第九讲
在大脑两半球皮质内神经过程的扩展与集中:甲、在个别的分析器(皮肤分析器与声音分析器)内的制止过程的扩展与集中
诸位!直到现在,可以说,我们主要地研究了大脑两半球的外在活动,而这种活动是树立生物个体与外界两者间最复杂的、最微妙的关系的。第一,大脑两半球从自然界的无数动因里,只把自然界里比较不多的一些动因对生物个体变成信号,而这些信号化了的少数动因,有些是对个体直接有益的,有些是有害的,都使该个体发生相当的反应。以后如果这些具有条件性作用的各动因不能适应于现实的关系,就不断地受着大脑两半球的矫正,在一定条件之下,永远地或者一时地丧失它们的作用。最后,与不断地、复杂地动摇着的自然界相调和着,成为条件刺激物的这些动因受了大脑两半球的作用,或者被分解而对于生物个体成为极微细的成分(被分析),或者互相融合而成为种种的复合刺激物(被结合)。
现在我们注意于大脑两半球机能的内在机制吧。在大脑两半球发挥上述机能的场合,基本的两种神经过程,即兴奋与制止的两种过程,是怎样进展呢?
第一,此处最先引起我们注意的,就是这些过程如何进行(即运动)的问题,这也就是我们将要研究的问题。
在生理学方面,往往需要研究一组互相近似的许多现象中的各不相同的成员,而其时某一个成员的研究却是对于研究者,较之其他一个成员比较地更便利,这是常有的情形。在我们的研究的场合,内制止过程的研究是占着优势的。同时这制止过程运动的研究也鲜明地说明皮肤分析器对于生理学者所提供的极大利益,因为皮肤分析器具有巨大的、完全可以接近的感受性表面。
现在举出一个基本的出发性的实验。由于这个实验,开辟了大脑两半球生理学的一个重要的新篇章。这个实验和其次的若干实验都是克拉斯诺高尔斯基(Н.И.Красногорский)所做的。
实验 在实验狗一侧后腿上装上一系列的刺激器(5个),以给予皮肤机械性刺激。各刺激器是从最下者起算,以3、9、15、22厘米的距离从足部向上排列,最下的刺激器是应用于制止性作用的,而其他4个上方的刺激器引起阳性效力。这些各部位的反射是这样形成的,就是与通常情形相同地,起初在一个部位形成了阳性食物性条件反射。因为初期驯化的缘故,其余的4个部位也多少具有阳性作用。