第一部分 人类的由来或起源
· The Descent of Otigin of Man·
我曾着眼于两个明确的目的,其一,在于阐明物种不是被分别创造的,其二,在于阐明自然选择是变化的主要动因,虽然它大部分借助于习性的遗传效果,并且小部分借助于环境条件的直接作用。然而,过去我未能消除我以往信念的影响,当时这几乎是一种普遍的信念,即各个物种都是有目的地被创造的;这就会导致我不言而喻地去设想,构造每一细微之点,残迹构造除外,都有某种特别的、虽然未被认识的用途。一个人如果在头脑里有这种设想,他自然会把自然选择无论过去或现在所起的作用过分夸大。有些承认进化论但否定自然选择的人们,当批评我的书时似乎忘记了我曾着眼的上述两个目的;因此,如果我在给予自然选择以巨大力量方面犯了错误——这是我完全不能承认的,或者我夸大了它的力量——在其本身来说这是可能的,那么我希望,至少我在帮助推翻物种被分别创造的教条方面作出了有益的贡献。

▲雌极乐鸟的色彩暗淡而且缺少任何装饰物,反之,雄极乐鸟大概是所有鸟类中最精于装饰者,其装饰如此多种多样,以致见者无不赞叹。
第一章 人类起源于某一低等生物类型的证据
有关人类起源证据的性质——人类和低于人类的动物的同源构造——有关一致性的各点——发育——残迹(退化)构造,肌肉,感觉器官,毛发,骨骼,生殖器官等——这三大类事实同人类起源的关系
如果一个人想要决定人类是否为某一既往生存类型的变异了的后裔,他最初大概要问,人类在身体构造和心理官能(mental faculties)方面是否变异,哪怕是轻微的变异;倘如此,则这等变异是否按照普遍适用于低于人类的动物的法则遗传给他的后代。还有,就我们贫乏知识所能判断的来说,这等变异是否像在其他生物的场合中那样,乃是同样的一般原因的结果,并且受同样的一般法则所支配;例如,受相关作用,使用和不使用的遗传效果等等法则所支配?作为发育受到抑制、器官重复等等的结果,人类是否会变成同样的怪相,并且人类的任何畸形是否表现了返归某一先前的、古远的构造型式?自然还可以这样问,人类是否像如此众多的其他动物那样,也产生彼此仅有微小差异的变种(varieties)和亚族(sub-races),或者产生差异如此重大的种族(race)而必须把它们分类为可疑的物种?这等种族如何分布于全世界;而且,当他们杂交时,他们在第一代和以后各代彼此发生作用吗?此外,还可追问其他各点。
追问者其次将问到重要之点,即,人类是否以如此迅速的速度增加,以致不时引起剧烈的生存斗争;结果导致无论身体或心理方面的有益变异被保存下来了,而有害的变异被淘汰了。人类的种族或种(species)无论用哪个名词都可以,是否彼此侵犯,相互取而代之,因而有些最终归于灭绝?我们将会看到,所有这些问题一定可以按照对低于人类的动物的同样方式得到肯定的回答。就大多数问题来说,的确显然如此。但对刚才所谈到的几个需要考虑的问题暂时推迟予以讨论,可能是方便的。我们先看一看,人类的身体构造在多大程度上或多或少明确地显示了一些痕迹,以说明他来自某一低等类型。在以后数章,将对人类的心理能力(mental power)在与低于人类的动物的心理能力的比较下加以考察。
人类的身体构造
众所周知,人类是按照其他哺乳动物同样的一般形式或模型构成的。人类骨骼中的一切骨可以同猴的、蝙蝠的或海豹的对应骨相比拟。人类的肌肉、神经、血管以及内脏亦如此。正如赫胥黎和其他解剖学者所阐明的,在一切器官中最为重要的人脑也遵循同一法则。比肖夫(Bischoff) (1) 是一位站在敌对方面的见证人,连他都承认人类的每一个主要的脑裂纹和脑褶都同猩猩(Drang-outang)的相似;但他却接着说,它们的脑在任何发育时期中都不完全一致;当然也不能期望它们完全一致,否则它们的心理能力就要一样了。于尔皮安(Vulpian) (2) 说:“人脑和高等猿类的脑的差别极其轻微。我们对这种关系不应有错觉。就脑部的解剖性状来看,人类之比类人猿,不但较近于类人猿之比其他哺乳动物,而且较近于类人猿之比其他猿类,如绿背猿(Des guenons)和猕猴(Des macaques)。”但是,在这里进一步详细地指出人类在脑的构造和身体其他一切部分上同高等哺乳动物的一致性,则是多余的了。
可是,对于构造没有直接或明显关系的少数几点加以详细说明,还是值得的,这种一致性或彼此关系借此会得到很好的阐明。
人类容易从低于人类的动物那里染上某些疾病,如恐水病 (3) 、天花、鼻疽病、梅毒、霍乱、疱疹等 (4) ,而且容易把这些病传给它们;这一事实证明了它们的组织和血液既在细微构造上也在成分上都密切相似, (5) 这比在最优良的显微镜下或借助于化学分析来比较它们还要明显得多。猴类像我们那样,常患许多同样的没有传染性的疾病。例如伦格尔(Rengger) (6) ,曾在巴拉圭卷尾猴(Cebus azarae)的原产地对它进行过仔细的观察,它容易患黏膜炎,具有通常的症状,如经常复发,就会导致肺结核病。这种猴还患中风、肠炎和白内障。幼猴在乳齿脱落时常死于热病。药物对它们产生的效果,同对我们一样。许多种类的猴对茶、咖啡、酒都有强烈的嗜好,我自己亲眼见到,它们还吸菸取乐。 (7) 布雷姆(Brehm)断言,非洲东北部的土人把装有浓啤酒的器皿放在野外,使野狒狒(baboons)喝醉以捕捉它们。他曾看到他自己圈养的几只喝醉的狒狒,他对它们的醉态和怪相做过引人发笑的描述。醉后翌晨,它们非常易怒而忧郁;用双手抱住疼痛的脑袋,作最可怜的表情;当给它们啤酒或果子酒时,它们就厌恶地躲开,但对柠檬汁却喝得津津有味。 (8) 一只美洲蛛猴(Ateles),当喝醉了白兰地酒之后,就会永远不再碰白兰地酒,这样,它就比许多人都更聪明了。这些琐事证明了人类和猴类的味觉神经是多么相似,而且他们的全部神经系统所受到的影响又多么相似。
人类的内脏会感染寄生虫,时常因此致死,并且受到外部的寄生虫的侵扰,所有这等寄生虫同感染其他哺乳动物的寄生虫都属于同属(genera)或同科(families),至于疥癣虫,则属于同种。 (9) 人类有如其他哺乳动物、鸟类甚至昆虫那样,受一种神秘的法则所支配 (10) ,这一法则使某些正常过程,如妊娠、成熟以及各种疾病的持续,均按月经期进行。人类的创伤按照同样的愈合过程得到恢复,人类截肢后的残余部分,特别是在胚胎早期,也像在低等动物的场合中那样,有时具有某种再生的能力。 (11)
像物种繁殖这个最重要机能的全部过程,从雄者的最初求偶行为 (12) 到幼仔的出生和哺育,在所有哺乳动物中都是显著一样的。猴在幼仔时不能自助的情况几乎同我们的婴儿一样;在某些属中,猴仔在外貌上完全不同于成猴,犹如我们的子女不同于他们的充分成熟的父母一样。 (13) 有些作者极力主张,作为一种重要的差别,人类幼儿的成熟期要比任何其他动物迟得多。但是,如果我们注意看一看居住在热带地方的人类的种族,其差别就不大了,因为,猩猩据信在10~15岁时才达到成年。 (14) 男人同女人在身材大小、体力、体毛多少等方面以及在精神方面都有差别,许多哺乳动物的两性也是如此。因此,人类同高等动物,特别是同类人猿在一般构造上、在组织的细小构造上、在化学成分上以及在体质上的一致性是极其密切的。
胚 胎 发 育
人是从一个卵发育成的,卵的直径约为1英寸的1/125,它在任何方面同其他动物的卵都没有差别。人类胚胎在最早时期同脊椎动物界其他成员的胚胎几乎无法区分。此时动脉延伸为弓形分支,好像要把血液输送到高等脊椎动物现今不具有的鳃中,虽然在他们的颈部两侧还留有鳃裂(图1,f,g),标志着它们先前的位置。在稍晚时期,当四肢发育时,正如杰出的冯·贝尔(Von Baer)所指出的,蜥蜴类和哺乳动物的脚,鸟类的翅膀和脚,以及人的手和脚,都是由同一个基本型式发生的。赫胥黎教授说:“在相当晚的发育阶段,人类幼儿才同幼猿有显著的差别,而猿在发育中同狗的差别程度,正如人在发育中同狗的差别程度一样大。看来这一断言好像要令人一惊,但它的真实性是可以证明的。” (15)
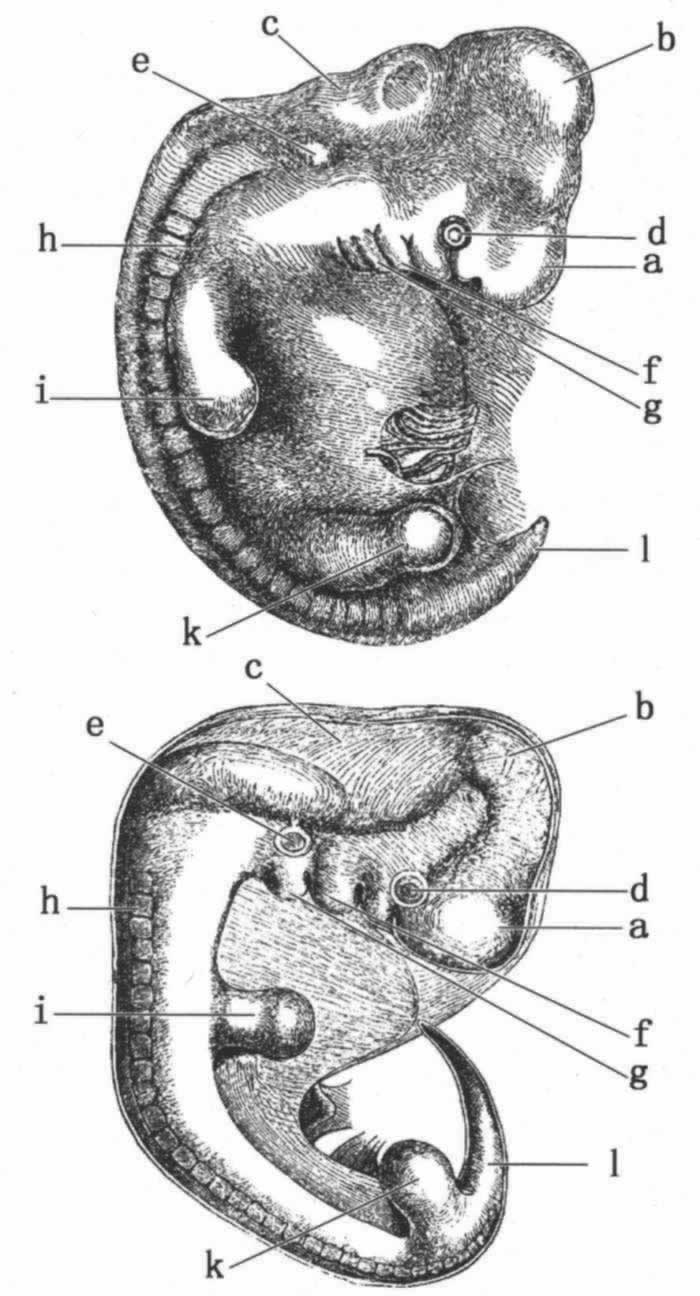
图 1 上图:人的胚胎(采自埃克);下图:狗的胚胎(采自比肖夫)
a.前脑:大脑半球等;b.中脑:四叠体;c.后脑:小脑,延髓;d.眼;e.耳;
f.第一鳃弓;g.第二鳃弓;h.在发育过程中的脊椎和肌肉;i.前肢;k.后肢;l.尾或尾骨。
由于本书的读者可能从来没有看过有关胚胎的绘图,所以我刊登了一幅人的和一幅狗的胚胎图,约在同一早期发育阶段,这幅图是从两部无疑正确的著作 (16) 中仔细复制的。
当举出了如此高水平的权威人士所做的以上叙述之后,我再借用别人的详细材料来阐明人类的胚胎密切类似于其他哺乳动物的胚胎,就是多余的了。然而却可以补充说明,人类的胚胎同某些成熟的低等类型在构造的种种方面同样也是类似的。例如,心脏最初仅是一个简单的搏动管,排泄物由一个泄殖道(cloacal passage)排出,尾骨(Os coccyx)突出像一条真尾,“相当地延伸到残迹腿之外”。 (17) 在所有呼吸空气的脊椎动物胚胎中,被称为吴耳夫氏体(corpora wolffiana)的某些腺与成熟鱼类的肾相当,并且像后者那样活动。 (18) 可以观察到,甚至在较晚的胚胎时期人类和低于人类的动物也有若干显著的相似性。比肖夫说,整七个月的人类胎儿的脑旋圈(convolutions)与成年狒狒的处于同样的发育阶段。 (19) 正如欧文教授所指出的,“大脚趾当站立或行走时形成一个支点,这在人类构造中恐怕是一个最显著的特性” (20) ;但是在约一英寸长的胚胎中,怀曼发现“大脚趾较其余脚趾都短;不是同其余脚趾平行,而是从脚的一侧斜着伸出,这样,它就与四手类动物(Quadrumana)这一部分的永久状态相一致了”。 (21) 我用赫胥黎的话来做结束,他问道:“人类是按照不同于狗、鸟、蛙或鱼的途径发生的吗?”然后他说,“立刻可以作出肯定的回答:毫无问题,人类的起源方式以及他们的早期发育阶段同在等级上直接处于其下的动物是完全相同的;毫无疑问,人类与猿类在这等关系上远比猿类与狗要近得多”。 (22)
残 迹 器 官
这个问题虽然在本质上不及上述两个问题重要,但由于几点理由,还要在这里更充分地予以讨论。 (23) 我们举不出任何一种动物,它们的某些器官不是处于残迹状态的,人类也不例外。必须把残迹器官同新生器官(nascent organs)区别开来,虽然在某些场合中把它们区别开是不容易的。残迹器官要么是绝对无用的,如四足动物雄者的乳房或反刍动物的永远不会穿出齿龈的切齿(incisor teeth);要么就是对其现今所有者仅有如此微小的作用,以致我们无法设想它们是在现今生活条件下发育出来的。处于后一状态的器官并不是严格残迹的,但有趋于这个方面的倾向。另一方面,新生器官虽然不是充分发育的,对其所有者却是高度有用的,而且能够进一步向前发展。残迹器官变异显著,关于这一点,可以部分地得到理解,因为它们无用,或近于无用,所以不再受自然选择所支配。它们往往完全受到抑制。当这种情形发生时,它们还是容易通过返祖而偶尔重现——这是一个十分值得注意的情况。
致使器官成为残迹的主要动因,似乎是由于在这个器官被主要使用的那个生命时期(这一般是在成熟期)却不使用它了,同时还由于在相应的生命时期的遗传。“不使用”这个名词不仅同肌肉活动的减少有关,而且包括血液流人某一部分或器官的减少在内,后一情形是由于压力交替较少或者由于其习惯性活动以任何方式变得较少所致。然而,在某一性别中表现为正常的那些器官,在另一性别中却可能成为残迹;这等残迹器官,像我们以后将要看到的那样,常常以不同于这里谈到的那些残迹器官的发生方式而发生。在某些情况中,器官是由于自然选择而被缩小的,因为由于生活习性改变,它们变得对物种有害了。缩小的过程大概还常常借助于生长补偿和生长经济(compensation and economy of growth)这两项原理;但是,当不使用对器官的缩小完成了所有作用之后,而且当生长经济所完成的节约作用很小时, (24) 器官缩小的最后诸阶段是难于理解的。在已经成为无用的而且大大缩小了的一个部分最后地、完全地受到抑制的情况下,如果生长补偿或生长经济都不能发生作用,这大概只有借助于泛生论(pangenesis)的假说才可以得到理解。但是,关于残迹器官的整个主题,我在以前的几部著作中已经讨论过了,而且举出过例证, (25) 在这里我无须就此点再多加赘述。
已经观察到,在人类身体的许多部分中有各种处于残迹状态的肌肉, (26) 可以偶尔发现,正常存在于某些低等动物中的不少肌肉,在人类身体中却处于大大缩小的状态。每一个人一定都注意过许多动物,特别是马抽动皮肤的能力,这是由肉质膜(panniculus carnosus)来完成的。现已发现这种肌肉的残迹以有效的状态存在于我们身体的种种部分中,例如双眉借以抬起的前额肌肉。在我们颈部非常发达的颈阔肌肌样体(platysma myoides)就属于这个系统。爱丁堡的特纳(Turner)教授告诉我说,他曾偶尔发现五个不同部位——即肩胛骨附近的腋下等——的肌肉束(muscular fasciculi)都一定同肉质膜有关系。他还阐明了,“胸骨肌(musculus sternalis)不是腹直肌(rectus abdominalis)的延伸,而与肉质膜密切近似,在600个人中,3%以上有胸骨肌”。他接着说,关于“偶现的和残迹的构造特别容易变异的论述,这种肌肉提供了一个最好的例证”。 (27)
有少数人可以收缩头皮上的表面肌肉,这等肌肉是处于变异的和部分残迹的状态的。小德康多尔(M.A.de Candolle)写信告诉我一个有关长期连续保持或遗传这种能力并且异常发达的事例。有一个家族,其族长在幼年时能够仅靠头皮的动作就可以从头上把几本沉重的书扔开,他用耍这个把戏去打赌,而赢得赌注。他的父亲、叔父、祖父以及他的三个孩子都同样具有这种能力,而且均达到异常程度。这个家族在八代以前分为两支,所以上述一支的族长同另一支的族长是七世的从堂兄弟。这位远房的从堂兄弟在法国的另一地方居住,当问到他是否也具有同样的这种能力时,他立即作了表演。这个例子很好地说明了,一种绝对无用的能力可以多么持久地传递下去,这种能力大概来自我们遥远的半人类祖先,因为许多种类的猴都具有而且常常使用这种能力,它们可以上下自如地充分移动它们的头皮。 (28)
为移动外耳服务的外在肌肉(extrinsic muscles)和使不同部分活动的内在肌肉(intrinsic muscles)在人类中都处于一种残迹状态,而且它们都属于肉质膜的系统;它们在发育方面,至少在机能方面也是易于变异的。我曾见过一个人能够把整个耳朵向前拉;另外一些人能够把耳朵向上拉;还有一个人能够把耳朵向后拉; (29) 根据其中一人向我说的,只要经常触动我们的耳朵,这样把我们的注意力引向它们,大多数人在反复试行中大概都能恢复移动耳朵的一定能力。把耳朵竖起并使它们朝不同方向移转的能力对许多动物来说,无疑都是最高度有用的,因为,这样它们可以觉察危险来自何方;但我从来没听到过有充足的证据可以证明一个人所具有的这种能力对他会有什么用处。整个外耳以及各种耳褶和突起(如耳轮和对耳轮、耳屏和对耳屏等)都可以被看做是残迹的;在低等动物中,当耳朵竖起时,它们在不给耳增加很大重量的情况下起到加强和支持耳的作用。然而,有些作者猜想,外耳的软骨有向听神经(acoustic nerve)传导振动的作用;但是,托因比(Toynbee)先生在搜集了所有关于这个问题的已知证据之后,断定外耳并没有独特的用途。 (30) 黑猩猩(chimpanzee)和猩猩的耳同人类的耳异常相似,而且其特有的肌肉同样也是非常不发达的。 (31) 伦敦动物园的饲养员们向我保证,这等动物从来不移动或竖起它们的耳朵;所以,就其机能而言,它们的耳处于和人类的耳相等的残迹状态。为什么这等动物以及人类的祖先失去了竖立耳朵的能力,我们还无法说明。可能是,由于它们在树上生活的习性而且力量大,它们面临的危险很小,因此在长时期内它们很少移动耳朵,这样就逐渐失去了移动它们的能力,但我并不满意于这种观点。这大概同下述事例是类似的,即,那些大型而笨重的鸟类由于居住在海洋岛上,不面临食肉兽的攻击,因而失去了使用双翅来飞翔的能力。然而,人类和几种猿虽不能移动耳朵,却可以从在水平面上自由移动头部以捕捉来自各方的声音而部分地得到补偿。有人断言,唯独人类的耳具有耳垂;但“在大猩猩(gorilla)中发现有它的残迹” (32) ;我听普瑞尔教授说,黑人不具耳垂者并不罕见。
著名的雕塑家伍尔纳(Woolner)先生告诉我说,他时常在男人和女人中观察到外耳有一个小特征,而且他觉察到这很有意义。最初引起他注意这个问题的是当他雕塑莎士比亚剧中顽皮小妖精(Puck)的时候,他曾给这只小妖精雕塑了一个尖耳。这样,他被引导去考察各种猴的耳,此后又更加仔细地对人类的耳进行了考察。这一特征为一个小钝点,突出于向内折叠的耳边,即耳轮。如有这个特征,在婴儿一生下来它就是发达的,按照路德维格·迈耶(Ludwig Meyer)教授的说法,具有这一特征的,男多于女。伍尔纳先生为其制作了一个精确的模型,并把本书的附图见赠(图2)。这个突出点不仅朝着耳的中心向内突出,而且常常稍微突出于它的平面之外,因而从正前方或者从正后方去看头部时,都可以见到这个突出点。它们在大小上是有变异的,在位置上也多少有点变异,稍微高一些或者低一些,且有时呈现于一耳,而不见于另一耳。并不仅限于人类才具有这一特征,因为我看到伦敦动物园里的一只蛛猴(Ateles beelzebuth)也具有这种特征。兰克斯特(E.Ray Lankester)先生告诉我说,汉堡动物园里的一只黑猩猩是另一个例子。耳轮显然是由向内折叠的最外部耳边形成的,这一折叠部分似乎多少同整个外耳被持久地压向后方有关联。在许多等级不高的猴类中,如狒狒和猕猴属(Macacus)的一些物种 (33) ,耳的上部是微尖的,而且耳边全然不向内折叠;但是,如果耳边向内折叠的话,那么一个微小的点必然要朝着耳的中心向内突出,而且可能稍微地突出于耳的平面之外。我相信,在许多场合中这就是它的起源。另一方面,迈耶教授在其最近发表的一篇富有才华的论文中主张,整个情形不过仅仅是变异性的一种而已;那个突出点并不是一个真的突出点,而是由于那个突出点两侧的内软骨没有充分发育所致。 (34) 我十分乐意承认,对许多事例来说,这是一个正确的解释,如在迈耶教授所绘的图中,耳轮上有若干微小的点,即整个耳边是弯进弯出的。通过唐恩(L.Down)博士的好意帮助,我曾亲自看到一个具有畸形小脑袋的白痴人的耳,在耳轮的外侧、而不是在向内折叠的边上,有一个突出点,所以这个点同既往存在的耳的尖端并无关系。尽管如此,在许多场合中我的本来观点,即那个突出点是既往存在的直立而尖形的耳之顶端,在我看来大概还是很可能的。我之所以这样认为,是由于它们的屡屡出现,而且由于它们的位置同尖耳顶端的位置一般是符合的。有一个例子——我曾得到它的照片:那个突出点如此之大,以致遮盖了全耳的整整三分之一,倘若按照迈耶教授的观点,则必须假定,软骨要在耳边的全部范围内有同等发育,才能使这样的耳完成。我还从通信中得知两个例子,一个发生在北美,另一个发生在英国,表明上部耳边全然不向内折叠,而是尖形的,因此,它在轮廓上同一只普通四足动物的尖耳密切类似。此二例之一,为一个幼儿的耳,他父亲把这个幼儿的耳同我在一幅图中 (35) 举出过的一种猴、即黑顶猿(Cynopithecus niger)的耳作了比较,说道,他们的轮廓是密切相似的。在这两个例子中,如果耳边以正常方式向内折叠,那么一个内向的突出点一定会形成。我还可以补充另外两个例子,表明耳的轮廓依然留有稍微尖形的残迹,虽然上部耳边是正常向内折叠的——其中一只向内折叠得很狭。下面的木刻图(图3)是依据一只猩猩胎儿相片的原样仿制的[蒙尼采(Nitsche)博士好意见赠],从这幅图中可以看出,这一时期的耳的尖形轮廓同其成长时期的状态是多么不同;当成长时,它的耳同人的耳一般是密切相似的。显然,这样一只耳的尖端折叠起来,除非它在进一步发育中发生重大变化,将会形成一个向内突出的点。总之,我依然觉得,所讨论的那个突出点在无论是人类或猿类的某些事例中很可能都是既往状态的残迹。
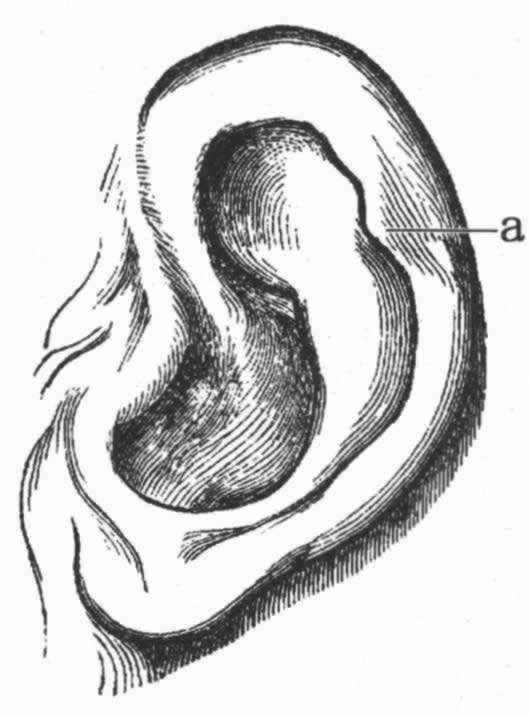
图 2 人类的耳
(伍尔纳先生雕塑并制图)
a.突出点
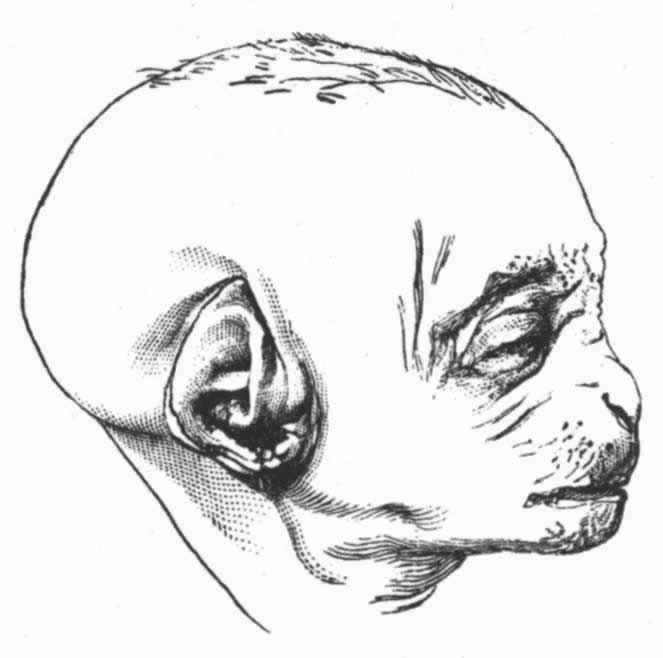
图 3 一只猩猩的胎儿
(表明这一生命早期的耳的形态)
