第二十讲
大脑两半球的病态,当做受手术作用的结果:丙、光分析器活动的障碍;丁、皮肤机械性分析器活动的障碍;戊、大脑额叶摘除后的障碍;己、皮肤温度性分析器的障碍;庚、梨状回转摘除后的障碍;辛、运动分析器的障碍
诸位!现在讨论光分析器的活动。虽然我们有关这分析器活动的研究是比较不多的,但是生理学内现有的材料使我们可能在大脑两半球内各有关部分被摘除的场合,发现光分析器的活动与声音分析器的活动具有很类似的关系。
从高儿兹关于除去大脑两半球的狗的实验而又可得结论:光线所引起的、最一般的方位判定的运动反应也是属于皮肤下核的。说明不过就仅是如此。在大脑两半球被摘除的狗的场合,没有其他著者指明任何其他光性机能的存在。在我们的一只实验狗(泽廖尼实验)的场合,上述的这种初步性反应也不曾显然地发生过。这就意味着,我们有理由地把我们实验动物的全部其他视觉性活动,可以认为是只属于大脑两半球的东西,这是与口腔分析器和声音分析器的活动相类似的。而在口腔分析器和声音分析器的方面,这个结论是具有充足的实验基础的。口腔分析器在大脑两半球摘除以后虽然还保持着分析性的机能(我们除去大脑两半球的狗不肯吃进许多不能吃的东西),可是不能形成一时性的条件性联系,那么,由于皮质下核而形成对光线刺激的条件反射的可能性,当然是更不可想象的。我们和以前的其他大脑两半球的研究者[包括明可夫斯基(М.А.Минковский)]都看见了(这必须当做确定的事实看待),在大脑两半球后头叶(枕叶)的一侧或两侧受了一定损伤的场合,与此相当地,会在一侧或两侧视野内有一定的狭窄,或者是侧方的狭窄,或者是上方或下方的狭窄。这样,进入于视野残存部分的事物会对动物引起相当的作用,但是如果外方的事物稍与视野残存部分不相配合,该动物就依然不发生通常的反应。
从这个事实必须作如下的一个结论,就是在后头叶(枕叶)内有光分析器核心的存在,使最复杂的复合性光性刺激的存在成为可能,所以也使复合性光性刺激的分析成为可能。我们的相当多数的狗,在其两侧大脑后头叶被摘除以后,不管它们在手术后活着多么长久(其中的一只在手术后活了3年),可是从来不曾显出辨别物体(即对象性视觉)的视力。人、其他的动物、食物都不是这些狗的光分析器所能辨别的。我们好几次把食物小片撒放在地面上,或者用线把食物小片以种种的高度挂起来,可是不管这些狗怎样饥饿,没有一只除去大脑后头叶的狗会因为光的刺激而做向食物方面的运动;只是利用气味刺激或皮肤机械性刺激,它们才能辨别食物所在的方位。在大脑后头叶所受损伤各不相同的场合,视野狭窄化的情形也各不相同。由于这个事实不能不结论地说,在对象视觉丧失的场合,事情的本质是在于光性刺激有关的最高综合力及分析力的缺乏,而不是在于视力辅助机能的调节作用及适应作用(аккомодацияиадаптация)的障碍。我们除去大脑后头叶的各实验狗,都是同样地不能辨别事物,大的或小的,在远或近距离的,很强地、中等地、很弱地被照亮的事物都是同样地不能辨别的。这样看来,光性分析器的中枢、核心,即光性刺激最高综合的及分析的器官,是存在于大脑两半球的后头叶内的。然而这并不是光性分析器的全部。这分析器是远远广布的,可能广布到大脑全部的实质之中。在大脑两半球生理学的成绩辉煌的时代(19世纪70年代),若干的研究者已经树立了一个假定,就是大脑两半球前头部也与视力有关系,可是并没有积极性的事实能做充足的证明。当时只能将大脑两半球机能讲义第二十讲 一切归纳于远方的制止性的作用。