蛙类的性别与性转化
自从Pflüger1881—1882年的工作以后,大家都知道幼龄蛙的性率特别,并且在从蝌蚪到蛙的变态时期内,生殖腺往往表现中间状态。这一类的个体究竟是雌是雄,引起了很多争论。近年来证明了:这种中间类型往往变成雄性,甚至有人主张许多族的雄性都经过这一阶段。
Richard Hertwig的实验证明,推迟蛙卵的受精时间,便会大大增加雄性的比例,并且在极端情况下,甚至所有的个体都变成了雄性。把推迟受精时间同染色体的改变联系起来的企图,还没有成功。
进一步的研究证明了,由于没有认清异族的蛙在精巢和卵巢的发育上表现出了显著的差别,于是以前研究的结果便模糊起来了。Witschi指明欧洲山蛤(Rana temporaria)一般分为两类或两族。一类的精巢和卵巢直接由早期生殖腺分化出来。这一族住在山岳地带和遥远的地方。另一族生活在山谷和中欧,其成雄个体的生殖腺经过一个中间阶段,腺内有他认为是未成熟卵子的大细胞。以后另有一组新的生殖细胞取而代之,并且变化成真正的精子。这些族称为未分化族。
Swingle也找到两类或两族的美国产喧蛙,大概说来,其中一种的精巢和卵巢很早便从原生殖腺分化出来。另一些族则分化较晚。在第二类雌蛙里,原生殖腺内的大细胞以后都发育成为真正的卵子,但是雄蛙的原生殖腺,在雌蛙分化以后,还继续存在一些时候。后者的大细胞可以分化为精子。不过这些精子后来大半被吸收,而一些尚未分化的细胞却变成了真正的精子。Swingle把雄性蛙原生殖腺内的大细胞不当为卵子,而看成是雄性的精母细胞。他证明这些细胞,经过一次流产的成熟分裂以后,便大部分被破坏了。换句话说,雄蛙并不经过雌蛙阶段,不过好像是在它进行第二次成熟分裂以后、分化之前,作出了一次形成精子的不成功的尝试罢了。
无论对原生殖腺内的大细胞作什么解释,目前讨论中的要点是,外在条件中或内在条件是否可以这样地影响预定雌蛙的原生殖腺,以致它后来产生有作用的精细胞。Witschi的证据支持在中性族内有过这样的转化。
Witschi汇集了德国和瑞士各地观察者所报道的性率,如下表。表的右边一行表示雌性所占的比例。由此可见前两群(第一、第二两群)性别比率接近1∶1,后三群(第三、第四、第五三群)的雌性百分率较高,有的区域最高达到一对雌雄所生的全部个体可以完全是雌性(100%)。它们都属于中性族。