瞬膜(nictitating membrane),即第三眼睑(third eyelid)及其附属的肌肉和其他构造在鸟类中十分发达,而且对它们有很大的机能重要性,因为它能迅速地把整个眼球遮盖起来。有些爬行动物和两栖动物,还有某些鱼类如鲨鱼,也有瞬膜。在哺乳动物的两个低等的门类(division)、即单孔目(Monotremata)和有袋目(Marsupials)中,以及在少数某些高等哺乳动物、如海象(walrus)中,瞬膜也是十分发达的。但是,在人类、四手类以及大多数其他哺乳动物中,如所有解剖学者所承认的,瞬膜不过是一种被称为半月褶(semilunar fold)的残迹物而已。 (36)
嗅觉对大多数哺乳动物来说,都是最高度重要的——如对反刍动物,用于警告危险;对肉食动物(Carnivora),用于搜索所要捕食的动物;还有,如对野猪,则上述两种意义兼而有之。但是,嗅觉甚至对黑色人种,如果还有一点用处的话,也是极其微小的,而黑色人种的嗅觉远比白色人种的嗅觉还要发达得多。 (37) 尽管如此,嗅觉并不为黑人警告危险,也不引导他们去找食物;它不阻止爱斯基摩人睡眠于恶臭的空气之中,也不阻止许多未开化人吃半腐烂的肉。在欧洲人中,各个人的这种能力差别很大;这是一位卓越的自然学者向我保证的,他具有高度发达的嗅觉,而且注意过这个问题。那些相信逐渐进化原理的人们不会轻易地承认,现今状态的嗅觉乃是由人类最初获得的。他从某一早期祖先遗传了这种处于衰弱而残迹的状态下的能力;对其早期祖先来说,它是高度有用的,而且要不断地使用它。在那些嗅觉高度发达的动物中,例如狗和马,对于人和地方的记忆是同它们的气味高度联系在一起的;这样,我们恐怕就能理解,如莫兹利(Maudsley)博士所正确指出的,为什么人类的嗅觉“在生动地追忆已经忘却的景色和地方的概念和影像时是异常有效的”。 (38)
人类几乎裸而无毛,这是同所有其他四手类的显著差别。但是在男人的大部分身体上还有少量散在的短毛,在女人身体上也有纤细的绒毛。不同种族在毛的多少上差别很大,同一种族中各个人的毛不仅在多少上,而且在部位上都是高度变异的。例如,有些欧洲人的肩部完全无毛,而另外一些人的肩部却生有茂密的丛毛。 (39) 这样散在于全身的毛乃是低等动物的均匀一致的皮毛的残迹,则是没有多大疑问的。这一观点从下述事实来看就越发可能是确实的了,即,我们知道,四肢和身体其他部分的“纤细的、短的、淡色的毛”,当在长久发炎的皮肤附近受到异常营养时,偶尔会发育成为“茂密的、长的、粗而黑的毛”。 (40)
詹姆斯·佩吉特(James Paget)爵士告诉我说,一个家族常有几个成员,他们的眉毛中有几根要比另外的长得多;所以说,甚至这种微小特性也是遗传的。这种长眉毛似乎也有它们的代表,因为黑猩猩和猕猴属的某些种在其眼的上方裸皮上生有相当于我们眉毛的很长的散毛;在某些狒狒中,有相似的长毛突出于眉脊(superciliary ridge)毛皮之外。
人类胎儿在六个月的时候,全身密布羊毛般的细毛,这就提供了一个更加奇妙的事例。在五个月的时候,眉端和脸上的毛、特别是口部周围的毛开始发育,口部周围的毛比头上的毛还要长得多。埃舍里希特(Eschricht)曾观察到一个女胎儿生有这种小胡子, (41) 但这件事情并不像最初看来那样令人惊奇,因为在生长早期男女两性的一切外在性状一般都是彼此类似的。胎儿身体所有部分的毛的趋向和排列同成年人的一样,不过受更大的变异性所支配。整个皮肤表面,甚至前额和双耳,都有毛密布其上;但有一个意味深长的事实,即,手掌和足蹠则完全是裸而无毛的,有如大多数低等动物的四个足蹠一样。因为这简直不能是一种意外的巧合,所以人类胎儿的羊毛般的覆毛大概代表着那些生来就是多毛的哺乳动物的最初永久性的毛皮。关于人生下来在其整个体部和面部就密布着细而长的毛,曾经记载过三四个事例;这一奇怪的状态是强烈遗传的,而且同牙齿的畸形相关。 (42) 亚历山大·勃兰特(Alex.Brandt)教授告诉我说,他曾将一位具有这样特性的35岁男人的面毛与一个胎儿的胎毛做过比较,发现它们在组织上是完全相似的;所以,如他所指出的,这种事例可以归因于毛的发育受到抑制以及它的继续生长。儿童医院里的一位外科医生向我保证说,许多病弱的儿童在背部生有十分长的细毛;这等事例大概可以纳入同一个问题之下。
最靠后的那个臼齿,即智齿,在人类比较文明的种族中好像有变为残迹的倾向。这等齿比其他臼齿小得多,黑猩猩和猩猩的相应齿也是如此;而且它们只有两个分叉的牙根。到17岁左右,它们才穿出牙龈,有人向我保证,它们远比其他齿容易龋坏,而且脱落也要早得多;不过有些著名的牙医否认这一点。它们还远比其他齿容易变异,无论在构造上或是在它们的发育时期上都是如此。 (43) 在黑色人种(Melanian races)中,智齿通常具有三个分叉的牙根,而且一般健全;它们同其他臼齿在大小上还有差别,不过其差别要比在高加索种族(Caucasian races)中要小。 (44) 沙夫豪森(Schaaffhausen)教授以文明种族的“颚的后齿部一直在缩短” (45) 来解释各种族之间的这种差别,我设想,可以把这种缩短归因于文明人惯常地吃软的和煮过的食物,这样,他们就较少使用颚部。勃雷斯(Brace)先生告诉我说,在美国把儿童的某些臼齿拔掉,已成为十分普通的常事,因为颚部长得不够大以容纳完全发育的正常齿数。 (46)
关于消化道,我看到一则报道,记载着唯一的残迹物,即盲肠的蛆型附属物。盲肠为肠部的一个分支或膨部(diverticulum),末端成一盲管(Cul-de-Sac),在许多以植物为食的低等哺乳动物中,它是极其长的。在有袋的树袋熊(koala) (47) 中,其盲肠实际上要长于整个体部的三倍以上。 (48) 它有时延长而成为一个长的逐渐变细的尖端,而且有时部分阻塞。看来好像是食物或习性的改变,致使各种动物的盲肠才大大地缩短了,蛆型附属物作为缩短部分的残迹物而被留下来了。我们从这一附属物的小型以及根据卡内斯垂尼就人类盲肠变异性所搜集的证据 (49) ,可以推论出这一附属物是一种残迹物。偶尔它完全不存在,偶尔却非常发达。其通道全长的一半或三分之二已完全闭塞,末端为一扁平实心的膨胀体。猩猩的这种附属物是长而盘曲的;人类的这种附属物从短的盲肠一端长出,其长度通常为4~5英寸,其直径仅为l/3英寸左右。它不仅是无用的,而且有时是致死的原因;关于这样的事,我最近听到两个例子;这是由于小而硬的东西,例如种子,进入它的通道而引起炎症所致。 (50)
在某些低等的四手类中,在狐猴科(Lemuridae)动物中,在食肉类动物中,以及在许多有袋类动物中,有一个孔道位于上膊骨(humerus)的下端附近,叫做上髁状突起孔(Supra-condyloid foramen),前肢的大神经从此孔通过,大动脉也常常从此孔通过。人类的上膊骨一般都有这一孔道的残迹,它有时发育得相当良好,由一个下垂的钩状骨突形成它的一部分,并由一束韧带使其成为一个完善的孔。曾经密切注意过这个问题的斯特拉瑟斯(Struthers)博士 (51) 现在阐明这一特性有时是遗传的,因为有一位父亲有此特性,在他的7个孩子中不下4人也有此特性。当这一孔道存在时,大神经一律要通过那里,这就明显地表示了,它是低等动物上髁状突起孔的同源部分和残迹物。特纳教授估计,如他告诉我的,现今人类骨骼有这一孔道的约占1%。但是,如果人类这一构造的偶尔发育是由于返祖——看来这似乎是可能的,那么它是返归到很远古的状态,因为它在高等四手类中是不存在的。
人类的上膊骨偶尔还有另一个孔,可以称为髁间(inter-condyloid)孔。这个孔发生于各种类人猿以及其他猿类 (52) ,但不经常,许多低于人类的动物也有此孔。值得注意的是,人类有此孔的在古代要比在近代多得多。关于这个问题,巴斯克(Busk) (53) 先生搜集了如下的证据:布罗卡(Broca)教授谈到,“在巴黎的南方墓地中搜集到的臂骨中,具有这个孔的占4%~5%;奥罗尼洞窟(Grotto of Orrony)的遗物属于青铜器时代,那里的32只上膊骨中就有8只具有这个孔;不过他认为这一异常大的比例可能是由于这个洞窟是一种‘家族墓地’。还有,杜邦(M.Dupont)在属于驯鹿时代(Reindeer period) (54) 的莱塞(Lesse)山谷的洞穴中发现有30%的骨有这个孔;勒盖(M.Leguay)在阿尔让特伊(Argenteuil)的史前墓的遗迹(dolmen)中看到25%的骨有这个孔;普律内尔-贝(M.Pruner-Bey)发现来自沃雷阿尔(Vauréa1)的骨有这个孔的占26%。可不要忽视普律内尔-贝所说的,在瓜契(Guanche)的骨骼中这种状态是普遍的”。在这个场合或另外几个场合中,古代种族比近代种族更加时常呈现一些类似于低于人类的动物的构造,这是一个有趣的事实。一个主要的原因似乎是,古代种族在漫长的系统线上距他们遥远的动物般的祖先,站得多少要近一些。
人类的尾骨以及下述某些其他椎骨,虽然已经没有作为尾巴的功能,却明显地代表着其他脊椎动物的这一部分。在胚胎的早期,它是游离的而且超出足部之外;如人类胚胎图(图1)所示。在某些罕见的、异常的场合中, (55) 据知甚至在降生后,还会形成一个尾状的外在小残迹物。尾骨是短的,通常只包含四个椎骨,所有都胶和在一起;这些椎骨都处于残迹状态,因为除去基部的一节外,其余仅由椎体(centrum)构成 (56) 。它们附有一些小肌肉;特纳教授告诉我说,其中的一块小肌肉曾被锡尔(Theile)明确描述为尾部伸肌(extensor)以残迹状态而重现,这块肌肉在许多哺乳动物中是非常发达的。
人类的脊髓仅仅伸延到最后一个脊椎(dorsal vertebra),即第一腰椎(1umbar vertebra);但一种线状构造(中尾丝,filum terminale)却沿着脊髓管的荐骨部分的轴、甚至沿着尾骨之背,向下伸延。这种线状体的上部,如特纳教授告诉我的,无疑是同脊髓同源的,而其下部显然纯粹是由软脑脊膜(pia mater)、即脉络被膜(vascular investing membrane)构成的。甚至在这时还可以说尾骨具有像脊髓这样一种重要构造的残迹,虽然它已不再被关闭在骨道之中了。下述事实仍承蒙特纳教授告知,它阐明尾骨同低等动物的真尾是多么密切地一致;卢施卡(Luschka)最近发现在尾骨之端有一个很特别的卷曲体,与中部的荐动脉(Sacral artery)相连接;这一发现引导克劳斯(Krause)和迈耶对一只猴(猕猴)和一只猫的尾进行了考察,发现在二者之中都有相似的卷曲体,但不是位于尾端。
生殖系统提供了各式各样的残迹构造,但这等残迹构造在一个重要方面同上述事例有所不同。这里我们所涉及的并不是某一物种的处于无效状态的那一部分的残迹,而是在某一性别中是有效的、在另一性别中却仅仅是残迹的那个部分。尽管如此,根据各个物种是分别创造的信念,此等残迹物的出现有如上述事例,还是难于解释的。此后我势必还要谈到这些残迹物,并将阐明它们的存在一般仅仅依靠遗传,这就是说,依靠某一性别所获得的部分曾被不完全地传递给另一性别。在这里我所举出的不过是几个有关这等残迹物的事例而已。众所周知,所有哺乳动物的雄者,包括男人在内,都有残迹的乳房。在几个事例中,这等残迹的乳房变得十分发达,而且分泌丰富的乳汁。它们在男女两性中本质上是相等的还可由下述事实得到阐明,即,在感染麻疹期间,男女双方的乳房都偶尔呈交感的增大。可以观察到,许多雄哺乳动物都有前列腺囊(vesicula prostatica),现已普遍承认它同雌者的子宫以及与其相连接的管道都是同源的。读过洛伊卡特(Leuckart)对这个器官所做的富有才华的描述和他的推论,而不承认他的结论的正确性,是不可能的。这在那些具有分叉的真正雌性子宫的哺乳动物中尤其明显,因为这等雄性哺乳动物的前列腺泡同样也是分叉的。 (57) 在这里还有另外一些属于生殖系统的残迹构造 (58) 可以引述。
现在列举的这三大类事实的意义是清楚明白的。但是,再反复陈述我在《物种起源》中详细提出来的一系列论点,就完全是多余的了。如果我们承认同科的诸成员来自一个共同的祖先,并且承认它们继此之后曾适应于多种多样的外界条件,那么,同科诸成员整个身体的同源构造就是可以理解的了。根据任何其他观点,则人或猴的手、马的足、海豹(Seal)的前肢、蝙蝠的翼等等之间的模式何以相似,就是完全不可解释的了。 (59) 断言他们一切都是按照一个同样的理想计划而造成的,并不是一个科学的解说。关于发育,根据这样的原理,即变异是在很晚的胚胎时期中随后发生的,而且是在相应的时期中遗传的,那么,我们就能清楚地理解为什么那些差异大得惊人的类型,其胚胎依然多少完全地保留着它们共同祖先的构造。人的、狗的、海豹的、蝙蝠的、爬行动物的胚胎彼此之间最初简直无法被区别开,对这样奇异的事实,从来没有过任何其他解释。为了理解残迹器官的存在,我们只能假定先前的一位祖先曾具有完善状态的这等部分,并且在生活习性改变了的情况下大大地缩小了,这或者是由于简单的不使用,要么就是由于那些最少是受多余部分之累的个体受到了自然选择,而且得到上述其他手段的帮助。
这样,我们就能理解,为什么人类和其他脊椎动物都是按照同样的一般模型被构成的,为什么他们都通过同样的早期发育阶段,而且,为什么他们都保留着某些共同的残迹物。因此,我们就应该坦白地承认它们的由来的共同性;如果接受其他观点,则无异于承认我们的构造以及我们周围的所有动物的构造仅仅是设下的一个陷阱以诱使我们的判断落入其中。如果我们注意一下整个动物系统的成员,并且考虑一下从它们的亲缘关系或分类、它们的地理分布和地质上的演替所得到的证据,上述结论就被大大加强了。使我们祖先宣称他们是从半神半人传下来的后裔,并且引导我们去反对上述结论的,不过是我们蒙昧的偏见和骄傲自大而已。但是,终有一天不久会到来,到那时,十分熟悉人类和其他哺乳动物的比较构造和比较发育的自然学者们如果还相信各个物种乃是分别创造作用的结果,那就会被认为是奇怪的事了。
第二章 人类自某一低等类型发展的方式
人类身体和心理的变异性——遗传——变异性的原因——人类的变异法则和低于人类的动物的一样——生活条件的直接作用——各部分增强使用和不使用的效果——受到抑制的发育——返祖——相关变异——增长的速度——对增长的抑制——自然选择——人类是世界上最占优势的动物——人类身体构造的重要性——导致人类变为直立的诸原因——由于直立而发生的构造变化——犬齿的缩小——头骨的增大及其形状的改变——无毛——无尾——人类不能自卫的情况
显然,人类现今依然受强大的变异性所支配。在同一个种族中没有任何两个人是完全相像的。我们不妨把无数面孔加以比较,而一个面孔一个样。在人类身体各部分的比例和大小方面也有同等大量的多样性,腿的长度是最易变异的诸点之一。 (60) 虽然在世界的某些地区一种长头颅是普遍的,在另外一些地区一种短头颅是普遍的,但是,甚至在同一个种族的范围内头的形状还有巨大的多样性,如美洲和澳洲南部地区的土著居民就是这样——后一种族的“血统、风俗以及语言在现存各种族中大概是最纯粹、最均一的”,甚至像区域如此狭窄的桑威奇群岛(Sandwich Islands)上的居民也是如此。 (61) 一位著名牙科医生向我保证说,牙齿的巨大多样性差不多同面貌的一样。主动脉如此常常地在歧路上循行,以致发现从1040具尸体中计算出循行路线的出现次数对解决外科问题是有用的。 (62) 肌肉是显著容易变异的,例如,特纳教授发现,在50具尸体中没有两具尸体的足部肌肉严格地相似,在有些尸体中其离差是相当大的。 (63) 他接着说,司掌运动的能力一定适当地按照若干离差而有所改变。伍德(J.Wood)先生曾做过如下记录:在36具解剖用的尸体中,有295个肌肉变异,在另一组同样数目的解剖用尸体中不少于558个变异,而且在身体两侧发生的变异只作为一个计算。 (64) 在后一组36人中,“查明并无一人与解剖学教科书中所做的肌肉系统的标准描述完全一样”。其中一个尸体竟有25个独特的畸形肌肉,数目之大,令人惊奇。同一块肌肉有时以多种方式发生变异,例如,麦卡利斯特(Macalister)教授描述过副掌肌(palmaris accessorius)的独特变异不少于20个。 (65)
著名的老一辈解剖学者沃尔夫(Wolff)坚决主张,内脏比外部器官更容易变异,“人体内部没有一部分不变异的”。 (66) 他甚至写过一篇专论,陈述如何选择内脏的典型标本作为代表。他讨论了肝、肺、肾等的至美,犹如人类外貌的至美一样,这一讨论听起来够奇怪的了。
人类同一种族心理官能(mental faculties)的变异性或多样性是如此为大家所熟知,以致无须在这里多赘,至于人类不同种族之间的更大差异,就更不必谈了。低等动物也是如此。所有管理过动物园的人们都承认这一事实,而且我们在家狗以及其他家养动物中可以明显地看到这一点。布雷姆特别坚决主张,他在非洲驯养的那些猴中,每一个猴都有它自己特殊的气质和脾气;他提到有一只狒狒,以它的高度智力而著称;伦敦动物园管理员曾向我指点过一只属于新世界(New World) (67) 的猴,同样以它的智力而著称。伦格尔(Rengger)也坚决主张,他在巴拉圭所养的同种的猴的各种心理特征(mental characters)也是多式多样的,他接着说,这种多样性一部分是先天的,一部分是它们受得何种待遇或教育的结果。 (68)
关于遗传的问题,我在他处 (69) 已经做过非常充分的讨论,在这里简直没有什么再需要补充的了。关于人类最细微的以及最重要的性状之遗传,我们所搜集到的事实比对任何低等动物的都多;虽然关于后者的事实也足够丰富的。至于心理属性(mental qualities)亦复如此,它们在家狗、家马以及其他家养动物中的遗传也是显著的。除了特别的嗜好和习性以外,一般的智力、勇气、坏脾气和好脾气等等肯定都是遗传的。至于人类,我们在差不多每一个家族中都可以看到相似的事实;通过高尔顿(Galton) (70) 先生的令人钦佩的工作,我们现在知道,天才也倾向于遗传,所谓天才就是高度才能的异常复杂的结合;另一方面,同样地,癫狂以及退化的心理能力肯定也在一些家族中得到遗传。
关于变异性的原因,就所有情况来说,我们都是很无知的;但我们还能够领会,在人类和低等动物中变异性的原因同各个物种在若干世代间暴露于其中的外界条件有某种关系。家养动物比那些处于自然状况下的动物更多变异;这显然是由于支配它们的外界条件性质的多样化和变化所致。在这方面,不同的人类种族同家养动物相似,同一个种族的各个人当居住在像美洲那样的辽阔地域时也同家养动物相似。我们看到在比较文明的民族中多样化的生活条件所产生的影响;因为属于不同阶层的而且从事不同职业的成员比野蛮民族的成员有更大的性格差距。不过,未开化人的一致性往往被夸大了,而且在某些场合中简直不能说有这种一致性存在。 (71) 即使我们仅注意到人类所暴露于其中的外界条件,要说人类远比任何其他动物更加“家养化” (72) ,也是一种错误。有些未开化种族,如澳洲人,并不及分布范围辽阔的许多物种的生活条件更为多样化。在另一个远为重要的方面,人类与任何严格家养的动物都大大不同;因为人类的生育从来没有通过有计划的或无意识的选择而长期受到控制。没有一个人类的种族或个人会被另外的人所完全征服,以至某些个人由于以某种方式在对主人有用方面胜过他人而被保存下来,这样便受到了无意识的选择。除去普鲁士掷弹兵那个著名的事例外,没有某些男人和女人被有意识地挑选出来而令其婚配;在普鲁士掷弹兵这一事例中,就像可以预期的那样,人服从于有意识选择的法则;因为有人断言,在掷弹兵及其高个子妻子所居住的村庄中曾经养育出许多高个子的人。在斯巴达(Sparta),也采用过一种选择方式,因为曾经颁布过这样的法律:所有婴儿在降生后不久就应受到检查,外貌良好而健壮的被保存下来,其余的则任其死亡。 (73)
如果我们把所有人类种族都视为单独一个物种,那么其分布范围是非常广阔的;不过有些与世隔离的种族,如美洲印第安人和波利尼西亚人(Polynesians),也有很广阔的分布范围。有一条法则是众所周知的,即分布范围广阔的物种远比分布范围狭窄的物种更加容易变异得多;把人类的变异性同分布范围广阔的物种的变异性相比较,比把他们同家养动物的变异性相比较,更加准确可靠。