然而现在,我们却能举出积极性的事实而主张,甚至于具有显著的机能的光性分析器,实在是存在于大脑两半球的前半部之内的,其限界线是从S状回转(g.sygmoidei)直接的上后方开始,斜行向后地达到雪儿维氏回转(g.sylviaticius)的前角,以后沿着雪儿维氏裂(fissura fossae Sylvii)而直到大脑两半球的底部。我们摘除大脑后头叶的全部狗(卡立谢的实验也与此相同)都对于室内光线容易形成条件反射,并且对于室内光线相当精微的浓淡也能形成很明了的分化相。这个事实对于蒙克所谓精神性盲给予了一个简单而有自然科学根据的说明。在大脑两后头叶被摘除以后,已受显著损伤的光分析器,只能与这光的极基本的机能,即只能对于光刺激强度动摇的反应,形成条件性的联系。因为如此,在被照亮的空间里,狗能够回避在阴影中的物体而向开放的门、光亮的地方走去。因为这样,如果心理学地说,就也许应该说,狗是能懂的,可是懂得不多,不能充分地看见。可是,当然这样方式的说明是多余的。显然,事情的本质是在于分析性机能的受了限制。这些事实以后的继续研究,完全证实了,对于事物的这样纯粹的客观的观点是科学地有益的。我们的某一只狗,也只保存上述限界线内的大脑两半球前头部,可是它也能以光分析器比较高级的机能而形成条件反射。这就是上述的那一只手术后还活了3年的狗,并且在上一讲里,这只狗是当做复合声音刺激的分析力永远丧失的例子而被引用过的。我要详细地说明用它所做的光性刺激的各实验(库得林实验)。两侧大脑半球的手术是相隔一个月而施行的。第二次的手术在1909年5月5日施行。在手术以前,在黑暗无光的室内用一百支光的电灯开亮形成了食物反射,在第二次手术后的第五天,这个反射就明了地恢复了,而直到了第11天,反射量甚至于反而超过手术以前。在这以后,这个反射不再被应用,而声音性刺激的研究却继续下去。在同年的9月7日,开始用暗室内影幕上照亮的十字形的运动(用幻灯将十字反映)形成了条件反射。当然,反射的形成是迅速的,并且在1周后是显著的。从9月28日起,放映的这十字形是在影幕上不动的。反射继续存在,不过反射量却若干减少了。现在着手于同等大小的同样照亮的圆形与十字形的分化,就是说,十字形的出现并用食物强化手续,而圆形的出现却不并用食物。从圆形的第七次应用,开始发现它的分化相,可是这实验以后中辍了,只在半年以后才又开始做。十字形的反射依然是残存的。分化相的出现是迅速的,并且不久就成为完全的分化相。这些实验的最后结果如下。

大脑手术后过了三年,施行这只狗的解剖,证实了手术的完全性,因为大脑两半球的后半部没有任何残留的踪迹。
不能怀疑的事实是,这只狗大脑两半球前半部内残存的光性分析器的一部分,不仅对于光亮强度的摇动,而也对于照亮的和遮暗的各种事物各不相同的形态,能够形成条件刺激物。并且同时如上所述地,在这只狗和其他的各狗后头叶被摘除的场合,直到它们生存的最后,都不能发现对于各个别事物有条件反射的形成。如果注意,只在这只狗手术后经过了4个月,才能最初检验了它对各种形态的条件反射的形成,这就意味着,这些条件反射的形成,可能是发生更早的,可是再过了两年半,辨别物体的视力还是不曾出现。因此就不能不结论地说,在手术后的第三年,光性分析器的状态是最后的、不可恢复的状态。同时也许还应该注意于如下的事实。究竟为什么,我们的这只实验狗既然在实验架台上能够辨别形态,却不能在不受拘束而自由的地区辨别事物的形态呢?但是在实验架台上的环境内,和在自由地位于各不相同的许多事物之中,这两种情形有很大的区别。在动物不受拘束而行走的时候,动物的前面有着极多事物的形态,并且这些形态对于动物的眼睛是不断地或多或少地变化着的,因为若干物体本来就是在运动着,而动物的本身对于一切物体也是运动着的。不仅如此,由于若干事物的移动和动物本身对各事物的相对性移动,光亮是动摇不定的,所以各种形态境界部的浓度也不是保持一定不变的。如果与通常的现实各条件相比较,在我们实验的架台上,一切情形就完全不同,是非常简单化了的。似乎是,也许需要很长时期的、渐渐的练习,辨别形态的残存机能才会对于动物具有生活上的实际意义。
