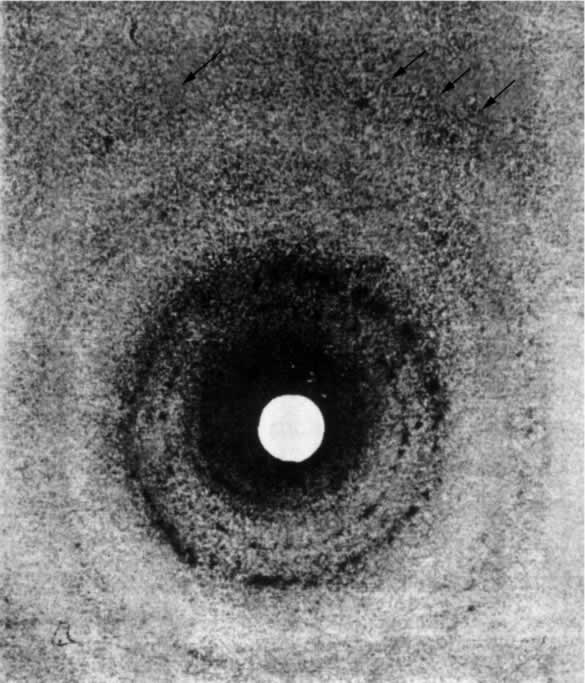
图13 排列不整齐的转移RNA的衍射图像
看到具有相当于从RNA单晶体反射的点的衍射环,箭头指向从平面反射到约6埃的距离。
第二个鼓舞人心的观察是从DNA得到的X射线资料,几乎完全限于解决不定向的DNA纤维晶体。DNA的强度资料说明DNA的温度因子(B=4埃)同简单的化合物相同。这就看出,DNA晶体具有很完整的结晶性,而一旦能得到DNA的单晶性,强度资料必能适用于精确测定DNA所有的原子位置(非周期性碱基顺序除外)。
我们正在研究获得DNA单晶体的可能性,但更值得注意的问题是,获得在晶体完整性方面同DNA相等的转移RNA的单晶体,从而可以分析碱基顺序。目前,RNA晶体的完整性要比DNA差得多。但是,我们大多数实验所用的RNA是对不同氨基酸特异的RNA混合物。我们很少使用只对一种氨基酸十分特异的RNA。我们希望或许能得到这种只有一种分子的RNA的良好制剂。我们可以指望这种RNA形成的结晶像DNA那样的完整。如能做到这一点,直接分析分子的全部结构,包括碱基顺序和在螺旋端的折叠,将毫无障碍。我们或许过于乐观,但X射线衍射分析在核酸和蛋白质领域内新的和有些出乎意料的成功使我们有理由持乐观态度。
参考文献
[1]Caspersson, T., 1941. Naturwiss.,29 , 33.
[2]Brachet, J., 1942. Arch. Biol. Liege,53 , 207.
[3]Avery, O. T., MacLeod, C. M., and MeCarty, M., 1944. J. Exp. Med.,79 , 137.
[4]Signer, R., and Schwander, H., 1949. Helv. Chim. Acts.,32 , 853.
[5]Hershey, A. D. and Chase, M, 1952. J. Gen. Physiol.,36 , 39.
[6]Chargaff, E., 1950. Experientia,6 , 201.
[7]Signer, R., Caspersson, T., and Hammarsten, E., 1938. Nature,141 , 122.
[8]Schmidt, W. J., 1937. Die Doppelbrechung von Karyoplasma, Zytoplasma, und Metaplasma, Borntraceger, Berlin.
[9]Pattri, H. O. E., 1932. Z. Zellforsch. Mikroskop. Anat.,16 , 723.
[10]Astbury, W. T., 1947. Symp. Soc. Exptl. Biol., I. Nucleic Acid, Cambridge Univ. Press, p. 66.
[11]Gulland, J. M. and Jordan, D. O., 1947. Symp. Soc. Exptl. Biol., I. Nucleic Acid, Cambridge Univ. Press.
[12]Gulland, J. M., 1947. Cold Spring Harbor Symp. Quant. Biol.,12 , 95.
[13]Watson. J. D. and Crick, F. H. C., 1953a. Nature,171 , 737.
[14]Watson. J. D. and Crick, F. H. C., 1953b. Nature,171 , 964.
[15]Hoogsteen, K., 1959. Acta Cryst.,12 , 822.
[16]Hamilton, L. D., Barclay, R. K., Wilkins, J. H. F., Brown, G. L., Wilson, H. R., Marvin, D. A., Ephrussi-Taylor, H. and Simmons, N. S., 1959. J. Biophys. Biochem. Cytol.,5 , 397.
[17]Rinne, F., 1933. Trans. Faraday Soc.,29 , 1016.
[18]Wilkins, M. H. F. and Randall, J. T., 1953. Biochim. Biophys. Act.,10 , 192.
[19]Langridge, R., Wilson, H. R., Hooper, C. W., Wilkins, M. H. F. and Hamilton, L. D., 1960. J. mol. Biol.,2 , 19.
[20]Wilkins. M. H. F., 1961. J. Chim. Phys.,58 , 891.
[21]Doty, P., 1961. Biochem. Soc. Symp., 21 , 8.
[22]Rich, A., 1959. in A Symposium onmolecular Biology (Ed. Zirkle), Univ. Chicago Press, 47.
[23]Rich, A. and Watson, J. D., 1954. Nature,173 , 995.
[24]Spencer, M., Fuller, W., Wilkins, M. H. F. and Brown, G. L., 1962. Nature,194 , 1014.
噬菌体、大肠杆菌素和大调节现象 【100】
萨尔瓦多·卢瑞亚(S. E. Luria) 【101】
(美国 麻省理工学院,1969年)
有关噬菌体生长、突变和重组方面的早期工作,有幸能为分子生物学发展到它目前这种令人满意的状况提供一条通路。今天,不必再细述这些早期工作的情况了,在这些早期工作中,我很幸运地与Max Delbrück和Alfred Hershey进行了友好且成功地协作。由此去研究系统发育的原因是一个更困难的问题。这个问题把早期的噬菌体工作引导到对病毒繁殖、基因复制、基因功能及其调节等方面的现代知识水平上来了。最使我满意的是我的学生和同事们在这些工作中起到了重要的作用,以及我本人也获得了与许多科学探险家们进行协作的经验。
噬菌体的研究已在很多方面形成分支,每一分支都为分子生物学大厦做出了部分贡献。最著名的一个分支是基因的功能及其调节。在这个领域中,噬菌体研究做出的主要贡献是由André Lwoff和F. Jacob进行的溶源性研究,这些工作导致F. Jacob和J. Monod提出了操纵子理论的模型。从这一理论考虑,调节现象关系到单个基因或基因组的功能。在本篇演讲中,我想讨论在细胞调节的某些方面与大调节现象有关的几种途径。这里所说的大调节现象,意思是功能上的改变能影响到活细胞的某些主要生命过程,如DNA、RNA、蛋白质的合成、能量的代谢、细胞膜的选择透性功能。
对青霉素、链霉素等抗菌素的研究,对以某种方式(克分子)作用于细胞过程的一些化学因素的研究,为阐明细菌细胞壁生物合成,或细菌蛋白质合成的机制等问题,都曾起到过重要的作用。当一种因素(如噬菌体),或某种别的大分子化学因素以单一颗粒的形式起作用,造成一种主要的细胞功能改变时,情况就更复杂化了。因为一些扩增机制必定介于各个单位因素和有关细胞受到影响的成分之间。对一种病毒因素来说,扩增机制可能就是病毒复制或其遗传潜能的表达。对一种蛋白质因素(如一种细菌素)来说,扩增机制必定是某些细胞结构或某些细胞控制系统功能整体性的一种改变。在这两种情况中,弄清这些因素对主要的细胞过程的作用模型,就可能揭示这个细胞机器在功能上的重要性。
在我的实验室中,我们通常是用噬菌体和细菌素来探索细菌细胞的大调节现象的。在这个领域中,除了遗传转录的调节研究外,其他方面还没有很大的进展。因此,即使是一种对目前工作的叙述,至少在说明“我们正在探求什么”这一点上还是有价值的。
噬菌体和大调节现象
噬菌体具有作为细胞功能控制者的潜在作用,早期工作就观察到:受了照射的噬菌体T2,在失去它的复制能力之后,仍保留其杀死寄主和进行干扰的能力[1 ] ,细菌不是全部破碎而是死亡。几年后,对噬菌体在引起致死作用前形成侵染作用的生化途径,可以根据生理学机制(即在大分子合成水平上的特异性抑制)来解释。现在我们知道,某些烈性噬菌体,包括T系偶数列大肠杆菌噬菌体,能造成它们寄主细胞的蛋白质、RNA和DNA合成的迅速停止。其他噬菌体在这些过程中造成的影响不那么猛烈,或较为短暂。但我们对这些抑制机制的了解进展得很慢,慢得出乎意料。
噬菌体侵染和寄主的DNA合成
让我们举例说明噬菌体侵染对寄主DNA的影响。T系偶数列噬菌体的情况看来应该是最简单的。这些噬菌体的DNA中没有胞嘧啶[2 ] ,只有羟甲基胞嘧啶(HMC),这些噬菌体DNA决定着一种酶——脱氧胞苷三磷酸酶——的产生,这种酶能破坏寄主DNA的一种特异性前体物,即dCTP(见Cohen的总结[3 ] )。细菌受到侵染后,其DNA很快被破坏,变成酶溶性的断片,最终变成单个的核苷酸。已知在细菌DNA中,双链的断裂就会停止细菌DNA的复制[4 ] ,但在导致这样的断裂中,噬菌体的作用还不清楚。噬菌体T4的某些突变体不能把寄主DNA转变成酸溶性产物[5 ] ,但还能发生初步的断裂,使寄主DNA断成大的断片。与这些断裂有关的核酸酶,对含有胞嘧啶的DNA是特异性的;尚未发现噬菌体哪个基因的突变能防止寄主DNA的断裂。
枯草杆菌 e噬菌体的情况更引人感兴趣,这在我们的实验室中,已由David Roscoe和Menashe Marcus研究过了。 e是一种在其DNA中含有羟甲基尿嘧啶(HMU),而不含有胸腺嘧啶的噬菌体,在侵染过程中,有一系列酶的改变,这些酶的变化使DNA合成途径从合成细菌类型的DNA转变到合成噬菌体类型的DNA[6 ] 。这些酶是一种dUMP羟甲基化酶;一种胸苷三磷酸核苷酸水解酶(dTTPase);一种胸苷酸合成酶抑制物;一种dTMP核苷酸酶;一种dCMP脱氨基酶;可能还有一种脱氧核苷酸激酶。 e噬菌体侵染后几分钟寄主DNA合成就停止了。Roscoe[7 ] 曾经指出,寄主的DNA仍是完整的,或者至少双链的断裂不会发生到在数目上可以测出来的程度。在有胸腺嘧啶存在的条件下,采用胸腺嘧啶缺陷的寄主细胞,或者采用dTTPase缺陷的突变噬菌体,就可以避免酶与dTTP合成之间的这种干扰作用。在这些条件下,产生的噬菌体DNA中至少10%(也可能是20%)的HMU被胸腺嘧啶代替了;但寄主DNA的合成仍被停止。因此我们认为,必定还有某种和这种停止有关的更特异的机制存在。这种机制可能不是因为HMU核酸酶抑制了寄主DNA合成酶,因为有一种噬菌体的突变体,它在决定dUMP羟甲基化酶或dTTPase能力上都是缺陷的,但它也能使细菌的DNA合成停止。此外,噬菌体侵染后,寄主DNA合成的停止需要蛋白质合成;由此看来,噬菌体有一些特异的、能抑制细菌DNA合成的功能。目前,我们正试图证实这种噬菌体功能,它可能是在自我复制的水平上,或在一些尚未认识的调节水平上发挥作用。
寄主蛋白质和RNA
现在我们来讨论噬菌体侵染对RNA和蛋白质合成的影响。至少在T系偶数列噬菌体中,寄主蛋白质合成的停止看来是mRNA合成停止的次级作用[8 ] 。然而,直接影响已有信使的翻译,也是可能的,无疑这些影响和某些动物病毒侵染的影响是一样的。
一年前,发现[9 ] 至少某些噬菌体(包括T系偶数列和T7大肠杆菌噬菌体[10 ] )引起RNA聚合酶的改变,使其特异性发生变化,在这一重大发现之前,对RNA合成停止的模型一直不清楚。噬菌体基因的极早期位点(侵染后立即转录的那些位点[11 ] )的转录需要一种聚合酶成分——σ因子,这种因子可能也是细菌基因的转录所需要的,它在噬菌体侵染后受到破坏或发生了改变。然后,由于一些新的因子的出现,使噬菌体其他基因得到转录,这些新的因子赋予寄主聚合酶稳定的核心酶以新的特异性[12 ] 。一种合乎情理的推测是:σ因子使聚合酶具有一种识别启动子的特异性,引起它在特异的DNA位置上起始mRNA的合成。
在这种情况下,大调节现象不是在某种纯调节机制的水平上引起的,而是在本身操纵机构的水平上引起的。噬菌体通过改变RNA聚合酶的特异性停止全部基因位点的表达。
哈佛大学R. Losick和我的学生A. L. Sonenshein一起证实了这种调节不是噬菌体侵染所特有的。Sonenshen和Roscoe[13 ] 观察到:枯草杆菌 e噬菌体,当它侵染处在孢子形成过程中的细菌时,它不能生长,也不能表达其功能。据此,Losick和Sonenshein推断:孢子形成和很多蛋白质合成的停止以及一些新蛋白质的出现是有关的,临界步骤可能是在RNA聚合酶特异性上有变化,这和大肠杆菌经T系偶数列噬菌体侵染后见到的情况类似[9 ] 。事实上他们已成功地证实了[14 ] ,有一种类似于σ的因子,是营养细菌细胞的RNA聚合酶的一部分,它在孢子形成过程中发生了改变,或被除去了,这样导致该细菌聚合酶的模板特异性有所改变。更明显的是在试管中,把大肠杆菌的σ因子加入这种枯草杆菌聚合酶的核心酶中,便可恢复它原来的活性!