看来人类和低于人类的动物的变异性不仅是由同样的一般原因所诱发的,而且二者身体的相同部分也以密切近似的方式受到影响。这一点已由戈德隆(Godron)和夸垂费什(Quatrefages)充分详细地予以证明了,所以在这里我只要提一下他们的著作 (74) 就行了。逐渐成为轻微变异的畸形在人类和低于人类的动物中同样也是如此相似,以致同样的分类和同样的名词可以通用于二者,小圣·伊莱尔(Isidore Geoffroy St.-Hilaire) (75) ,对此已有所阐明。我在有关家养动物变异的那部著作中,曾试图以粗略的方式把变异的法则安排在如下的各个项目中:改变了的外界条件直接而一定的作用,这种作用可以由同一物种的一切个体或几乎一切个体在同样环境中按照同样方式发生变异而被显示出来。各个部分长期连续使用和不使用的效果。同源部分的结合。复合部分的变异性。生长补偿。不过关于这一法则,我还没有找到有关人类的良好事例。某一部分对另一部分的机械压迫的效果,如婴儿的颅骨在子宫中所受到的骨盆压迫。发育的抑制,导致诸部分的缩小或其生长受到抑制。通过返祖,长久亡失性状的重现。最后,相关作用。所有这些所谓的法则可以同等地应用于人类和低等动物,其中大多数法则也可以应用于植物。在这里对所有这些法则一一加以讨论将是多余的。 (76) 不过其中有几项法则是如此重要,以致还必须以相当篇幅加以讨论。
改变了的外界条件直接而一定的作用
这是一个最错综复杂的问题。不可否认,改变了的外界条件对所有种类的生物都会发生某些作用,有时是相当大的作用;最初看来很可能是,如果有充足的时间,这将是必然的结果。但是我没有能够得到支持这一结论的明显证据;而在相反方面却可以提出正当的理由,至少有关适应于特殊目的的无数构造是如此。然而无可怀疑,改变了的外界条件可以引起几乎无限的彷徨变异,而整个组织因此在某种程度上变为可塑的了。
在美国,参加最近一次战争的一百万以上的士兵受到了身体测量,而且对他们的降生和成长时所在的州进行了登记。 (77) 根据这一数量大得惊人的观察,可以证明某种地方性影响对身材直接发生作用;我们进一步认识到,“大部分身体成长时所在的州以及表明其祖先系统的降生时所在的州似乎对身材有显著影响”。例如,已经证实,“当成长时居住在西部各州,有使身材增高的倾向”。另一方面,海军生活会延缓其成长,正如下述所阐明的那样,“十七八岁的陆军士兵和海军士兵的身材有巨大差别”。古尔德先生力图查明这样对身材发生作用的影响因素的性质,但他只得到了反面的结果,即,它们同气候、土地高度、土壤没有关联,甚至同生活的富裕和贫困也不以任何支配的程度相关联。这后一结论用维勒美(Villermé)根据法国不同地方应征士兵身高的统计所得出的结论直接相反。如果我们把波利尼西亚酋长和同岛低层人民的身材差别加以比较,或者把肥沃的火山岛居民和同海洋 (78) 低处的荒瘠珊瑚岛居民的身材差别加以比较,再把生活资料很不相同的火地岛(Tierra de Fuego)东海岸和西海岸居民的身材差别加以比较,那就不可避免地会得出如下结论:较好的食物和较大的生活舒适确可对身材产生影响。但是,上面的叙述阐明了要得出任何确切的结果是何等困难。比多(Beddoe)博士最近证明,关于英国居民,城市生活和某种职业对其身高有一种退化的影响;并且他推论这一结果在一定程度上是遗传的,在美国同样也有这种情况。比多博士进一步相信,“如果一个种族的身体发达到最高顶点,其身体精力和精神活力也要升到最高峰”。 (79)
外界条件对人类是否产生任何其他作用,现在还不知道。可以预料,气候的差异将会发生一种显著影响,因为肺和肾在低温下的活动要加强,而肝和皮肤在高温下也是如此 (80) 。以前认为,皮肤的颜色和毛发的特性是由光或热来决定的;虽然简直无法否认由此产生的某种效果,但几乎所有观察家们现在还一致认为这种效果是很小的,即使多年暴露于其中也是一样。在我们讨论人类不同种族的时候,还要对这个问题进行更适当的探讨。关于家养动物,有理由相信寒冷和潮湿对毛的生长有直接影响;但是,在人类的场合中,我还没有遇到任何有关这个问题的证据。
各部分增强使用和不使用的效果
众所周知,使用可以使一个人的肌肉加强,而完全不使用,或破坏其专有的神经,则可使肌肉减弱。当眼受到破坏时,视神经常常变得萎缩。当一条动脉被结扎时,其侧脉管的直径不仅在增大,而且管壁的厚度和强度也有所增加。当一个肾因病停止作用时,另一个肾就要增大,而且加倍地工作。骨如果负担较大的重量,不仅厚度、而且长度都有所增加。 (81) 经常从事不同职业可导致身体各部分的比例发生变化。例如,美国“联邦委员会”(United States Commission) (82) 查明,参加最近这次战争的海军士兵的腿比陆军士兵长出0.217英寸,虽然海军士兵平均要矮些;而海军士兵的手臂却短1.09英寸,所以,就其矮缩的身高而言,手臂的减短已越出了比例。海军士兵手臂的减短显然是由于它们的使用较多所致,这是一个料想不到的结果;但海军士兵的手臂主要用于拉牵,而不是用于支持重量。海军士兵的颈围和脚面厚度均较陆军士兵的为大,但其腰围、胸围和臀部则较小。
如果在许多世代中都遵循同样的生活习性,那么,上述几种变异是否会变为遗传的,还不知道,但这是可能的。伦格尔 (83) 把巴拉瓜河流域的印第安人(Payaguas Indians)的细腿和粗臂归因于他们一代连一代地几乎在独木舟中过一辈子,而下肢无所运动。另外一些作者对相近的事例作出了相似的结论。按照曾同爱斯基摩人长期在一起生活的克兰兹(Cranz) (84) 的说法,“当地人相信捕捉海豹时的机灵和敏捷(他们最高的技艺和美德)是遗传的,这确乎有些道理,因为一位著名的海豹捕捉手的儿子虽然幼年丧父,也显示了他的英雄本色”。在这一场合中,看来心理能力和身体构造都同等多地得到了遗传。有人断言,英国工人婴儿的手在降生时大于贵族婴儿的手。 (85) 根据四肢发育和颚部发育之间所存在的相关作用——至少在某些场合中是如此, (86) 那些不大用手和脚劳动的阶级,其颚部由于这种原因可能缩小。优雅文明人的颚部一般小于辛勤劳动者或未开化人的颚部,乃是确定无疑的。但是,关于未开化人,如赫伯特·斯宾塞(Herbert Spencer) (87) 先生所说的,在咀嚼粗糙的、未烹调的食物时较多地使用颚部,将会以一种直接方式对咀嚼肌及其所附着的骨发生作用。远在降生以前的胎儿,其足蹠的皮肤比身体其他任何部分的皮肤都厚; (88) 简直不能怀疑,这是由于压力在一长系列世代中的遗传效果。
众所周知,钟表匠和雕刻匠容易近视,常过户外生活的人,特别是未开化人,一般是远视的。 (89) 近视和远视肯定都有遗传的倾向。 (90) 同未开化人相比,欧洲人的视力以及其他感官都较差,这无疑是在许多世代中减少使用之积累的和遗传的结果;因为伦格尔说过,他曾反复地观察过同未开化的印第安人一起长大的、并同他们一起度过终生的欧洲人;尽管如此,这些欧洲人的感官在敏锐性上还不能同印第安人的相比。 (91) 同一位自然科学者还观察到,美洲未开化人头骨上容纳几种感觉器官的腔比欧洲人的为大;这大概暗示着这等器官本身在大小上的相应差异。布鲁曼巴哈也曾谈到美洲未开化人头骨上的鼻腔很大,并且把这一事实同其显著敏锐的嗅觉能力联系在一起了。按照帕拉斯(Pallas)的材料,北亚平原上的蒙古人具有异常完善的感官;普里查德相信,他们的穿过颧骨(zygomas)的那一部分头骨非常宽阔,系由于他们的感觉器官高度发达所致。 (92)
奎丑印第安人(Quechua Indians)居住在秘鲁的巍峨高原上;杜比尼(Alcide d'Orbigny)说,由于不断地呼吸稀薄的空气,他们获得了异常大的胸和肺。其肺部细胞也比欧洲人的大而多。 (93) 这些观察材料曾受到怀疑,不过福布斯(D.Forbes)先生对一个近似种族亚马拉人(Aymaras)进行过多次身体测量,他们也在10000到15000英尺的高地上生活;他告诉我说,他们在身体的粗细和长短方面都同他所看见过的所有其他种族的人有明显的差别。 (94) 在他的测量表格中,每一个人的身高定为1000,其他测量数据则按此标准缩减。该表说明,亚马拉人的伸直的双臂比欧洲人的为短,比黑人的更短。同样地,他们的双腿也较短;他们表现了这样一种显著的特征,即每一个受到身体测量的亚马拉人,其股骨(femur)比胫骨(tibia)为短。平均计算,股骨长度同胫骨长度之比为211∶252,而同时受到测量的两个欧洲人,其股骨长度同胫骨长度之比则为244∶230,在三个黑人中,其比例为258∶241。同样地,前肢的肱骨也相对的要比前臂短些。和身体最近的四肢那一部分的这样缩短,如福布斯先生向我提示的,似乎是同躯干长度大大增加有关的一种补偿的情形。亚马拉人还呈现一些其他独特的构造之点,例如,脚后跟的凸出部分很小。
这些人如此彻底地适应了他们的寒冷而高峻的居住地,先前西班牙人把他们带到低下的东部平原时,现在为高工资所诱、下来淘金时,以致死亡率有了可怕的提高。尽管如此,福布斯先生还找到了少数幸存了两代的纯粹家族;他观察到,他们依然遗传了其固有的特性。但是,甚至用不到测量,也可明显看出这些特性完全缩小了;通过测量,发现他们的身体已不像居住在高原者的身体那样长;同时他们的股骨却变得多少长一些,胫骨也有所增长,但程度较轻。至于实际测量数据,查阅福布斯先生的研究报告便知。根据这些观察,我以为毫无疑问的是,在一个非常高的地方居住了许多世代,直接地和间接地有诱使身体比例发生遗传的变异的倾向。 (95)
人类在其后期生存阶段,虽然通过诸部分的增强使用或减弱使用没有发生很大变异,但以上所举的事实阐明,人类在这一方面的倾向并没有消失。我们确知,同样的法则也适用于低等动物。因此,我们可以推论,当人类的祖先在远古时代处于变迁状况之下时,并且当他们由四足动物变成两足动物时,身体不同部分的增强使用或减弱使用的遗传效果很可能对自然选择起了很大的帮助作用。
发育的抑制
受到抑制的发育同受到抑制的生长有所不同,诸部分在前一状况下继续生长、同时依然保持其早期状态。各种畸形可以纳入这一项目之下,有些畸形,如裂口盖(cleft-palate)据知是偶尔遗传的。对我们的目的来说,只要谈谈沃尔格的研究报告 (96) 中所描述的畸形小头白痴的受到抑制的脑部发育就足够了。他们的头骨较小,而且脑旋圈(convolutions of the brain)不及正常人的复杂。额窦(frontal sinus),即眼眉上部的突起,非常发达,颚部以“异常”的程度向前突出,所以这等白痴同人类的低等模式多少相类似。他们的智力以及大多数心理官能,都极其薄弱。他们不能获得说话的能力,而且完全不能长久地注意,但很善于模仿。他们是强壮的,而且显著地活泼,不停地嬉戏、跳跃和做鬼脸。他们爬楼梯时常手脚并用,而且非常喜欢攀登家具和树木。这就使我们想起,几乎所有的小孩都喜欢爬树;这又使我们想起,原本为高山动物的小羔羊和小山羊多么欢喜在小丘上跳来跳去,不管这小丘是多么小。白痴在其他一些方面也同低于人类的动物相类似,例如,他们在吃每一口食物之前,都要小心地嗅味,对此已有几个事例的记载。有一个白痴被描述常用口帮助双手去捉虱子。他们的习性往往是猥亵的,没有礼貌的感觉;他们的身体显著多毛, (97) 关于这一点曾经发表过几个事例。
返 祖
这里所举的事例有许多大概可以纳入上述项目之下。如果一种构造在其发育时受到了抑制,但仍继续生长,直到它和同一类群(group)的某些成熟的低等成员的相应构造密切类似,那么,在某种意义上就可把这一构造看做是一个返祖的事例。一个类群的低等成员对其共同祖先大致是如何构成的,向我们提供了某种概念;简直不能相信一个复杂的部分在胚胎发育的早期阶段受到抑制后,还会继续生长到终于可以执行其固有功能,除非它在某一较早的生存期间获得了这种能力,而现今异常的,即受到抑制的构造在那时还是正常的。一个畸形小头白痴的简单头脑,就其同一只猿的头脑相类似来看,在这种意义上可以说它提供了一个返祖的事例。 (98) 还有另外一些事例可以更加严格地纳入现在这个返祖项目之中。在人类所属于的那一类群的低等成员中正常发生的某些构造,偶尔也会在人类中出现,虽然在正常的人类胚胎中并没有发现过这等构造;或者,这等构造如果正常存在于人类胚胎中,但它们都变得异常发达,竟达到这一类群的低等成员的那种情况,虽然在后者这是正常的。下述例证将会使这些论述更加清楚明白。
各种哺乳动物的子宫都是由一个具有两个明显的孔和两个通道的双重器官,如在有袋动物中那样,渐渐变为一个单独的器官;它除去有一个微小的内褶以外,如在高等猿类和人类中那样,一点也不是双重的了。啮齿动物显示了这两个极端状态之间的一个完整的累进系列。所有哺乳动物的子宫都是由两个简单的原始的管发展而来的,在这两个管下方的部分形成了两个角;按照法尔(A.Farre)博士所说的,“这两个角在其下端的愈合,形成了人类的子宫体;在没有子宫中央部分或子宫体的那些动物中,这两个角依然保持不相愈合的状态。在子宫发展的进程中,这两个角逐渐变短,最后终至消失,或者可以说,它们被吸收入子宫体之中。”甚至像低等猿类和狐猴类那样的高等动物,其子宫依然具有两个角。
且说,成熟的子宫具有两个角或者部分地分为两个器官的这等异常事例在妇女中并不罕见;按照欧文的说法,这等事例再现了某些啮齿动物所达到的那种“集中发育(concentrative development)的阶段”。关于胚胎发育受到简单的抑制,以及继之而来的生长和完全的功能发育,我们在这里所看到的恐怕就是这种事例;因为这种局部双重的子宫每一边都能执行固有的妊娠功能。在另外一些更罕见的事例中,两个明显的子宫腔被形成了,每一个腔都具有它的固有的孔和通道。 (99) 在正常的胚胎发育期间从来不通过这等阶段;很难相信,虽然这也许不是不可能的,两个简单的、微小的、原始的管会知道如何(如果可以使用这一名词的话)生长成两个明显的子宫,每个子宫具有一个构造良好的孔和通道,而且还具有无数的肌肉、神经、腺和血管,如果它们不是像在现存有袋动物的场合中那样地曾在以往经历过一个相似的发育过程。谁也不会妄想,像妇女的畸形双重子宫那样的一种如此完善的构造仅仅是偶然的结果。但返祖原理——据此一种长久亡失的构造会被召回重生——大概可以用来说明其充分的发育,即使这种发育是在间隔了非常悠久的时光之后进行的。
卡内斯垂尼教授在讨论了上述事例以及各种相近的事例之后,得出了同上述一样的结论。他提出另外一个例子,是关于颧骨的, (100) 这种骨在某些四手类动物以及其他哺乳动物中正常是由两个部分构成的。当人类胎儿在两个月的时候,颧骨就是这种状态;通过发育的抑制,有时在成年人中、特别是在突颚的低等种族中还保留着这种状态。因此,卡内斯垂尼断定,人类某些古代祖先的这种骨一定正常地分为两个部分,而在以后却变得融合在一起了。人类的额骨是由单独一片构成的,但在胚胎中、在小孩中而且在差不多所有低等哺乳动物中,额骨是由两片构成的,由一条明显的缝分开。在人类达到成熟期之后偶尔还多少明显地保留着这条缝;这种情形在古代的头盖骨比在近代的头盖骨中更加常见,特别是如卡内斯垂尼所观察的,在那些从冰碛(drift)中发掘出来的、属于短头模式的头盖骨中尤其常见。在这里就像在颧骨的近似事例中那样,他再次得出了同样的结论。在这个事例中,以及在就要谈到的另外一些事例中,古代种族比近代种族在一定性状上,更加常常接近低等动物的原因,看来是由于后者在漫长的系统线上距离他们的早期半人类祖先多少要远一点儿。
人类的同上述多少相似的各种其他畸形,曾被不同作者提出来作为返祖的事例;不过对此等事例似乎还有不少疑问,因为,在我们发现这等构造正常存在以前,我们势必在哺乳动物系统中下降到极低的地位。 (101)
人类的犬齿是完全有效的咀嚼工具。但他们的真正犬齿性状,如欧文 (102) 所说的,为“齿冠呈圆锥形,其末端为一钝点,外面凸形,内面扁平或稍凹,内面基部有一个微小的突起。黑色种族、特别是澳洲土人最好地显示了这种圆锥形齿冠。犬齿较切齿埋植得更深,而且牙根更强固”。然而,对人类来说,这个齿已不再是为了撕裂敌物或猎物的特殊武器了;所以,就其固有的机能而言,不妨把它视为残迹的。在人类头骨的任何大型采集品中,如赫克尔 (103) 所观察的,总可以找到犬齿相当突出于其他齿之外的一些头骨,其方式就像类人猿的犬齿一样,不过程度较轻而已。在这等场合中,一颚的齿间空位是留待容纳另一颚的犬齿。瓦格纳(Wagner)所绘的卡菲尔人(Kaffir)头骨的齿间空位异常宽阔。 (104) 同近代头骨比较,古代头骨受到检查的非常之少,但至少已有三例表明前者犬齿非常突出,这确是一个有趣的事实;据说脑雷特人(Naulette)的颚是异常大的。 (105)
在类人猿中,仅雄者具有充分发达的犬齿;但在大猩猩(gorilla)中并且程度较轻地在猩猩(orang)中,雌者的犬齿也相当地突出于其他齿之外;所以,我所确信的妇女有时有相当突出的犬齿这一事实,对于相信人类犬齿偶尔非常发达乃是返归猿类般的祖先,并不是一个严重的障碍。如果有人轻蔑地拒绝相信他自己的犬齿形状以及其他人的偶尔非常发达的犬齿乃是由于我们早期祖先曾经装备有这等可怕的武器,那么他大概在冷笑中却揭示了他自己的由来。因为,人类虽然不再打算或者没有能力再使用这等齿作为武器,但还会无意识地收缩他的“嗥叫肌”(snarling muscles,贝尔爵士命名 (106) ),以便把犬齿露出,准备动作,就像一只狗准备咬架那样。
四手类或其他哺乳动物所固有的许多肌肉偶尔也会在人类发育。沃拉克威契(Vlacovich)教授 (107) 检查过40个男性尸体,发现其中19人具有一种被他称为“坐耻肌”(ischiopubic)的肌肉;3人具有代表这种肌肉的韧带(1igament);其余18人连一点这种肌肉的残迹都没有。在30个女性尸体中,仅有2人在两侧具有这种发达的肌肉,另外3人只有残迹的韧带。所以,这种肌肉在男性中看来远比在女性中普遍得多;根据人类起源于低等生物类型的信念,这个事实就是可以理解的了;因为在几种低于人类的动物中,曾发现过这种肌肉,凡是具有这种肌肉动物,它的唯一作用即在于帮助雄者的生殖行为。
伍德先生在他的一系列有价值的论文中, (108) 详细地描述了大量的人类肌肉变异,这等肌肉都同低等动物的正常构造相类似。同我们最近亲属四手类动物所正常存在的肌肉密切类似的那些肌肉真是多得举不胜举。有一具男性尸体,体格强壮,头骨构造良好,在其身上观察到的肌肉变异不下七处之多;所有这些都明显地代表了各种猿类所固有的肌肉。僻如说,这具男尸在颈部两侧各具一块直正的、强有力的“锁骨提肌”(1evator claviculae),就像我们在所有猿类中所看到的那样,据说在60具人类尸体有一具有这种肌肉。 (109) 再者,这具男尸还有“一块特殊的趾蹠骨展肌(abductor of the metatarsal bone),正如赫胥黎教授和弗劳尔(Flower)先生所阐明的,高等的和低等的猿类普遍具有这种肌肉”。我仅提出两个补充的事例;在所有低于人类的哺乳动物中都可以找到肩峰底肌(acromiobasilar muscle),而且它似乎同四足行路的步法相关, (110) 在60具人类尸体中有一具有这种肌肉。布雷德利(Bradley)先生 (111) 发现一个男人的两只脚都有一块“第五蹠骨展肌”(abductor ossis metatarsi quinti);在此之前没有记载过人类具有这种肌肉,但它在类人猿中却是永远存在的。手和臂——这是人类特性非常显著的部分——的肌肉极端容易变异,结果变得同低于人类的动物的相应肌肉类似。 (112) 这种类似或是完全的,或是不完全的;在不完全的场合中,它们显然具有过渡性质。某些变异在男人中较普遍,另外一些变异则在女人中较普遍,对此还不能举出任何理由。伍德先生描述了大量变异之后,作出如下意义深远的陈述:“在诸器官的沟中或沿着各个方面延伸的肌肉构造显著脱离正常的模式,一定暗示有某种未知的因素,这对于一般的、科学的解剖学的全面知识有极大的重要性。” (113)
这一未知因素乃是返归以往的生存状态,可以被认为是有最高度可能性的。 (114) 人类和某些猿类之间如果没有遗传的连接关系,要说一个人的畸形肌肉同猿类的肌肉相似者不下于七处之多,系出于偶然,则是完全不可相信的。另一方面,如果人类是从某一猿类般的动物传下来的,那么就举不出任何有根据的理由来说明某些肌肉为什么不会经过成千上万的世代之后而突然重现,其方式就像马、驴、骡经过数百代、更可能经过数千代在腿部和肩部突然重现其暗色条纹一样。
这等各式各样的返祖事例同第一章中所举出的残迹器官事例的关系是如此密切,以致把其中的许多事例放在那里或这里,都无可无不可。例如,一个具有角的人类子宫可以被说成是以一种残迹状态代表某些哺乳动物同一器官的正常状态。人类的某些残迹部分,如男女两性的尾骨以及男性的乳房,是一直存在的;还有另外一些残迹部分,如髁上孔(supracondyloid foramen),仅偶尔出现,因而可以被纳入返祖项下。这几个返祖的构造以及那些严格是残迹的构造揭示了人类是从某一低等类型以准确无误的方式传下来的。
相 关 变 异