以后这4个上方的部位的刺激,都不断地并用食物而强化,直到这4个部位具有几乎同等的或完全同等的效果时为止;最低部位的刺激,相反地,虽然重复地被应用下去,却不并用食物的强化手续,直到该部位失去一切阳性作用而为制止作用所代替时为止。在下述的记录表里,条件刺激继续作用30秒以后,再并用无条件刺激。5个部位各用数字号码加以表示。由下向上地,最低的部位是0,其余的4个部位是1、2、3、4,同时上方4个部位与最下方相隔的距离,用括弧内数字加以表示。当然,阳性刺激都并用了食物,而制止性刺激却不并用食物。
我们在这个记录里看见了什么?3个不同的阳性刺激物的试验都显出相等的垂涎分泌的效力,就是30秒钟的分泌量都是5滴。其次,在完全相等的时间条件之下,就是说,在制止性刺激物三次被应用以后过1分钟,再个别地试验各阳性刺激物。于是这些阳性刺激物的效力就完全不同。与制止性刺激物最接近的部位的效力,一次是完全不发生,另一次也几乎没有作用(不到一滴)。其次的上方部位几乎失去了效力的一半,而最上的两个部位,或者保持原有的效力,或者作用反而增大若干。这个实验的意义是很明了的:因为皮肤的各点当然就是大脑两半球皮质内与此相当各点的射影(проекция),所以根据上述实验和与此相同的实验而不能不达到一个结论,就是№0(注:见附录六,戈绍龙学习笔记的第九讲(本书p.313)。)部位由于刺激所引起的制止过程,在大脑皮质内由与此相当的一点向该点的邻接各点扩布,而各点与制止过程发生点距离越远,所受的影响也就越微弱。对于距离最远的各点,几乎完全没有什么影响。这就意味着,在我们面前的现象是制止过程由出发点向大脑两半球实质的扩展。
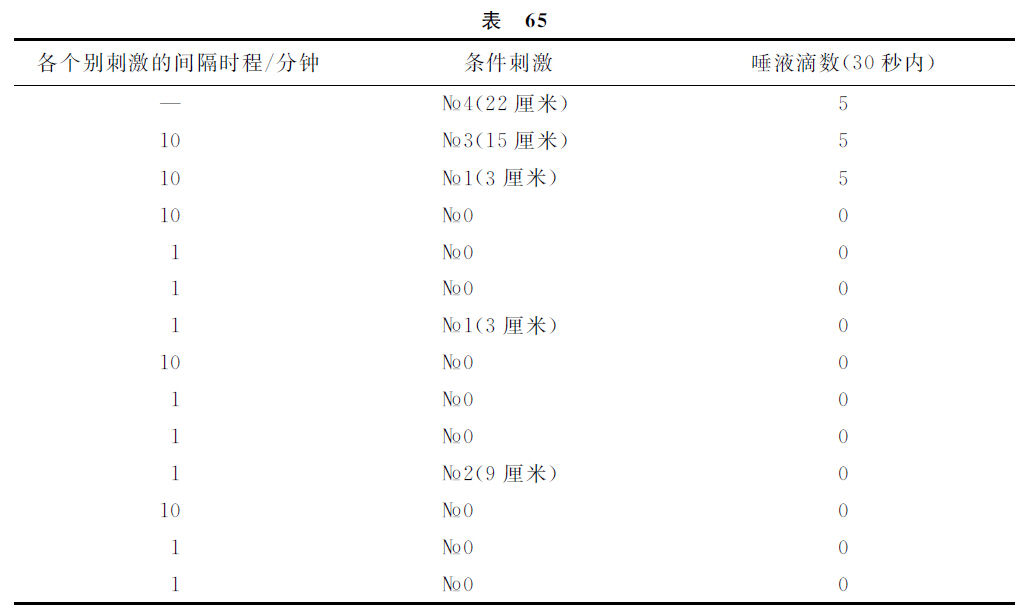
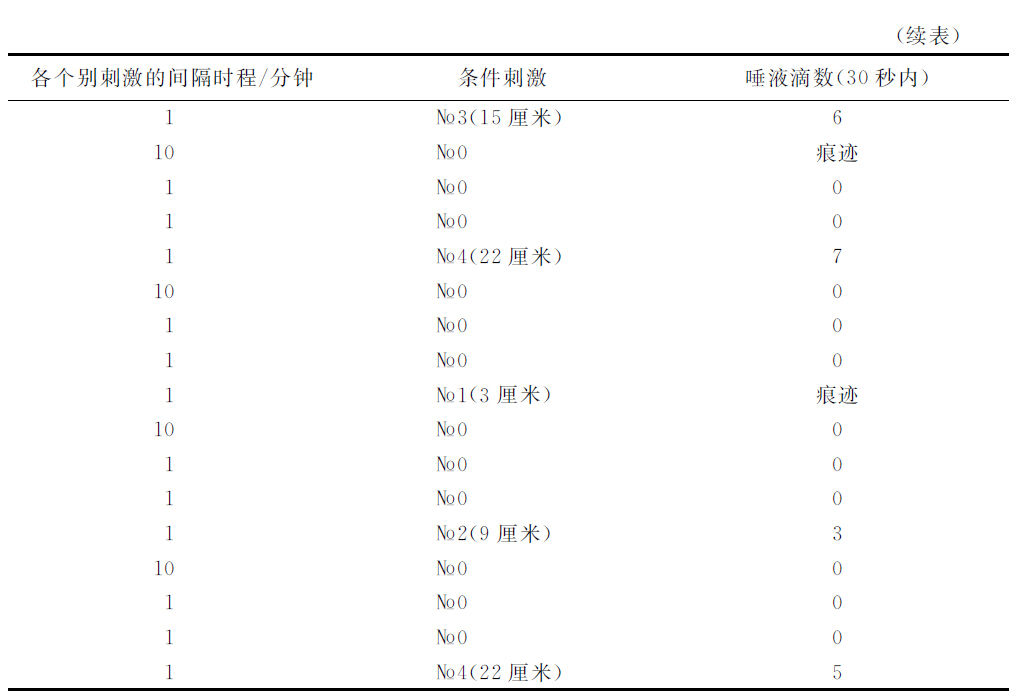
如果我们应用制止过程的另一个强度,该制止刺激物作用的累积化(суммировать)不是三次而是或多或少的次数,并且所采取的制止过程发展终了瞬间与阳性刺激物开始瞬间两者的间隔时程是与上述实验相异的,那么,当然我们所获得的唾液分泌量的数字是别样的,然而意义却完全相同。与此有关的另一个实验的记录如下。

这样看来,在间隔时程较小的场合,较强的制止过程,也对于最远隔一点发生显著的影响。