值得注意的是在对孢子形成的这种研究中,使用噬菌体不是研究它在细胞中引起的变化,而是作为一种探索手段,去揭示和一种细菌的细胞周期中主要分化有关的一种调节现象——从营养生长到孢子形成在合成上的变化。RNA聚合酶变化与更普遍的大调节变化之间的关系,为解决高等生物的分化问题,提出了一种有意义的假说[12 ,15 ] ,它可能为研究细胞分化问题提供新途径。
大调节和大肠杆菌素
下面我将讨论另一个大调节的途径,为了能进一步搞清细菌细胞的功能机构,近来我们已转向研究这一途径。这涉及某些大肠杆菌素作用模型的研究。虽然大肠杆菌素研究的历史与噬菌体研究的历史是紧紧交织在一起的,但是详细说明这条迂回的道路是很有益处的,目前,我对大肠杆菌素的兴趣就在这方面。Iseki和Sakai[16 ] 发现的由温和噬菌体引起的沙门氏杆菌体细胞抗原转变,也来源于对噬菌体的研究。1956年,Hisao Ueteke博士到我的实验室来,和我一起研究了由ε噬菌体引起的从抗原10到抗原15的转变[15 ,17 ] 。1960年,Takahiro Uchida博士从Uetake的实验室来到麻省理工学院(M. I. T,)参加我的工作,使这种协作又继续了下去。幸运的是抗原转变的问题引起我的同事Phillips Robbins博士的注意。Robbins和他的同事是如何在生化水平上解决这个问题,以及在这个过程中是如何发现和阐明在多糖合成中类脂载体的作用[18 ] ,这些问题不需要在这里详细叙述了。然而,因这个工作的关系,引起了我对膜的兴趣,特别是对细菌细胞质膜的某些显著特性的兴趣。
在细菌细胞中,细胞膜是仅有的细胞器。它含有酶和一些别的组分,它不仅在渗透和主动运输中起作用,而且在细菌细胞壁的大分子成分[如肽聚糖和其他多糖(包括肠道细菌的脂多糖)]的生物合成中也起作用。另外,细胞质膜是末端呼吸机构的场所,在DNA复制过程以及在细胞分裂时,细胞质膜对DNA拷贝的分离也可能起到一种决定性的作用[19 ] 。这个重要结构的功能,至今还不清楚。据推测,为了功能上的有效性,一个指定的生化过程中有关的每一组酶和载体分子,应该是按精确的形式彼此紧挨着排列在适当的位置上。我们不知道是否这种超分子结构完全是由各种成分的内在性质来决定的,而这些成分在离体情况下是能改变功能结构的(就像用单体蛋白质来装配病毒的外壳或细菌的鞭毛一样);或者是否在一个生长细胞的膜中,这种预先存在的分子组织的构型在有规律地添加新的功能成分时,起到某种作用——引发作用或甚至是催化作用,例如,使无活性的前体物转变成有活性的成分。在某些复合病毒[20 ] 蛋白质外壳的装配中,对这样一些酶促步骤的出现,已有一些提示性的证据。一种更引人感兴趣的可能性是:膜的结构可能不仅起到规定位置的作用,而且还可以起到活性组分(例如,通过传递构型的信号)的作用。这可以提供另一种水平的细胞功能调节。
这就是大肠杆菌素有趣的地方。大肠杆菌素是蛋白质类抗菌素,它能杀死大肠杆菌型细菌的敏感菌株,它是由这类细菌的另一些带有相应的遗传决定子或叫大肠杆菌素源因子的菌株所产生的。很早就知道,一些大肠杆菌素能停止敏感细胞大分子成分的合成[21 ] 。一个主要的进展是发现了不同的大肠杆菌素引起不同的生化变化[22 ] ,通过用胰酶从细胞受体上把大肠杆菌素降解掉,可以使某些大肠杆菌素的致死作用得到逆转[22 ,23 ] 。这种来自外界的作用,并结合大肠杆菌素致死的单击动力学,可以表明在细胞被膜表面的一种单一的大肠杆菌素分子,通过细胞被膜本身的扩增机制,可以起到抑菌或杀菌的效能。因此,Nomura[22 ] 主张:一种附着在适当受体上的大肠杆菌素,通过引起细胞质膜的某种特异成分的功能改变,作用于一个特异性的生化靶子上。Nomura[22 ] 和我[24 ] 曾经考虑到作为一个整体来看,扩增机制可能是通过细胞膜的构型改变来传递的。Changeux和Thiery[25 ] 基于对膜蛋白之间变构作用的考虑,以一种更特异性的方式提出了相同的看法。
Nomura[22 ] 识别大肠杆菌素作用的3种类型是:
(1)DNA合成停止,DNA被断裂,这是大肠杆菌素E2的典型作用。
(2)抑制蛋白质合成,这是大肠杆菌素E3的特性,这种作用可以追溯到是30S亚基某种成分上的特异性改变[26 ] 。
(3)大分子合成全部停止,这是很多大肠杆菌素(E1、K、A、I)的共同作用机制。
在大肠杆菌素E2和E3的情况中,生化效应的大小完全取决于增殖能力,而致死作用(根据失去生长能力来确定)是严格单击动力学类型的。因此,存在着一些问题,诸如观察到的特异性效应,到底是初级的还是次级的。然而,对大肠杆菌素K和E1来说,致死效应和抑制增殖能力与初级效应之间具有很好的相关性,观察到的生化现象与初级效应之间可能具有更直接的关系。
一个大肠杆菌素分子怎样抑制所有大分子的合成呢?一个重要的发现(F.Levinthal和C. Levinthal,私人通信)是当大肠杆菌素E1与在严格无氧条件下培养的大肠杆菌起作用时,蛋白质或核酸的合成是不受抑制的;放入空气,引起了一种迅速的但可逆的抑制。观察到的上述现象,与RNA和蛋白质合成的抑制是同时发生的,并没有前后顺序,这一事实使Levinthals提出了大肠杆菌素E1的初级作用,是在氧化磷酸化(一种细胞质膜的功能)上起作用的。ATP水平严重下降,几乎达到0。
根据这些情况,再加上我们对细菌膜中的大调节机制的兴趣,我和我的合作者打算协作弄清大肠杆菌素的作用与膜性质(如透性和输送)改变的相关性。我打算只做关于大肠杆菌素E1和K方面的工作,对此我有一些成功的把握。
我和Kay Fields首先对用大肠杆菌素处理过的大肠杆菌细胞[27 ] ,在β-D-半乳糖苷输送和积累方面可能发生的改变进行了研究。结果表明,依赖能量的积累过程受到严重抑制,而邻位-硝基苯基半乳糖苷(ONPG)的输送速率(用完整细胞的半乳糖苷酶水解ONPG的速率来测定)几乎不受影响。因此,ONPG是透不过细胞的。α-甲基葡萄糖苷的积累(靠磷酸烯醇丙酮酸而不是靠ATP[28 ] 来推动)对大肠杆菌素E1或K都是不敏感的,这证明在受大肠杆菌素抑制的细胞中(糖原)酵解能继续进行。
当我们用受大肠杆菌素处理过的细胞来继续进行葡萄糖命运的研究时,出乎意料地发现了一个我们所期待的证据——膜透性的一种特异改变[29 ] 。用大肠杆菌素处理过的这种细胞,向培养基中分泌出几乎达1/3的葡萄糖衍生碳源,例如6-磷酸葡萄糖、1,6-二磷酸果糖,二氢丙酮磷酸盐和3-磷酸甘油酸盐。其他中间产物的分泌量测不出来。此外,葡萄糖分解代谢的主要快速产物是丙酮酸盐而不是乙酸盐和CO2 。这并不是由于丙酮酸盐的渗漏引起的,而是因为当大肠杆菌素处理过的细胞,乳酸脱氢酶的水平高时,丙酮酸盐可以转变成乳酸盐。这个分解代谢反应产生丙酮酸盐,而不产生乙酸盐。这反映了丙酮酸盐氧化作用直接或间接地受到特异性的抑制。
还了解到,对能量代谢的影响要比对刚才说的那种氧化磷酸化作用的抑制更为复杂。如果在适当条件下,而不是在严格无氧条件下,把大肠杆菌在葡萄糖中发酵培养,它的蛋白质和核苷酸的合成对大肠杆菌素抑制作用的敏感性与需氧细胞中是一致的。在不完全缺氧条件下,即使受空气强烈抑制的氯高铁血红素缺陷的突变体,对大肠杆菌素也是敏感的。
这些观察揭示了,这些大肠杆菌素的早期效应可能是一种(可逆的)细胞质膜的变化,这种变化需要有氧存在,通过限制ATP的利用效力,使依赖ATP的生化过程发生阻断。这可能是由于ATP的形成减少了,或ATP的分解增加了。另外,不管剩余的ATP水平有多高,生物合成的过程也要受阻,其中部分原因可能是由于AMP的积累,而使AMP/ATP的比值升高造成的[30 ] 。事实上,AMP激酶热敏感的一种大肠杆菌突变体,在高温下的行为与受大肠杆菌素抑制的细胞十分相似[31 ] 。
在进一步探索大肠杆菌素对膜的影响的工作过程中,一位客人David Feingold,去年在我们的实验室中研究了在有羰基氰化物m-叶绿素腙(CCCP)——能促进H+ 渗透的氧化磷酸化作用的一种强解偶联剂——存在时,大肠杆菌素E1对细菌吸收质子的影响[32 ] 。大肠杆菌素本身不能引起对质子的渗透性增加;事实上,用洗过的正常的细胞观察到,它能防止缓慢的pH值上升。即使经低倍数的大肠杆菌素处理,也能使细菌变得对CCCP敏感,以至于只要在细菌悬浮液中加入小至10-6 M的CCCP时,几乎立即发生平衡。这样看来,在大肠杆菌中;大肠杆菌素E1的作用很像缬氨霉素对革兰氏阳性细菌的效应[32 ] 。Hirata等人[33 ] 也分别得到了类似的发现。Feingold实验室正在研究这种大肠杆菌素的效应,是抑制能量代谢的次生作用呢?还是对渗透性的一种特异效应呢?例如当K+ 和H+ 通过CCCP的作用彼此接近时,K+ 能与H+ 发生交换,从而改变K+ 的渗透性。
耐大肠杆菌素的膜突变体
另一些观察使我们能将细菌对大肠杆菌素的反应与细菌被膜的功能特性结合起来。我和Rosa Nagel de Zwaig[34 ] 一起研究了一类耐某些大肠杆菌素的细菌突变体,它们能吸附大肠杆菌素,但不受抑制。在其他几个实验室中,也研究了相似的耐性或抗性突变体。膜对大肠杆菌素的作用与所预料的一致,我们满意地发现:我们测定过的所有耐性突变体,都有一些膜的缺陷。某些类型的突变体是很脆弱的,以至于很多菌体在生长过程中自发溶解了,仿佛这些细胞被膜的合成是缺陷的。像肠道细菌其他的被膜缺陷突变体一样,这些耐性突变体对脱氧胆酸盐也是十分敏感的,可能是因为这些突变体的被膜变得易受这种表面活性剂的影响。更有趣的是,还有一类耐性突变体,对很多有机染料,主要是阳离子染料,如吖啶、溴化乙啶以及亚甲基蓝,都是非常敏感的。我们可以证明,对染料的这种敏感性,是因为突变体细胞快速地摄取染料造成的,而正常细胞对这些染料几乎无透性。因此,这种耐大肠杆菌素的突变与膜透性的一种特异性改变是相关的 【102】 。