人类就像低于人类的动物那样,他的许多构造如此紧密关联,以致当某一部分发生变异后,另一部分也要跟着发生变异;在大多数场合中,我们对此还不能举出任何理由。我们无法说,是否这一部分支配那一部分,或者,是否二者受某一较早发达的部分所支配。各种畸形,正如小圣·伊莱尔所反复坚决主张的那样,就是这样紧密地连接在一起的。同源构造特别容易一齐变异,就像我们在身体两侧和上下肢所看到的情形那样。梅克尔很久以前就曾说过,当手臂肌肉脱离其固有模式时,它们几乎永远模拟腿部的肌肉;相反的,腿部肌肉也是如此。视觉器官和听觉器官,齿和毛,皮肤和毛的颜色,体色和体质,或多或少都是相关的。 (115) 最初是沙夫豪森教授注意到显然存在于肌肉结构和眶上脊(supra-orbital ridges)之间的关系,眶上脊乃是人类低等种族的显著特性。
除了多少可能地纳入上列项目中的变异以外,还有一大类可以暂时称为自发的变异,因为就我们的无知程度来说,它们似乎是在没有任何激发的原因下发生的。然而可以阐明,这等变异无论具有微小个体差异,或者具有强烈显著而突然的构造离差,其决定于生物体质远比决定于它所处的外界条件性质要多得多。 (116)
增 长 速 度
文明国家的人口据知在适宜条件下,如在美国,25年可增加一倍;按照尤勒(Euler)的计算,12年稍微多一点就可增加一倍。 (117) 如果按照前一项增长速度,则现今的美国人口在657年间就会如此稠密地布满整个水陆形成的世界,以致四个人只能占一平方码的面积。对人类不断增长的首要的或基本的抑制,是获得生活资料以及舒适生活的困难。根据我们所看到的,可以推论情况确系如此,例如在美国,那里生活容易,而且有大量房屋。如果在大不列颠(Great Britain)这等生活手段突然加倍,那么我们的人口也会迅速加倍。在文明国家里,这种首要的抑制主要是以限制婚姻来完成的。在最贫困的阶级中,婴儿的较大死亡率也是很重要的;拥挤而蹩脚的房屋中的一切年龄的居民,由于各种疾病同样也有较高的死亡率。在位于适宜条件下的国家里,严重的流行病和战争很快会起到平衡作用,甚至超过平衡。移民也有助于暂时的抑制,但对极贫困的阶级来说,这在任何程度上都没有作用。
正如马尔萨斯(Malthus)所指出的,可以有理由设想,野蛮种族的生殖力实际上低于文明种族。关于这个问题,我们肯定不知道,因为对未开化人没有进行过人口调查;但从传教士以及其他同这等民族长久相处的人士所提出的一致证据来看,他们的家庭通常是小的,大家庭不多见。那里的妇女据信给婴儿哺乳的时期很长,上述情形由此或者可以得到部分的解释;但,高度可能的是,未开化人要经历很大的苦难,而且不会得到像文明人那样多的营养丰富的食物,他们的生殖力实际上大概要差一些。我在前一著作中 (118) 曾指出,所有我们家养的四足动物和鸟类以及所有我们的栽培植物,其能育性均比处于自然状况下的相应物种为高。动物突然被供给过剩的食物,或者长得很肥,以及大多数植物突然从很瘠薄的土地被移植到很肥沃的土地上,都会或多或少变为不育的;以此来反对上述结论是毫无根据的。所以,我们可以预期,文明人的生殖力要比野蛮人的为高,在某种意义上可以说文明人是高度家养的。文明民族的增高了的生殖力,就像我们的家养动物那样,变成一种遗传的性状,也是非常可能的;至少已经知道,人类产双胞胎的倾向在一些家族中是向下传递的。 (119)
尽管未开化人的生殖力低于文明人,毫无疑问他们还会迅速增加,如果他们的人数没有在某些方面受到严格限制。关于这个事实,桑塔尔人(Santali)或印度山区部落最近提供了一个良好的例证;因为,正如亨特(Hunter)先生所阐明的 (120) ,自从施行种牛痘、其他瘟疫有所缓和以及战争切实受到遏制之后,他们以异常的速度增加了。然而如果这等未开化人不是进入邻接地区做雇工,他们人口的增加大概是不可能的。未开化人几乎都结婚;但有某种谨慎的约束,因为他们一般都不在最早可能的年龄结婚。年轻人常常需要显示出他能够养活一个妻子后才可以结婚;他们一般需先挣得她的身价,以便从她父母那里把她买来。获得生活资料的困难,对未开化人远比对文明人以更加直接得多的方式偶尔限制其人口的数量,因为所有部落都周期地遭受饥饿的危害。这时,未开化人被迫去吃更恶劣得多的食物,他们的健康难免受到损害。关于他们在遭到饥饿之后或在饥饿期间肚子凸出和四肢消瘦,已经有过许多记载。于是,他们被迫到处游荡,如我在澳大利亚听到的,这时他们的婴儿要大量死亡。由于饥饿是周期的,主要决定于非常的季节,因而所有部落的人口数量一定波动很大。他们不能稳定地、有规律地增长,因为那里的食物供给不会人为地增加。未开化人当穷迫过甚时,就彼此侵犯领地,结果引起战争,其实他们同邻近部落的战争几乎不绝。他们在陆地和水上寻觅食物时,容易遇到许多意外事情;在一些地方他们受到猛兽的危害很大。在印度由于虎患,有些地区的人口减少了。
马尔萨斯曾讨论过这几种抑制,但他对很可能是其中最重要的一种抑制,即杀婴、特别是杀女婴以及堕胎,却强调得不够。今天,世界上许多地方都盛行此事;据伦南(M'Lennan)先生说,以往杀婴的规模还要更大。 (121) 此事的发生与其说由于未开化人认识到养活所有生下来的婴儿是困难的,毋宁说是不可能的。淫乱生活也可以加入上述抑制;不过这还是被作为生活手段所引起的;但有理由相信,在某些场合中(例如在日本),这是作为一种控制人口的手段而有意地受到鼓励。
如果我们回顾一下极其远古的时代,在人类还没有达到人的地位以前,他大概要比今天最低等的未开化人更多地受本能而更少地受理性所引导。我们早期的半人类祖先不会实行杀婴或一妻多夫制;因为低于人类的动物决不会如此违反常情, (122) 以导致它们经常地杀害自己的后代,或全然无所妒忌。那时婚姻不会受到谨慎的限制,男女双方在年龄很轻的时候就会自由结合。因此,人类的祖先就趋向于迅速增殖;但某种抑制,无论是间歇的或经常的,一定曾经使其数量下降,甚至比对现今的未开化人还要剧烈。这等抑制的确切性质是什么,我们还无法说出,就像我们对大多数其他动物的情况无法说出那样,我们知道,马和牛不是极其多产的动物,当最初纵放于南美时,它们便以极大的速率增殖。象,为所有已知动物中繁育最慢者,在几千年之内其子孙便可充满整个世界。各种猴的物种的增殖一定要受到某种方法的抑制;但不像布雷姆所说的,是由于猛兽的侵袭而受到抑制。谁都不会假设美洲的野马和野牛的实际繁殖力最初以任何明显的程度增大了,其后由于它们充满了各个地区,这同样的繁殖力便缩小了。毫无疑问,在这一场合以及在所有其他场合中,多种抑制同时发生作用,而且在不同的环境条件下有不同的抑制作用;因不良季节而发生的周期的饥饿可能是所有抑制中最重要的。对人类的早期祖先来说,亦复如此。
自 然 选 择
现在我们已经知道人类的身体和心理都是可变异的,这等变异就像在低等动物的场合中那样,是由同样的一般原因直接地或间接地所引起的,并且服从同样的一般规律。人类广布于地球的表面,在他们不断的迁徙期间 (123) ,一定接触到多种多样的外界条件。在这一半球的火地、好望角、塔斯马尼亚(Tasmania)的居民,以及在另一半球的北极地区(Arctic regions)的居民,当到达他们现今的家乡之前,一定经历过多种气候,而且多次改变了他们的习惯。 (124) 人类的早期祖先还像所有其他动物那样,一定趋向于增殖到超越他们的生存方法以外;所以他们一定不时进行生存斗争,因而要受严格的自然选择法则所支配。所有种类的有利变异将这样偶尔地或经常地被保存下来,而有害变异则被淘汰。我所指的并非强烈显著的构造离差,这只是间或发生的,我所指的仅是个体差异而已。例如,我们知道决定我们运动能力的手与足的肌肉,就像低等动物的那样 (125) ,是容易不断变异的。那么,如果居住在任何地方的、特别是居住在外界条件发生了某种变化的一处地方的人类祖先分为相等两部分,其中一部分的所有个人由于他们的运动能力最适应于获得生存资料或保卫自己,他们就会比天赋较差的另一部分平均存活的人数较多,而且生下来的后代也较多。
即使现今在最野蛮状况下生活的人类,也是这个地球上曾经出现过的最占优势的动物。他比任何其他高等生物类型分布更广;所有其他生物都屈服在他的面前。这种巨大的优越性显然归功于他的智能,而且归功于他的社会性——这引导他去帮助和保卫他的伙伴,同时还归功于他的身体构造。这等特性的异常重要已由生活斗争的公断所证明。通过他的智力,有音节的语言发展了;他的惊人的进步主要取决于此。正如昌西·赖特(Chauncey Wright)先生所指示的,“语言官能的心理分析阐明,语言的最小熟练程度比任何其他方面的最大熟练程度可能需要更多的脑力”。 (126) 他发明了而且能够使用各种武器、工具、陷阱等等,借此他保卫自己,杀死或捕捉动物,并且用其他方法去获得食物。他曾建造木筏或独木舟从事捕鱼,或渡海到邻近肥沃的岛屿。他曾发明取火的技术,借此,把坚硬而多纤维的植物根弄成可消化的,并且把有毒的植物根或根部以外的部分弄成无毒的。取火的发明始于有史以前,这大概是人类在语言以外的最大发明。这几种发明乃是他的观察、记忆、好奇、想象以及推理诸种能力的直接结果,处于最野蛮状况下的人类凭借这些发明就可以变为最优秀超群的了。所以,我不能理解华莱士(Wallace)先生为什么要主张:“自然选择只能把略优于猿类的脑赋予未开化人。” (127)
对人类来说,智力和社会性虽然具有最高的重要性,但我们必须不要低估他的身体构造的重要性,本章下余部分将专门讨论这个问题,关于智力和社会性即道德官能的发展,将在以下两章进行讨论。
甚至准确地使用锤子,也并非一件容易的事,每一个学过木工的人都会承认这一点。像火地人那样,把一块石头准确地投掷在目标上,以保卫自己或击毙鸟类,则需要手、臂、肩各种肌肉高度完善的协同动作,而且,进一步还需要敏锐的触觉。一个人投石或掷枪以及进行许多其他动作,必须双足站稳;这又需要许多肌肉的完善的相互适应。把一块燧石削成最粗糙的器具,或者用一块骨头制成钩枪或钓针,则需要使用完善的手;因为,正如最有才能的鉴定家斯库克拉夫特(Schoolcraft)先生 (128) 所指出的,把碎石片制成刀、矛或箭,表明要有“异常的才能和长久的实践”。原始人实行分工的事实在很大程度上证明了这一点;并非每一个人都制造他自己的石器或粗糙的陶器,而是某些人似乎专门从事这种工作,无疑用此来交换他人狩猎之所得。考古学者们相信,在我们祖先想到把削碎的燧石磨成光滑的器具之前,曾经历了非常悠久的岁月。几乎谁都不会怀疑,一种类人的动物,如果具有充分完善到可以准确投掷石块的手和臂,或者可以把一块燧石制成一种粗糙的器具,仅就机械技能而言,就能通过充分的实践制作文明人所能制作的差不多任何器物。在这方面,手的构造可以同发音器官的构造相比,猿类的发音器官用于发出各种带有信号的叫声,有一个属,可以发出音乐般的声调;但是在人类,密切相似的发音器官通过使用的遗传效果却适于发出有音节的语言了。
现在我们转来谈谈人类的最近亲属,也就是我们早期祖先最好的代表;我们发现四手类 (129) 的手是按照人手的同样一般形式构成的,但对多种多样用途的适应,则远远不够完善。用于行进,它们的手不及狗的脚;从黑猩猩和猩猩那样的猴类可以看到这种情形,它们用手掌的外缘或指关节(knuckles)行走。 (130) 然而它们的手却极好地适于爬树。猴类用拇指在一边、其余四指和手掌在另一边以抓住细树枝或绳索,其方式和我们的一样。这样,它们还能把相当大的东西,如瓶颈,举到嘴边。狒狒用手翻转石头,挖掘树根。它们可以用拇指对着其余四指抓住坚果、昆虫或其他小东西,这样,它们无疑还会从鸟巢中掏取鸟卵和小鸟。美洲猴用树枝碰打野生橙,直到果皮裂开,然后用双手的指头把果皮撕去。它们以一种狂暴的状态用石块把坚硬的果实砸开。其他猴用拇指把贝壳掰开。它们用手指拔出身上的树棘和果刺,而且彼此捉身上的寄生虫。它们从高处把石头滚下,或者向它们的敌对者投掷石块;尽管如此,它们在做各种这样动作时却非常笨拙,就像我亲眼所见的那样,它们完全不能准确地把石头投出去。
有人说,因为猴类“抓握东西非常笨拙”,所以“一种专化差得更多的抓握器官”对它们来说,其用处和它们具有现在那样的手是同等美好的; (131) 在我看来,这种说法非常不正确。相反,我看没有理由可以怀疑,一双构造更加完善的手,如果不致使它们因此爬树较差,大概对它们还是有利的。我们可以猜想,完善得像人类那样的手,大概不利于攀登;因为世界上大多数树栖的猴类,如美洲的蛛猴(Ateles)、非洲的疣猴(Colobus)以及亚洲的长臂猿(Hylobates),或者拇指缺损,或者足趾部分地结合,所以它们的四肢已变成纯粹用于把握的钩状物了。 (132)
灵长类(Primates)一大系的某些古代成员,由于谋生的方式发生变化,或者由于周围环境发生某种变化,一旦达到较少树栖的地步,它的惯常的行进方式就会跟着改变;这样,就要使它更加严格地四足行动或二足行动。狒狒出没于丘陵区或山区,只是在必要时,才攀登高树; (133) 所以它们获得了差不多像狗那样的步法。仅有人类变为二足动物,我以为,我们可以部分地了解他怎样取得最显著特性之一的直立姿势。没有手的使用,人类是不能在世界上达到现今这样支配地位的;他的手是如此美妙地适于按照他的意志进行动作。贝尔爵士坚决认为,“人手提供一切工具,手与智慧相一致便使人类成为全世界的主宰”。 (134) 但是,只要手和臂惯常地用于行进和支持身体的全部重量,或者,如上所述,只要手和臂特别适于爬树,那么,它们就几乎不能变得完善到足以制造武器或把石头和矛枪准确地投掷到目标上的程度。手的这种简单使用,还会使触觉变钝;而手的妙用大部分取决于触觉。仅仅由于这些原因,变为二足动物对人类也是有利的;不过双臂和整个身体上部的自由,对许多动作的完成乃是必不可少的;而且为了这个目的,他必须稳固地用脚站立。为了得到这种巨大利益,人类的脚变得扁平了;而且大足趾发生了特殊的改变,但这使它几乎完全失去了把握的能力。因为手变得完善到适于把握,脚就应变得完善到适于支承和行进,这同通行于整个动物界的生理分工原理是相符合的。然而在有些未开化人中,脚还没有完全失去它的把握能力,他们的爬树方式以及手的其他用法阐明了这一点。 (135)
无可怀疑,用脚稳固地站立以及手与臂的自由对于人类是一种利益,这已由他在生活斗争中的卓越成功所证明,那么,要说人类变得愈来愈直立或二足行动对他的祖先没有利益,我看是没有任何理由的。这样,他们便能用石块或棍棒去防卫自己,攻击他们所要捕食的动物,或用其他方法获取食物。从长远观点来看,构造最好的个体将会取得最大的成功,而且大量生存下来。如果大猩猩和少数亲缘关系密切的类型灭绝了,那么,可能会有这样的争辩:一种动物不能由四足的逐渐变为二足的,因为处于一种中间状态的所有个体都非常不适于行走;而这一争辩是具有巨大的说服力和明显的真实性的。但是我们知道(这是值得好好思考的),类人猿现在实际上是处于一种中间状态;而且总的看来,无疑它们是很适应于它们的生活条件的。例如,大猩猩以左右摇摆的蹒跚脚步奔跑,但在行进时通常是用两只弯垂的手来支撑。长臂猿有时把双臂用做好像拐杖一般,它们的身体在两臂之间悬摇而前,某些种类的长臂猿在不经教导的情况下就还算能迅速地直立而行或奔跑;然而它们行动笨拙,远远不及人走得稳当。总之,在现存的猴类中,我们看到一种介乎四足动物和二足动物之间的行进方式;但是,正如一位没有偏见的鉴定家 (136) 所坚决主张的,类人猿在构造上距离二足动物比距离四足动物更近。
由于人类的祖先变得越来越直立,他们的手变得越来越适于把握和其他用途,他们的脚和腿同时变得适于稳固地支撑和行进,所以构造上其他无穷的变化就是不可避免的了。骨盆势必加阔,脊骨特别弯曲,头安置在已经改变的位置上,一切这等变化都是人类所曾经完成的。沙夫豪森教授主张,“人类头骨上强有力的乳头状突起就是他的直立姿势的结果”; (137) 猩猩、黑猩猩等都没有这等突起,大猩猩的比人类的为小。在这里还可以接着谈谈与人类直立姿势有关联的各种其他构造。很难确定,这等相关变异有多大程度是由于自然选择的结果,有多大程度是由于某些部分增强使用的效果,或者,有多大程度是由于某一部分对另一部分所起的作用。毫无疑问,这等变化方式经常是协同进行的。例如,某些肌肉及其所附着的骨节当由于惯常使用而扩大时,这就阐明了某些动作是惯常进行的,而这等动作一定是有益的。因此,进行动作最好的个体乃有较多数量生存下来的倾向。
臂和手的自由使用,部分是直立姿势的原因,部分是其结果,这似乎以一种间接的方式导致了构造的其他改变。如前所述,人类的早期男性祖先大概具有大型的犬齿;但是,由于他逐渐获得了使用石块、棍棒或其他武器以同敌对者或竞争者进行战斗的习惯,他们就越来越少地使用他们的颌(jaws)与牙。在这种情况下,颌与牙将会缩小,无数近似的事例使我们感到差不多确实如此。在此后一章中,我们将会遇到非常类似的例子,表明雄性反刍动物犬齿的缩小和完全消失显然同角的发达有关系,而马类,则同它们用门牙和蹄进行斗争的习惯有关系。
正如吕蒂迈尔(Rütimeyer) (138) 和其他人所坚决主张的,在成年的雄类人猿中,正是由于颌肌的非常发达对头骨所产生的效果,才使它在许多方面同人类有如此重大差异,并且使这种动物的容貌确实可怕。所以,在人类祖先的颌与牙缩小之后,成年者的头骨就会越来越同现存人类的相类似。正如我们以后将要看到的,雄者犬齿的缩小几乎肯定要通过遗传对雌者牙齿发生影响。
由于心理官能的逐渐发达,脑几乎肯定要变大。我推测没有人会怀疑,人脑体积与其身体的比例大于大猩猩或猩猩的脑体积同其身体的比例,是与人类的高度心理能力密切关联的。在昆虫方面,我们遇到密切近似的事实:蚁类的脑神经节(Cerebral ganglia)异常之大,所有膜翅目(Hymenoptera)的脑神经节比智力较差的目、如甲虫的脑神经节要大许多倍 (139) 。另一方面,没有人会想象任何两种动物或两个人的智力可以由脑壳的容积准确地测定出来。肯定的是,有极小的一点纯粹的神经物质,就可进行非凡的心理活动。例如,蚁类令人吃惊的各种各样的本能、心理能力以及感情是众所周知的,而它们的脑神经节还不及一个小针头的四分之一那样大。从这个观点来看,蚁脑乃是这个世界上的最不可思议的物质原子之一,也许比人脑更加不可思议。
关于人脑的大小同智能的发达之间有某种密切关系这一信念,得到了未开化人和文明人的头骨比较以及古代人和近代人的头骨比较的支持,而且得到了从整个脊椎动物体系所看到的相似现象的支持。伯纳德·戴维斯(J.Barnard Davis)博士 (140) 根据许多仔细的测量证明了,欧洲人头骨的内容积为92.3立方英寸;美洲人为87.5;亚洲人为87.1;澳洲人仅为81.9立方英寸。布罗卡教授 (141) 发现,巴黎坟墓中的19世纪头骨大于12世纪墓穴中的头骨,其比例为1484:1426;而且根据测量所确定的,增大部分完全在头骨的前额——智能的活动中心。普里查德相信,不列颠的现代居民比古代居民具有“宽阔得多的脑壳”。尽管如此,还必须承认,有些极其远古的头骨,如尼安德特人(Neanderthal) (142) 的一个著名的头骨,也是非常发达而且宽阔的 (143) 。关于低等动物,拉脱特(M.E.Lartet) (144) 根据对同一类群的第三纪哺乳动物和近代哺乳动物的颅骨比较,作出如下值得注意的结论,即在较近代的类型中,一般脑要较大些,脑旋圈要较复杂些。另一方面,我曾指出,家兔的脑体积同野兔或山兔的脑体积相比较,前者是相当地缩小了; (145) 这大概可以归因于它们被严密地禁闭了许多代,因而很少运用它们的智力、本能、感觉以及随意运动(voluntary movements)。
脑和头骨重量的逐渐增加一定会影响作为支柱的脊骨的发达,特别是当变得直立的时候尤其如此。当带来这种姿势变化之后,脑的内压又要影响头骨的形状;因为许多事实阐明了头骨会多么容易地受到这样影响。人种学者相信,婴儿所睡的摇篮种类就会使头骨改变。肌肉的经常痉挛以及严重烧伤的疤痕,都会使面骨永久改变。青年人的头由于疾病向一边偏歪或向后歪,一只眼睛就要改变位置,而且头骨形状显然由于脑压朝着新方向发生作用而有所改变。 (146) 我曾阐明,关于长耳兔,甚至像一支耳朵向前垂下这样一种微小的原因,也会把几乎每一个头骨都朝着那一边向前拉;因而相对一侧的头骨就不严格对称了。最后,如果任何动物在一般体积上,大幅度地增加或缩减而心理能力不发生任何变化,或者,如果心理能力大幅度地增加或缩减而身体体积不发生任何重大变化,那么其头骨形状几乎肯定要发生改变。我是根据对家兔的观察作出这一推论的,有些种类的家兔变得比野兔大得很多,还有一些种类的家兔保持了同野兔差不多的大小,但是无论在哪一种情况下,它们的脑同身体体积相比,都大幅度地缩小了。当我最初发现所有这等家兔的头骨都变长了、即长头(dolichocephalic)的时候,使我大吃一惊;例如一只野兔的头骨和一只家兔的头骨,其宽度差不多相等,但前者的长度为3.15英寸,而后者的长度却为4.3英寸。 (147) 不同人类种族之间最显著的区别之一,就是有些种族的头骨是长形的,有些是圆形的;家兔事例所提供的解释,在这里也适用;因为韦尔克尔(Welcker)发现,矮个子“常倾向于短头(brachycephaly),而高个子则倾向于长头”。 (148) 高个子的人可以同身体越来越大的家兔相比拟,所有这等家兔都是“长头”的。