Witschi所发现的最重要的事件涉及分化族与未分化族两者性率差异的遗传。Hertwig用异族的雌性和雄性进行杂交。

杂交(1)的子代雌性,全是未分化型;杂交(2)的子代雌性,则分化很早。Witschi断言分化族的卵子,比较未分化族的卵子,有着更强的决定雌性的作用。

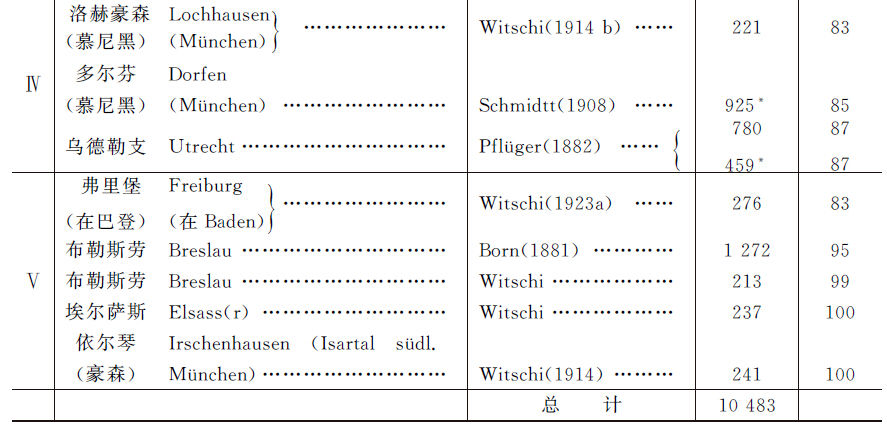
在另一个实验里,Hertwig用“决定雌性能力”(Kraft)、强弱不同的各个未分化族进行杂交。他的结论是:弱卵子同强精子受精的结果,与强卵子同弱精子受精的结果,是一致的。“同一类型的卵子或成雄精子,具有相同的遗传组合”。
蛙类的染色体成分问题已经争论了好几年了,不仅涉及究竟有多少条染色体,而且也涉及有二型配子的,究竟是雄性还是雌性。几个物种的染色体最可能的数目似乎是26条(n=13)。也有过其他数目(24、25、28)的报导,按照Witschi最近的记载:山蛤有26条染色体,包括雄性的一对大小不同的X、Y在内(图145)。如果这点得到证实,那么,雌性为XX(同型配子),而雄性为XY(异型配子)。
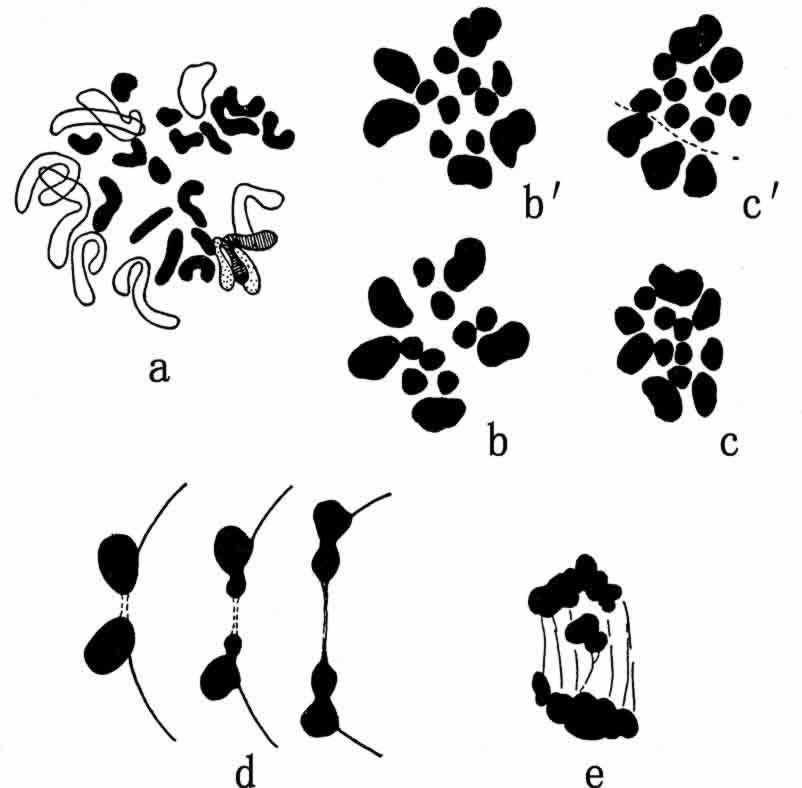
图145 山蛤(Rana temporaria)的染色体群。a.二倍型雄性染色体群;
b和b′.第一次精母细胞分裂的后期,各有13条染色体;c和c′.同前;d.第一次精母细胞X和Y在第二次精母细胞分裂中的分离(仿Witschi)
Pflüger(1882)、Richard Hertwig(1905)和以后的Kuschakewitsch(1910)已经证明过熟的卵子提高雄性的百分率。不过因为这些实验不是用同一个雄性和同一组卵子受精来进行的,所以实验结果也就可疑了。Hertwig本人指出了低温效果同过熟效果很相类似。许多胚胎是畸形的。Witschi证实了Hertwig的结果(用Irschenhausen族)。估计过熟到80~110h之久的卵子,发育成为74 ,21♀和20条中性蝌蚪(注:蝌蚪死亡率为20%,幼蛙死亡率为35%。)。
,21♀和20条中性蝌蚪(注:蝌蚪死亡率为20%,幼蛙死亡率为35%。)。