正常细菌和耐性突变体细胞被膜的一些初步分析研究,并没有揭示出它们之间有任何的明显差异。肠道细菌细胞被膜在化学上是极复杂的,对它了解得还很少。即使发现有化学变化,也不容易确定它们是否直接与所研究的现象有关。上述情况确实存在,例如,在大肠杆菌素处理过的细菌中报道了有磷脂组分的变化[35 ] 。然而,令人鼓舞的是有关某些大肠杆菌素的研究和耐大肠杆菌素突变体的研究,都已集中到我们所注意的焦点上,即大肠杆菌素作用的位点与细菌膜功能之间的相关性。目前,这种关系还只是推理性的,并无可靠根据。但是,这些观察是非常鼓舞人心的,它增强了我们的信心,这就是对大肠杆菌素的研究,可以在膜的水平上揭示出控制着细菌细胞的某些主要功能是如何形成的。
跋
目前大肠杆菌素研究的状况与早在20世纪40年代噬菌体研究的情况之间,有着有趣的相似之处。在这两种情况中,都是由先驱的研究者先提出现象,再由一个工作小组,对有关的新目标进行检验。在噬菌体研究中,目标是要得到增殖的基本现象,希望病毒的增殖将有助于阐明遗传物质的复制。在大肠杆菌素的研究中,目标是要揭示细菌细胞质膜的功能,推测某些发现可能有助于阐明细胞膜功能的普遍性问题。在这两种情况中,都是使用简单的细菌系统,通过突变体来研究各自的遗传学和膜学规律。
如同25年前噬菌体研究的情况一样,今天从事大肠杆菌素研究的人员甚少,他们互相协作,对成功充满信心地开展工作——但也有点担心,成功可能也会使这个平静的研究领域转变成一种臃肿的学院式研究[37 ] 。还是如同噬菌体研究一样,我们知道,只有当我们正在研究的这些问题具有可靠的生化手段时,才能使问题得到圆满解答。这个问题可能和已弄清楚的基因功能和复制一样,是一门新颖的生物化学课题。今后也许我们能再次发现某个意义深远的、鼓舞人心的重大问题。
参考文献
[1 ]Luria, S. E., and Delbrück, M., 1942. Arch. Biochem.,1 , 207.
[2 ]Wyatt, G. R., and Cohen, S. S., 1952. Nature,170 , 1072.
[3 ]Cohen, S. S., 1961. Virus-induced Enzymes, Columbia University Press, New York.
[4 ]Cairns, J., and Davern, C. I., 1966. J. mol. Biol.,17 , 418.
[5 ]Kutter, E. M., and Wiberg, J. S., 1968. J,mol. Biol.,38 , 395.
[6 ]Aposhian, H. V., 1968. in H. Fraenkel-Conrat (Ed.),molecular Basis of Virology, Reinhold, New York, 497.
[7 ]Roscoe, D. H., 1959. Virology,38 , 527.
[8 ]Kaempfer, R, O. R., and Magasanik, B., 1967. J. mol. Biol.,27 , 453.
[9 ]Burgess, R. R., Travers, A. A., Dunn, J. J., and Bautz, E. K. F., 1969. Nature,221 , 43.
[10 ]Summers, W. C., and Siegel, R. B., 1969. Nature,223 , 1111.
[11 ]Hosoda, J., and Levinthal, C., 1968. Virology,34 , 709.
[12 ]Travers, A. A., 1969. Nature,223 , 1107.
[13 ]Sonenshein, A. L., and Roscoe, D. H., 1969. Virology,39 , 205.
[14 ]Losick, R., and Sonenshein, A. L., 1969. Nature,224 , 35.
[15 ]Britten, R. J., and Davidson, E. H., 1969. Science,165 , 349.
[16 ]Iseki, S., and Sakai, T., 1953. Proc. Japan Acad.,29 , 121.
[17 ]Uetake, H., Luria, S. E., and Burrous, J. W., 1958. Virology,5 , 68.
[18 ]Wright, A., Dankert, M., and Robbins, P. W., 1965. Proc. Natl. Acad. Sci. USA, 54 , 235.
[19 ]Ryter, A., Hirota, Y., and Jacob, F., 1968. Cold Spring Harbor Symposia Quant. Biol.,33 , 669.
[20 ]Wood, W. B., Edgar, R. S., King, J., Lielausis, I., and Henninger, M., 1968. Federation Proc.,27 , 1160.
[21 ]Jacob, F., Siminovitch, L., and Wollman, E., 1952. Ann. Inst. Pasteur,83 , 295.
[22 ]Nomura, M., 1963. Cold Spring Harbor Symposia Quant. Biol.,28 , 315.
[23 ]Reynolds, B. L., and Reeves, P. R., 1963. Biochem. Biophys. Res. Communs.,11 , 140.
[24 ]Luria, S. E., 1964. Ann Inst. Pasteur,107 , 67.
[25 ]Changeux, J. P., and Thiery, J., 1967. J. Theoret, Biol., 17, 315.
[26 ]Koniskey, J., and Nomura, M., 1967. J. mol. Biol.,26 , 181.
[27 ]Fields, K. L., and Luria, S. E., 1969. J. Bacteriol.,97 , 57.
[28 ]Kundig, W., Ghosh, S., and Roseman, S., 1964. Proc. Natl. Acad. Sci. USA, 52 , 1067.
[29 ]Fields, K. L., and Luria, S. E., 1969. J. Bacteriol,97 , 64.
[30 ]Atkinson, D. E., 1966. Ann. Rev. Biochem., 35 , 85.
[31 ]Cousin, D., 1967. Ann. Inst. Pasteur, 113 , 309.
[32 ]Harold, F. M., and Baarda, J. R., 1969. J. Bacteriol.,96 , 2025.
[33 ]Hirata, H., Fukui, S., and Ishikawa, S., 1969. J. Biochem.,65 , 843.
[34 ]Nagel de Zwaig, R., and Luria, S. E., 1967. J. Bacteriol.,94 , 1112.
[35 ]Cavard, D., Rampini, C., Barba, E., and Polonovski, J., 1968. Bull. Soc. Chem. Biol.,50 , 1455.
[36 ]Nagel de Zwaig, R., and Luria, S. E., 1969. J. Bateriol.,99 , 78.
[37 ]Stent, G. S., 1969. Science,166 , 479.
核糖核酸指导的脱氧核糖核酸合成 【103】
霍华德·特敏(H.M.Temin) 【104】
(美国 威斯康星大学McArdle实验室,1972年)
一
现代生物学的主要目标是研究遗传信息在分子结构内是如何编码的,以及在生物系统中如何从一种分子传递到另一种分子。揭示控制这种传递的规律,对于了解动物和植物体内胚细胞如何分化成上百种不同类型的细胞,以及正常的健康细胞如何变成癌细胞,都具有重要的意义。
最近20年来,已经知道在所有活细胞内,遗传信息都编码在由两条DNA长链构成的双螺旋分子内。每种生物的遗传信息都是由4个字母(即4种不同的碱基)的文字写成的。在正常细胞内,遗传信息短句(单个基因)从DNA转录到密切相关的单链RNA分子上,然后,代表一个基因的一段RNA链再被翻译成一种特异的蛋白质,蛋白质是由20个字母(即20种不同的氨基酸)的文字构成的。细胞分裂时,DNA双链中每一条链上的遗传信息都进行复制,从而保证子代细胞具有亲代的全部遗传性状。