如果在一个实验里,试验各个不同阳性刺激物的作用,而制止性刺激物作用终了后至其次位刺激物的间隔时程是各不相同的,那么,可以完全明了地看见,制止过程起初扩布很远,其次渐渐地离开稍远的各点,最后也离开与出发点最接近的各点。
现在举出与此有关的一个实验。
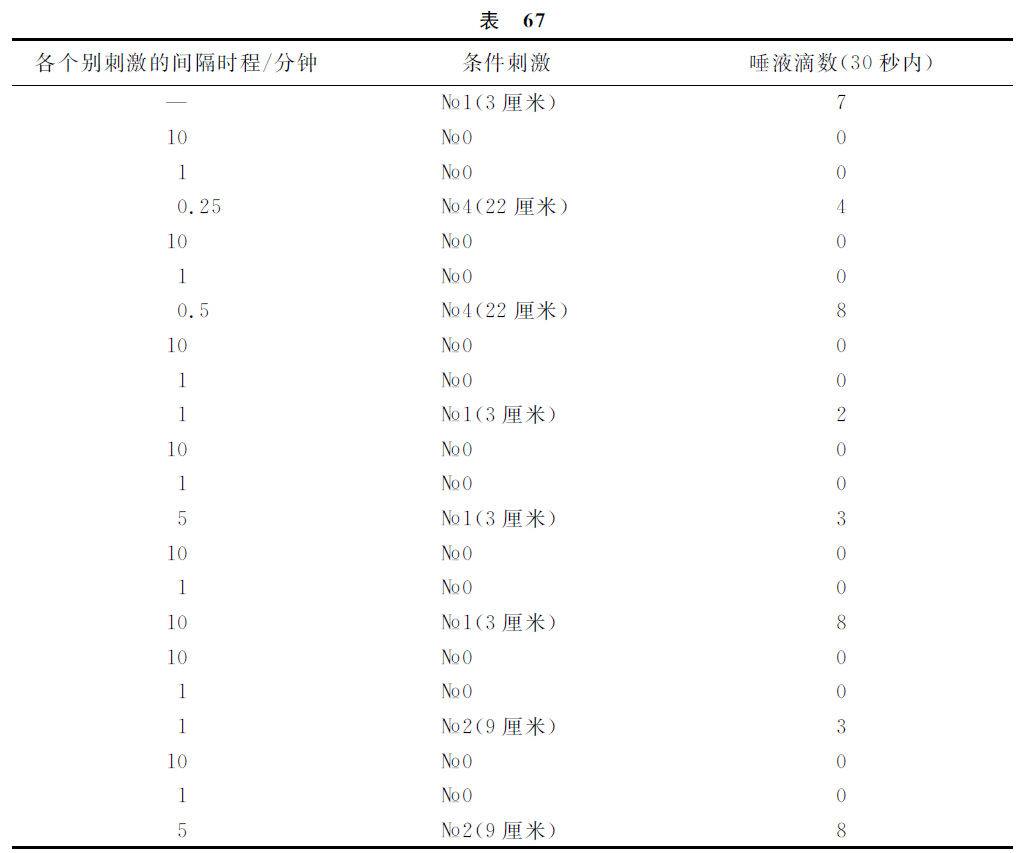
这样,№4在0.5分钟以后,№2在5分钟以后,№1在10分钟以后,都解除了制止过程。
在几天或几周以内,分化相的实验越频繁,远隔各点制止过程的解除就越快,并且在一个实验里,如果阳性刺激与阴性刺激多次重复地被应用,上述的结果就会有时发生。
值得提及,这些实验多次在许多生人的面前,都有过供览的证明,并且在多人参加的一个医学协会的开会时期也完全顺利地成功。
这种制止过程逐渐从其所暂时保持的各点离开的事实,应该怎样解释呢?这是各点上制止过程的毁灭或消失吗?或者这是遭遇着另外的某一种过程的影响,因而该制止过程仿佛是回到原有的出发点去而集中起来吗?我们注意于一个反复发生的正确而恒常的事实罢。在分化相的应用回数越多及越坚立的场合,完全与此并行地,该分化相的后继性制止就在时间和空间两方面都会减小。由于这个事实的存在,我们自然会倾向于第二个假定,就是说,我们可以假定,这些事实是意味着扩展过程的对立过程——即是过程的浓集(концентрирование),而不是意味着扩展过程的消散或减弱(рассеяниеилиослабление),所以这是向某一点的集中或增强(сосоедоточениеилиусиление)。以后我们将举出有利于这个结论的一些重要事实。
像我们刚才看见的,在克拉斯诺高尔斯基的一些实验里,浓集过程进行是缓慢的,是需要几分钟的。可是在这些相同的实验里,扩展过程的进展却是极快的,因此克拉斯诺高尔斯基不曾能够捉摸住这一扩展的进程,也不曾发现它。
当然,制止过程具有向两方面进行的性质,这是很重要的,所以我们对于这个主题必须利用各种变式的实验而进行研究。第一,引起我们注意的是在条件反射消去的场合所发生的一种制止过程[高冈(В.А.Коган)的实验],并且我们也利用了相同的皮肤分析器和皮肤机械刺激,而实验的开始手续却是与克拉斯诺高尔斯基的实验相同的。准备先在某一个部位形成酸的阳性皮肤机械刺激性条件反射。其次,该条件反射以驯化的性质,在身体一侧的全皮肤表面都尽可能地形成了同等强度的反射,于是才着手做预定的试验。