Richard Hertwig比较正常卵子和延迟卵子(相隔67h)两者的性率,结果如下:由正常受精而来的49日龄幼虫(在变态之前),有46个中性♀;由延迟受精卵而来的幼虫则有38个中性♀和39 。在大约150日龄的正常蛙当中,有些是分化型的雌性,有些在生殖腺方面是中性的雌性,有些是雄性(数目不详);从延迟受精的卵子当中,则得出45个中性♀和313个 。一岁的蛙有6♀和1 (正常受精)以及1♀和7 (延迟受精)。这里,过熟作用似乎可以促进雄性的分化,其次,则使中性个体(这里列入未分化雌性一类)转化为雄性。
卵子过熟的结果,究竟应怎样解释,还是很不明白的。表面上,这些结果似乎表明正常应该形成雌性的,可以变成雄性。这种雄蛙的精子,其性别决定的性质究竟怎样,一直还没有进行过遗传学检验。理论上,这些精子应该是同型配子。但在自然条件下,这样的个体似乎是很难生存和发挥作用的,否则过熟一定不会稀少,何以实际上没有看到正常的雄性产生100%的雌性呢?Witschi已经指出过过熟卵分裂异常,而且他所检查过的少数胚胎表现了内部的缺陷,不过这些缺陷同雌性向雄性转化之间,有什么关系,却不明了。
Witschi的实验(1914—1915)已经证明:生殖腺(或原生殖腺)尚未分化,或者雌雄同体的生殖腺尚未成熟时,这样的个体是有可能被外在因素转化成雌性的。
Ursprungtal族的蝌蚪很有可能属于未分化族。这族的蝌蚪在10℃时,有23个雄性和44个雌性;在15℃时,有131个雄性和140个雌性;在21℃时,有115个雄性和104个雌性。显然,这一族蝌蚪的性别不受温度的影响。
另一方面,Irschenhausen族的蝌蚪在20℃下饲养时,有241个未分化型雌性;在10℃下饲养的六批里,有25个雄性和438个雌性。Witschi由此得出结论:低温是一种雄性决定因子,不过不应当忽视,有许多这种所谓的雌性后来发育成了雄蛙。他以后谈到这些实验时说道:“低温使雄性变成了雌性早熟的幼龄雌雄同体,这一般也是未分化族的正常现象”。
因此,这里除了真正的雄性状态延缓以外,是否尚有别的因素,似乎还有问题。
目前只可能根据现有的证据,作出一个暂时的结论:未分化族的半数个体正常应该变成雌性,看来,它们的生殖细胞可以变成精细胞,也可以被来自另一来源并且以后变成精子的细胞所代替。换句话说,通常足够产生雄性或者雌性的那种基因间的平衡,也可以被环境因子“取消”,一个染色体平衡应该产生雌性的个体可以发生精巢。换句话说,这意味着每一只蛙能够发生精巢和卵巢;在正常情况下,XX个体只发生卵巢,而XY个体只发生精巢;但在非常条件下,XX型的雌蛙可以发生精巢。相反方向的变化是否可能,还没有证实。