DNA双螺旋结构的发现人之一克里克(F.Crick),最初提出的假说认为,遗传信息可以从核酸传到核酸,从核酸传到蛋白质。但是,遗传信息一旦传到蛋白质,就不可能逆向得到它。也就是说,遗传信息不能从蛋白质传到蛋白质,也不能从蛋白质传到核酸。这些概念简称为分子生物学的“中心法则”。根据中心法则,遗传信息连续地从DNA传到RNA,从RNA传到蛋白质(见图1)。尽管克里克的原始公式并不排斥遗传信息从RNA到DNA的逆向传递,但自然界中的生物体似乎并不需要这样一种逆向传递,而且,有很多生物学家认为,一旦揭示出这种逆向传递,就会动摇中心法则。
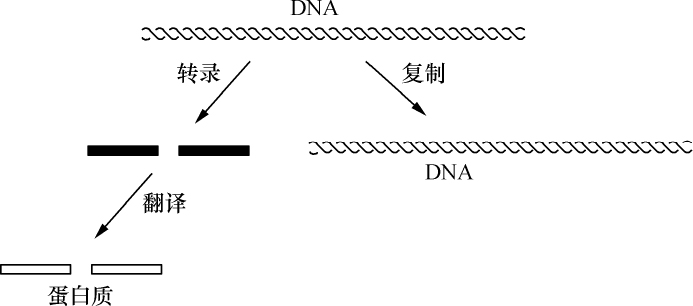
图1 中心法则
最初由克里克提出的分子生物学的“中心法则”认为:生物体内遗传信息可以从DNA传递到DNA,或者从DNA到RNA再到蛋白质,但不能从蛋白质传到蛋白质,也不能从蛋白质到它们的DNA和RNA。虽然Crick最初提出的公式没有排除遗传信息从RNA到DNA的逆向传递,但许多分子生物学家认为,将来一旦发现这样一种逆向传递,就会动摇中心法则。
下面我介绍一下最初揭示出遗传信息从RNA传递到DNA的有关实验。这些实验不仅有力地证明了遗传信息的逆向传递确实存在,而且也只有用逆向传递才能解释一大类动物病毒的反常行为,这些动物病毒的遗传信息编码在RNA分子上,而不是在DNA分子上。这类病毒中有许多能诱发动物致癌。虽然目前尚未证实这些病毒与人癌有关,但是,揭示这类病毒在活细胞内将遗传信息从RNA传递到DNA的机制,有可能为统一两种过去似乎是互不相干的人类致癌假说,即遗传假说和病毒假说作出贡献。
病毒分为两大类,即由DNA组成基因组(或者说整套基因)的病毒和由RNA组成基因组的病毒。在病毒感染的细胞中,DNA病毒能进行复制,产生新的DNA,并把遗传信息从DNA传递到RNA,再从RNA传到蛋白质。大多数的RNA病毒,例如引起脊髓灰质炎的各种病毒、普通感冒病毒和流感病毒,则直接把RNA复制成新的RNA,并把遗传信息从这些RNA分子传递到蛋白质,在它们的RNA复制过程中,不直接涉及DNA。
近几年来,已经清楚,各种RNA肿癌病毒(又称白血病病毒或劳斯氏病毒)是通过另一种信息传递方式进行复制的。劳斯氏病毒除了能用在细胞或DNA病毒中发现的信息传递方式外(从DNA到DNA、DNA到RNA、RNA到蛋白质),还能利用从RNA到DNA的信息传递方式。劳斯氏病毒与其他RNA病毒不同,不是把信息从RNA传到RNA。由于在劳斯氏病毒复制中存在RNA到DNA的信息传递方式,因此,有人建议把病毒分为三大类:DNA病毒、RNA病毒和RNA-DNA病毒(见图2)。
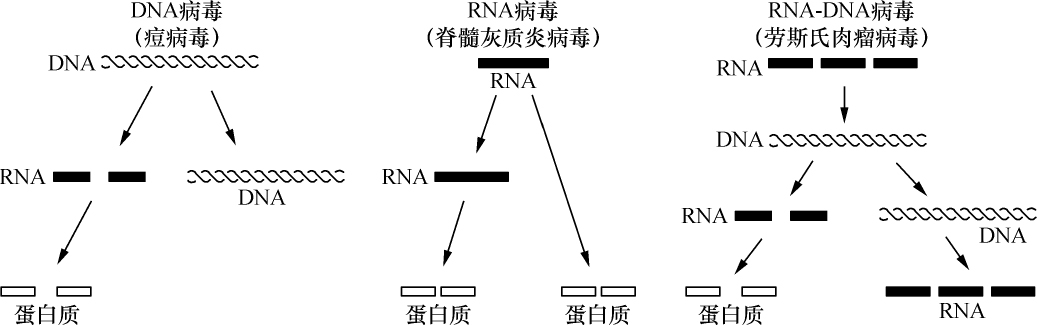
图2 病毒可以分为三大类
(1)DNA病毒(左边的),它的基因组(或者说整套基因)是由DNA组成的;(2)RNA病毒(中间的),它的基因组是由RNA组成的;(3)RNA-DNA病毒(右边的),它是最近发现的、基因组由RNA和DNA交替组成的病毒。图中括号内为各类病毒的原型肿瘤病毒。图解表示每一类病毒复制时所特有的信息传递方式。
61年前,劳斯氏在洛克菲勒医学研究所发现了RNA肿瘤病毒的原型——鸡的劳斯氏肉瘤病毒。实际上,早在劳斯氏之前,哥本哈根的V. Ellerman和O.Bang已经发现了RNA肿瘤病毒,但由于这种病毒能引起鸡的白血病,又比劳斯氏肉瘤病毒更难对付,因此,他们没有对这种病毒进行深入的研究。Rous研究了在Plymouth种鸡身上移植肿瘤,开始他观察到,通过移植细胞,可使肿瘤移植。1911年他又发现,用无细胞滤液仍然可以使肿瘤移植。一般都认为能通过无细胞滤液进行传播的疾病都是由病毒引起的。现在,用Rous发现的病毒后代进行研究工作的实验室遍布世界各地。但当时大家还不相信Rous的发现。过了10年,Rous自己也停止了肿瘤的研究工作。大约过了30年,美国纽约市Bronx区的退役军人医院的L. Gross发现了RNA肿瘤病毒引起小鼠白血病,这样劳斯氏(Rous)病毒的研究才普遍开展起来。
现在已经知道,和劳斯氏病毒同一大类的病毒或它的亲缘病毒,不仅能在雏鸡和小鼠身上引起肿瘤,而且在大鼠、仓鼠、猴子以及各种动物身上,也能引起肿瘤。从蛇类等非哺乳动物中也分离出了这类病毒。但是,到现在为止,还没有发现真正的人类劳斯氏病毒。看来,这类病毒的某些种,例如某些“辅助病毒”并不致癌。
二
20世纪50年代,随着细胞培养法开始应用于动物病毒学研究,对劳斯氏肉瘤病毒的组织培养也发展起来了。首先由R. A. Manaker和V. Groupé在Rutger大学开拓了应用组织培养物研究劳斯氏肉瘤病毒的研究工作,随后,H. Hubin和我在加利福尼亚理工学院也进行了这方面的研究工作。其方法就是把病毒悬液加到由鸡胚体壁细胞形成的稀薄的细胞培养物中,劳斯氏肉瘤病毒能感染某些细胞,并把它们转化成肿瘤细胞。这些转化了的肿瘤细胞,在形态和生长特性上都与正常细胞不同,结果就产生了变异细胞群。用劳斯氏肉瘤病毒感染由火鸡、鸭、鹌鹑及大鼠得来的细胞,更进一步发展了这种试验方法。用其他转化的病毒也建立起了类似的试验方法。
转化细胞群数目与加到细胞培养物中的有感染性的病毒颗粒数成正比。这为测定劳斯氏肉瘤病毒提供了一种迅速、而且重复性也好的试验方法。这种试验法的利用导致以下发现,即在与细胞相互作用的方式上,劳斯氏肉瘤病毒与当时已经研究过的其他病毒是不同的。多数病毒的增殖与细胞的分裂是对抗的。换言之,病毒引起感染细胞死亡。而感染劳斯氏肉瘤病毒的雏鸡细胞,不仅存活下来了,而且继续分裂并产生新的病毒颗粒(见图3)。当用劳斯氏肉瘤病毒感染大鼠细胞时,细胞和病毒间的相互作用与上述情况有所不同,大鼠细胞转化成了癌细胞,这些细胞仍能分裂,尽管可以证明,这些转化了的细胞中有病毒基因组(DNA)存在,但不产生劳斯氏肉瘤病毒。如果雏鸡的正常细胞和大鼠的转化细胞发生融合,即可引起劳斯氏肉瘤病毒的形成。
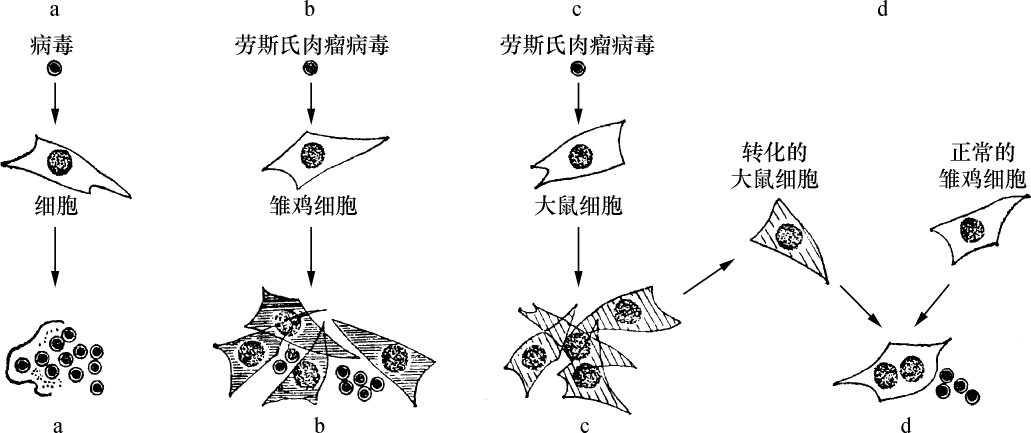
图3 病毒与细胞的相互作用
由于大多数病毒与细胞分裂是对抗性的,因此,病毒与细胞的相互作用,通常导致感染细胞的死亡(a)。但劳斯氏肉瘤病毒以不同的方式与细胞相互作用,因此,感染劳斯氏肉瘤病毒的雏鸡细胞,不仅活下来了,而且转化成了能继续分裂并产生新病毒颗粒的癌细胞(b)。感染劳斯氏肉瘤病毒的大鼠细胞,转化成能分裂的癌细胞,但不产生新的病毒颗粒(c)。把转化的大鼠细胞与正常的雏鸡细胞融合,可以诱发劳斯氏肉瘤病毒颗粒的形成(d)。
20世纪60年代初期,发现抗菌素放线菌素D,可以用来阐明在RNA病毒感染的细胞内遗传信息的传递方式。这种抗菌素能抑制以DNA为模板的RNA合成,但不影响以RNA为模板的RNA合成。因此,除了与病毒基因组有特殊关系的RNA外,这种抗菌素停止了RNA病毒感染的细胞中其他一切RNA的合成。用这一新方法,很容易确定,哪种RNA分子对于病毒是特异的。
当我把放线菌素D加到形成劳斯氏肉瘤病毒的细胞培养物中,发现这种抗菌素抑制了全部RNA的合成。而正像我们所期待的那样,以一种RNA病毒基因组为模板复制RNA,可以无阻碍地连续进行(见图4)。这一结果首次直接证明了,劳斯氏肉瘤病毒复制的分子生物学与其他RNA病毒是不同的。从观察到上述结果的时候起,放线菌素D能抑制劳斯氏病毒的复制被公认为是这些病毒的确定特性之一。放线菌素D的实验向我们揭示了,劳斯氏肉瘤病毒可能通过一种DNA中间物而进行复制,这种假说被称为DNA前病毒假说。
图4 RNA的合成
在正常细胞中,以DNA为模板合成RNA(a)。加入抗菌素放线菌素D会抑制这一合成(b)。由于放线菌素D并不影响以RNA为模板合成RNA,因此,在大多数RNA病毒感染的细胞中,这种抗菌素不抑制与病毒基因组有关的RNA分子的合成(c)。在产生劳斯氏肉瘤病毒的细胞中,放线菌素D能抑制所有RNA的形成(d)。这一发现是说明劳斯氏肉瘤病毒与其他RNA病毒复制的分子机制不同的第一个直接证据。放线菌素D的实验使作者提出了DNA前病毒假说,假说主张像劳斯氏肉瘤病毒这样的劳斯氏病毒是通过一种DNA中间物进行复制的。
我和J. P. Bader在美国国立癌症研究所中所作的进一步实验表明,如果在细胞接种劳斯氏肉瘤病毒之后,立即抑制细胞的DNA合成,就能保护细胞不被感染。抑制剂有:氨甲蝶呤、氟代脱氧尿嘧啶核苷、阿拉伯糖胞嘧啶核苷。这些实验似乎支持了认为感染细胞必须以RNA为模板合成新病毒DNA的想法。但是这种解释是很含糊的,因为劳斯氏肉瘤病毒的增殖需要细胞在被感染之后能正常分裂。因此,在感染之后抑制DNA的合成,就会抑制劳斯氏肉瘤病毒的增殖,因为这不仅封阻了新病毒DNA的合成,而且也抑制了细胞的正常分裂。
三
为了解决这个问题,我采用了以劳斯氏肉瘤病毒感染静态(或称为不分裂的)细胞培养物的方法。培养物中的细胞繁殖需要血清中的某些特殊因子,如果从培养基中去掉血清,细胞就停止分裂。如果将这种培养细胞暴露于劳斯氏肉瘤病毒中,细胞就会被感染,但是,在重新加入血清并使细胞再次分裂之前,并不发生病毒的增殖和细胞形态变化。在静态细胞中,加进DNA合成的抑制剂,由于这些细胞不合成DNA,因而并不死亡。在静态细胞中,同时加入劳斯氏肉瘤病毒和DNA合成抑制剂,细胞既不死亡,也不被感染(见图5)。
图5