根据上述的资料,光性分析器和声音性分析器已损害的机能各最重要的阶段,可以认为在一定的程度上是彼此相当的。光性分析器内的小障碍是正常视野的狭窄化,而声音性分析器内的最小障碍,如果根据我们唯一实验,是复合音中各个别音的消除。光性分析器比较更显著的障碍是物体辨别力的缺乏,就是说,形态、明暗、色彩(个别的狗往往具有色的兴奋性)等等复合物的辨别力的缺乏。换句话说,光性刺激物最高的综合力与分析力都消失了。声音性分析器同样的比较更显著的障碍,显然是各种声音复合物的辨别力的丧失,譬如言语辨别力的丧失,这又意味着声音性刺激最高综合力及分析力的丧失。姑且暂不论及两分析器的完全损坏,这两种分析器最大障碍的表现就是只有各别地有关于光性刺激和声音性刺激的强度的区别。在机能障碍的极大极小的各阶段之间还有一些场合是这样的,就是在光性分析器内光的各种强度上,还可以辨别由许多发亮点而成立的各种不同的组群,而在声音性分析器方面,却能辨别各种不同的音响,譬如击声、杂音及各种不同的音。
在我们的前面还有一个问题:在大脑实质内皮肤机械性分析器的分配是否与上述的光性及声音性分析器在大脑实质内的情形相同呢,就是说,除有关这类机能的高级特殊的区域以外,关于比较非高级的机能,有没有比较更广泛地散在的皮肤机械性感受器成分的存在呢?虽然还不能作最后的答复,可是根据我们的材料,我们倾向于肯定性意义的答复。现在我们举出与此有关的、已有的和现在的多数实验材料,这在上述问题以外也还是有兴趣的。老早[替霍密洛夫(Тихомиров)实验]我们已经看见,大脑两半球前部的摘除会引起条件性皮肤机械性反射的消失,而其他分析器有关的各动因的反射却是同时并不丧失。最近的实验确实资料证明了(克拉斯诺高尔斯基实验),运动区域是与皮肤机械性分析器的特殊区域多少相隔离的,并且大脑内的这个领域的各一定部位是与皮肤各个别部位相一致的。举一个例子如下:一只狗,除其他分析器一些动因的条件反射以外,具有一个泛化及于全皮肤表面的条件性皮肤机械性的酸反射。现在摘除了这只狗左侧冠状回转(gg.coronarius)及外雪儿维氏回转(ectosylvius)。在手术后的第四天,其他各分析器的条件反射都已经出现。条件性皮肤机械性反射在第八天出现,不过只在左侧出现,并且迅速地达到正常量。在第十天,右侧的皮肤机械性反射也出现,但只出现于躯干的中部。在前腿、后腿、躯干的肩、腰各部,这反射是完全缺乏的,并且有这条件反射的部位与无这反射的部位两者间的境界是非常显然的。这反射出缺乏继续到手术后的第90天,以后由上向下顺次地开始恢复。在这实验的场合,除本分析器部位与明可夫斯基所指明的部位相一致以外,富于特殊兴趣的是如下的。上述皮肤部位的条件反射既然丧失了阳性作用,同时这些部位还显出强烈的制止性作用(在有关睡眠的一讲内这是已经指明过的)。制止性作用的表现是这样的,就是这些皮肤部位的机械性刺激似乎是不具有作用的,但如果与皮肤其他部位具有阳性效果的刺激相复合,或与其他分析器具有阳性效果的刺激相复合,就会使后者两种阳性效果减弱或消失。