有过许多关于“雌雄同体”成蛙的记载(图146)。Crew列举了40个新近的例子。这些雌雄同体的蛙是否同上面所谈到的性转化有某种关系,还不知道。也许重要的是,在上述实验中也报道了几个雌雄同体的个体。另一方面,有些雌雄同体的个体也可能有不同的起源。但是它们的附属性器不对称的很少,生殖腺组织往往不整齐地分布,所以要说它们是性染色体排除后所造成的雌雄体或嵌合体,也找不出很多证据来。其次,支持雌雄同体型的精子和卵子都是同型配子的证据如果有效,那么,把染色体的排除作为一个可能的解释,也就失去了它的根据。
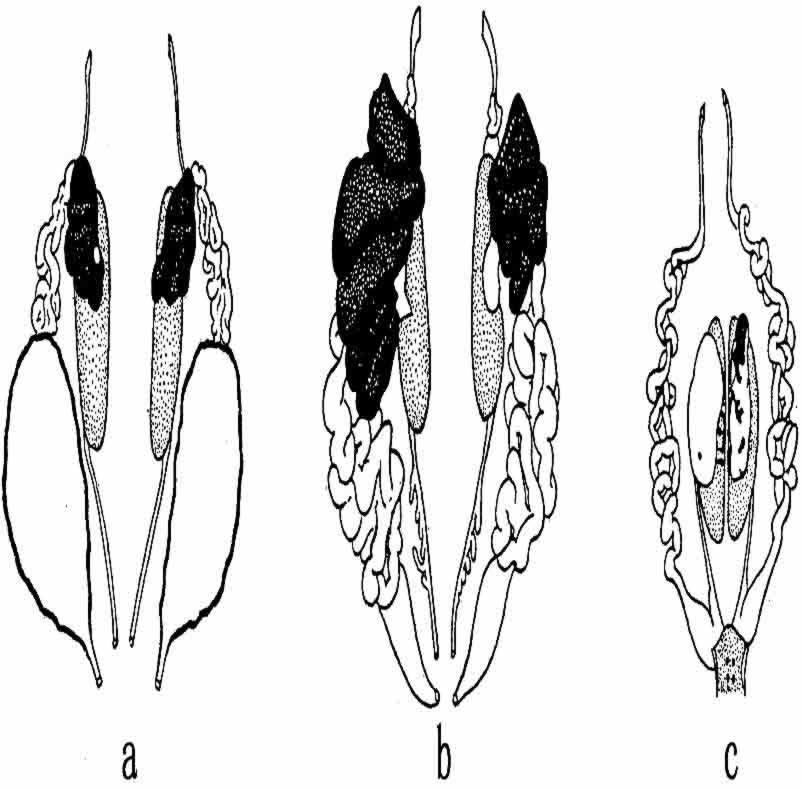
图146 三只雌雄同体型的蛙(示泌尿生殖系)(仿Crew和Witschi)
Witschi从一个雌雄同体(Hh)的个体里,成功地取出了成熟的精子和卵子,并用另一个分化族的精子和卵子来进行检验,结果如下:
(1)分化族雌性的卵子 配 雌雄同体型的精子=♀♀
(2)雌雄同体型的卵子 配 分化族雄性的精子=50%♀+50%
雌雄同体型的卵子又和同一个体的精子受精,得到45♀和一个雌雄同体的个体,例如
(3)雌雄同体型的卵子 配 雌雄同体型的精子=45♀+1雌雄同体
以上结果意味着原来的雌雄同体型雌蛙是XX。每个成熟卵子有一条X。同样,每个有作用的精子一定也有一条X。这样就不可避免地得出下面任一个结果,或者是每条精子有一条X,或者是一半精子有X,另一半精子没有X,不过后一种精子在母体内死亡了(即从来没有过作用)(注:Crew(1924)报道:用雌雄同体型的精子同正常雌蛙的卵子受精,获得成功。各个蝌蚪的生殖腺都是直接发育出来。发育过程达到可以判明性别的一切子代(774),都是雌性。母蛙可以被看成是一个真正的XX型雌型,其所产生的卵子和精子各有一条X染色体。
Witschi(1928)把七周龄的Rana sylvatica雌性蝌蚪放在32℃下面,使所有雌性的卵巢都转化成了精巢(内有原精细胞)。雄性没有变化。(1928年添注)。)。