作者和J.B.Bader在美国国立癌症研究所进行的实验,支持了下述假说,假说认为用劳斯氏肉瘤病毒感染细胞,需要以RNA为模板合成新的病毒DNA。把病毒加到正常分裂的细胞培养物中,细胞转化为癌细胞,这种癌细胞能进行分裂并产生新的劳斯氏肉瘤病毒(a)。如果在细胞接种了劳斯氏肉瘤病毒之后,立即加入细胞DNA合成的抑制物,能够保护细胞免受感染(b)。作者在后来的实验中,使用了静态的(或者说不分裂的)细胞培养物,当在培养物中加入血清时,细胞开始正常分裂(c)。但是,如果这种静态细胞先暴露于劳斯氏肉瘤病毒,虽然细胞受到感染,但直至重新加入血清细胞重新分裂之前,并不产生病毒颗粒和形态变化(d)。当细胞同时暴露于劳斯氏肉瘤病毒和DNA合成的抑制物时,细胞没有死亡,也没有被感染;当把DNA合成的抑制物除去并加入血清时,细胞正常分裂,不被感染,不发生转化,也不产生病毒。
如果这时去掉DNA合成的抑制剂,加入血清,能促使细胞重新分裂。我们发现细胞依然不被感染,这些细胞不转化成肿瘤细胞,也不产生病毒。这些实验支持了下述假说,即细胞被劳斯氏肉瘤病毒感染之后,就合成新的病毒DNA,这种病毒DNA合成不同于细胞正常的DNA合成。显然,这种新的病毒DNA是以病毒RNA为模板合成的。
我的学生D. E. Boettiger,为了解决劳斯氏肉瘤病毒复制问题,又发展了上述研究。Rochester大学医疗和牙科学校的P. Balduzzi和H. R. Morgan也独立地进行了这方面的研究。另一些研究人员发现,如果将DNA组分胸腺嘧啶的类似物——5-溴代脱氧尿嘧啶,参入到DNA中去,DNA即敏化,以至于能被光失活。在同样的条件下,光不能使正常的、含胸腺嘧啶的DNA失活。因此,Boettiger在有5-溴代脱氧尿嘧啶存在的情况下,使静态细胞与劳斯氏肉瘤病毒接触,然后再用光照射这些细胞,光照并不杀死细胞,而且这一处理反而保护细胞免受病毒的感染。重新加入血清,使细胞能够分裂,但这些细胞并不发生转化,也不产生病毒(见图6)。
图6
作者的学生D. E. Boettiger(Rochester大学的P. Balduzzi和H. R. Morgan也独立地进行了这些实验)所做的实验说明,在有5-溴代脱氧尿嘧啶(DNA组分胸腺嘧啶的一种类似物)存在的情况下,用劳斯氏肉瘤病毒感染静态细胞,当5-溴代脱氧尿嘧啶参入DNA, DNA即敏化,致使在光照下失活。左图是对照,处理过的细胞开始不经过光照,在加入血清之后,这些细胞重新分裂,同时转化成了癌细胞,并开始产生病毒颗粒。右图,另一组处理过的细胞经过光照,细胞未被杀死,这种处理保护了这些细胞免受病毒的感染,重新加入血清,使这些细胞能够分裂,但它们不能发生转化。
Boettiger进行的与此有关的一个实验表明,细胞被劳斯氏肉瘤病毒感染后的失活速率取决于感染每个细胞的病毒数目。当感染每个细胞的病毒数目增加时,他发现感染增加了对光失活作用的抵抗能力。我们对这些实验的解释如下:感染细胞的每种病毒,产生一种新的特异DNA,侵入每个细胞的病毒数目越多,产生新病毒DNA的分子也就越多。看来这一实验有力地排除了下述假说,即感染病毒引起某种原有的细胞DNA的新合成。
遗憾的是,到目前止,谁也没能确切证明:在劳斯氏肉瘤病毒感染的细胞中存在新合成的DNA。要检查这些期望存在的新的微量病毒DNA,现在所用的这些技术显然是太粗糙了。但是,已有报道说用转化细胞取得了一定的结果。方法之一就是把感染细胞的DNA分子同标记的病毒RNA混合,观察两种分子的单链是否会结合成双链杂种分子。当DNA中的碱基顺序与RNA中的碱基顺序互补时,这种杂种分子是很容易产生的,这说明DNA和RNA带有相同的遗传信息,因而每种分子都能够由另一种分子转录产生。
迄今为止,已经报道过的杂交实验结果引起了激烈地争论。虽然有些实验(最著名的是洛杉矶加利福尼亚大学的M. Baluda和D. P. Nayak等的一些实验)好像说明感染细胞中存在与病毒RNA互补的DNA,但是,这个结果还没有被普遍承认。找到一种中间物病毒DNA,是稳固确立DNA前病毒学说最重要的证据。
与此同时,各种不同类型的实验都为这一假说提供了有力的证据。1969年,曾写过关于细菌病毒博士论文的S. Mizutani,来到我们实验室进行博士学位后的进修。我们决定提出这样一个问题:以病毒RNA为模板合成前病毒DNA的酶(一种蛋白质)的来源是什么?Mizutani发现,在有蛋白质合成抑制物存在的条件下,用劳斯氏肉瘤病毒处理静态细胞培养物,细胞仍能受到感染,我们认为这个实验说明,能以病毒RNA为模板合成DNA的酶,在细胞受到感染之前就已经存在了。
不久前,有人分离出了病毒颗粒(这种真正的病毒颗粒与细胞内病毒所具有的形态是不同的),以及发现了RNA聚合酶,这种酶催化由4种不同的核糖核苷三磷酸合成RNA。1967年,普林斯顿大学的J. Kates和B. R. McAuslan以及Roswell Park研究所的W. Mungon、E.Paoletti和J. T. Grace在一种大型DNA病毒——痘病毒中,发现了RNA聚合酶。另外有人在一种肠道呼吸道病毒(一种双链RNA病毒)中发现了另一种RNA聚合酶。因此,我们决定,在劳斯氏肉瘤病毒颗粒中寻找能以病毒RNA为模板的DNA聚合酶。经过几个月的初步实验,我们成功地证明了在纯化的劳斯氏肉瘤病毒颗粒中,存在一种DNA聚合酶。
在讨论这个结果之前,我想先暂离本题描述一下劳斯氏肉瘤病毒的结构(见图7)。劳斯氏肉瘤病毒颗粒的直径约为100纳米,比引起脊髓灰质炎的病毒颗粒大些,比引起天花的病毒颗粒小些,劳斯氏肉瘤病毒颗粒是由含脂被膜(由细胞膜凹陷而成)、内膜以及含有病毒RNA和某些蛋白质的类核(或叫核心)构成的(见图8)。
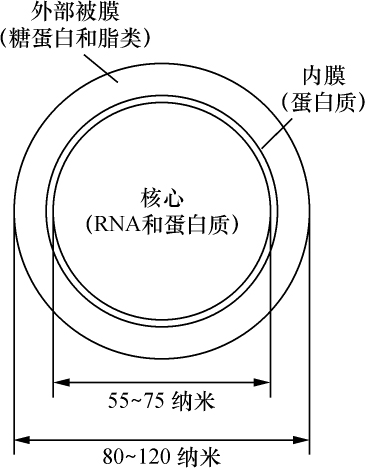
图7 RNA-DNA病毒颗粒结构模式图
图8 RNA-DNA病毒颗粒的电子显微镜照片(直径放大700000倍)
RNA-DNA病毒是一种动物肿瘤病毒,在这种病毒中,除了有正常细胞和别的病毒所用的正常的遗传信息传递方式以外,它还能把遗传信息从RNA传递到DNA。在电子显微镜超薄切片中看到的这种特殊的RNA-DNA病毒颗粒,能引起小鼠产生白血病,它和本文中所讨论的劳斯氏肉瘤病毒在结构和功能上都是十分相似的。图7就是这种类型的病毒颗粒的结构模式图。
为了证明劳斯氏肉瘤病毒确实含有一种能以RNA为模板合成DNA的聚合酶,我们先用去垢剂处理病毒颗粒,使它的含脂被膜发生破裂,然后,在这种已经裂解的病毒中加进构成DNA的原材料——4种脱氧核糖核苷三磷酸。其中一种脱氧核糖核苷三磷酸带有放射性标记。
当混合物在40℃保温时,放射性标记就参入到不溶于酸的物质中,这种不溶于酸的物质通常作为鉴定DNA的物质,它对碱和能破坏RNA的核糖核酸酶是稳定的;相反,能破坏DNA的酶,也能破坏这种物质,使它成为碎片。我们用核糖核酸酶(一种能破坏RNA的酶)预处理已经裂解的病毒颗粒进行重复实验时,只产生极少量的DNA,或不产生DNA,这说明合成DNA需要用完整的病毒RNA作为模板(见图9)。
图9 说明RNA聚合酶存在的实验
作者和他的同事S. Mizutani的实验(麻省理工学院的D. Baltimore也独立地进行了这一实验),证明了在RNA肿瘤病毒中存在一种能以RNA为模板合成DNA的聚合酶。Mizutani和作者进行的实验,先用去垢剂处理纯的劳斯氏肉瘤病毒颗粒,使其含脂被膜破碎,然后在已经裂解的病毒颗粒中加入构成DNA的材料——4种脱氧核糖核苷三磷酸,将混合物进行保温,带有放射性标记的那种脱氧核糖核苷三磷酸参入不溶于酸的物质,这种物质在有核糖核酸酶(一种能分解RNA的酶)存在的条件下是稳定的,但能被脱氧核糖核酸酶(分解DNA的酶)破坏形成碎片。用核糖核酸酶预处理过的已经裂解的病毒颗粒进行重复试验时,不产生或者产生极少量的DNA,这说明合成DNA需要有完整的病毒RNA作为模板。
1970年5月在休斯敦举行的第10届国际肿瘤会议上,我报告了这些结果之后,我们得知麻省理工学院的D. Baltimore独立地用小鼠白血病病毒颗粒作材料也发现了同样的现象。记述这些发现的两篇文章同时发表在1970年6月27日出版的一期英国科学周刊《自然》上,这两篇文章的发表引起了大量的研究工作,其高潮尚未出现。
四
在我们早期的文章中,把这种新的病毒酶叫做依赖于RNA的DNA聚合酶,因为RNA是模板,而DNA是产物。后来,我们和别人都发现,这种酶也可以用DNA为模板合成DNA。因此我们决定把“依赖”这个词换成“指导”。所以现在我们把这种酶叫做RNA指导的DNA聚合酶。这个更正过的新名,没有指出酶的来源和它与其他聚合酶的关系。《自然》杂志最先把这种酶单独称为逆转录酶,由于它的含义不明确,我不喜欢这个名字,但它已得到了广泛地传播。
后来的这些研究都证实了最初的发现,即RNA肿瘤病毒颗粒含有一种DNA聚合酶系统,用去垢剂处理病毒颗粒,可以使该酶系统得到活化,这个酶系统对核糖核酸酶是敏感的。这种病毒颗粒酶只有DNA聚合酶的功能,而没有RNA聚合酶的功能。正像我们已经指出的那样,其他一些无关的RNA病毒也都含有一种RNA聚合酶。
如果将RNA指导的DNA聚合酶分离出来,除去蛋白质,就可以在超速离心机中用蔗糖密度梯度离心法测定这些DNA分子的大小。这些分子极微小,它们的长度小于人们预计的病毒RNA一个完整拷贝应有长度的十分之一。它们为什么这样小,原因尚不清楚。分离出来的DNA在硫酸铯密度梯度中离心(硫酸铯能根据RNA和DNA密度的不同把它们分开),我们发现这种产物具有DNA所特有的密度(见图10)。进一步的鉴定(例如,用专门作用于单链或双链的特异酶进行处理)证明,这种DNA聚合酶系统的产物是双链DNA。从这些研究可以推断,病毒颗粒DNA聚合酶系统合成小片段的双链DNA。
图10 在RNA指导的DNA聚合酶系统中合成的DNA产物的硫酸铯密度梯度离心(硫酸铯根据RNA与DNA密度的不同把它们分开)实验