根据这几个事实,我们在一定程度上可以理解人类如何获得了大的而多少圆形的头骨,而人类同低等动物相比,这正是最显著不同的性状。
人类和低等动物之间另一个最显著的差异为人类的皮肤无毛。鲸和海豚(鲸目,Cetacea),儒艮(海牛目,Sirenia)以及河马都是无毛的,这对它们滑游于水中可能是有利的;而且这不会散失体内热量而对它们有害;凡栖息在寒带的物种,都有一厚层脂肪保护身体,其效用同海豹和水獭的毛皮一样。象和犀牛几乎是无毛的;以往曾在极其寒冷地区生活过的某些绝灭种却被有绵状毛或茸毛,因此这两个属的现存种失去它们的毛被似乎是由于暴露在炎热之中的缘故。因为印度的象生活于高寒地带者比生活于低地者被有较多的毛 (149) ,所以上述好像越发可能了。那么,我们是否可以这样推论,人类之所以失去他们的毛是由于原本居住在某一热带地方吗?现今男人主要在胸部和面部保存有毛,无论男人和女人还在四肢同躯干连接处保存有毛,这就支持了人类在直立以前就失去了毛的这样一种推论;因为现在毛保存得最多的部位,正是那时保护得最好而不受太阳热辐射危害的部位。然而,头顶却提供了一个奇特的例外,因为无论在任何时候它一定都是最暴露的部分之一,而它却密被头发。可是,人类属于灵长类,而灵长类的其他目(order)的成员虽然栖息于各式各样的热带地方,却周身有毛,一般朝上的表面最厚 (150) ,这一事实同人类通过日光作用而变得无毛的假设恰恰相反。贝尔特(Belt)先生 (151) 相信,在热带地方,无毛对人类是一种利益,这样可以避免大群的扁虱(螨,acari)和其他寄生虫,这些寄生虫常常侵扰他,而且不时引起溃烂。但是,这种弊害是否会大到足可以通过自然选择而导致他身体无毛,尚可怀疑,因为在栖息于热带的许多四足动物中,据我所知,没有一种获得了解除这种痛苦的手段。在我看来,最可能的观点是,男人、更确切地说是女人最初失去他们的毛,如我们将要在论“性选择”中所看到的,是由于装饰的目的;按照这一信念,人类同所有其他灵长类动物在毛发方面表现有如此重大差异,就不足为奇了,因为,通过性选择获得的性状在关系密切的类型中,其差异往往达到异常的程度。
按照普通的印象,以为尾的缺损乃是人类的显著特点;但是,同人类关系最近的那些猿类也没有这一器官,因此它的消失并非专与人类有联系。在同一个属内,尾的长度常常有巨大差别。例如猕猴属的某些物种的尾比它们的整个身体还要长,由24块椎骨形成;而在另外一些物种中,它仅是一个刚刚看得见的德根,只包含3~4块椎骨。有些种类的狒狒,它们的尾包含25块椎骨,而山魈(mandrill)的尾只有10块很小的、发育不全的尾椎,或者,按照居维叶(Cuvier)的说法,有时只有5块尾椎。 (152) 尾无论长的或短的,几乎永远在末端逐渐变细;我假定这是由于末端肌肉通过不使用而萎缩的结果,一齐萎缩的还有它的动脉和神经,因而导致了末端椎骨的萎缩。但是,关于它的长度常常发生的巨大差异,现在还无法提出解释。然而,这里我们所特别关注的却是尾的外部完全消失。布罗卡教授 (153) 最近阐述了,所有四足动物的尾均由两个部分组成,一般彼此截然分开;基部所包含的椎骨就像正常椎骨那样地具有多少完善的骨沟和骨凸(apophyses);而端部则不具骨沟,几乎是平的,简直不似真正的椎骨。虽然看不见人类和类人猿外部有尾,实际上却是存在的,而且其基部和端部以完全一样的形式构成。形成尾骨的端部椎骨完全是残迹的,其体积和数量大大缩减。基部椎骨同样也很少,牢固地结合在一起,发育受到抑制;但它们因此比其他动物的尾的相应椎骨宽阔得多而且扁平得多,它们构成了布罗卡所谓的副荐椎(accessory sacral vertebrae)。对于支持某些内在部分和在其他方面,它们具有机能上的重要性;而且它们的变异同人类以及类人猿的直立姿势或半直立姿势直接相关联。由于布罗卡以前持有不同的观点,而现在他已放弃,所以这一结论更可信赖。因此,人类以及高等猿类的基部尾椎的变异是直接地或者间接地通过自然选择而完成的。
但是,关于尾端部残迹的并且容易变异的椎骨,即形成尾骨者,我们将说些什么呢?有一种见解曾经常常受到嘲笑,无疑今后还会受到嘲笑,即认为尾的外部的消失同摩擦多少有些关系,而这一见解最初看来好像并不那样荒谬可笑。安德森(Anderson) (154) 博士说,褐猴(Macacus brunneus)的极短的尾是由11块椎骨形成的,嵌在肉里的基部椎骨也包括在内。尾端是腱质的,并不含椎骨;继此之后为5块残迹的椎骨,它们如此之小,其长度合在一起也不过一“赖因”(1ine) (155) 半,而且永久弯向一边成钩状。尾的自由部分的长度仅为一英寸稍强,只包含4块更小的椎骨。这个短尾可以直竖,但其全长的约四分之一向内折叠于左方;包括钩状部分在内的这一末端用于“填充老茧皮上方分开部分的间隙”;这种动物坐于其上,这样便使它成为粗糙的并且起老茧。于是安德森博士总结其观察所得如下:“在我看来,对这等事实只能有一种解释;由于这种尾是短的,当猴坐下来的时候,便可随心所欲地放置它,当猴取这种坐势时经常把它置于其下;因此尾不能伸出坐骨隆(ischial tuberosities)的末端之外,最初好像按照这种动物的意愿,将其尾弯成圆形置于老茧皮间的空隙,以避免在地面和老茧皮之间受到挤压,当弯曲变为永久性的时候,把它坐在下面自能适合。”在这种情况下,尾的表皮变得粗糙和起老茧,就不足为奇了,穆里博士 (156) 在伦敦动物园里曾仔细观察过这个物种以及另外三个尾巴稍长的密切近似的类型,他说,当这种动物坐下来时,它们的尾“必定要伸到臀部的某一边;无论它是长的或短的,尾根因而都容易受到摩擦或擦伤”。关于肢体损伤有时会产生遗传效果, (157) 现在我们已有证据;因此,在短尾猴中,尾的突出部分既在机能上无用,且由于不断地受到摩擦或擦伤,经历许多世代之后变为残迹的和弯曲的,看来并非是很不可能的事。我们看到褐猴尾的突出部分就是这种状态,而叟猴(M.ecaudatus)以及几种高等猿的尾的突出部分则是绝对发育不全的。最后,就我们所能判断的来说,人类和类人猿的尾是由于其末端在悠久的岁月里受到摩擦的损伤而消失了;嵌在肉内的基部缩小了而且变异了,以致可以适于直立的或半直立的姿势。
现在我已尽力阐明了,人类某些最独特的性状多半是直接地、或者更加普通的是间接地通过自然选择而获得的。我们应该记住,构造或体质的变异,如果不能使一个有机体适应于它的生活习性、它所消费的食物,或者被动地适应于环境条件,就不能这样获得之。但决定什么变异对每种生物是有用的,我们切不可过于自信:我们应该记住,对于许多部分的用途,或者对于血液或组织中的何种变化可以使一种有机体适合于新的气候或新的食物种类,我们所知道的是何等之少。我们也一定不要忘记,相关作用的原理,如小圣·伊莱尔在人类场合中所阐明的那样,把构造的许多奇特离差都束缚在一起了。与相关作用无关,某一部分的一种变化通过其他部分的增强使用或减弱使用,常常会导致具有一种完全意想不到的性质的其他变化。对于下述事实加以思考将是有好处的,如一种昆虫的毒物可以招致一些植物奇妙地生长树瘿,饲喂某些鱼类或注射蟾蜍的毒物可以使鹦鹉的羽衣颜色发生显著变化; (158) 于是我们可以知道,组织系统的体液如果为了某种特殊目的而发生改变,就会引起其他变化。我们应该特别记住,为了某种有用的目的在过去时期内获得的而且不断使用的变异,大概会牢稳地固定下来,而且会长久地被遗传下去。
这样,就可使自然选择的直接的和间接的结果扩展到巨大而无限定的范围;读了内格利(N geli)的有关植物的论文以及各位作者的有关动物的议论,特别是读了布罗卡教授最近写的那些文章之后,现在我承认我在《物种起源》最初几版中,也许归功于自然选择或最适者生存的作用未免过分了。我对《物种起源》第五版已作了一些改动,以便把我的论述局限在构造的适应性变化方面;但是,甚至最近几年所得到的事实也使我确信,在我们看来现今似乎无用的很多种构造今后将被证明是有用的,因而将会处在自然选择的范围之内。就我们现在所能判断的来说,有些构造的存在既是无益的也是无害的,对此我以前没有给予充分的考虑,我相信在我的著作中这是迄今所发觉的最大失察之一。作为某种借口,或者可以允许我这样说:我曾着眼于两个明确的目的,其一,在于阐明物种不是被分别创造的,其二,在于阐明自然选择是变化的主要动因,虽然它大部分借助于习性的遗传效果,并且小部分借助于环境条件的直接作用。然而,过去我未能消除我以往信念的影响,当时这几乎是一种普遍的信念,即各个物种都是有目的地被创造的;这就会导致我不言而喻地去设想,构造每一细微之点,残迹构造除外,都有某种特别的、虽然未被认识的用途。一个人如果在头脑里有这种设想,他自然会把自然选择无论过去或现在所起的作用过分夸大。有些承认进化论但否定自然选择的人们,当批评我的书时似乎忘记了我曾着眼的上述两个目的;因此,如果我在给予自然选择以巨大力量方面犯了错误——这是我完全不能承认的,或者我夸大了它的力量——在其本身来说这是可能的,那么我希望,至少我在帮助推翻物种被分别创造的教条方面作出了有益的贡献。
geli)的有关植物的论文以及各位作者的有关动物的议论,特别是读了布罗卡教授最近写的那些文章之后,现在我承认我在《物种起源》最初几版中,也许归功于自然选择或最适者生存的作用未免过分了。我对《物种起源》第五版已作了一些改动,以便把我的论述局限在构造的适应性变化方面;但是,甚至最近几年所得到的事实也使我确信,在我们看来现今似乎无用的很多种构造今后将被证明是有用的,因而将会处在自然选择的范围之内。就我们现在所能判断的来说,有些构造的存在既是无益的也是无害的,对此我以前没有给予充分的考虑,我相信在我的著作中这是迄今所发觉的最大失察之一。作为某种借口,或者可以允许我这样说:我曾着眼于两个明确的目的,其一,在于阐明物种不是被分别创造的,其二,在于阐明自然选择是变化的主要动因,虽然它大部分借助于习性的遗传效果,并且小部分借助于环境条件的直接作用。然而,过去我未能消除我以往信念的影响,当时这几乎是一种普遍的信念,即各个物种都是有目的地被创造的;这就会导致我不言而喻地去设想,构造每一细微之点,残迹构造除外,都有某种特别的、虽然未被认识的用途。一个人如果在头脑里有这种设想,他自然会把自然选择无论过去或现在所起的作用过分夸大。有些承认进化论但否定自然选择的人们,当批评我的书时似乎忘记了我曾着眼的上述两个目的;因此,如果我在给予自然选择以巨大力量方面犯了错误——这是我完全不能承认的,或者我夸大了它的力量——在其本身来说这是可能的,那么我希望,至少我在帮助推翻物种被分别创造的教条方面作出了有益的贡献。
就我所能知道的来说,所有生物,包括人类在内,可能均有无论过去或现在都是毫无用处的。因而不具任何生理重要性的构造特点。我们还不知道各个物种的诸个体之间的无数微小差异何以产生,因为返祖只不过把这个问题向后推移了少数几步,但每一个特点一定都曾经有过它的生效的原因。不管这等原因是什么,如果它们在一个长久时期内比较一致地和有力地发生作用(没有理由可以反对这一点),其结果大概不是仅仅的微小个体差异,而是十分显著而稳定的变异,虽然它们不具生理重要性。变化了的构造如果完全是无益的,就不能通过自然选择而保持一致,虽然变化了的有害构造将因此而被淘汰。然而,性状的一致性自然是起于激发原因的假定一致性,同样也是起于众多个体的自由杂交。在连续的时期内,同一个有机体可能以这种方式获得连续的变异,只要激发原因保持不变而且自由杂交如故,则这等变异将以差不多一致的状态被传递下去。关于激发原因我们所能说的,就像谈到所谓自发变异(spontaneous variation)时那样,只是,它们同变异着的有机体体质的关系要比同其外界条件性质的关系密切得多。
结 论
我们在这一章里已看到,人类在今天,就像每一种其他动物那样,容易发生多种多样的个体差异,即微小的变异,人类的早期祖先无疑也是如此;这等变异在以往同现在一样,都是由同样的一般原因所引起的,并且受同样的一般而复杂的法则所支配。由于所有动物的增殖都有超出其生活资料的倾向,所以人类的祖先一定也是如此;这就要不可避免地导致生存斗争和自然选择。后一过程大大受助于身体诸部分增强使用的遗传效果,这两种过程彼此相作用,永无止息。还有,如我们以后将要看到的,人类似乎是通过性选择获得了各种不重要的性状。此外还有无法解释的变化,只好把它们留给那些假定的未知力量的一致作用,这种作用在我们家养生物中偶然会引起强烈显著而突发的构造离差。
根据未开化人以及大多数四手类的习性来判断,原始人而且甚至人类的似猿祖先大概都是过社会生活的。关于严格社会性的动物,自然选择不时通过保存有利于群体的变异而对个体发生作用。一个群体如果包含大量禀赋良好的个体,就会增加其数量,而且就会战胜其他天赋较差者;即使个别成员并不优于同群的其他成员,也是如此。例如,群居昆虫所获得的许多奇异构造,如工蜂的花粉采集器或蜇针,兵蚁的巨大颚部,对于个体来说都是用处不大或者毫无用处的。关于高等社会性的动物,我还不知道有任何构造专为群体的利益而发生变异,虽然有些构造变异对于群体具有第二位的用途。例如,反刍动物的角、狒狒的大型犬齿,由雄者获得似乎是作为进行性竞争的武器,但也用于保卫兽群。至于某些心理能力,如我们在第五章将要看到的,情况就完全不同了;因为这等能力的获得主要是甚至专门是为了群体的利益,而个体不过因此同时间接地得到了利益而已。
上述这等观点常常遭到反对,谓人类乃是世界上最不能自助和自卫的一种动物,在其早期和不甚发达的状态下,他还要更加不能自助。例如,阿盖尔公爵(Duke of Argyll) (159) 坚决主张,“人类的体制同兽类的构造之分歧,是在身体的较大的不能自助性和软弱性那个方面的。这就是说,在其他一切分歧中,这是最不能把它归因于单纯的自然选择的”。他提到,身体无毛和无保护的状态,缺少用于自卫的大型牙齿或爪,人类的力气小而且速度慢,以及用嗅觉去发现食物或避免危险的能力薄弱。在这些缺点中似乎还可以加上一个更为严重的缺点,即人类不能迅速登攀以逃避敌对者。体毛的消失对热带居民来说大概不是什么重大损害。因为我们知道,不穿衣服的火地人在恶劣气候下也能生存。当我们以人类的不能自卫状态同猿类相比较时,我们必须记住,猿类所具的大型犬齿,只是在其充分发育时专为雄者所有,而且主要用于与其他雄者争取雌者的斗争;雌者虽不具此,也照样生存。
关于体格大小或体力强弱,我们还不知道人类究竟是从黑猩猩那样的某一小型物种传下来的呢,还是从强有力的大猩猩那样的物种传下来的;所以我们不能说,人类较其祖先变得更大更强些,还是变得更小更弱些。然而我们应该记住,正是体格大的、力量强的而且凶猛的、像大猩猩那样可以保卫自己不受一切敌对者危害的一种动物,也许未曾变为社会性的:恰恰是这一点最有效地阻止了高级心理属性——如对其伙伴的同情和热爱——的获得。这对于人类发生于某种比较软弱的动物,乃是一种巨大的利益。
人类的力气小、速度慢,本身不具天然武器等等,可由下列几点得到平衡而有余,即,第一,通过他的智力,他为自己制造了武器、器具等,即使依然处于野蛮状态下,也能如此。第二,他的社会性导致了他和同伴们相互帮助。世界上没有一处地方像南非那样地充满了危险的野兽,没有一处地方像北极地区那样地呈现了可怕的物质艰难,然而,一个最弱小的种族——布什门族(Bushmen) (160) 屹立于南非;矮小的爱斯基摩人(Esquimaux)则屹立于北极地区。毫无疑问,人类的祖先在智力方面,大概也在社会性方面,均劣于现存的最低等未开化人;但完全可以想象得到,如果他们在智力方面进步了,同时逐渐失去了他们的野兽般的能力,如爬树等,他们也会生存下来,甚至繁盛起来。如果这些祖先当时居住在温暖的大陆或大岛如澳大利亚、新几内亚(New Guinea)、婆罗洲(Borneo) (161) ——这些地方正是猩猩的现在故乡,即使他们远比任何现存的未开化人更加不能自助和自卫,也不致遭遇任何特别的危险。在上述那样广阔的区域里,由部落与部落之间的竞争而引起的自然选择,再加上习性的遗传效果,在适宜的条件下足可以把人类提高到现今他在生物等级中所占据的那样高上位置。
第三章 人类同低于人类的动物的心理能力比较
最高等猿类同最低等未开化人在心理能力上的巨大差异——某些共同的本能——各种情绪——好奇心——模仿性——注意力——记忆力——想象力——理性——向前改进——动物使用工具和武器——抽象作用,自我意识——语言——审美感——神的信仰,心灵作用,迷信
我们在以上两章中看到,人类在其身体构造上带有来自某一低等类型的明显痕迹;但也许可以这样说:由于人类在其心理能力(mental power)上同所有其他动物的差别是如此之大,因而这一结论一定还存在某种错误。毫无疑问,这一方面的差别是巨大的,即使我们把一个最低等未开化人——他没有表示四以上数目的任何字眼,并且对普通事物或感情也几乎不会使用任何抽象的名词 (162) ——的心理同一只最高等猿的心理加以比较,也是如此。纵然一种高等猿类改进或开化到像一只狗超出其祖先类型狼或豺(jackal)那样的程度,二者之间的差别无疑还是巨大的。火地人可以列为最低等的野蛮人,在英国皇家军舰“贝格尔”(Beagle)号上有三个火地土人,他们曾在英国住过几年,并且能说一点英语,这三个人在气质和大多数心理官能(mental faculties)上同我们如此密切相似,以致经常使我感到惊奇不已。如果除了人类以外没有一种生物具有任何心理能力,或者,如果人类的心理能力性质完全不同于低于人类的动物的,那么我们永远不能使自己相信人类的高等智能乃是逐渐发展而来的。但可以阐明,二者基本上没有这种差别。我们还必须承认,一种最低等鱼类如七鳃鳗(1amprey)或文昌鱼(1ancelet)同一种高等猿类在心理能力上的间隔要比猿类同人类在这方面的间隔广阔得多,而这一间隔是被无数级进(gradations)填补起来的。
就道德倾向(moral disposition)来说,像老航海家拜仑(Byron)所描述的那个野蛮人,因其子倾落一篮海参,就把他撞死在岩石上,以之比霍伍德(Howard)*或克拉克森(Clarkson) (163) ,其间的差别诚然不小。就智力来说,一个几乎不会使用任何抽象名词的野蛮人和牛顿(Newton) (164) 或莎士比亚(Shakspeare) (165) 之间的差别,亦复如此。最高等种族的最高等人士和最低等未开化人之间的这种差别,彼此是由最细小的等级连接起来的。因此,它们由这一端变化和发展到另一端,是可能的。
这一章的目的在于阐明,在心理官能上人类和高等哺乳动物之间并没有基本差别。这个题目的每一部分都可以扩充为一篇单独的论文,但在这里只能简短地加以讨论。因为关于心理能力还没有一种普遍被接受的分类方法,所以我将按照最适于我的目的的顺序来安排我的论述;并且选用那些给我印象最深的事实,我希望它们对读者会产生一些影响。
关于等级很低的动物,我将在讨论“性选择”时补充一些事实,以阐明它们的心理能力之高远远超出我们的意料之外。同一物种中诸个体的心理官能变异性,对我们来说是一个重要之点,所以要在这里举出少数例证。但关于这个问题,我不准备详加讨论,因为我根据多次调查得知,所有那些长期对许多种类动物甚至鸟类注意观察过的人们都一致认为,个体之间的每一种心理特性,都有重大差别。要问心理能力在最低等有机体中最初是以怎样方式发展起来的,就如同问生命本身是怎样起源的一样,目前还是没有希望得到解答。如果这些是人确能解决的问题,那也有待于遥远的未来了。
由于人类具有和低于人类的动物同样的感觉,所以人类的基本直觉(intuitions)一定也是同样的。人类和低等动物还有某些少数共同的本性,如自保,性爱,母亲对新生儿女的爱,新生儿女吸乳的欲望,等等。不过人类所具有的本能也许比低于人类的动物所具有的本能要稍微少一些。东印度群岛的猩猩以及非洲的黑猩猩,均筑平台作为宿所,由于这两个物种遵循这同样的习性,或许可以这样辩说:这是出于本能,但我们无法肯定,这不是由于这两种动物有相似的需要而且有相似的推理能力的结果。像我们所设想的那样,这等猿类不吃许多种热带的有毒果实,而人就没有这种知识。但是,我们的家养动物当被带到异地时,在春季第一次把它们放出去之后,常常会吃毒草,不过以后它们就避免吃了;我们还无法肯定,猿类不会从它们自己的经验中或者从它们双亲的经验中去选吃什么样的果实。然而,像我们即将看到的那样,猿类肯定有怕蛇的本能,并且可能还有怕其他危险动物的本能。同低等动物的本能相对照,高等动物的本能显著地比较少而简单。居维叶主张本能和智力彼此成反比,有些人以为高等动物的智能是从它们的本能发展而来的。但普歇(Pouchet)在一篇有趣的论文 (166) 中阐明,这种反比实际上是不存在的。具有最奇异本能的那些昆虫肯定是最有智力的。在脊椎动物的系列中,智力最差的成员如鱼类和两栖类,都没有复杂的本能;在哺乳动物中,以其本能著称的动物如河狸(beaver),则有高度的智力,每一个读过莫尔根(Morgan)先生的优秀著作 (167) 的人都会承认这一点。