此外,上述部位的重复刺激,尤其如果是较长时间的刺激,每次都很快地引起动物的瞌睡或完全睡眠,而动物以前在实验架台上不仅从来不曾睡着过,而且也不会瞌睡过。睡眠只发生于这些刺激的时候。在同一的环境里,如果没有这一类的刺激,动物就保持活泼的状态。在比较不久以前,这样的实验也重复地做过,结果一般地也是相同的(拉仁可夫实验)。在对这只狗因大脑手术而受损害的皮肤部位(与克拉斯诺高尔斯基的实验相同,脑皮质的同一回转部位受了手术的损伤,不过是一部分的损伤)应用刺激以前,其他各分析器的反射部是存在的。但是在有障碍的皮肤部位一受了刺激以后,动物就在该实验的其余的全部时间以内会变成瞌睡,并且其他一切的条件反射都成为零。于是就发生值得研究的有趣的问题:没有任何方法可以发动这刺激的阳性作用的成分吗?阳性作用的成分出现了(也是拉仁可夫的实验)。直到当时,孤立的条件刺激的时间,就是说,在与无条件刺激物相结合以前的时间,是继续30秒钟的。在新的变式实验的开始,皮肤无作用部位的孤立的条件刺激,每次只应用5秒钟,而在应用数次以后,到了实验的最后,才又与寻常相同地继续30秒钟。在这些实验条件之下,也能够发现了这刺激的阳性作用。这样的阳性效果出现得很快,并且效果不很大,而最重要的是,这阳性效果在刺激的时间以内就会消失,可是在其他各刺激物的场合,这样阳性效果通常在孤立刺激的最后是会增强的。与此有关的实验如下。
这只狗的各阳性条件刺激物是:拍节机响声,哨笛音,电灯开亮及皮肤机械性刺激。在这实验里,对于手术后成为无作用的前腿一个皮肤点施行了刺激。唾液分泌量按照测量管的划度计算,5个划度与1滴量相当。这只狗的唾液分泌总是不多的。在手术以后,不曾发现这只狗的痉挛发作。
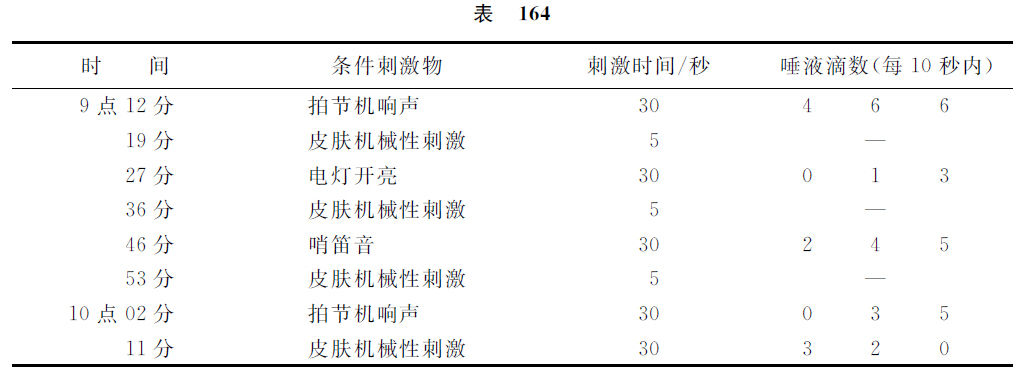
在对于通常不发生效力的皮肤部位给予刺激的场合,也可以利用若干其他的处理方法而获得同样小量的阳性作用,就是利用正性诱导、解除制止法及咖啡因溶液的注射等等。显然,与以前有关其他事例的说明相同的,这是最大的兴奋性薄弱的现象。
在我们全部的实验里,包括最近的实验(菲耀道洛夫实验),通常不发生效力的皮肤部位,迟早与时间的经过并行地,会又恢复原有的正常机能,这是与以前各实验报告者的声明相同的。在我们面前发生一个问题,也就是其他各研究者所提出的问题:这样机能的恢复,是怎样地,与什么东西有关系地而发生呢?当然,除联系皮肤各部位与大脑的两侧交叉性通路以外,起先可以想及的是直接通路。因此,对于若干狗,完全除去了一侧的大脑半球,检查了相对侧皮肤机械性条件反射,直到实验动物死亡为止(若干的实验动物在手术后活了一年以上,并且没有痉挛的发作)。最近的实验是用4只狗做的。这四只狗的大部分具有食物条件反射,一部分具有对于酸注入的防御性反射,及与脑手术相对一侧皮肤应用弱电流的防御反射[弗尔西柯夫(Фурсиков)与贝可夫(Быков)的实验]。虽然做了各种变式的实验,所获得的结果在一切的场合都是否定的。为了兴奋性的提高,应用了番木鳖素和咖啡因的注射(弗尔西柯夫),并且用种种的方式,检查受了损伤的身体一侧皮肤刺激的可能的制止性作用(贝可夫)。后者的试验方法坚持地被应用了多次,这是因为在大脑半球一部分障碍的场合有关于皮肤分析器的上述资料的缘故。制止性作用对于各种不同的阳性条件反射并不曾出现,并且在健康的体侧各皮肤部位,在后继地应用场合和同时应用的场合,情形也是如此。完全相同地,对于受损伤一侧的皮肤所给予的刺激,也没有任何催眠性作用的出现。当然受伤体侧的这样刺激,即使在应用上述若干处理方法的场合,却也不能变为阳性,而在皮肤分析器一部分被除去的场合,上述这些方法的应用,会使无效力的皮肤机械性刺激成为阳性。所以那些刺激(注:受损伤部位的刺激)在条件反射性活动的关系上,完全是无关性的刺激;换句话说,按照我们现在实验的资料判断,身体一侧的皮肤与同一侧大脑半球的直接联系是不存在的。