从这个实验发现,产物具有DNA的密度。根据这一结果再结合其他发现,可以得出以下结论:劳斯氏肉瘤病毒颗粒的DNA聚合酶系统能合成小片段的双链DNA。
很多研究人员业已证明,RNA指导的DNA聚合酶系统合成的DNA,具有与病毒RNA互补的碱基顺序(见图11)。这个结论是从退火实验和杂交实验得出来的。处理由病毒颗粒聚合酶反应得到的标记DNA,使DNA的双链解离;将单链DNA加到未标记的病毒RNA中,将混合物保温,使互补的链能形成杂种复合物。然后再将混合物在硫酸铯密度梯度中超速离心,大约DNA产物总量的一半所形成的带,具有RNA或者DNA-RNA杂种分子的密度特性,而不具有DNA的密度特性。这一实验是很特异的,它说明了病毒颗粒的DNA聚合酶把病毒RNA的碱基顺序复制成DNA的碱基顺序。但是,这一实验仍未证实有类似的复制过程在劳斯氏肉瘤病毒感染的细胞中发生。
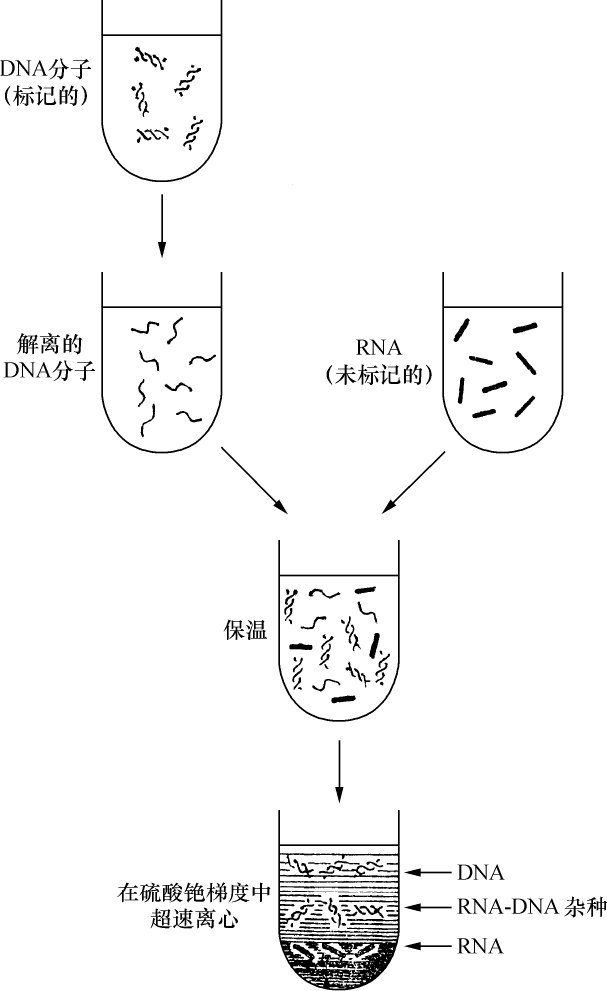
图11 分子杂交实验
分子杂交实验表明,病毒颗粒中,RNA指导的DNA聚合酶系统的DNA产物,把病毒RNA碱基顺序复制成为DNA的碱基顺序。从病毒颗粒的DNA聚合酶反应中产生的标记DNA,首先经过处理使双链解离。然后,将单链的DNA与未标记的病毒RNA混合,混合物在高温下保温,使互补链形成杂种复合体。如果把这种退火得到的混合物在硫酸铯密度梯度中超速离心,结果约有一半的产物DNA形成一个具有RNA-DNA杂种分子密度特性的带。
C. Todaro所领导的小组在美国国立癌症研究所以及J. M. Coffin在威斯康星大学我的实验室所进行的实验,证明了在病毒颗粒的核心中有病毒DNA聚合酶存在(见图12)。用去垢剂处理劳斯氏肉瘤病毒颗粒破坏它的被膜,然后将已经裂解的病毒在蔗糖密度梯度中离心。发现大部分病毒RNA、大约20%的蛋白质以及大部分RNA指导的DNA聚合酶活力,都一起沉降在核心部分(核心这个术语指的是比整个病毒颗粒密度较大的那部分结构)。进一步的研究指出,当病毒颗粒受到更严重的破坏时,病毒DNA聚合酶就能和病毒RNA分开,并被提纯出来。纯化的酶能在各种各样的模板上(例如,合成的DNA、天然的DNA、RNA、DNA-RNA杂种)合成DNA。
图12 J. M. Coffin在作者的实验室所做的工作证明,劳斯氏肉瘤病毒的核心中存在病毒DNA聚合酶
中图的曲线表示整个病毒颗粒各种放射性标记组分的密度分布。右图的曲线表明用去垢剂处理使病毒颗粒的被膜裂解之后,离心测定相同组分的密度分布。裂解病毒颗粒的大部分病毒RNA(曲线1),约20%的蛋白质(曲线2)和大部分RNA指导的DNA聚合酶活性(曲线3)沉降在一起,其密度高于整个病毒颗粒的相应组分,说明这些组分集中在病毒颗粒的核心中。
很多实验室的研究得出了一个普遍性结论,即劳斯氏病毒DNA聚合酶与上述的其他DNA聚合酶是十分相似的,后者是在大家更熟悉的那些生物系统中存在的,并能催化以DNA为模板的DNA合成。换言之,劳斯氏病毒DNA聚合酶能以RNA作为模板合成DNA,这并不是它的独特性质(几年前首先由Sloan-Kettering研究所的S. L. Huang和L. F. Cavalieri提出)。至于在劳斯氏病毒复制中RNA指导的DNA合成所起到的那种显著的生物学作用,至今来说还是独一无二的。
我的实验室做的进一步工作表明,在纯化的劳斯氏肉瘤病毒颗粒中,还含有另外一些与DNA复制有关的酶,其中最罕见的一种是多聚核苷酸连接酶,它能修复DNA分子的断裂。有一种十分有吸引力的假说认为:这种连接酶的功能是把病毒DNA和宿主细胞的染色体DNA连接起来,从而病毒基因组和细胞基因组就整合在一起了。整合之后,该病毒的遗传信息将随着宿主细胞遗传信息的复制而复制,并从亲代细胞传递给子代细胞。肉瘤病毒的病毒颗粒还含有很多作用完全不清楚的其他的酶。我们不知道它们是参与病毒的整个生活史呢,还是在病毒颗粒形成时偶然混进来的杂质。
五
在RNA肿瘤病毒颗粒中,第一次发现了DNA聚合酶之后,又检查了很多其他的RNA病毒,以了解它们是否含有类似的DNA聚合酶系统。最初发现原来属于RNA肿瘤病毒类的全部病毒都含有这样的酶系统。这类RNA病毒,既包括引起肿瘤的劳斯氏病毒,也包括不引起肿瘤的劳斯氏病毒。更有趣的是,发现有两类与RNA肿瘤病毒不属同一类的病毒,也含有这种DNA聚合酶系统。其中之一是绵羊髓质脱失性脑炎病毒,它能引起绵羊发育缓慢的神经疾病。在绵羊髓质脱失性脑炎病毒颗粒中确定了存在DNA聚合酶后,K. K. Takemoto和L. B. Stone在美国国立卫生研究所的工作指出,这种病毒能引起培养中的小鼠细胞癌化。因此,绵羊髓质脱失性脑炎病毒现在可以认为是转化的劳斯氏病毒。另一类被发现具有DNA聚合酶系统的病毒是“泡沫”病毒,或叫做合胞体形成病毒。这些从猴子和猫身上分离出来病毒,还不知道它们与哪种特殊的疾病有关,但常常是细胞培养物中的污染物。到现在为止,还没证实它们具有引起肿瘤和癌化的能力。
在RNA肿瘤病毒中存在的DNA聚合酶,不仅说明了这些病毒如何引起受它们感染的细胞的稳定癌化,而且解释了某些病毒的潜伏现象,即病毒在感染机体之后随即消失,而数月或数年后又重新出现的现象。一种RNA病毒一旦把它的遗传信息传递给DNA,它就在细胞内潜伏起来,并由复制和修复细胞DNA的细胞酶系统来复制。以后,当受到某种激活作用时,又能以有感染力的病毒颗粒形式重新出现(见图13)。
图13 RNA病毒的潜伏现象
RNA病毒感染机体后的潜伏现象,可能和存在于这些病毒核心中的DNA聚合酶系统有关。RNA病毒将其遗传信息传递给细胞核中的DNA之后,就在细胞中潜伏起来,它借助于复制和修复细胞DNA的细胞酶系统来完成其本身的复制,以隐蔽的形式留在细胞中(左中、右中)。数月或数年后,由于某种激活作用可引起有感染性的RNA病毒颗粒的重新出现(右上)。
大约一年之前,报道了在人的肿瘤细胞中发现了依赖于RNA的DNA聚合酶,这一报道引起了普遍的重视。根据上面谈到这方面工作的大部分研究结果,现在得出的普遍性结论是在适当的条件下,所有DNA聚合酶都能把遗传信息从RNA转录到DNA。目前,我们还没有公认的标准来确定这样的合成是否具有生物学意义,以及是否与劳斯氏病毒有关。
我们实验室曾采用了稍微不同的方法来研究细胞内RNA指导的DNA合成问题。我们曾用去垢剂的激活作用和核糖核酸酶的敏感性,来作为在各种动物细胞中粗查DNA聚合酶的指标。我们在细胞中寻找和在病毒核心中类似的DNA聚合酶系统。Coffin曾在正常的、未受感染的大鼠胚细胞中发现了这样的DNA聚合酶系统。到目前为止,我们还不清楚这一发现的全部意义,但是它说明除了肿瘤细胞和被病毒感染的细胞外,正常细胞中也存在对核糖核酸酶敏感的DNA聚合酶系统。
多年来,我一直有这样一个想法,即RNA指导的DNA合成在正常的细胞过程,尤其是和胚细胞分化有关的过程中,起着重要的作用。这个思想发展到形成前病毒假说(见图14)。它的大概意思是在正常细胞中,DNA上有一些区段可以作为合成RNA的模板,而这种RNA反过来又是合成DNA的模板。合成的这种DNA,以后又与细胞DNA整合。DNA的某些区段可能通过这种方式进行扩增。再借助其他一些方法可以使这种扩增DNA发生变化,在不同的细胞中,这种扩增DNA发生的变化是不同的。这种差异可以用作区分不同细胞的手段。
图14 前病毒假说
作者提出的前病毒假说,具体体现了下述设想:RNA指导的DNA合成,可能在正常的细胞过程中起着重要作用。根据这一观点来看:a.在正常细胞中,DNA的某些区段能作为合成RNA的模板;b.这些RNA反过来又作为合成DNA的模板;c.这样合成DNA以后与细胞DNA发生整合;d.由于重复这一过程而引起的某些DNA区段的扩增,这种扩增作用以及扩增DNA变化的其他过程,在胚胎的细胞分化中共同起着重要作用。
那么,这方面工作对预防和治疗人体癌症有什么普遍意义呢?我们只能这样推断,某些生物系统能利用以前未被发现的方式,即RNA到DNA的方式传递遗传信息。这种新的遗传信息传递方式,在引起肿瘤的病毒中首先发现,这只是有趣的巧合。但我们不能说RNA指导的DNA合成是这类病毒的独特性状。RNA指导的DNA合成的发现,是否意味着我们现在已有了简单的生化方法可以测定以下问题:(1)新发现的人类病毒是否和引起动物肿瘤及细胞癌化的RNA病毒属于同一大类;(2)在人类癌细胞中寻找与这些病毒有关的遗传信息。现在我们还不能说,RNA指导的DNA合成的抑制剂对人癌会有什么效果。在引起动物肿瘤的劳斯氏病毒中,新病毒DNA的合成看来只是在癌化初期(而不是在以后)是重要的。
对于人癌的了解来说,这一发现最重要的意义可能就是它消除了关于癌症起源的病毒学说和遗传学说之间的矛盾对立。过去,一种学说认为,基因是由DNA构成的,而且只有突变才能使它发生改变;另一种学说则认为,大多数已知的动物致癌病毒却是由RNA构成的,因此在那个时候,很难设想病毒学说和遗传学说会有共同之点。现在,我们已有充分证据说明,致癌RNA病毒能从病毒RNA转录出DNA来,我们很容易推导出以下假说:与病毒RNA有关的因子与细胞基因组结合,并遗传下去,在将来某一时期被激活,从而引起“自发”癌。旨在检验这一假说的实验,目前正在世界各地的许多实验室中进行。
注释
【1】 梁宏译自F. Miescher. On the chemical composition of pus cells. Great Experiments in Biology , 1957: 233-239.