在预定的某一天,使皮肤的某一个点蒙受刺激的作用,记载其1分钟内的唾液分泌效力,但这条件性刺激并不与无条件反射并用,这样,条件反射就消去了。在每个很短的间隔时程(2分钟)以后,上述的处理手续重复被应用下去,直到刺激效果变成零的时候为止。于是在零相刺激成立后的各种不同时期以内,或者刺激皮肤的这一点,或者刺激另一点,并将1分钟的刺激效力记载下来。这些其他部位的各点,在一定的时间条件之下,也或多或少地成为被制止的状态。引起反射消去的皮肤第一个点,我们把它叫做第一级的消去(在第四讲曾经提及这个问题),而其他各点的消去被叫做第二级消去。所以,消去性制止并不是停留于受一定外来刺激影响的大脑内的某一点,而是向大脑实质的各部扩散、扩展的,就是说,此时所发生的过程是与我们在分化性制止过程的场合所见相同的。
很显然,在每个个别的实验里,我们必须更主动地使用皮肤的各部位,以做条件消去的实验,否则我们也许又有分化性制止的形成,会坚固地在该形成的部位保持长时期的效力(几个月),而不能获得消去性制止。消去性制止的效力在几分钟内,最多在1~2小时以内,在第一级消去的部位会消失。
现在举出说明这种情形的实验。
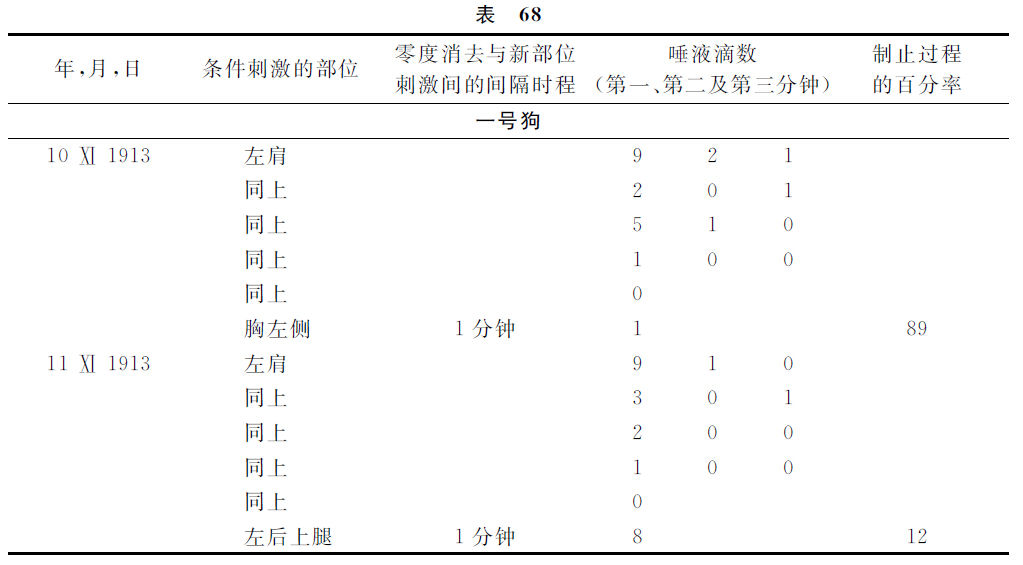
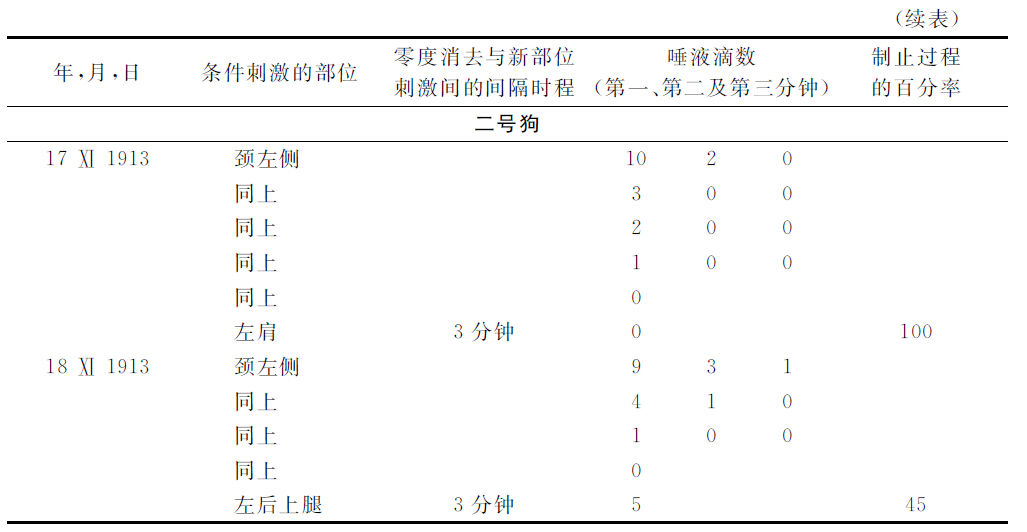
显然,如果与第一级被消去的地位相隔越远,第二级被消去的部位的制止过程也就越加微弱。
现在,如果在零相消去过程以后经过种种不同的间隔时程而刺激第二级消去的部位,那么,我们会看见,该间隔时程越大,该第二级消去部位的制止过程就会越小。如果条件相反,结果就相反。
现在举出说明这样情形的一些实验。


显然,上述的现象就是我们在分化性制止场合叫做制止过程集中的一种现象,因为制止过程与时并进地逐渐离开远隔的各点,而逐渐向出发点接近。
同时在上述实验里还有引起注意的一个详情。对于各只不同的动物,第二级消去各点制止过程消去的速度也大不相同。对于一号狗需要1分钟的事情,对于二号狗却需要15分钟。当然,这是事情很重要的一面,也就是用数字表现高级神经活动的特色。并且可证明这不是由任何偶然性而产生的影响,因为对于这三只狗做这些实验的时间有几个月之久,可是上述的区别是依然不变的。