这样,在反射一时性消失以后的恢复方面,直接通路(即不交叉的通路)具有关系的可能性是被排除了。那么,只剩了一个可能性,就是受了手术损伤的部位的活动为该同侧大脑半球内的残存部分所代替的可能性,这是其他各研究者们所假定过的和试验过的事情。所以我们先尽可能地除去了一侧大脑半球前部的各脑回转。可是起初很长久地消失了的条件性皮肤机械性反射,终于与时共进地逐渐恢复了。但是现在,还可能想象与第一次手术部位相接近的各部位具有代偿性的能力。然而这些与第一次手术相接近的部位再补充地受了手术的处置而破坏(尤尔曼实验),以后,分析器已经恢复的机能却不受任何影响。所以,代偿作用是由于远在的神经成分而发生的。在事情获得这种结果以后,第一个成为显著的问题,是我们皮肤机械性刺激应用方法的是否毫无缺点,该刺激的声音性的成分可能也使这情形变成复杂了。于是就作成了一种皮肤机械性的刺激器,其作用至少对于我们人类的耳器官是毫无声音的。可是没有对照试验,这也还是不够的。所以我们在刺激器与皮肤之间,插进了一种媒介物,这是必定能够除去刺激器的皮肤机械性刺激的,但是决不能除去刺激器的可能有的声音成分。在这样条件下检查该刺激器作用的场合,反射不发生。所以,在刺激器寻常被应用的场合,反射实在是由于刺激器对狗皮肤的刺激,而并非由于对狗耳的刺激所引起的。第二,我们按照与光性及声音性分析器的类似性而不能不假定,在第一次手术后所残存的皮肤分析器的部分与最初被除去的部分相较,不过只具有很有限的机能。我们打算应用复合性皮肤机械性刺激而加以检查,就是在对正常皮肤用向各种方位进行的皮肤机械性刺激形成了分化相以后,施行第一次手术,以后再检查一时地丧失反射性机能的各个皮肤部位。这样的实验不过是正在进行之中。但是在有关部位摘除后,皮肤反射恢复的各实验结果是与我们以前的这个同样实验不一致的,所以我们继续再做这些实验。
除有关皮肤机械性分析器的上述资料以外,我们还有一些实验,阐明这些分析器两侧彼此间的关系。如以前曾经提及过的,对于我们许多工作同人(克拉斯诺高尔斯基、安烈勃及其他诸人),出现了一个很显著的事实,就是在身体一侧各皮肤部位所形成的条件性皮肤机械性刺激,即刻正确可惊异地、对称地在身体另一侧的相同各部位也会自动地照样地发生。关于阳性和阴性条件反射,情形都是这样的。根据这个事实,不能不期望着,身体两侧相对称皮肤部位的分化必定是一个比较困难的过程,而事实也是如此(贝可夫,柏德可琶叶夫及格里高洛维区的实验)。这个意外的事实是怎样发生的呢?当然可以想及大脑联合的联系(комиссуральнаясвязь)。并且在实际上,胼胝体(corpus callosum)的切断使这个事实不能成立了。在胼胝体切断以后,两侧的条件性皮肤机械性反射就成为完全彼此无关的了(贝可夫实验)。所应用的条件性皮肤机械性反射是:食物性反射,对于口腔内酸的注入及对于皮肤弱电流的应用所形成的防御性反射。所用的电流强度是这样的,就是仅能引起动物腿部的急屈和探索反射,但不致引起动物的吠唤及与刺激器的斗争。所做的实验是采取很多不同方式的,但结果都是相同。已形成的各条件反射只存在于身体的一侧。在身体的另一侧,不能不重新特别地施行形成反射的手续。在身体两侧相对称的各部位,无困难地形成了相对立的反射。