虽然按照赫伯特·斯宾塞先生 (168) 的说法,智力的最初端绪是通过反射作用(reflex actions)发展而来的,虽然比较简单的本能逐渐变为反射作用而且二者几乎无法区别,如幼小动物的吮乳,但更加复杂的本能的起源,似乎还是与智力无关。然而我绝不是否认本能活动会失去其固定的和不学自会的特性并且可以由自由意志(free will)所助成的其他特性所代替。另一方面,有些智力活动进行了几代之后,还会转变成本能而被遗传下去,如海洋岛上的鸟类学会避人就是这样。于是这等活动可以被说成是特性的退化,因为这种活动进行不再通过理性或经验了。但是,大多数比较复杂的本能似乎是以一种完全不同的方式被获得的,即由于比较简单的本能活动的变异受到了自然选择。这等变异似乎是由作用于脑组织的同样未知原因而发生的,引起身体其他部分发生微小变异或个体差异的就是这等原因;由于我们的无知,这等变异常常被说成是自然发生的。我以为,关于比较复杂的本能的起源,我们还作不出任何其他结论,如果我们考虑一下不育的工蚁和工蜂的不可思议的本能,而它们却不留后代以承继它们的经验和改变了的习性的效果,就可想而知了。
虽然我们从上述昆虫和河狸认识到高度的智力同复杂的本能确是共存的,虽然最初随意学得的动作不久可以通过习性以一种反射作用迅速而准确地进行之,但自由智力(free intelligence)和本能之间还有一定程度的抵触——后者含有脑的某种遗传变异。关于脑的功能,我们所知者甚少,但我们能够觉察到,当智力变得高度发达时,一定有最自由沟通的而且极其错综复杂的渠道把脑的各部分连接在一起;因此,每一个独立部分恐怕要较差地适于以一种确切的和遗传的——即本能的——方式去回答特殊的感觉或联想(associations)。甚至在智力的低级程度和形成固定的、但不是遗传的习性的强烈倾向之间似乎也存在着某种关系。因为一位有洞察力的医生告诉我说,稍微有点低能的人每一行动都倾向于按照常规、即习性,如果给他这种鼓励,就会使他非常高兴。
我以为这种离题之论还是值得一提的,因为,当我们把高等动物、特别是人类的以记忆力、预见力、推理力和想象力为基础的心理能力活动和低于人类的动物以本能来执行的完全相似的活动加以比较时,我们也许容易地对前者的心理能力估价得过低;在低等动物的场合中,执行这等活动的能力是通过心理器官在各个连续世代中的变异性和自然选择逐步被获得的,而与动物所表现的任何有意识的智力无关。正如华莱士先生 (169) 所辩说的,人类所完成的很多智力工作无疑是由于模仿,而不是由于理性;但人类的活动和低于人类的动物的许多这等活动之间的重大差别,即在于此。这就是说,人类不会通过他的模仿力在最初一试中就能制造比如说一只石斧或一条独木舟,人类必须通过实践去学习工作;另一方面,一只河狸筑造它的堤堰或水道 (170) ,一只鸟筑造它的巢,在最初一试中其完善程度就可以像它年老而有经验时一样,或者差不多一样,而一只蜘蛛在最初一试中所织成的网同其年老而有经验时所织成的就完全一样地完善了。 (171)
现在回到本题上来:低等动物像人那样也会感到快乐和悲伤,幸福和苦难。幼小动物如小狗、小猫、小羊等在一起玩耍时和我们的小孩一样,没有比它们在这时所表现出的幸福感更加明显的了。甚至昆虫,如卓越的观察家于贝尔(Huber) (172) 所描述的,也像许多种类小狗那样地在一起玩耍,他曾看到一些蚁相互追逐,彼此假相咬啮。
低于人类的动物可以被和我们同样的感情所激动,这个事实已经如此充分地得到证明,以致没有必要再详加说明而引起读者厌烦。恐怖对它们发生作用的方式就同对我们一样,会引起肌肉颤抖,心脏跳动,括约肌(sphincters)松弛以及毛发竖立。猜疑是畏惧的产物,它是大多数野生动物的显著特性。坦南特(E.Tennent)爵士关于用做诱捕其他象的雌象行为写过一篇报道,我想凡是读过这篇报道的人不可能不承认这些雌象是有意识地在玩弄欺诈,而且深知它们在干什么。勇敢和怯懦在同一物种的诸个体中是极端容易变异的属性,这在我们养的狗中有明显的表现。有些狗和马的脾气坏,容易生气,还有一些狗和马的脾气好,这等属性肯定是遗传的。谁都知道动物多么容易狂怒,而且表达得多么明显。关于各种动物经过长久期间后还会狡猾地进行报复,已经发表过许多逸事,看来这大概是真实的。伦格尔和布雷姆 (173) 说,他们所养驯的美洲猴和非洲猴确会施行报复。动物学家安德鲁·史密斯(Andrew Smith)爵士的严格认真是众所周知的,他给我讲过一个他亲眼所见的故事:在好望角有一位军官经常虐待一只狒狒,某星期日当这只狒狒看到他列队前进时,便把水倒入一个小坑里,急忙和些稠泥,当这位军官走近时,它熟练地把稠泥向他猛砸过去,于是逗得许多旁观者发笑。很久以后,每当这只狒狒看到这位受害者的时候,还表现出胜利的欢欣。
狗对主人的爱是众所周知的,一位往昔的作者 (174) 富有风趣地说道:“在这个世界上,狗是爱你甚于爱它自己的唯一动物。”据知,狗在临死的极度痛苦中还抚爱它的主人,大家都听说过,正在被解剖中的一只狗还去舐解剖者的手;除非这次解剖确可增加我们的知识,要不,除非解剖者心如顽石,否则他必将悔恨终生。
休厄尔(Whewell) (175) 有理由地问道:“一切民族的妇女的母爱同一切雌性动物的母爱如此经常地联系在一起,以致读过这等动人事例的人,能够怀疑在这两种场合中的行为原则不是一样的吗?”我们看到在微小细节上所表现出来的母爱,例如,伦格尔观察到一只美洲猴(卷尾猴,Cebus)小心地把打扰母猴的幼儿的蝇子赶跑;迪沃塞尔(Duvaucel)看到一只长臂猿(Hylobates)在一条小河边为它的幼儿洗脸。雌猴失去它们的幼儿时,其悲痛是如此剧烈,以致布雷姆在北非圈养的某些种类必定因此而死去。早孤的幼猴总是由其他雄猴和雌猴收来抚养,并且受到小心保护。有一只雌狒狒,它的心肠如此宽宏,不仅收养其他物种的幼猴,而且还偷取小狗和小猫,随时把它们带在身边。然而,在把它的食物分给受抚养的幼猴方面,它就不那样仁慈了,这使布雷姆感到惊异,因为它养的猴总是把每一件东西十分公平地分给它亲生的幼猴。一只受抚养的小猫把这只富有深情的狒狒抓破了,这只狒狒的智力肯定是敏锐的,因为它对被抓破感到非常惊讶,随即检查小猫的脚,立刻把它的爪咬去。 (176) 伦敦动物园的一位管理员告诉我说,在那里有一只老狒狒(C.chacma),它抚养一只猕猴(Rhesus monkey),但是,当把一只幼山魈(drill)和西非山魈放进槛笼时,它似乎觉察到这两只猴虽属于异种,却是它的较近亲属,于是它立刻弃去那只猕猴,而收养了幼山魈和西非山魈。我看到这只小猕猴对于受到这样遗弃,表示非常不满,它像一个顽皮儿童那样地给小山魈和小西非山魈找麻烦并攻击它们,每当它能安全地这样干的时候它就这样干,这种行径激起了老狒狒的很大愤慨。按照布雷姆的说法,猴类当其主人受到任何侵犯时都会保护他,就像主人所养的狗当他受到别的狗侵犯时对他进行保护一样。但我们在这里触及了同情和忠诚的问题,以后我还要讨论这一点。布雷姆养的有些猴以各种巧妙的方法戏弄它们所厌恶的一只老狗和其他动物,由此而感到非常高兴。
大多数比较复杂的情绪是人类和高等动物所共有的。众所周知,如果一只狗的主人对任何其他动物表示过分地亲热,这只狗会多么妒忌;关于猴,我曾观察到同样的事实。这阐明动物不仅会施爱于他,而且有受爱的欲望。动物显然有好胜心,它们喜欢受到称赞。一只狗为它的主人携带一只篮子,就会表现出高度的自满或骄傲。我以为当狗过于频繁地乞求食物时,无疑它会感到羞耻,这同恐惧有别,而接近于谦逊。大狗对小狗的吠叫表示蔑视,这或者可以被称为宽宏大量。若干观察家说过,猴类肯定厌恶别人拿它取笑,而且有时它们幻想这是受到攻击。我在伦敦动物园看到一只狒狒,每当它的饲养员拿出一封信或一本书向它高声朗读时,它总是暴怒,它是如此怒气冲冲,以致有一次我亲眼看到它咬自己的腿,直到流血。狗有一种名副其实的幽默感,这和单纯的游戏有所不同。如果把一小截树枝或其他类似物品丢给一只狗,它常常把这件东西带到不远的地方,然后蹲在它的近前等候着,直到主人完全走近来拿这东西的时候,于是它抢先衔住这东西,耀武扬威地猛奔而去,它重复地玩弄这同样的花招,并且显然享受这种开玩笑的乐趣。
现在我们谈谈更近于理智的情绪和官能,这是高等心理能力发展的基础,故很重要。动物显然喜兴奋,而恶无聊,所以看到狗有这种情形,伦格尔说猴也有这种情形。所有动物都有惊异感(wonder),有许多动物还显示好奇心(curiosity)。它们不时因后一属性而受害,因为当猎人玩弄滑稽动作时,它们就会这样受到诱惑;我亲眼看到,鹿是这样,谨慎的岩羚羊(Chamois)是这样,某些种类的野鸭也是这样,布雷姆有过如下的奇妙报道:他养的猴对蛇表示了本能的畏惧;但它们的好奇心如此之重,以致不能打消一看的念头,不时把蓄蛇箱的盖子掀开,以饱享恐怖之乐,这很像人类的风尚。我对他的报道感到非常惊奇,所以我把一条人造的、盘卷的蛇标本扔进伦敦动物园的猴房,由此而引起的激动是我平生所看到的最奇妙景象之一。有三种长尾猴(Cercopithecus)最为惊恐,它们在笼内冲来冲去,并且发出为其他猴所明白的带有危险信号的尖锐叫声。少数幼猴和仅有一只老阿努比斯狒狒(Anubis baboon)对这条蛇不予注意。于是我把这个人造的标本放到一间较大的猴房地上。这一回,所有的猴都集到一起围成一个大圈,目不转睛地注视着那条蛇标本,面貌极其滑稽可笑。它们变得极度神经紧张,有一只它们经常玩的木球,部分埋在麦草内,不料它从那里滚出来,弄得它们立刻惊散。当把一条死鱼、一只鼠、 (177) 一只活龟以及其他新奇物件放进它们的笼内时,这些猴的表现就大不同了;虽然它们最初被吓一跳,可是很快就走近这些东西,触摸它们而加以检查。这时我把一条活蛇放入一个纸袋内,袋口微闭,然后把它放在一间较大的猴房里。有一只猴随即走近,小心地把袋口打开一点,向内窥视,立刻猛冲而去。于是我亲眼见到布雷姆所描述的那种情况:诸猴相继而来,把头抬得高高地,而且扭向一侧,忍不住向这个直立的袋内偷看一下那个安静地卧在袋底的可怕之物。好像猴类对动物学的亲缘关系也有某种概念,因为布雷姆所养的猴对无害的蜥蜴和蛙表示了一种奇异的、虽然是错误的本能恐惧。据知,猩猩最初一看到龟也非常惊恐。 (178)
人类的模仿性(imitation)很强,如我亲自观察的,未开化人的模仿性尤其强。在脑部患有某种病症的状况下,这一倾向被扩大到异常的程度:有些半身不遂的患者以及其他脑部初期炎性软化的患者,不自觉地模仿别人说的每一个字,无论这是本国语言还是外国语言,而且模仿他们所看到的每一种姿势或动作。 (179) 德索尔(Desor) (180) 曾说,没有一种动物自愿地模仿人类的动作,直至上升到猴类的等级,都是如此;众所周知,它们是可笑的模仿者。然而,动物不时彼此模仿对方的动作:例如,有两种由狗养育起来的狼,它们学狗叫,就像豺不时所做的那样, (181) 不过这是否可以被称为自愿的模仿还是另一个问题。鸟类模仿其双亲的鸣声,有时还模仿其他鸟类的鸣声;鹦鹉以善于模仿它经常听到的任何声音而著称。马尔(Dureau de la Malle) (182) 做过如下报道:有一只由猫养育起来的狗,它学着模仿猫的一种出名的动作,用舌舐脚爪,然后洗其双耳和脸,著名的博物学者奥杜因(Audouin)亲自见过这种情形。我收到过几篇这方面的确实报道,其中之一表明,有一只猫把一只狗同几只小猫一齐带大了,但它并没有吃过猫的奶,可是这只狗就这样获得了上述习性,而且此后在它一生的13年中一直这样做。马尔养的一只狗同样地也从小猫那里学会用前爪扑打着球,使它滚来滚去。一位通信者向我保证说,他家有一只猫惯于用前爪伸人牛奶罐内醮奶偷吃,因为罐口太狭,容不进它的头。这只猫生养的一只小猫很快就学会了这个诡计,此后只要有机会它就这样干。
许多动物的双亲依靠其幼儿的模仿性、特别是依靠其本能的或遗传的倾向,或者可以称为对它们进行教育。当老猫把一只活鼠带给它的小猫时,我们就可以看到这种情形了。马尔就他对鹰的观察写过一篇奇妙的报道(见上述引用的文章):鹰用以下的方法去教小鹰学会敏捷以及对距离的判断,即首先把死鼠和死麻雀从空中丢下来,但小鹰一般捉不到它们,然后把活鸟带给小鹰,再纵放它们飞去。
对人类智慧的进步来说,几乎没有任何智能比注意力(attention)更重要的了。动物明确地显示了这种能力,如猫守候在鼠穴旁,准备向鼠扑去。野生动物有时如此集中注意力,以致这时人可以容易地接近它们。巴特利特(Bartlett)先生给过我一个奇妙的例证以说明这种能力在猴类中多么容易变异。有一位驯猴做戏的人,惯常从“动物学协会”购买普通的种类,每只付价五镑;但是如果让他把三四只猴养上少数几天,再从其中选出一只,他就愿付出双倍的价钱。当问他怎么能够那样快地判断出被选定的猴是否会成为一个好的表演者,他答道,这完全决定于它们的注意力。当他向一只猴说话和解说任何事物的时候,如果它的注意力容易分散,僻如说把注意力转向墙上的一只苍蝇或其他细小物件,那么这种情形就没有希望了。如果他试着用责罚来使注意力不集中的猴做戏,它就要发怒。另一方面,有些猴小心地注意着他,这些猴肯定可以被训练好。
动物对人和地点都有极好的记忆力(memories),对此已不必多加赘述。安德鲁·史密斯爵士告诉我说,在好望角有一只狒狒,在他离去九个月之后还认识他,并表示了喜悦之情。我养过一只狗,它对所有生人都嫌恶而且凶悍十足,在离开五年零两天之后,我特意试过它的记忆力。我走近它的窝,按照我的老样子呼喊它,它虽没有表示喜悦,但立即跟着我出去散步,并且服从我的指挥,好像我和它刚分开半小时一样。休眠达五年之久的一连串联想,就这样立即在它的头脑中被唤醒了。正如于贝尔 (183) 所明确阐述的,甚至蚁类和同群的伙伴分开四个月之后,还能彼此认识。动物肯定能以某种方法去判断再发事件的间隔时间。
想象力(imagination)是人类所拥有的最高特权之一。凭借这种官能,而不是依赖意志,他就能把先前的意象(images)和观念(ideas)联合在一起,并由此得到灿烂而新奇的结果。正如吉恩·保罗·里歇特(Jean Paul Richter) (184) 所说的,一位诗人“如果必须思考他要塑造的人物究应说‘是’,还应说‘否’——见他的鬼去吧;这个人物只能是一具愚蠢的僵尸”。做梦这件事可以使我们有一个关于想象力的最好概念,吉恩·保罗还说过,“梦乃是一种无意识的诗之艺术”。我们想象力的产物的价值当然决定于我们的印象的数量、准确性和清晰度,决定于我们在取舍无意识的印象组合时所作的判断和所表现的爱好,并且在一定程度上还决定于我们有意识地组合它们的能力。因为狗、猫、马、可能一切高等动物乃至鸟类 (185) 都有清晰的梦,它们在睡眠中的动作和发出的声音阐明了这一点,所以我们必须承认它们具有某种想象力。一定有某种特殊的原因致使狗在夜间,特别是在月夜中以一种异常的、忧郁的声调吠叫。并非所有狗都夜吠,乌泽说,它们不是对着月亮吠叫,而是对着接近地平线的某一固定地点吠叫。 (186) 乌泽以为它们的想象力被周围物体的模糊轮廓扰乱了,于是在它们面前呈现出幻想的意象,倘真如此,则它们的感觉差不多可以被称为迷信了。
我设想,在人类的所有心理官能中,理性(reason)可以被承认处于顶峰。现在只有少数人对动物具有某种推理能力还有疑问。随时可见,动物会踌躇、审慎和下决心。一位博物学者对任何特殊动物的习性研究得越多,他就把习性归因于理性者越多,而归因于无意识的本能者越少 (187) ,这是一个值得注意的事实。在以下几章中将会看到,某些等级极低的动物显然也显示一定程度的理性。理性的能力和本能的能力无疑常常是难以区别的。例如,海斯(Hayes)博士在他的《开放的北极洋》(The Open Polar Sea)一书中屡次提到,当他的狗把雪橇拉到薄冰上的时候,它们就不继续采取密集队形,而是彼此散开,以便它们的重量可以比较平均地分布。这常常是旅行者们所得到的最先警报:冰已经变薄而且有危险了。那么,狗的这种行为是来自各个个体的经验呢,或是来自比较年长而且比较聪明的那些狗的示范呢,还是来自一种遗传的习性,即本能呢?这种本能可能发生于很久以前当地居民用狗来拉雪橇的时候,或者,爱斯基摩狗的祖先——北极狼已经获得了这样一种本能,迫使它们不要在薄冰上密集地去攻击它们所要捕食的动物。
我们只能根据完成行为时所处的环境条件去判断这些行为是由于本能、或是由于理性、还是由于观念的联合。默比斯(M bius) (188) 教授举过这样一个奇妙的事例:有一只狗鱼(pike)在水族箱内被玻璃板隔开,玻璃板的另一侧养着一些鱼,它常常如此猛烈地撞向玻璃板,试图捉对面的鱼,以致不时被撞晕过去。这条狗鱼这样继续干了三个月,但最后学会慎重,停止乱撞了。这时把玻璃板移去,它不再攻击原来的那些鱼,却吞食此后放进去的鱼,在它的薄弱心理中,一种猛烈冲撞的观念与捕食以前邻居的试图如此强有力地联合在一起了。如果一个从来没有见过大厚玻璃窗的未开化人,甚至只在窗上撞过一次,长久以后他还会把冲撞和窗框联想到一起。但和狗鱼大不相同,他大概要对障碍的性质进行思考,并且会在相似情况下加以注意。关于猴类,如我们即将看到的那样,只要有一次由于一种行为而得到痛苦的,或者仅仅是不适意的印象,有时这就足可以阻止这种动物再去重复它。如果我们把猴和狗鱼的这种差别完全归因于猴比狗鱼的联合观念的能力强得多而且持久得多,虽然狗鱼所受到的损害常常严重得多,那么在人类的场合中,我们能够主张一种相似的差别是意味着他具有一种基本不同的心理吗?乌泽 (189) 说,当在得克萨斯穿过一处广阔而干燥的平原时,他的两条狗非常之渴,它们冲下凹地去找水,不下三四十次。这些凹地并非溪谷,那里没有一棵树,而且植被也没有任何其他差别,况且那里是绝对干燥的,所以不会有一点湿土的气味。狗有这样的行为,好像它们知道低凹的地势可以为其提供找到水的最好机会,乌泽还经常亲眼见到其他动物也有这种同样的行为。
bius) (188) 教授举过这样一个奇妙的事例:有一只狗鱼(pike)在水族箱内被玻璃板隔开,玻璃板的另一侧养着一些鱼,它常常如此猛烈地撞向玻璃板,试图捉对面的鱼,以致不时被撞晕过去。这条狗鱼这样继续干了三个月,但最后学会慎重,停止乱撞了。这时把玻璃板移去,它不再攻击原来的那些鱼,却吞食此后放进去的鱼,在它的薄弱心理中,一种猛烈冲撞的观念与捕食以前邻居的试图如此强有力地联合在一起了。如果一个从来没有见过大厚玻璃窗的未开化人,甚至只在窗上撞过一次,长久以后他还会把冲撞和窗框联想到一起。但和狗鱼大不相同,他大概要对障碍的性质进行思考,并且会在相似情况下加以注意。关于猴类,如我们即将看到的那样,只要有一次由于一种行为而得到痛苦的,或者仅仅是不适意的印象,有时这就足可以阻止这种动物再去重复它。如果我们把猴和狗鱼的这种差别完全归因于猴比狗鱼的联合观念的能力强得多而且持久得多,虽然狗鱼所受到的损害常常严重得多,那么在人类的场合中,我们能够主张一种相似的差别是意味着他具有一种基本不同的心理吗?乌泽 (189) 说,当在得克萨斯穿过一处广阔而干燥的平原时,他的两条狗非常之渴,它们冲下凹地去找水,不下三四十次。这些凹地并非溪谷,那里没有一棵树,而且植被也没有任何其他差别,况且那里是绝对干燥的,所以不会有一点湿土的气味。狗有这样的行为,好像它们知道低凹的地势可以为其提供找到水的最好机会,乌泽还经常亲眼见到其他动物也有这种同样的行为。
我曾在伦敦动物园里看见过,我敢说别人也曾在那里看见过,当把一个小物件扔到一头象钩不到的地面上,它就会用鼻子向着小物件那边的地面上吹气,所以从四面八方反射回来的气流,可以把那个物件吹至它能钩到的范围之内。再者,一位著名的人种学家韦斯特罗普(Westropp)先生告诉我说,他在维也纳看到一只熊用它的前脚去拍打其笼子栏杆前面的一汪水,造成水流,以便把一片漂浮的面包引至它能钩到的范围之内,简直不能把象和熊的这等行为归因于本能,即遗传的习性,因为这对处于自然状态下的动物一点也没有用处。那么,当一个未开化人也有这等行为时,它们同一种高等动物的这等行为有什么区别吗?