在这一系列的实验里(高冈实验),我们能够看见了制止过程扩展的进程,就是说,从出发点开始,制止过程会渐渐地扩散开来。现在举出一些实验。在这些实验里,我们试验了与第一级消去点有种种距离的皮肤各部位的制止过程的状况,并且在第一级消去点完全消去以后,就即刻试验。
现在举出与此有关的实验。
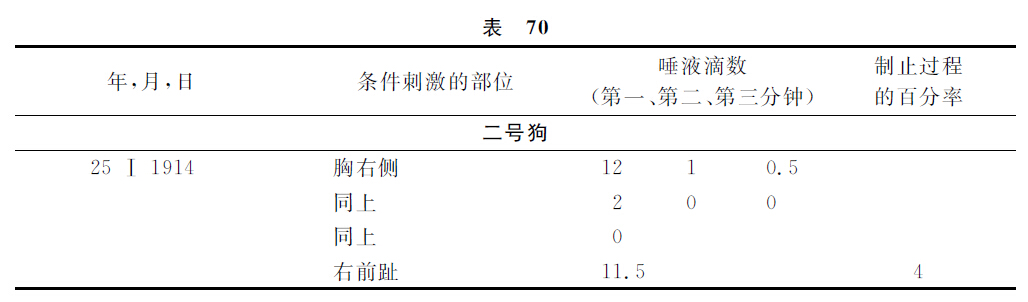
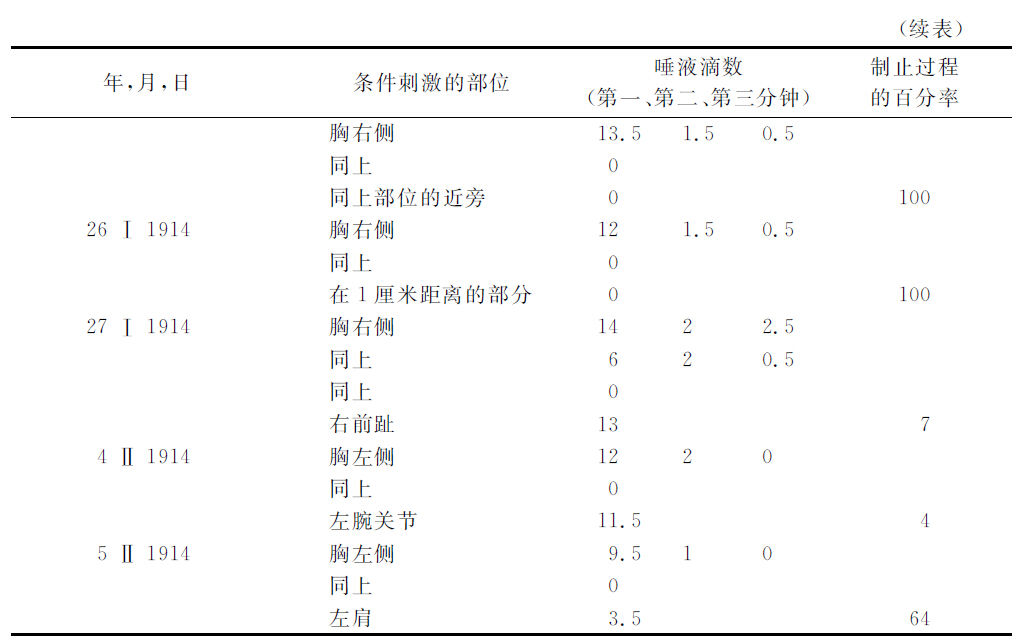
将这些实验的结果与对这同一只狗所做的以前实验的结果比较起来,就显然地证明着,直接地在第一级消去部位的制止过程完全发展以后,只在与此点最接近的各部位也有同样的完全的制止过程,而在较远的部位却几乎没有制止过程的发生,即几乎不能察觉。令人感兴趣的是,应用于这些实验的三只狗,由于所选定的条件的关系,其远隔各点的状况也是很不相同的。一号狗的远隔点不但不被制止,反而表现很增强的阳性效力,而三号狗的远隔点已经有明显的制止状态。
举例如下。
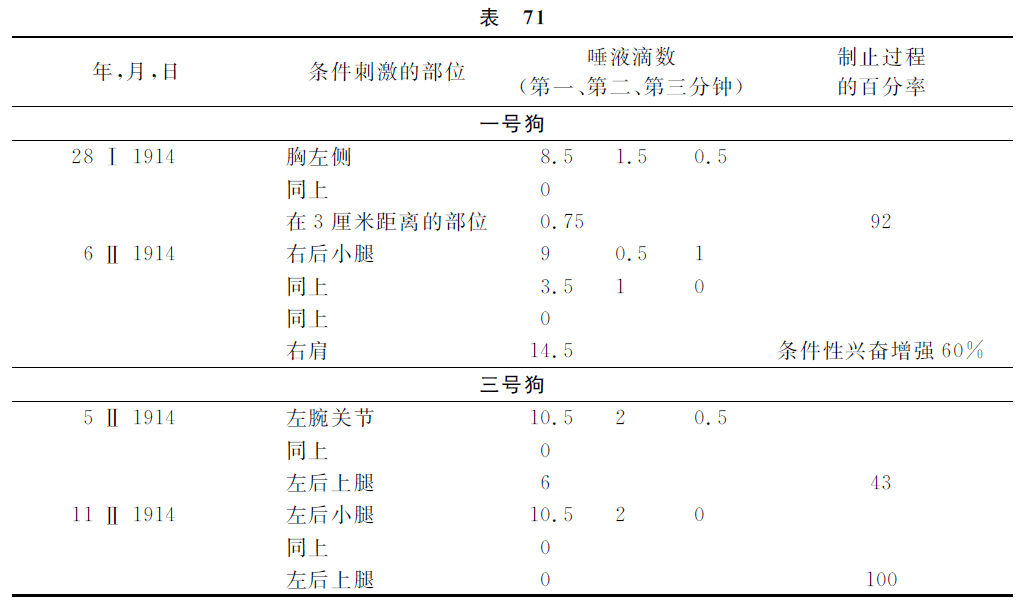
以后高冈聚集了很多的数字材料,这当然更详细地说明这三只狗的制止过程的一切进行状况。按照这些资料,情形大致是如下的。一号狗的扩展过程大约需要了20秒钟,而集中过程的时间继续到75秒钟。二号狗的扩展过程在3分钟的时候完结,而集中过程的进行需要了15分钟。三号狗的扩展是4~5分钟,而集中是20分钟。这样看来,这三只狗的制止过程进行的继续时程很互有差异,但扩展与浓集两过程时间的对比却是几乎恒常的,就是浓集比扩展多4~5倍。然而与此有关的这些材料,不能认为是完全不可非难的,因为各条件反射出发点的反射量往往是很不相同的,而且在一些场合,并不曾能够确定这些数字差异的原因。
关于克拉斯诺高尔斯基及高冈两人的实验,请诸位注意于如下的情形。在克拉斯诺高尔斯基若干实验的场合,特别在高冈一号狗的实验的场合(参看1914年2月6日的实验),很屡屡地在第一级消去部位消去过程完全成立以后,检查远隔部位的场合,该远隔部位的阳性效力不但不曾消去,反而或多或少地,并且有时很显著地增大起来。在再下一讲内,我们会详细地认识这个特殊的现象。