举例如下。这只狗具有食物性反射。在右侧后上腿受刺激的场合,有阳性反射,而右侧肩部受刺激的场合,有阴性反射。左侧的反射正与此相反。

全部的4个反射都是不能不个别地形成的。完全与此相同的是身体一侧一切新异的皮肤刺激(50摄氏度,弱电流等在食物反射及酸反射的场合),能引起探索反射,只制止身体同一侧的各条件性皮肤机械性反射,但不影响于身体另一侧的相同的各反射。这些实验是用3只动物做的。
与皮肤分析器的特殊实验并行地,可以提及只除去大脑两侧额叶的实验(巴勃金实验)。额叶的摘除是沿着两侧前十字沟(sulcus praecruciatus)及前雪儿维氏沟(sulcus praesylvius)直达脑底的线上施行的,所以嗅叶(lobi olfactorii)也同时损坏了。有时手术刀,并且几乎一定常有的手术后的病变,都涉及位于比较后方的各脑回转,这是在解剖以后所证明的。这样实验是对于4只狗施行的。在这全部的四个实验例,在手术以后,眼的与耳的各条件反射都或迟或早地恢复了或形成了。可是机能障碍,尤其显著的机能障碍,只发生于皮肤分析器及运动分析器。在动物的生前(手术后动物活着1~6个月,最后因痉挛而死亡),大部分是阳性条件性皮肤机械性反射不能形成的,尤其躯干部的皮肤刺激是这样的。有时在四肢上却能形成阳性皮肤机械性反射。以条件制止物的形式,各阴性反射是可能在皮肤的任何部位形成的。同时,几乎必定存在的是皮肤表面兴奋性的普遍增强,于是若干狗在实验的时候不能忍受实验架台上的足绳,只在除去足绳以后,它们才会安静地站在实验架台上。我们也观察过动物的一时性运动紊乱症,其表现是异常的姿势(头部下垂,背部向上方弯曲),四肢有麻痹现象及痉挛。特别强烈的是口部运动的障碍。在手术后不久的时期内,各狗摄取坚硬的食物特别地艰难,所以不能不用手喂给它们吃。除上述的这些情形以外,在动物的行动方面,并没有什么特殊的现象。
从皮肤温度性分析器的一些动因的条件反射的少数实验而言,似乎可以结论说,这个分析器与皮肤机械性分析器在大脑皮质内的部位并不完全互相一致。在摘除前十字脑回转(g.praecruciatius)的场合(仕序洛实验),后肢的条件性皮层机械性反射恢复很快(大约1周),而后肢的条件性温度反射(寒冷刺激及45摄氏度的刺激)的恢复是相对地很慢出现的,需要了4周。
利用条件反射的方法,我们打算获得若干研究者所指示过的梨状回转(g.pyriformis)对于气味分析器有关的资料[惹华德斯基(Завадский)实验]。对于几只动物,预先确立了多数的运动性和分泌性的条件反射,而且这些反射是无条件性的和条件性的两方都有的,尤其条件性的反射却是人工的和天然的条件反射都有的。在两侧的梨状回转及海马角(cornus ammonis)完全被除去以后,恰恰最早出现的是气味性反射。在手术后的第二天至第三天,对于气味的鼻翼运动就出现了。在第三天和第四天,实验狗能够只利用鼻子而从包有肉和香肠的纸包无错误地取出内容物。从第六天起,出现了对于肉粉的唾液反射。对于樟脑气味的人工性食物反射在手术后的第14天起就比其余的皮肤、耳、眼各方面的条件反射更早地、显著地出现。人工性气味性反射在手术后第一次试验的时候就已经出现,就是说,这是反射的恢复,而不是新的形成。