【2】 约翰·米舍尔(Johan Friedrich Miescher, 1844—1895),瑞士生物学家。他通过对脓细胞的分析发现了一种含磷量极高、还含有氮和硫的成分,这种成分来自细胞核。米舍尔把它命名为核素(nuclein),后来被称为脱氧核糖核酸(DNA)。他认识到核素可能在遗传信息的传递中起重要作用。为证明自己的看法,他进行了大量实验,遗憾的是未能获得理想证据。但为后人(O. T. Avery)证明DNA是传递遗传信息的载体铺平了道路。
【3】 梁宏译自W. Flemming. Contribution to the knowledge of the cell and its life phenomena. Great Experiments in Biology , 1957: 240-245.
【4】 沃尔瑟·弗莱明(Walther Flemming, 1843—1905),德国解剖学家。他研究了细胞分裂,发现了染色体及其有丝分裂过程,成为细胞遗传学的创建者。
【5】 梁宏译自E. V. Beneden. Researches on the maturation of the egg and fertilization. Great Experiments in Biology , 1957: 245-248.
【6】 埃德瓦·贝内登(Edouard Van Beneden,1846—1910),比利时动物学家。他证明了生物体各部分的细胞染色体数相同,染色体数目具有种的特异性。他以染色体数很少的巨头蛔虫(Ascaris megalocephala)为材料进行了研究,发现生殖细胞减数分裂时染色体数减半,并通过受精而恢复原数。
【7】 梁宏译自W. Johannsen. Heredity in populations and pure lines. Classic Papers in Genetics , 1962: 20-26.
【8】 威廉·约翰森(Wilhelm Ludwig Johannsen, 1857—1927),丹麦植物生理学家、遗传学家。他提出了基因、基因型和表型等术语,并在这个基础上对遗传现象进行了系统的分析。在进行纯系研究中引入变异统计而提出纯系内选择无效,证明纯系内的个体差异不是遗传上的变异。
【9】 Die Mutationstbeorie,1 ,p.97.
【10】 德弗里斯的《突变理论》的最后部分是在这本著作的编辑过程中形成的(第503~504页),他以最巧妙的方法证明突变是怎样在多数情况下首先表现出来。从这里得到一个非常重要的例子来解释刚刚讨论的关系。
【11】 梁宏译自W. S. Sutton. The chromosomes in heredity. Biological Bulletin , 1903, 4: 231-251.
【12】 沃尔特·瑟顿(Walter Stanborough Sutton, 1877—1916),美国动物学遗传学家。他研究了染色体的行为和在遗传中的重要性,证明了染色体携带遗传单位基因,并成对出现,将染色体和遗传联系起来,而染色体在减数分裂中的行为是随机的。瑟顿的工作为染色体遗传学说奠定了基础。
【13】 Montgonery于1901年首次得出联会是父本和母本染色体成对联合的结论。
【14】 有许多有机体并非如此。
【15】 考虑到不可能在任一个联会对中辨别其母本和父本成员,要在一个纯种得到绝对的证明是不可能的。但Moenkhaus所得到鱼的杂种能饲养到性成熟,就可以指望在这一点上得到绝对的证明。这种观察能在某些杂种鱼的早期细胞中进行,根据染色体形态上的差别来辨别它是母本的还是父本的,如在成熟分裂时也能这样做,染色体在减数时的分配问题就成为一种非常简单的观察了。
【16】 任何物种单个个体生殖产物的可能组合数可用简单的公式2n 表示。此处n代表减数系统中染色体的数目。
【17】 不管副染色体,它不参加联会。
【18】 Bateson的名词。
【19】 Montgomery(1901)研究过的Protenor和非直翅目昆虫一样,其X染色体和副染色体非常相似,在减数分裂中分开,而不是在均等分裂中分开。这是很值得注意的事实,因为在Protenor中,就和所有已作过研究的半翅目-异翅亚目(Hemiptera)-(Heteroptera)一样,减数在第一次成熟分裂中完成。
【20】 梁宏译自E. B. Wilson. The chromosomes in relation to the determination of sex in insects. Science , 1905, 22: 500-502.
【21】 埃德蒙·威尔逊(Edmund Bee-cher Wilson,1856—1939),美国动物学家、细胞生物学家。他将细胞学与孟德尔学说进行了综合,大大推进了对染色体的研究和理解,开创了染色体学说。
【22】 作者认为副染色体(accessory chromosome)、异向染色体(heterotropic chromosome)或性染色体(idiochromosome)都相当于性染色体。
【23】 直到1932年,C. Bridges发表了一项大量的分析才使此预言得以证实,该分析支持下述假设:可以把性别恰当地看做是一种数量上的连续性,而雄性和雌性只代表了这种数量连续上两个不同的点。已发现某一有机体的性别的实际程度决定于在一种完全可以预测的方式下常染色体对性染色体的比率。果蝇常染色体对雄性为净倾向,而X染色体则对雌性为净倾向。正常的雌性,即含有常规二倍体常染色体组和性染色体,同带有一额外Y染色体的相似雌性,在它们的外观上是无法区别的。
【24】 梁宏译自A. Kossel. The chemical composition of the cell nucleus. Nobel Lectures Physiology or Medicine , 1901-1921: 394-405.
【25】 阿尔布雷希·科塞尔(Albrecht Kossel, 1853—1927),德国生物化学家。他因在蛋白质和核酸成分分析方面的杰出成果而荣获1910年诺贝尔生理学或医学奖。
【26】 梁宏译自T. H. Morgan. Random segregation versus coupling in Mendelian inheritance. Science , 1911, 34: 384.
【27】 托马斯·摩尔根(Thomas Hunt Morgan, 1866—1945),美国遗传学家、动物学家和胚胎学家。他创立了关于遗传基因在染色体上作直线排列的基因理论,并把果蝇400多种突变基因定位在染色体上,制成染色体图谱(即基因的连锁图);确立了伴性遗传规律;发现位于同一染色体上的基因之间的连锁和交换等现象,建立了遗传学的第三定律——连锁交换定律。他获得了1933年诺贝尔生理学或医学奖,被称为“遗传学之父”。后人为了纪念他,将染色体图中基因之间的遗传距离单位叫做“摩尔根”。
【28】 梁宏译自A. H. Sturtevant. The linear arrangement of six sex-linked factors on drosophila, as shown by their mode of association. Journal of Experimental Zoology. 1913, 14: 43-59.
【29】 埃利弗利德·斯特蒂文特(Alfred Henry Sturtevant,1891—1970),美国遗传学家。他是埃德蒙·威尔逊的学生,后到摩尔根实验室工作。他于1913年构建了第一张果蝇的遗传图谱,对摩尔根基因学说的发展作出了重要贡献。
【30】 在这方面,阅读Lock对这个问题的讨论(1906)是有意思的。
【31】 本文送交发表后,观察到一个CR距离内发生3次交换的例子。
【32】 梁宏译自C. B. Bridges. Sex in relation to chromosomes and genes. American Naturist , 1925, 59: 127-137.
【33】 卡尔文·布里奇斯(Calvin Blackman Bridges, 1889—1938),美国遗传学家。他是协助摩尔根完成著名果蝇实验的三个(A. H.Sturtevant, C. B.Bridges, H. J.Muller)得意门生之一,是他首先发现了第一只白眼睛的雄果蝇,并与其他两人合作完成了历时8年的著名果蝇实验,为摩尔根创建基因学说作出了重要贡献。
【34】 括弧内为译者注。
【35】 梁宏译自H. J. Muller. Artificial transmutation of the gene. Science , 1927, 66: 84-87.
【36】 赫曼·缪勒(Hermann Joseph Muller , 1890—1967),美国遗传学家。他在摩尔根指导下用X射线对果蝇进行辐射诱变获得了果蝇白眼及其他突变体,为摩尔根的基因学说提供了证据。缪勒因对X射线的生理和遗传效应的研究取得的杰出成果获得1956年诺贝尔生理学或医学奖。
【37】 梁宏译自K. Lansteiner. On individual differences in human blood. Nobel Lectures Physiology or Medicine , 1922-1941: 234-245.
【38】 卡尔·兰德施泰纳(Karl Lansteiner, 1868—1943),美国免疫学家。他于1901年发现了抗红细胞抗体,后来建立了人类ABO血型系统;1926年他与Philip Levine共同发现了MNP血型系统,1940年与Albert Wiener共同发现了Rh系统。他的发现使人类得以成功地享受正确地血液移植带来的一切好处,兰德施泰纳也因此获得1930年诺贝尔生理学或医学奖。
【39】 系指1930年时的国名。——译者注
【40】 梁宏译自H. B. Creighton, B. McClintock. A correlation of cytological and genetical crossing-over in zea mays. Proceedings of the National Academy of Science. 1931, 7: 492-497.
【41】 哈里特·克赖顿(Harriet Baldwin Creighton, 1909—2004),美国植物学家和遗传学家。她在康奈尔大学读博士期间与巴巴拉·麦克林托克一起在玉米中发现了基因在染色体上的转移现象。
【42】 巴巴拉·麦克林托克(Barbara McClintock, 1902—1992),美国杰出的细胞遗传学家。20世纪30年代她在玉米中发现了基因在染色体上的转移现象。麦克林托克因她在转座基因方面的杰出贡献被授予1983年诺贝尔生理学或医学奖。
【43】 梁宏译自T. S. Painter. A new method for the study of chromosome rearrangements and the plotting of chromosome maps. Science , 1933, 78: 585-586.
【44】 西奥菲勒斯·喷特(Theophilus Shickel Painter,1889—1969),美国动物遗传学家。他首先从染色体(果蝇唾液腺的)中鉴定出了基因;他开创了染色体重排和作图的研究,为动物遗传学作出了重要贡献。
【45】 王斌译自G. W. Beadle, E. L. Tatum. Genetic control of biochemical reactions in Neurospora. Proc. Nat. Acad. Sci. USA. 1941, 27(11): 499-506.
【46】 乔治·比德尔(George Wells Beadle, 1903—1989),美国遗传学家。他因发现基因在细胞生化反应中的调控作用,与E. L. 塔图姆、乔舒亚·莱德伯格共同获得1958年诺贝尔生理学或医学奖。
【47】 在本文原稿送去印刷时,进一步证实了不能合成噻唑和对氨基苯甲酸的两种突变株都是能遗传的,它们与正常菌株也只有单个基因的差别。
【48】 我们打算进一步研究用链孢霉菌株在上述那种玻璃管中的生长情况来检验维生素,为了确定这个方法的可靠性和重复性,因此必须作这样一些辅助试验。
【49】 这3种突变体中的每一种,都可以从正常菌株的提取物中得到它所需要的维生素,这个事实说明这3种维生素都是正常菌株生长所必需的。
【50】 王斌译自O. T. Avery, C. M. MacLeod, M. McCarty. Studies on the chemical nature of the substance inducing transformation of pneumococcal types. J. Exp. Med. 1944, 79(2): 137-158.
【51】 奥斯瓦尔德·埃弗里(Oswald Theodore Avery, 1877—1955),美国内科医生,分子生物和免疫生物化学的先驱。在1944年他和Colin MacLeod及Maclyn McCarty共同发现并证明了DNA是遗传物质,是组成基因和染色体的物质。
【52】 物质或元素的当量是一个比值。在生产或科研上,为了使用方便,用克为单位表示当量。NaOH=23+16+1=40克,则它的克当量为20。这里0.1克当量就是2克。
【53】 以克计的化合物或元素的重量,其量在数值上等于该化合物或元素的分子量。
【54】 即指DNA解聚酶和能破坏转化因素的酶。——译者注
【55】 指蛋白质、荚膜多糖和体细胞多糖。——译者注
【56】 王斌译自J. Lederberg, E. L. Tatum. Novel genotypes in nixed cultures of biochemical mutants of bacteria. Cold Spring Harbor Symp. Quant. Biol. 1946, 11: 113-114.