关于我们所谓条件性制止的进行[安烈勃(Анреп)实验],我们也在皮肤分析器上做过实验。当机械性刺激以条件刺激的性质在皮肤的不同部位形成了条件反射,并且各条件反射量相等的时候,如果只在一个部位使这阳性刺激与其他一个分析器有关(电铃)的刺激互相复合,重复地应用下去,并且不并用无条件反射的强化处理,于是这一个部位的阳性刺激就会变成阴性制止性的刺激。在颈部与胸部左侧境界的一个部位构成了这个制止点(№0),而在肩部(№1)、前腿趾部(№2)、胸与腹的交界部(№3)、腰部(№4)、后肢小腿(№5)及后肢趾部(№6)的各部位,依然都是阳性作用的各点。实验的进行情形是如下的。在一个预定的实验里预先检查了一个一定部位的完全阳性条件反射量。其次就应用制止性复合物,以后以各种的间隔时程,重复地应用这阳性刺激物。在个别的各实验里,对于其他的各阳性部位,也用同样的手续加以处理。阳性刺激和制止性刺激的继续时间都是30秒钟。结果列于下表。
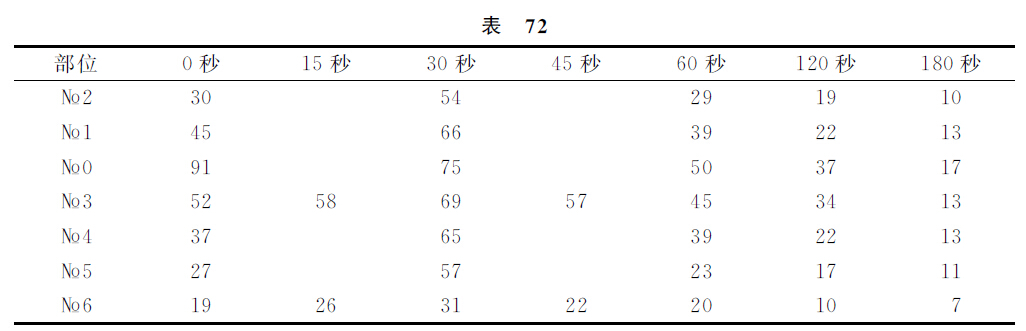
第一个直行是皮肤受刺激的各部位的号码。其他的各直行表示各部位的制止作用量,但这些数字是制止量对于阳性刺激量的百分率。最上的一个横列表示制止性刺激终了后的间隔时程。
如我们在上表里所看见的,在某一个一定点上形成了条件性制止的场合,制止过程起先扩布到一个分析器的全部,而在距离出发点越远的部位,制止过程也就越弱,并且各点的最大制止过程都只在30秒钟以后才能达到,以后各部位的制止过程同时渐渐地开始减弱。只有№0点却是例外地即刻达到最大的数值,而不是逐渐增强的。当然,即在这些实验里,偶然的一些新异刺激也发挥紊乱性的作用,或多或少地,有时制止了阳性的反射,有时使制止性反射解除制止化了。
上述的关于制止过程进行所做的三个系列实验,是与分化性制止、消去性制止以及所谓条件性制止的三种过程进行的问题有关的。由于这些实验而确定了一个事实,第一是,在某一点上以适当的处理而形成的制止过程,会扩展于一个分析器的全部,第二是,这扩展的制止过程又会慢慢地消失。可是在这个事实的详情方面,还有些显著的区别,有时这些区别甚至是本质性的。在克拉斯诺高尔斯基分化性制止的场合,制止过程的扩展是发生于瞬间的,是实验者所不能捉摸的,不过集中过程的进行却需要一定的时期。