最后,我们自己又提出一个有关大脑两半球皮质的所谓运动区域的性质的问题。这个运动区域是什么?大脑皮质的其他各区域都是从外方向有机体进行的各种刺激的分析器;与此相同地,在有机体做复杂精微工作的时候,大脑皮质运动区域是不是与有机体的骨骼运动性装置有关的各刺激的分析器呢?抑或这运动区域是与其他一切的效验性的脑皮质区域具有区别呢?换句话说,在生理学机能上,是不是这皮质运动区域与脊髓的前半部或后半部相当呢?如众所周知的,这个问题是这样古老的,就是与脑皮质运动区域被确定的事实本身同样古老的,并且直到现在,有不少的人主张,这问题的解决是应该依据上述第一个解释的。我们打算利用条件反射的方法获得更多有关这问题解决的资料。我们利用一定的运动作为条件刺激物,并且实验地确定脑皮质内与该运动有关的区域。我们的实验是如下进行的(克拉斯诺高尔斯基实验)。因为与这些实验有关的是与大脑两半球一般构造有关的一个重要问题,并且因为从实验的进行而言,这些实验与其他的一切条件反射的实验相较,是相当复杂的,所以我认为自己有理由尽可能地详细说明这些实验,并且需要充分地利用实验记录而加以说明。
条件性食物刺激物是胫跗关节和跖趾关节的屈曲运动。屈曲运动是如下形成的。一侧后肢的大腿及小腿都用装拆自由的石膏绷带加以固定,而这石膏装置是固定于一个金属的支柱之上的。该金属支柱用螺丝钉被拧紧在实验架台上。胫跗关节的运动就是利用这个设备的。如果要利用跖趾关节的运动,也可以固定跖趾于特殊的金属支柱上。这部分的屈曲运动可以用器械或用手施行。
在左后肢的胫跗关节的屈曲运动形成反射的时候,就试验右侧同关节反射的有无。在右侧,这反射也自然形成了,正如我们在皮肤条件性刺激的场合所见的情形一样。其次,我们着手于左后肢跖趾关节从胫跗关节屈曲运动的分化,其时只在跖趾关节屈曲的时候使动物摄取食物。在跖趾关节屈曲运动受过42次强化处理以后,分化就成功了,在胫附关节不受无条件刺激物的强化手续而屈曲74次以后就也形成了分化,并且分化自然地在右下腿也出现了。当然,这还不是我们所需要的事情。在屈曲运动的时候,皮肤机械性刺激必定也同时发生,这也是能单独地产生条件的效力的。必须将这可能的皮肤机械性刺激与屈曲运动互相分化。所以应用了一切可能方式的皮肤机械性刺激:皮肤的接触,压迫,提捏及关节一侧节奏性的牵伸和关节另一侧的皮肤叠皱等等刺激(因为这是大约与屈曲运动时发生的情形相等的)。最后的这个处理方法具有最强的作用。只重复地应用这种皮肤刺激的方法,同时却不并用食物强化的手续和屈曲运动,终于这些刺激方法都成为完全无效了。但屈曲运动单独的作用却是继续存在的。然而这还不能确证唯一屈曲运动的本身必定是条件刺激物的事实。因为可能的是,我们虽然应用了种种方式的机械性刺激,但还不能完全作成与屈曲运动时相同的皮肤刺激。必须还有另一个比较地无可非难的证据,证明屈曲运动的本身实在是条件刺激物。我们想这样地获得这个证明,就是除去皮层分析器某一部分,才可以完全除去皮肤性的成分,而我们已经知道,与后肢皮肤性刺激有关的分析器就是冠状回转(gg.coronarius)与外雪儿维氏回转(ectosylvius)。准备好先形成了后肢5个部位的皮肤机械性刺激的条件性食物反射,并且以每秒钟500次的振动音也形成了条件性食物反射。在左侧的大脑该部分手术后的第七天,音的条件反射第一次恢复了。在手术后的第八天,右侧屈曲的第一次试验,还是不显作用。但是在同一天的实验内,右侧屈曲的第二次试验已经在30秒内获得了2滴唾液,在第十天获得了3滴。但是在这第十天,同侧下肢5个皮肤机械性刺激器同时试验的结果,还不曾引起唾液的分泌。在第12天,屈曲的反射量是5滴,而5个皮肤机械性刺激器的刺激、关节皮肤的牵伸及叠皱,依然都是零的结果。