未开化人和狗往往在平地的低处发现过水,这种发现水时的情况总是彼此一致的,这种一致的情况在它们的心理中便联系起来了。文明人也许对这个问题可以提出某种一般的命题,但根据我们所知道的未开化人的一切情况来说,他们是否也能这样做,确系一个极大的疑问,狗肯定不能这样做。但是,未开化人乃至狗还能按照同样的方式去找水,虽然他们屡屡感到失望;未开化人的、或者狗的这种行为似乎同等都是理性的,无论是否有任何一般的命题有意识地置于心理之中。 (190) 象和熊造成气流和水流的那种情况,也是如此。未开化人肯定不会理解,也不会关心依据什么法则才能完成所期望的运动;但他的行为受到一种粗略的推理过程的引导,的确就像一位哲学家在他的一大串演绎中所做的那样。毫无疑问,未开化人和高等动物之间的差别在于:未开化人注意极其细小的境况和条件,并且以其极少的经验来观察这二者之间的任何关联,而这一点则具有至高无上的重要性。我对我的一个小孩的行为曾逐日做过记录,当他长到11个月左右的时候,在他还不能说一个单字之前,他就能迅速地把所有种类的事物和声音在他的精神中联系在一起,其迅速的程度超过我所知道的最聪明的狗,这一情况屡屡给我留下了深刻印象。但是,高等动物同狗鱼那样的低等动物之间在联想力、推理力和观察力方面的差别也完全如此。
美洲猴的下述行为很好地阐明了通过很短的经验之后就能激起理性的活动,而美洲猴在灵长类中处于低级的地位。一位最谨慎的观察家伦格尔说道,当他在巴拉圭第一次把一些鸡蛋给他所养的猴时,它们把鸡蛋打碎了,因而大部分蛋黄和蛋白都流失掉了;其后它们就把鸡蛋的一端轻轻地向一种坚硬的东西击撞,并且用手指剥去一点碎壳。只要它们被任何锐利的工具割伤一次之后,它们以后就不再触动它,或者非常小心地去拿它。伦格尔常常把糖块用纸包好后再给它们;有时他在纸包中放一只活黄蜂,当它们急着打开纸包时就被蜇到了;只要经过这样一次之后,它们总是首先把纸包放在耳朵旁边,侦查一下其中是否有任何动静。 (191)
下述是关于狗的一些事例。科尔库杭(Colquhoun)先生曾用枪射伤两只野鸭的翅膀,它们落在一条小河另一边较远的地方;他的“拾物猎狗”(retriever)试图一次把两只同时叼回来,但没有成功;于是它审慎地咬死一只,把另一只带过河后,又回去带那只死的,但在此之前它从来没有损伤过野鸭一根羽毛。哈钦森(Hutchinson)上校 (192) 叙述,他曾用枪同时射到两只鹧鸪,一只被射死,一只受伤;受伤的那只逃走,但被拾物猎狗捉到,当它回来的时候又跑到那只死鹧鸪处;“它停了下来,显然非常为难,试了一两次之后,发现它无法把死鸟带走而不让伤鸟逃去,考虑片刻之后,它就狠狠地给伤鸟一口,把它咬死,然后把两只一齐带走”。这是它“故意伤害任何猎物的唯一事例”。在这只拾物猎狗先去捉伤鸟然后又回过头来带死鸟的例子中,以及在那两只野鸭的例子中,我们看到了理性,虽然这并不是十分完全的。我之所以列举上述两个例子,因为它们是以两位彼此无关的目睹者所提出的证据为基础的,并且因为在这两个事例中“拾物猎狗”经过深思熟虑之后竟然打破了它们所遗传下来的一种习性(不咬死拾取的猎物),同时还因为它们显示了其推理力多么强有力地克服了固定的习性。
我愿引用杰出的洪堡 (193) 的一段议论作为这个问题的结束。他说:“南美的赶骡人说道,‘我不给你一头走得最平稳的骡子,我给你一头理性最好的骡子’”;接着洪堡又说:“根据长期经验所表达出来这种通俗言辞,反驳了动物乃是有生命的机器系统那种说法,恐怕它比思辨哲学的所有论点都好。”尽管如此,有些作者甚至现在还否认高等动物具有一点理性的痕迹,而且他们力图凭借看来仅仅是一些冗词滥调 (194) 把上述一切事实巧辩过去。
我想,现在我已经阐明了人类和高等动物、特别是和灵长类动物有一些少数共同的本能。它们都有同样的感官、直觉以及感觉——相似的热情、情感以及情绪,甚至更复杂的,如嫉妒、猜疑、争胜、感激以及宽宏大量;它们都会玩弄欺诈和实行报复;它们有时对受到嘲笑都敏感,甚至还有一种幽默感,它们都有惊奇感和好奇心;它们都具有同样的模仿、注意、深思熟虑、选择、记忆、想象、观念联合、理性等各种官能,虽然其程度有所不同。同一物种的诸个体在智力上有许多等级,由绝对低能一直到高度优秀。它们也有患精神错乱的,但这种情形远比在人类场合中为少。 (195) 尽管如此,许多作者还坚决主张,人类和一切低于人类的动物在心理官能方面是被一道不可逾越的障壁分开的。以前我曾搜集过大量有关上述的警句,但几乎都是没有什么价值的,因为其内容彼此差异极大,而且数量过多,证明这种试图如果不是不可能的,也是困难的。有人断言,只有人类能够向前改进;只有他能利用工具和火,驯养其他动物,或者拥有财产;任何动物都没有抽象力、即形成一般概念的能力,都没有自我意识和自知之明;任何动物都不能使用语言;只有人类有审美感,不容易解释的怪想,感激之情,神秘感等;人类信仰上帝,并且有良心。我愿就其中比较重要而有趣的几点贸然提一点意见如下。
大主教萨姆纳(Sumner) (196) 以前主张,只有人类才能向前改进。人类比其他任何动物的改进都无比之大而且无比之快,对此已无争辩的余地了;这主要是由于他有说话的能力,并且能把他获得的知识传下去。关于动物,我们首先看一看个体,每一个对设置陷阱有点经验的人都知道,小动物比老动物容易被捉到;而且敌对者接近它们也比较容易。关于老动物,甚至不可能在同一地点和用同一种类的陷阱捉到许多,或者用同一种类的毒药把它们全都毒死;它们大概不可能都一齐吃过毒药,或者一齐被陷阱捕捉过。它们一定由于看到同伴的被捕捉或被毒害而学会警惕。所有观察家们一致证明,在北美,毛皮动物长期受到追捕,因此它们所显示的机智、小心以及狡猾几乎到了难以置信的程度;但是在那里设置陷阱已经进行了如此之久,以致遗传性业已起了作用并非是不可能的。我曾收到几份报道,指出当在任何地区初设电报时,许多鸟由于飞撞电线而致死,但经过几年之后,它们似乎看到同伴因此而死的情况,便学会了避免这种危险。 (197)
如果我们考虑到连续的世代或考虑到种族,毫无疑问,鸟类以及其他动物对人类或其他敌对者的警惕是逐渐地获得和失去的 (198) ;肯定地,这种警惕大部分是一种遗传的习性或本能,但一部分乃是个体经验的结果。一位优秀的观察家勒鲁瓦(Leroy) (199) 述说,在有大量猎狐的地方,小狐在最初离开它们的穴时,其警惕性不可否认地远远超过那些猎狐不多的地方的老狐。
我们家养的狗是从狼和豺传下来的 (200) ,虽然它们在狡诈方面可能无所得,在警惕和猜疑方面也许有所失,但它们在某些道德品质方面,如仁爱、忠诚、温良,而且大概在一般智力方面,却向前发展了。在整个欧洲,在北美的一部分地方,在新西兰,最近在中国,普通鼠已经战胜和打倒了另外几个物种。斯温赫(Swinhoe)先生 (201) 描述过中国内地和台湾地区的这种情况,他把普通鼠之所以能够战胜一种大型家鼠(Mus coninga)归因于前者有较大的狡诈性;这种属性的获得大概可以归因于它们避免人类扑灭的一切能力惯常地受到了锻炼,并且可以归因于差不多一切狡诈较差或智力薄弱的鼠类不断地被它们所消灭。然而,普通家鼠的取胜可能是由于它们在同人类接触之前就已经具有了优于同时存在的其他物种的狡诈性了。不以任何直接证据为依据,而主张没有任何动物经历悠久岁月的过程在智力或其他心理官能方面曾经有所前进,这无异用未经证明的假定对物种进化问题进行狡辩。根据拉脱特的叙述,我们已经知道,属于若干“目”的现存哺乳动物的脑大于其第三纪的古代原型的脑。
经常这样说,动物不会用任何工具;但是,在自然状况下的黑猩猩却会用一块石头把一种好像胡桃似的当地果实打碎。 (202) 伦格尔 (203) 容易地教会一只美洲猴用石头把一个硬棕榈坚果击破,此后它就会主动这样把其他种类的坚果甚至箱子击破。它还会这样去掉味道不适口的软果皮。另一只猴被教会用一根木棍把一个大箱子盖撬开,此后它就会把木棍作为杠杆去移动沉重的物体;我曾亲自见到一只小猩猩把一个木棍插入裂缝,用手握住另一端把箱子撬开,它把木棍当做杠杆用的方式是恰当的。众所周知,印度的驯象会折取树枝,用以赶跑蝇子;曾经观察到在自然状态下的一头象也会这样干。 (204) 我曾看到一只小猩猩自以为要受鞭打,便用毡子或麦草来掩护自己。在这几个事例中,石头和木棍是被当做工具用的,但它们同样地还把这些东西当武器用。布雷姆 (205) 说,根据著名旅行家席佩尔(Schimper)的权威叙述,在埃塞俄比亚(Ethiopia),当一种狮尾狒狒(C.gelada)成群结队从山上下来掠夺田野的时候,它们时常同另一种埃塞俄比亚鼯猴(C.hamadryas)相遇,这时便要发生战斗。狮尾狒狒把大石头滚下来,埃塞俄比亚鼯猴设法躲开,然后双方大声喧嚣,彼此凶猛地冲击。布雷姆陪伴科堡-哥达公爵(Duke of Coburg-Gotha)曾在埃塞俄比亚的门沙(Mensa)隘道用火器助攻一群鼯猴;作为报复,这群鼯猴从山上滚下来这么多的石头大如人头,以致攻击者不得不迅速退却;而且隘道实际上为之堵头,堵塞了一段时间,致使货车不得通过。值得注意的是,这些鼯猴是协同动作的。华莱士先生曾三次见到一些携带着幼子的雌猩猩“以非常狂怒的容貌折断榴莲树(Durian tree)的枝条和大刺果,掷如雨下,有效地防止了我们走到树的近旁”。 (206) 我曾屡次见到黑猩猩把手边的任何东西掷向来犯的人;还有,前文提到的好望角的那只狒狒准备好稠泥作为攻击之用。
伦敦动物园里有一只猴,它的牙齿软弱,经常用一块石头把坚果敲开,管理员们向我确言,它用毕那块石头,便把它藏在麦草下面,并且不许其他任何猴动它。于是我们在这里看到了所有权的观念;不过每一只狗对于一块骨头,以及大部分或全部鸟类对于它们的巢,全有这种观念。
阿盖尔(Argyll)公爵 (207) 说,制造适合于一种特殊目的的工具,绝对只有人类才能做到;他认为这在人类和兽类之间形成了难以计量的分歧。无疑这是一个很重要的区别;但是在我看来,卢伯克爵士 (208) 的意见还是相当正确的,他认为当原始人类最初为了达到任何目的而使用燧石时,可能偶然地把它们打成了碎片,这时他大概会选那些锐利的碎片来用。从这一步到有目的地弄破燧石,大概只有一小步;再经过不大的一步,就可以粗糙地使它们成形了。然而,在新石器时代人类开始琢磨石器以前,却经历了非常悠久的岁月,据此判断,上述后面那种进步大概也需要很长的时间。卢伯克爵士又说,当破裂燧石时,火花会发出;当琢磨石器时,热会生出:这样,“两种通常取火的方法便发生了”。在许多火山区,熔岩不时流过森林,那里的人对火的性质大概会有所了解。类人猿大概在本能的引导下,为自己建造临时的平台;但是,许多本能主要受理性的支配。所以像建造平台那样比较简单的本能大概会容易地变成一种自愿的和有意识的行为。据知猩猩在夜间用露兜树叶遮盖自己,布雷姆说,他养的狒狒经常把草席盖在头上以防太阳晒。在这几种习性中,我们大概看到了向着某些比较简单的技艺——如发生于人类早期祖先时代的那种粗糙的建筑和衣服——的最初步骤。
抽象作用,一般概念作用,自我意识,心理的个性
无论谁,即使学问远远超过我的人,要想决定动物呈现任何这等高级心理能力的痕迹到怎样程度,也是很困难的。这种困难起因于不可能判断在动物心理中所闪过的念头是什么,还有,作者们对上述名词所赋予的意义大不相同,这就招致了进一步的困难。如果根据最近发表的各种材料来判断,那么最强调的似乎还是在于假定动物完全没有抽象的能力,即没有形成一般概念的能力。但是,当一只狗在一段距离内看到另一只狗时,显然它抽象地察觉到那是一只狗;因为,当它走近时,另一只狗如果是一个朋友,它的全部举止就会突然改变。最近一位作者说,在所有这等事例中,断言人类和动物的心理行为在本质上具有不同的性质,乃是一种纯粹的臆测。如果任何一方把由感官所察觉到的归入一种心理概念,那么双方均可如此。 (209) 我以热切的声调向我的 (terrier) (210) 说(我如此试过多次),“嘿,嘿,它在哪里呢?”它立刻把这作为一种信号,表明有些东西有待猎取,一般先是急向周围注视,然后冲入最近的灌木丛,嗅寻是否有任何猎物,当什么都找不到的时候,它就向邻近的树上窥视,看看那里是否有松鼠。那么,这等行为不是明显地阐明了在它的心理中有一种关于某些动物有待发现和猎取的一般观念或概念吗?
(terrier) (210) 说(我如此试过多次),“嘿,嘿,它在哪里呢?”它立刻把这作为一种信号,表明有些东西有待猎取,一般先是急向周围注视,然后冲入最近的灌木丛,嗅寻是否有任何猎物,当什么都找不到的时候,它就向邻近的树上窥视,看看那里是否有松鼠。那么,这等行为不是明显地阐明了在它的心理中有一种关于某些动物有待发现和猎取的一般观念或概念吗?