在高冈实验的场合,制止过程也是慢慢地散布于整个分析器的,不过比集中过程的发生却是远远更快。这个区别也许可以认为是与发展中的制止过程的强度有关的。克拉斯诺高尔斯基通常地在制止性刺激物虽然即刻发挥完全作用以后(零相),还把制止性刺激物重复地应用了若干次。高冈在消去过程一达到零的时候就不再应用制止性刺激物了。可是在条件性制止场合的区别,是远远更复杂的、更本质性的。在安烈勃所做的条件性制止实验和克拉斯诺高尔斯基的分化性制止实验两个场合,制止过程都向一个分析器的全部扩展,不过扩布的程度却有不同,可是以后,安氏的实验的制止过程却与克拉斯诺高尔斯基的场合不相同地,在一定的时期以内,又同时在各点上逐渐加强,以达到某一个极大的数值。在极大数值以后,全部各点上的制止过程,又同样地同时渐渐开始减弱。这样看来,在条件性制止的场合,与分化性制止及消去性制止相异地,不曾能够发现从一点向另一点的移行,既不从一个方向移行到另一个方向,也不对出发点前进或后退。
因为在我们面前的这个新范围是越过越广大而越深入的,并且特别有关大脑两半球皮质内神经过程进行的问题是越过越复杂化的——这是我们以后会看见的——,所以我认为,我们材料的说明必须按照历史的顺序(即我们25年间的实验),因为现在即刻把研究的全部各点都归结于具有联系性的最后的结论,这还是不可能的。
在刚才所记载的安烈勃研究里,已经详细地引证了一个事实,其一部分是其他学者早已发现的。这个事实就是,如果在身体一侧的各个不同的部位形成了阳性或阴性条件反射,并且如果这些反射在制止过程进展的时候发生动摇(特别在安烈勃实验里制止过程的进展上),那么,在身体另一侧各个相对的部位(симметричныеместа)并不需要任何条件反射形成的手续,自然会非常正确地有同样的条件反射的出现。关于这个有趣的事实及其一部分的分析是以后会有记载的。
与皮肤分析器的制止过程进行有关的实验相同地,我们试做了耳(即声音)分析器的实验,因为我们假定着,在大脑两半球皮质里也有与耳分析器末梢部相当的射影。以这个目的,我们(马努伊洛夫及伊凡诺夫·斯莫连斯基)采用了各种不同的声音(各种的音、拍节机响声及嘘声)当做一些条件刺激物,先用其中的一个刺激物做反射消去的实验,以后再研究这已消去的刺激物对其他各刺激物的影响。