在第13天,关节部皮肤的牵伸和叠皱制止了同时应用的音刺激;就是说,皮肤机械性刺激的作用不是阳性的,而是制止性的。在第15天及第16天,左侧后肢已经完全不受脑手术的制止性影响,而该后肢的单独的皮肤机械性刺激,引起了显著的唾液分泌效力,但是右侧跖趾关节皮肤的机械性刺激和同时该部皮肤的牵伸及叠皱处置却不引起唾液的分泌。所以在右肢的方面继续地受着手术的影响,右肢的皮肤机械性刺激是制止性的,而不是阳性的。然而屈曲运动却是必定并有唾液的分泌。举出这些实验中的若干记录如下。
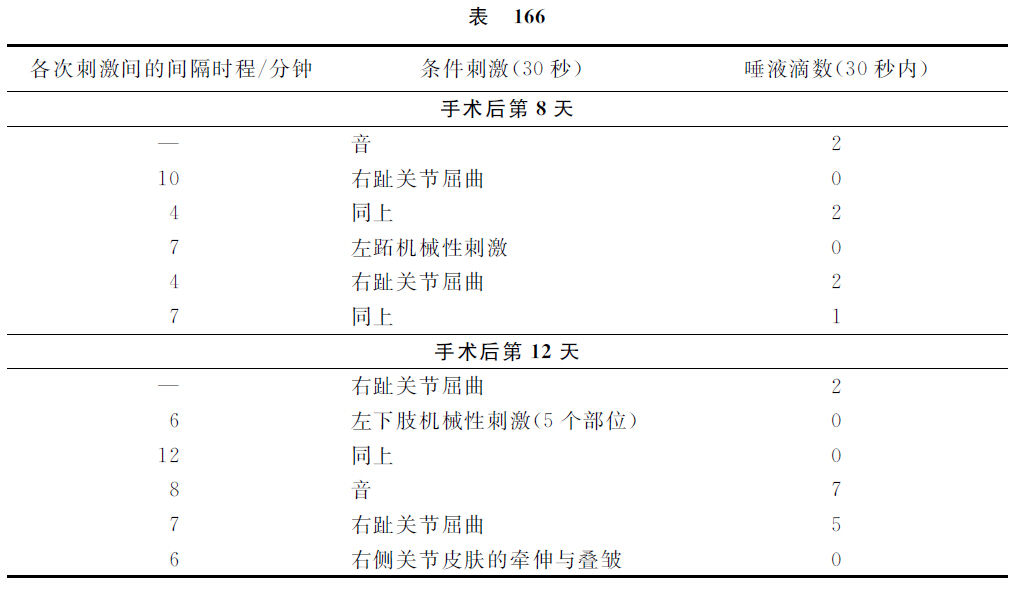
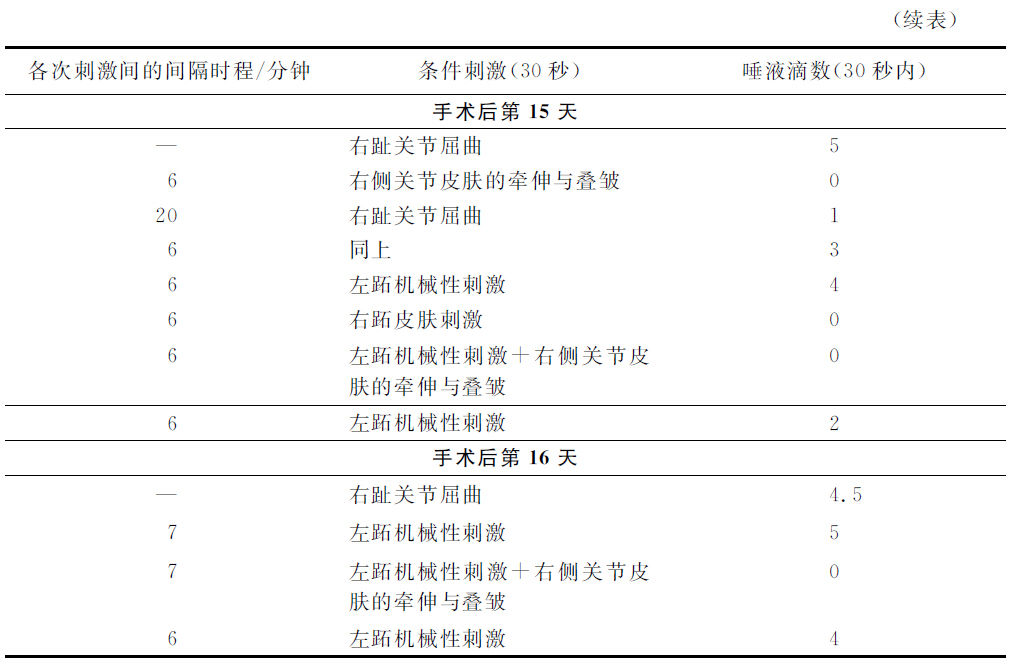
这些实验使我们有理由做两个结论。第一,仅仅运动性的动作而无皮肤刺激成分的参与,就可能成为条件刺激物。第二,运动性动作的刺激与皮肤机械性刺激在大脑皮质内各有不同的作用部位。我们知道大脑皮质内皮肤机械性的区域,究竟大脑皮质内的运动性区域却在什么地方?另一只狗的实验给了答复。
我们的一只狗的右侧S状回转(g.sygmoideus dexter)在2个月有余以前被完全除去了。在这手术以后,这只狗左侧前后两肢总是有着强烈的运动紊乱。左侧S状回转的损伤是很微弱的,右侧的前后两肢没有任何运动机能障碍的表现。全身皮肤表面的机械性反射是正常的。对于右侧,着手于趾关节屈曲运动的条件性食物反射的形成,这个反射迅速地成立了。于是开始使这反射与皮肤刺激成分分化开来,所应用的方法就是如上述地对这关节使用各种变式的皮肤机械性刺激,当然此时不并用无条件反射。一个月以后分化相是形成了,不过不一定是完全的。该分化相的例子如下。

左侧趾关节皮肤的牵伸与叠皱刺激也是因为上述的关系而完全无效的。同时左侧对于屈曲的运动也没有反射的发生。在屈曲运动受了强化手续的时候,随后地,仅仅皮肤机械性刺激也获得了分泌唾液的作用。不能不这样地解释这个实验,就是左侧的屈曲运动的本身并不具有条件性的作用,可是这屈曲运动的强化手续,使在屈曲时必然并存的皮肤机械性的成分解除制止化了。并且以后,虽然顽强地继续施行分化的手续,可是不曾能够与皮肤刺激无关而个别地获得屈曲运动的条件反射。只是皮肤刺激的效果一消失,屈曲运动的反射也就消失。在应用食物强化的场合,皮肤刺激的反射就会恢复。在右肢方面,关节屈曲运动与皮肤刺激之间,分化相总是完全明了地成立的。举例如下。

在这个实验内,每次各侧的屈曲运动都并用了食物强化手续,而皮肤刺激却不曾并用食物的强化。
这些实验昭示着,在骨骼运动性系统工作时,S状回转是来自这系统的刺激所趋向的皮质区域。可惜,以后我们不曾继续这些实验,也不曾进行种种方式的实验。当然,这些实验还需要重复地做,需要进一步证实。如果根据已做的这些实验说,就应该做如下的结论,即大脑皮质的运动性领域是生物个体骨骼肌肉活动能的分析器,这与大脑皮质其他某些领域是对于生物个体发生作用的各种外来能(внешняяэнергия)的分析器相似的。在这个见解上,大脑两半球是生物个体与外界和内界有关的一个壮丽的分析器。当然,关于骨骼运动机能所不能不做的假定,有理由地,可以更广泛地适用于生物个体的其他更巨大的活动,纵然不适用于其全部的活动。只是在假定一个生物全体活动有关的大脑皮质分析器的存在的场合,纵然这部分通常还是不够解析说明的、不够确定的,可是这样才可以从生理学的立场,把暗示具有巨大作用的许多异常的事例加以解释,譬如假想性妊娠(мнимаябеременность)就是。