【57】 对这些实验资料的解释以及另外的一些实验,发表在Gene recombination in Escherichia coli,Nature 153 ,558,1946。
【58】 乔舒亚·莱德伯格(Joshua Lederberg, 1925—2008),美国分子生物学家。因发现细菌的遗传重组,在他33岁时,与E. L. 塔图姆、乔治·比德尔共同获得1958年诺贝尔生理学或医学奖。
【59】 梁宏译自N. D. Zinder, J. Lederberg. Genetic exchange in salmonella. Journal of Bacteriology. 1952, 64: 679-699.
【60】 诺顿·津德尔(Norton David Zinder, 1928— ),美国分子遗传学家。他发现了噬菌体可以携带一个基因从一个细菌到另一个细菌,并把这种交换过程命名为“转导”。后来他又发现了RNA噬菌体及其介导的转导现象。
【61】 王斌译自A. D. Hershey, M. Chase. Independent functions of viral protein and nucleic acid in growth of bacteriophage. J. Gen Physiol. 1952, 36(1): 39-56.
【62】 阿尔弗雷德·赫希(Alfred Day Hershey, 1908—1997),美国微生物遗传学家。他因在噬菌体分子生物学方面的开拓性工作,与Max Delbrück、Salvador Luria共同获得1969年诺贝尔生理学或医学奖。
【63】 mmol/L。
【64】 王斌译自J. D. Watson, F. H. C. Crick. molecular structure of nucleic acid: Nature , 1953, 171(4356): 737-738.
【65】 詹姆斯·沃森(James Dewey Watson, 1928— ),美国分子生物学家和动物学家,美国科学院院士。他因与克里克共同发现了核酸分子的双螺旋结构及其在生物信息传递中的重大意义,获得1962年诺贝尔生理学或医学奖。
【66】 王斌译自J. D. Watson, F. H. C. Crick. The structure of DNA. Cold Spring Harbor Symp. Quant. Biol. 1953, 18: 123-131.
【67】 原文误为2.5%埃。——译者注
【68】 当时的认识有误,后来的研究证明G和C的配对是通过三对氢键完成的(见此处 图1)——译者注
【69】 梁宏译自L. J. Stadler. The gene. Science , 1954, 120: 811-819.
【70】 路易斯·斯塔德尔(Lewis John Stadler, 1886—1954),美国遗传学家。他一生中大部分精力用于研究各种射线对主要农作物的诱变效应及在遗传育种中的应用。为了纪念他,在他曾经工作过32年的密苏里大学每两年举办一次以他名字命名的国际遗传学研讨会(Stadler Genetics Symposium)。
【71】 梁宏译自S. Benzer. Fine structure of a genetic region in bacteriophage. Proc. Nat. Acad. Sci. U. S. A. 1955, 41: 344-354.
【72】 西摩尔·本泽(Seymour Benzer, 1921—2007),美国分子生物学家和神经学家,行为遗传学的创始人之一,他在噬菌体遗传学和果蝇行为遗传学两大领域都作出了开创性的贡献。
【73】 王斌编译自M. Grnnberg-Manago, P. J. Ortiz, S. Ochoa. Enzymatic synthesis of nucleic acidlike polynucleotides. Science , 1955, 122(3176): 907-910.
【74】 玛丽安娜·玛那哥(Marianne Grunberg-Manago, 1921— ),法国生物化学家。Arthur Kornberg和Severo Ochoa因揭示核酸(DNA和RNA)的合成机制,共同获得1959年诺贝尔生理学或医学奖,玛那哥对该项工作亦作出了重要贡献。
【75】 克分子:以克计的化合物或元素的重量,其量在数值上等于该化合物或元素的分子量。
【76】 我们Warner, R. C. 的实验,也曾指出了混合聚合物和相应的单一聚合物在电泳行为上存在着的某些差异。
【77】 在一株产碱杆菌(Alcaligenes faecalis,是由耶鲁大学G. B. Pinchot提供的)中也发现了这种酶。D. O. Brummond(未发表的实验)在一种梭状芽孢杆菌(Clostridum kluyveri)的提取液中,也发现了这种酶。
【78】 王斌译自H. Fraenkel-Conrat and B. Singer. Virus reconstitution. Ⅱ. Combination of protein and nucleic acid from different strains. Biochem. Biophys. Acta. 1957, 24: 540-548.
【79】 海因茨·康拉特(Heinz Ludwig Fraenkel-Conrat, 1910—1999),美国生物化学家、病毒学家。他最引人注目的研究是对烟草花叶病毒TMV的研究,并发现TMV病毒的繁殖是由位于病毒核心中的RNA控制的。1955年,他和生物物理学家用纯化的RNA和外壳蛋白建成有功能的RNA颗粒。1960年,公布了该病毒外壳蛋白的158个氨基酸全序列。
【80】 与HR和TMV类似的病毒,在粘毛烟草(N. glutinosa)上都形成局部病斑,但大小明显不同。根据在另一类烟草(N. sylvestris)上得到的反应类型来看,在分化上,TMV和M属一类,HR和YA属另一类。
【81】 TMV蛋白质(1毫克/毫升)吸收光谱形状与有助于它的吸收作用的各种氨基酸的混合物(每毫升中含有色氨酸30微克、酪氨酸40微克、苯丙氨酸65微克、半胱氨酸7微克、甘氨酸1毫克)的吸收光谱形状是显然不同的。但是,它们分别在波长为282和278纳米时吸光率最大值是相同的(O. D=1.25)。当用67%的醋酸制备蛋白质溶液和各种氨基酸混合物溶液时,那么它们的O. D值是相同的,而且吸收光谱形状也非常相似(吸收最大值分别在279和278纳米)。
【82】 王斌译自A. Kornberg. Biologic synthesis of deoxyribonucleic acid. Science , 1960, 131(3412): 1503-1508.
【83】 阿瑟·科恩伯格(Arthur Kornberg, 1918—2007),美国生物化学家。他因揭示人类DNA合成机制,与Severo Ochoa共同获得1959年诺贝尔生理学或医学奖。
【84】 王斌译自F. H. C. Crick, F. R. S. Leslie Barnett, S. Brenner and R. Watts-Tonin. General nature of the genetic code for proteins. Nature . 1961, 192(4809): 1227-1232.
【85】 弗朗西斯·克里克(Francis Harry Compton Crick, 1916—2004),英国分子生物学家、物理学家、神经学家。他因与沃森在1953年发现了核酸分子的双螺旋结构及其在生物信息传递中的重大意义,与沃森、威尔金斯(Maurice Wilkins)共同获得1962年诺贝尔生理学或医学奖。
【86】 王斌译自M. W. Nirenberg, J. H. Matthaei. The dependence of cell-free protein synthesis in E. coli upon naturally occurring or synthetic polyribonucleotides. Proc. Nat. Acad. Sci. USA. 1961, 47(10): 1588-1602.
【87】 马歇尔·尼伦伯格(Marshall Warren Nirenberg, 1927—2010),美国分子生物学家。他因破译遗传密码及其在蛋白质合成中的功能,与R. W. Holley、H. G. Khorana共同获得1968年诺贝尔生理学或医学奖。
【88】 王斌译自F. Jacob, J. Monod. Genetic regulatory mechanisms in the synthesis of proteins. J. mol. Biol. 1961, 3: 318-356.
【89】 弗朗索瓦·雅各布(Francois Jacob, 1920— ),法国生物学家。因与雅克·莫诺一起发现了细胞中通过对转录的反馈抑制控制酶的合成数量,并提出了操纵子假说,与雅克·莫诺、André Lwoff共同获得1965年诺贝尔生理学或医学奖。
【90】 道理以后会清楚。重要的是考虑下述假说是否有证据,假说认为:半乳糖苷酶和乙酰基转移酶活性可能与相同的基本蛋白质单位有关。因此我们应该正视下列观察:(1)有些突变体能形成半乳糖苷酶而不形成乙酰基转移酶,或者相反;(2)纯化过的乙酰基转移酶没有半乳糖苷酶活性;(3)这两种酶的特异性是完全不同的;(4)用分步沉淀的方法很容易把这两种酶全部分离开;(5)乙酰基转移酶是高度耐热的,在同样条件下半乳糖苷酶却是很不稳定的;(6)半乳糖苷酶抗血清不能沉淀乙酰基转移酶;乙酰基转移酶抗血清也不沉淀半乳糖苷酶。
因此,对于“半乳糖苷酶和乙酰基转移酶的活性与同一种蛋白质有关”这个问题的争论是没有理由的。
【91】 某些作用于外源底物的酶受到阻遏作用的控制。大肠杆菌的碱性磷酸酶不受磷酸酯的诱导,却受正磷酸盐的阻遏。假单孢杆菌(Pseudomonas)的脲酶受铵的阻遏。
【92】 也许我们会回想起,葡萄糖和其他碳水化合物能抑制很多作用于各种底物的诱导酶的合成这一众所周知的事实(Dienert,1900;Gale,1943;Monod,1942;Cohn和Horibate,1959)。这种非特异性的“葡萄糖效应”可能与特异性代谢产物阻遏效应有关,但这种关系还不清楚(Neidhardt & Magasanik,1956a、b)。本文将不讨论葡萄糖效应。
【93】 这一发现的意义还在研究中,为了分离组成型突变体,当然必须采用选择培养基,这种程序可能有利于双重突变体,在这个过程中半乳糖苷酸和透性酶的组成型是独立发生的。因此可以从i+ z+ y- (即不产透性酶)菌株中选择i+ →i- 的突变体。分离到50株i+ z+ y- 突变体,并进一步选出“隐藏组成”型i- z+ y- ,再经y- 回复突变,得到50个i- z+ y+ 的组成型菌株。这50个菌株被证实了透性酶都是组成型的。
【94】 对5-甲基色氨酸的抗性也可能由另外的机制引起,但与我们这里讨论的问题无关。
【95】 假定,当这种突变体失去了形成其有关诱导蛋白质能力时,这种阻遏物仍有活性,那么,这种模型就可以解释乳糖系统中调节基因iS (显性)突变体的性质。
【96】 我们再重申一下,在这个系统中,不管如何系统地筛选突变体,都未能分离到任何一种类型的非多效性突变体。
【97】 必须指出的是操纵基因位点和直接相邻的结构顺反子之间作用上的差别,只是建立在下述事实基础上的,即操纵基因突变可以影响受连锁顺反子控制的几种蛋白质的合成。这不排斥操纵基因位点实际上是和它相邻的结构顺反子的一部分的可能性。假如真是这样,可以预料某些组成型的操纵基因突变,可能涉及受这个相邻的顺反子控制的一种蛋白质的结构改变。目前,尚无可靠证据足以肯定或否定这种假设。
【98】 梁宏译自M. H. F. Wilkins. Themolecular configuration of nucleic acids. Nobel Lectures Physiology or Medicine , 1942-1962: 754-782.(童克忠校。)
【99】 莫里斯·威尔金斯(Maurice Hugh Frederick Wilkins, 1916—2004),英国物理学家和分子生物学家。他因在DNA结构研究工作方面的突出贡献,与克里克、沃森共同获得1962年诺贝尔生理学或医学奖。
【100】 王斌译自S. E. Luria. Phage, colicins, and macroregulatory phenomena. Nobel Lectures Physiology or Medicine , 1963-1970: 426-436.
【101】 萨尔瓦多·卢瑞亚(Salvador Edward Luria, 1912—1991),美国微生物学家。因和Max Delbrück、Alfred Hershey在噬菌体分子生物学方面的开拓性工作,共同获得1969年诺贝尔生理学或医学奖。
【102】 近来,以温度敏感的耐性突变体得到的某些发现说明,细胞被膜的行为和敏感位点与耐性位点嵌合体的行为一样,完全取决于合成各个位点时的温度[36 ] ,这些发现不能支持那种认为“大肠杆菌素作用的扩增机制是由于细菌细胞被膜整个构型变化引起”的朴素想法。
【103】 王斌译自H. M. Temin. RNA-directed DNA synthesis. Scientific American , 1972, 226(1): 25-33.
【104】 霍华德·特敏(Howard Martin Temin, 1934—1994),美国肿瘤学家。他因发现了逆转录酶,挑战了分子生物学的中心法则,与Renato Dulbecco、David Baltimore共同获得1975年诺贝尔生理学或医学奖。