此地举出伊凡诺夫·斯莫连斯基有关本问题的实验。对于一只狗,用4个音形成了个别的食物性条件反射。4个音都是利用马克斯·考尔(Max Kohl)的变音器(тонвариатор)而获得的。4个音是各由两对邻接音而成立的,而这些音的相隔的距离是3个八度音程(октава),就是大八度音程的si(H123次振动),小八度音程的do(C130次振动),第三个八度音程的do与re(D1161次振动,C1036次)。并且也用嘘音及拍节机响声(每分钟100次)形成条件刺激物。这实验是这样做的,就是把某一个条件反射消去而达到零相以后,或者即刻地,或者各经过1、3、5、7、10、12及15分钟,在各个不同的实验里,检查全部其余的各反射。全部的各条件反射都成为制止状态,不过被制止的程度却有不同。一部分的结果如下表。
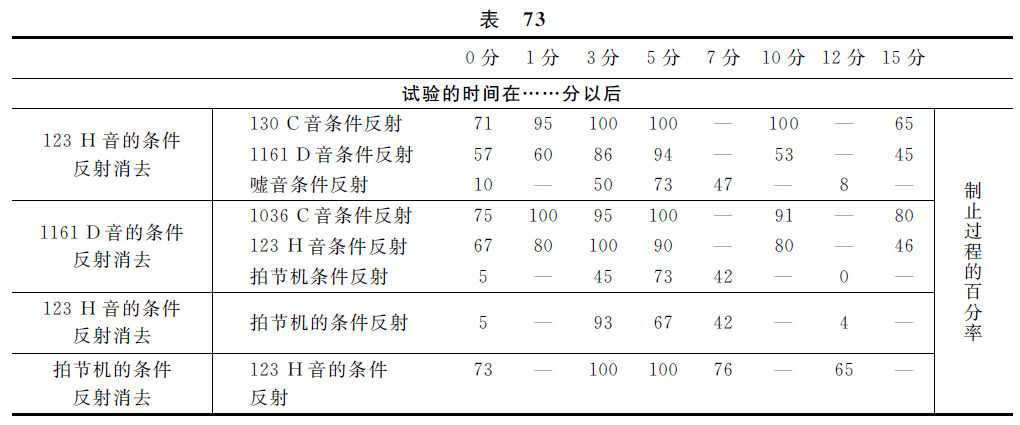
这个表指示着,在一对低音中的一个音被消去的场合,另一个低音的反射就比每个高音的条件反射,更迅速地达到最大的制止过程,更长久地保持这最大量,更慢慢地解除这制止过程。在一对高音中的一个高音被消去的场合,另一个高音的反射也比每个低音的反射,更迅速地达到最大的制止过程,更长久地保持这最大量,更慢慢地解除这制止过程。在任何一个音被消去的场合,嘘音反射的与拍节机反射的第二级制止在各种关系上的表现,都比各音反射的表现更远远地微弱,并且相反地,在嘘音反射及拍节机反射被消去的场合,其他各音反射的第二级制止的表现在一切关系上都是很强烈的。
当然,只从各音消去及消去后检查各音反射的实验说,我们可以多少有根据地主张,制止过程是在声音分析器的脑皮质终末部,即在考尔铁氏器官射影的脑皮质终末部内进行着的。消去了的各反射,一方面对于各个纯音发生影响,另一方面对于嘘音及拍节机响声发生影响——这种互相影响的结果如何,很像是由于这些不同的各种刺激的不同强度而决定的。
我们必须承认,在皮肤分析器方面明了地显现的情形,也在声音分析器的方面足够明了地显现出来,就是说,在一个分析器脑终末部内的制止过程由一点向其他一点的运动,也发生于其他各分析器之内。然而与此有关的证明,受着显然的技术条件的限制,是很困难的,或者甚至在目前是不可能的。如果把这个事实的详细情形能够确定,也许就有一个基础,以说明这些分析器基本性的构成。