如果自我意识这个名词的含义是,它会考虑他是从哪里来的、或者它将往哪里去、或者什么是生和死等等那样的问题,那么根据这个名词的这种含义,可以坦白地承认动物不具有自我意识。但是,一只老狗如果具有最好的记忆力和某种想象力,如它做梦所阐明的;我们总能肯定它决不会考虑它过去在追猎中的乐趣或痛苦呢?这大概就是自我意识的一种形态。另一方面,如比希纳 (211) 所说的,智力低下的澳洲未开化人的辛苦劳动的妇人只能说很少的抽象言辞,计数不能到四以上,她们所行使的自我意识或对其本身存在的考虑是何等之少。高等动物具有记忆力、注意力、联想力甚至某种想象力和推理力,已得到普遍承认。如果在不同动物中大不相同的这等能力能够改进,那么,通过比较简单智能的发展和结合,进化到比较复杂的智能、如抽象和自我意识等等的高级形态,似乎并没有很大的不可能性。有人认为不可能说出在上升阶梯的哪一点动物变得能够进行抽象等等,并以此极力反对这里所主张的观点;但是,有谁能说出我们的幼儿在什么年龄可具有这种能力吗?至少我们知道,幼儿的这等能力的发展是以不可觉察的程度进行的。
动物保有它们的心理个性是没有问题的。当我的声音唤起上述那只狗在心理中的一连串联想时,它一定保有它的心理个性,虽然它的每一个脑原子在这五年期间大概不止一次地发生了变化。也许有人要利用这条狗把最近发生的辩论向前推进以打垮所有进化论者,说道:“在所有心理状态和所有物质的变化中……我坚持认为,关于原子可以像遗产那样地把它们的印记留给落入它们所空出的位置中的其他原子的那种学说是与意识的表达相矛盾的,所以这种学说是虚假的;而这种学说正是进化论所必需的,因此进化的臆说也是虚假的。” (212)
语 言
这种能力已被公平地认作是人类和低等动物之间的主要区别之一。但是,正如一位高度有才能的评论家惠特利大主教(Archbishop Whately)所说的,人类“并不是唯一的动物能够利用语言来表达其心理上所闪过的东西,并且多少能够理解他人如此表达出来的东西。” (213) 巴拉圭的一种卷尾猴当激动时至少可发出六种不同的声音,这些声音对另外一些猴可以激起相似的情绪。 (214) 伦格尔以及其他人士宣称,猴类的面貌动作和姿势能为我们所理解,而且它们也能部分地理解我们的。还有一个更加值得注意的事实:狗自从被家养之后,至少学会叫出 (215) 4~5个不同的音调。狗的吠叫虽是一种新技艺,但是狗的野生祖先无疑会以各种不同的叫声来表达它们的情感。关于家狗,有热切的叫,如在追猎中那样;有愤怒的叫以及不平的叫;失望的狺狺叫或嗥叫,如在被关起来时那样;夜间的空叫;欢乐的叫,如在陪伴主人开始出去散步时那样;还有一种请求或哀求的很独特的叫,如在要求开门或开窗时那样。赫祖(Houzeau)特别注意过这个问题,他说,家鸡至少可发出12种有区别的声音。 (216)
惯常使用有音节的语言,为人类所专能;但是,他也用无音节的喊叫,辅以姿势和面部肌肉的动作,来表达他的意思,这同低于人类的动物无异。 (217) 当表达那些同我们高等智力很少关联的简单而活跃的情感时,尤其如此。我们的痛苦、恐怖、惊奇、愤怒的叫声,再加上恰如其分的动作,以及母亲对爱子的低沉连续的哼哼声,比任何言辞都富有表达力。人类和低于人类的动物的区别并不在于是否理解有音节的声音,因为,每一个人都知道,狗是理解许多字句的。在这方面,狗和10~12个月的婴儿处于相同的发育阶段,那时婴儿理解许多单字和短句,但连一个单字还不会说。我们区别于低等动物的特性并不仅仅在于有音节的语言,因为鹦鹉和其他鸟类也有这种能力。也不仅仅在于把一定声音和一定观念连接在一起的智能;因为有些鹦鹉当被教会说话之后,也可以准确地把字和物以及人和事连接在一起。 (218) 低等动物和人类之间的区别完全在于人类把极其多种多样的声音和观念连接在一起的能力几乎是无限大的,而这显然决定于其心理能力的高度发展。
宏伟的语言科学奠基人之一霍恩·图克(Horne Tooke)论述,语言是一种技艺,就同酿酒和烤面包一样;不过书写也许是一个更好的直喻。这肯定不是一种真正的本能,因为每一种语言都必须学而知之。然而,语言和一切普通技艺都大不相同,因为人类有一种说话的本能倾向,如我们幼儿的咿呀学语就是这样;同时却没有一个幼儿有酿酒、烤面包或书写的本能倾向。再者,现在没有一位语言学家还假定任何语言是被审慎地创造出来的;它是经过许多阶梯缓慢地、无意识地发展起来的。 (219) 鸟类发出的声音在若干方面同语言极为近似,因为同一物种的所有成员都发出同样本能的鸣叫来表达它们的情绪;而所有能够鸣叫的鸟类都是本能地发挥这种能力;不过真正的鸣唱,甚至呼唤的音调,都是从它们的双亲或其养母养父那里学来的。戴恩斯·巴林顿(Daines Barrington) (220) 已经证明,“鸟类的鸣声同人类的语言一样,都不是天生就会的”。鸟类最初鸣唱的尝试“可以同一个幼儿不完全的咿呀学语的努力相比拟”。幼小的雄鸟要继续练习,或如捕鸟人所说的,它们要“录音”达10~11个月之久。在未来的鸣唱中几乎没有最初试鸣的一点痕迹;但当它们稍稍长大的时候,我们还能觉察出它们所欲学者为何事,最后,它们便被称为“能够圆润地唱歌”了。学会不同物种鸣唱的雏鸟,如在蒂罗尔(Tyro1)训练的金丝雀,则把它们的新歌传教给其后代。栖息在不同地区的同一物种,它们的鸣唱有轻微的自然差异,如巴林顿所说的,这可以恰当地比做“各地方言”;虽然属于不同物种,但亲缘关系近似者的鸣唱或可以比做人类不同种族的语言。我之所以举出上述细节是为了阐明,求得一种技艺的本能倾向并非人类所专有。
关于有音节的语言起源,当我一方面读了亨斯利·韦奇伍德(Hensleigh Wedgwood)先生、法勒(F.Farrar)牧师以及施莱歇尔(Schleicher) (221) 教授的最有趣味的著作,另一方面又读了马克斯·米勒(Max Müller)教授的讲演集之后,我无法怀疑语言的起源应归因于:对各种自然声音、其他动物叫声以及人类自己的本能呼喊的模仿及其修正变异,并辅以手势和姿势。当我们讨论到性选择的时候将会看到,原始人类,更确切地说人类的早期祖先,大概最初用他们的声音来发出音乐般的音调,即歌唱,就像某些长臂猿今天所做的那样;根据广泛采用的类推方法,我们可以断定这种能力特别行使于两性求偶期间——它会表达各种情绪,如爱慕、嫉妒以及胜利时的喜悦——而且还会用于向情敌挑战。所以,用有音节的声音去模仿音乐般的呼喊,可能会引起表达各种复杂情绪的单字的发生。和我们亲缘关系最近的猴类,畸形小头的白痴 (222) ,以及人类的野蛮种族,都有一种强烈的倾向去模仿所听到的一切,这是值得注意的,因为同模仿问题有关。因为猴类理解人向它们说的话一定很多,而且在野生状况下会向其同伴发出作为危险信号的呼叫; (223) 还因为家鸡会发出地面危险和空中有鹰类危险的两种不同警告(这两种叫声以及第三种叫声皆能为狗所了解), (224) 那么某种异常聪明的类猿动物曾经模仿食肉兽的吼叫,并且以此来告诉其猿类同伴所料想的危险性质,难道是不可能的吗?这大概是语言形成的第一步。
由于声音的使用日益增多,发音器官通过使用效果的遗传原理将会强化和完善化;而且反过来这对说话的能力又会发生作用。但是,语言的连续使用和脑的发展之间的关系无疑更加重要得多。甚至在最不完善的语言被使用之前,人类某些早期祖先心理能力的发展一定也比任何现今生存的猿类强得多;不过我们可以确信,这种能力的连续使用及其进步,反过来又会对心理本身发生作用,促使其能够进行一系列的思想活动。一系列复杂思想,无论在说话时或不说话时,如果没有言辞的帮助是无法进行的,正如不使用数字或代数就无法进行长的计算一样。甚至一系列普通思想似乎也需要某种形式的语言,或者被它所大大推进,因为一个聋、哑、盲的少女劳拉·布里奇曼(Laura Bridgman)曾被看到在梦中还打手势。 (225) 尽管如此,没有任何形式的语言帮助,也可通过心理产生一连串活泼的和彼此联系的观念,因为从狗在梦中的动作可以作此推论。我们还知道,动物也能够进行一定程度的推理,这显然并不依靠语言的帮助。像我们现在这样发达的脑与说话能力之间的密切关系,从特别影响说话能力的那些脑病奇妙例子中得到了很好的阐明。例如,当记忆名词的能力失去之后,还能正确地使用其他单词,或者,还能记住某一类名词或全部名词,但忘记了这些名词的起首字母及其恰当的意义。 (226) 心理器官和发音器官的连续使用将导致它们在构造和功能上发生遗传的变化,这就像笔迹的情形那样,它部分地决定于手的形状,部分地决定于心理的倾向,而笔迹肯定是遗传的。 (227)
几位作者、特别是马克斯·米勒教授 (228) 最近极力主张,语言的使用意味着要有形成一般概念的能力;没有任何动物被假定具有这种能力,因此,这就形成了人类和动物之间的一个不可逾越的障碍。 (229) 关于动物,我已经尽力阐明了它们至少以一种原始萌芽的程度具有这种能力。就10~11个月的婴儿来说,我简直不能相信他们能够把某些声音和某些一般观念那样迅速地在头脑中连接在一起,除非这等观念已经在他们的头脑里形成了。同样的这种意见可以引申到智力较高的动物,如莱斯利·斯蒂芬先生 (230) 所观察的,“一只狗对猫和绵羊可以构成一般概念,而且可以像哲学家那样准确地知道与它们相称的字眼。理解的能力犹如说话的能力,很好地证明了运用语言进行表达的智力,虽然其程度较差”。
为什么现今用以说话的器官起始就已经为了这个目的达到了完善化的地步,而任何其他器官都不是这样,这并非难以理解,蚁类具有利用触角彼此交流信息的相当能力,胡伯尔已经阐明了这一点,他曾用整整一章来讨论蚁类的语言。我们可以用手指作为交流信息的有效手段,因为一个熟练此术的人能够把公共集会上说得很快的讲演词的每一个字用手势报告给聋人;但是这样被使用的双手一旦失去,必将造成严重的不便。所有高等哺乳动物都有发音器官,都是按照和我们同样的一般图式构成的,而且都是用做交流信息的手段,因此,如果交流信息的能力得到了改进,这等同样器官还会进一步发展,显然是可能的;相连的和十分适应的各部分、即舌和唇帮助了这一发展的完成。 (231) 高等猿类不会用发音器官来说话,无疑是决定于它们的智力还没有足够的进步。它们具有经过长期连续练习后才可用来说话的那些器官,但现在并没有这样用,这同具有适于鸣唱的器官但从来不鸣唱的鸟类事例是相似的。例如,夜莺和乌鸦都有构造相似的发音器官,前者能用它进行多种多样的鸣唱,而后者只能用它呱呱地叫。 (232) 如果问道,为什么猿类的智力没有发展到人类那样的程度,我们只能举出一般的原因作为回答;试想,我们对各种生物所经过的发展诸连续阶段几乎一无所知,却希望作出更加明确的任何回答,都是不合乎道理的。
不同语言的形成和不同物种的形成,以及二者的发展都是通过逐渐过程,其证据是异常相似的。 (233) 但是,对于许多词的形成比对于物种的形成,我们可以向前追踪得更远,因为我们能够察觉词实际上是怎样来自对各种声音的模仿的。我们发现,不同的语言由于起源的共同性而彼此一致,还由于相似的形成过程而彼此类似。当其他字母或发音有所变化时,某些字母或发音就要随之变化,其方式同生长的相关作用很相像。在这两种场合中都有诸部分的重叠、长期连续使用的效果等。无论在语言或在物种中都屡屡出现一些残留的遗迹,这就更加值得注意了。在“am”这个词中,m表示I的意思,因此在“Iam”这个词句中便保存了多余而无用的残留遗迹。还有,在词的拼法中也常常残留着作为古代发音形式遗迹的字母。语言有如生物,也可以逐类相分;既可以按照由来的系统进行自然分类,也可以按照其他特性进行人为分类。占有优势的语言和方言广为传播,并且导致其他语言的逐渐绝灭。一种语言有如一个物种,一旦绝灭,如莱伊尔爵士所说的,就永远不会再现。同一语言绝没有两个发源地。不同语言可以杂交或混合在一起。 (234) 我们知道每一种语言都有变异性,而且不断地产生新的词;但是,由于记忆力有一个限度,所以词就像整个语言那样,会逐渐绝灭。正如马克斯·米勒 (235) 所恰当指出的:“各种语言的词和语法形式都在不断地进行着生存斗争。较好的、较短的、较易的形式永占上风,它们的成功应归因于它们本身固有的优点。”某些词的生存除了有上述那些比较重要的原因之外,还可以加入对新奇和时髦的爱好;因为在人类的心理中对所有事物的微小变化都有一种强烈的爱好。在生存斗争中,某些受惠的词的生存或保存乃是由于自然选择。
许多野蛮民族的语言构造是完全规律而异常复杂的,这常常被提出以证明这些语言起源于神,或者证明这些语言的创始者具有高度的技艺和既往的文化。例如,冯·施勒格尔(F.von Schlegel)写道:“在那些看来似乎是智育程度极低的语言中,我们屡屡观察到在其语法构造上有很高程度的和精心制作的技艺。巴斯克语(Basque) (236) 和拉普语(Lapponian) (237) 以及许多美洲语言尤其如此。” (238) 但是,如果认为语言是被精心地和有条理地构成的,就把任何语言都说成是一种技艺,肯定是错误的。语言学者现已承认动词各种变化形式、词尾变化形式等等原本都是作为不同的单词存在的,后来才结合在一起了;这等单词表达了人和物之间的最明显的关系,因此,它们在最古时代为大多数种族的人所使用,就不足为奇了。下述的例证最好地阐明了我们在完善化这个问题上多么容易犯错误:一种海百合(crinoid)有时是由不下十五万个壳片构成的, (239) 所有壳片的排列都以放射线状而完全对称,但博物学者们并不认为这种动物比两侧对称的动物更为完善,后者身体的诸部分比较少,除了身体两侧的各部分彼此相像以外,其余部分都不相像。他公正地把器官的分化和专业化看做是对完善化的检验。关于语言,也是如此:最对称的、最复杂的语言不应被列在没有规律的、简略的以及混杂的语言之上,所谓混杂的语言就是从各种征服别人的种族、被征服的种族以及移入的种族那里借入了一些表达力强的词和语言构造的有益形式。
根据这些不完善的少数议论,我断言,许多野蛮人语言的极其复杂和极其规律的构造不足以证明,语言是起源于一种特殊的创造行为。 (240) 正如我们已经看到的,有音节语言的能力实质上也没有提供出任何不可排除的理由来反对人类是从某一低等类型发展而来的信念。
审 美 感
这种感觉曾被宣称为人类所专有。我这里谈到的只是关于由某些颜色、形状和声音所引起的愉快感,这或者可以恰当地被称为对美的感觉;然而对文明人来说,这等感觉是同复杂的观念和一系列的思想紧密地联合在一起的。如果我们看到一只雄鸟在雌鸟面前尽心竭力地炫耀它的漂亮羽衣或华丽颜色,同时没有这种装饰的其他鸟类却不进行这样的炫耀,那就不可能怀疑雌鸟对其雄性配偶的美是赞赏的。因为到处的妇女都用鸟类的羽毛来打扮自己,所以这等装饰品的美是毋庸置疑的。我们在以后几章中将会看到,蜂鸟(humming-birds)的巢和造亭鸟(bower-birds)的游戏通道都用鲜艳颜色的物件装饰得很优雅;这阐明它们见到这些东西后一定会感到某种愉快。然而,就我们所能判断的来说,大多数动物对于美的爱好仅限于吸引异性。许多雄鸟在求偶季节所鸣唱的甜蜜歌声,肯定会得到雌鸟的赞赏;关于这个事实的证据,以后再举。如果雌鸟不能够欣赏其雄性配偶的美丽颜色、装饰品和鸣声,那么雄鸟在雌鸟面前为了炫耀它们的美所作出的努力和所表示的热望,岂不是白白浪费掉了,这一点是不可能予以承认的。为什么某些鲜艳的颜色会激起快感,我以为所能解释的,不会比对于某些味道和气味何以会令人感到愉快的解释更多一点,但是,习性对于这个结果一定有些关系,因为有些东西最初使我们感官不舒适,但终于使它们舒适了,而且习性是遗传的。关于声音,为什么和声与某些音调令人感到悦耳,赫姆霍尔兹(Helmholtz)根据生理学原理在一定程度上对此提出了解释。但是,除此之外,在不规则的时间内经常翻来覆去的声音最叫人厌烦,凡是在夜间听过缆绳不规则地拍打船板的人都会承认这一点。同一原理似乎也适用于视觉,因为眼睛喜欢看到对称或规则地循环出现的图形。甚至最低等的未开化人也把这种图案用做装饰品;通过性选择,这等图案发展为某些雄性动物的装饰。对于这样来自视觉和听觉的愉快,不论我们能否提出什么理由,总归人类和许多低等动物都一样地喜欢同样的颜色、同样的优雅色调和形状以及同样的声音。
对于美的爱好,并非人类精神中的一种特殊本性,至少就妇女的美而论是如此;因为,在不同的人种中这种爱好大不相同,甚至在同种的不同民族中也不完全一样。根据最不开化人对丑陋的装饰品以及对同等丑陋的音乐的赞赏来判断,可以认定他们的审美能力还没有发展到某些动物,例如鸟类那样的高度。显然没有什么动物能够赞赏诸如夜晚的天空、美丽的山水那样的景色,或优美的音乐;但是,这等高尚爱好是通过教养才获得的,而且依靠复杂的联想,野蛮人或没有受过教育的人不会欣赏它们。
许多这等官能曾对人类向前的进步作出了不可估量的贡献,诸如想象、惊异,好奇的能力,没有界限的审美感,模仿的倾向,对刺激或新奇的喜爱,几乎不能不导致风俗和时尚发生不定的变化。我之所以提出这一点,是因为最近一位作者 (241) 奇怪地把不定性作为“未开化人和兽类之间的最显著的、最典型的差异之一”。但是,我们不仅能够部分地理解人类怎样由于各种相互冲突的影响而成为不定性的,我们还能部分地理解低等动物,如此后即将看到的那样,在其爱好、厌恶以及审美感方面也是不定的。还有理由来设想,它们也爱新奇,正是为了那是新奇的缘故。
神的信仰——宗教
还没有证据可以证明,人类本来就赋有对于一位万能上帝存在的崇高信仰。恰恰相反,有充分的证据可以证明,曾经有、现在依然有为数众多的种族没有一神或多神的任何观念,而且在他们的语言中从来没有表达这一观念的字。 (242) 当然,这个问题同是否存在有一位主宰宇宙的创造者和统治者那种更高的问题完全是两码事,而在最高级的知识界中有些人已经对后一问题作了肯定的答复:确是存在的。
如果我们把对灵魂世界或精灵作用的信仰包括在“宗教”这一名词之内,那就完全是另一回事了,因为文化较低的种族似乎普遍都有这种信仰。关于它是如何发生的,并不难说明。一旦想象、惊异、好奇那些重要官能以及某种推理能力部分地有所发展之后,人类自然会渴望理解在他周围发生的情况,而且还会对其本身的存在模糊地进行思考。伦南先生 (243) 曾经说过:“人一定要对生命现象为自己想象出某种解释,根据这种解释的普遍性来判断,人最初想到的最简单的臆说似乎曾经是,自然现象可以归因于在动物、植物和物品中,以及在自然界的力量中,都存在有主使运动的精灵,这种精灵同人自觉到自己有一种内在的精神力量而外发为种种活动一样。”正如泰勒(Tylor)先生所阐明的,梦境也许是发生精灵概念的起因,这也是可能的,因为未开化人不会很快地把主观印象和客观印象区别开。当一个未开化人做梦时,他相信出现在他面前的形影是从远方来的;并且监视他的;或者,“做梦人的灵魂在旅途中出了窍,把所见到的都记在心中而回到家里”。 (244) 但是,当想象、好奇、推理等等能力在人类精神中相当完善地发展之前,他的梦境不会引导他去相信精灵,这和狗在做梦后不会这样是相同的。
有一次我曾注意到一件小事情,也许它可以说明未开化人有一种倾向去想象给予自然物体或自然力量以生命的是精灵的或活的实体:我有一只狗,已达到成年,而且很聪明,在一个炎热而宁静的白天里它卧在一片草地上;在距它不远的地方,放着一把张开的阳伞,微风不时把它吹动,如果有人在阳伞的旁边,这条狗就完全不去理睬它。事实上,当阳伞旁边没有人时,无论什么时候只要阳伞稍微一动,这条狗就要凶猛地吠叫。我想,它一定以迅速而无意识的方式给自己推论出,没有任何明显原因的阳伞活动暗示了有某种奇怪的活力量存在,而且它认为陌生者没有权力在它的领域内停留。
对精灵作用的信仰将会容易地变为对一神或多神存在的信仰。因为未开化人自然会认为我们所感到的同样的情欲,同样的对复仇或简单形式的正义的喜爱以及同样的慈爱,均系精灵所赐。火地人在这方面似乎居于中间状态,因为,当“贝格尔”号舰上的军医射击一些幼鸭做标本时,火地人约克·明斯特(York Minster)以最严肃的态度宣称:“唉呀,拜诺(Bynoe)先生,要下大雨、下大雪、刮大风呀”;显然这是对糟蹋人类食物的一种报应的惩罚。他又说道,他的弟弟杀了一个“野人”,于是风暴肆虐很久,而且下了大雨和大雪。然而我们从来没有发现过火地人信仰我们所谓的上帝,或者实行任何宗教仪式;火地人吉米·布顿(Jemmy Button)以一种情有可原的骄傲态度坚定地主张,他的家乡没有魔鬼。他的这种主张更加值得注意,因为未开化人信仰恶的精灵远比信仰善的精灵更加普遍得多。
宗教信仰的感情是高度复杂的,其中包括爱、对崇高的和神秘的居上位者的完全服从,强烈的信赖感 (245) 、恐惧、崇敬、感激以及对未来的希望,也许还有其他要素。没有任何生物能够体验如此复杂的一种感情,除非他的智力和道德官能至少进步到中等高度的水平。尽管如此,我们还会看到狗对主人的深爱,结合着它的完全服从、某种恐惧心,也许还有其他情感已经遥遥地多少向着上述那种心理状态接近了。一只狗在离别后又回到主人那里的态度,我还可以接着指出,一只猴在离别后又回到它所喜爱的饲养员那里的态度,和对它们同群的态度大不相同。在离别后与同群再见时,欣喜若狂的劲儿似乎多少要小一些,而且在每一个动作中都显示了平等感。布劳巴哈(Braubach)教授甚至主张,狗把它的主人看成是一位神。 (246)
同样水平的心理官能最初引导人去信仰不可见的精灵作用,然后是信仰拜物教,多神教,最终是一神教;只要他的推理力保留在不发达的状态下,这种水平的心理官能一定会引导人产生各式各样奇怪的迷信和风俗。许多这等迷信和风俗真是骇人听闻——例如,把人作为牺牲献给嗜血的神;用服毒或探火的神裁法去审讯无辜的人;巫术等——对于这等迷信不时进行思考是有好处的,因为它们阐明了我们应该多么感激我们理性的进步、科学以及我们积累起来的知识所赐予的无限恩惠。正如卢伯克爵士 (247) 所正确观察的,“不必过多地说些什么就可明白,对于未知的灾祸所抱有的那种可怕的畏惧,就像一层厚云那样笼罩在未开化人的生活之上,而且更加重了他们的痛苦”。人类最高能力所产生的这等不幸的和间接的结果可以同低于人类的动物本能所附带发生的偶然错误相比拟。
第四章 人类同低于人类的动物的心理能力比较(续)
道德观念——基本命题——社会性动物的属性——社会性的起源——相反本能的斗争——人类是一种社会性动物——比较持久的本能战胜比较不持久的本能——未开化人唯独重视社会美德——自重美德是在较晚发展阶段获得的——同群公众对善恶行为评判的重要性——道德倾向的遗传——提要
有些作者 (248) 主张在人类和低于人类的动物之间的一切差异中,道德观念、即良心是最重要的;我完全同意这一判断。正如麦金托什(Mackintosh) (249) 所指出的,道德观念“理所应该地凌驾于其他任何人类行为的准则之上”;它的高深意义可以总结在简短而重要的“应尽义务”这个词中。它是人类所有属性中最高尚的一种属性,引导他毫不迟疑地冒着自己生命的危险去保护同伙的生命;或者,经过适当的深思熟虑之后,仅仅由于对权利和义务的深刻感觉,而被迫在某种伟大事业中牺牲自己的生命。康德(Immanuel Kant) (250) 喟然叹曰:“义务!不可思议之思想乎,其工作既不由献媚求宠,亦不由威胁恐吓,而仅仅由灵魂中所高举汝之无私法律,因此,汝如不能强取对汝永远遵从,亦将强取对汝永远敬畏;一切欲望无论如何秘密地进行反抗,在汝之前均哑然无声,汝果从何而发生乎?” (251)
许多才华横溢的作者 (252) 已对这个伟大问题进行了讨论,我触及这个问题的唯一可以原谅之处,仅在于不可能在这里对它略而不谈,而且还在于,就我所知道的来说,还没有人完全从博物学方面来探讨过这个问题。这一研究还有某种独立的趣味,可以作为一种尝试来看。对低于人类的动物的研究可以把人类最高心理官能之一说明至何种程度。
在我看来,下述命题是高度可能的——即,无论何种动物,只要赋有十分显著的社会本能 (253) (包括亲子之情),一旦其智力发展得像人类的那样完善,或者差不多那样完善,就必然会获得一种道德观念,即良心。这是因为,第一,社会本能可以导致一种动物以和其同伙营社会生活为乐,对其同伙有一定程度的同情心,并且为其同伙进行各种服务。这种服务可能具有一种明确的和显然是本能的性质;或者可能只是一种希望和思想准备,如大多数高等社会性动物以某些一般的方式去帮助它们的同伙那样。但是,这种感情和服务仅施于它们的同伙,决不会扩大到同一物种的所有个体。第二,一旦心理官能变得高度发达之后,所有过去的行为和动机的意象将不断地在各个个体的头脑中通过;如我们以后就要看到的,由任何不满足的本能而必然发生的不满足的感情、甚至痛苦,像常常被觉察到的那样,将会引起持续而永在的社会本能让位给较强的某种其他本能,但后者的性质并非是持续的,也不给后来留下很鲜明的印象。显然,许多本能的欲望,如饥饿,在性质上其持续是短暂的;而且一旦得到满足之后,就不会容易地或者鲜明地被回忆起来。第三,当语言能力被获得并且公共愿望能够被表达之后,各个成员为了公共利益应该如何行动的舆论,自然会成为指导行为的最高准则。但是,应该记住,不论我们认为舆论力量有多么大,我们对于同伙的称赞和非难还决定于同情心;如我们即将看到的,同情心形成了社会本能的主要部分,而且确是它的基石。第四,个体的习性在指导各个成员的行为方面,起了很重要的作用;因为,社会本能连同同情心,就像其他任何本能那样,大大地被习性所强化了,因而就要遵从公众的愿望和评判。现在必须对这几个从属的命题进行讨论,有些还要以相当篇幅进行之。
最好预先声明一下,我并非要主张,任何严格社会性动物的智能如果变得像人类的那样灵敏,那样高度发达,它就会获得和人类完全一样的道德观念。各种动物都有审美感,虽然它们所赞美的对象大不相同,同样地,各种动物大概都有是非感,虽然由此而导致遵从的行为界线大不相同。举一个极端的例子来说明,例如,人的养育条件如果同蜜蜂的完全一样,那么几乎无可怀疑的是,未婚妇女就会像工蜂那样把杀死她们的兄弟视为神圣的义务,同时母亲们也要努力杀死其能育的女儿,而且不会有任何同类想到去进行干涉。 (254) 尽管如此,我们为蜜蜂或任何其他社会性动物在我们那个假定的场合中将会获得某种是非感或良心。因为各个个体都有一种内觉(inward sense),这种内觉具有某些较强的或较持久的本能以及不甚强的或不甚持久的本能;所以对于遵从何种冲动(impulse),将经常进行斗争;而且,由于过去的印象当不断通过头脑时要进行比较,因而将会感到满足,不满足,或者甚至痛苦。在这种情况下,内在的告诫者将告诉这种动物遵从某一冲动会比遵从另一冲动为好。某种行动方向应该被遵从,另外的行动方向不应被遵从;某种行动方向是正确的,另外的行动方向是错误的;不过关于这些问题。以后还要谈及。
社 会 性
许多种类的动物都是社会性的,我们发现甚至不同物种也在一起生活。例如,某些美洲猴类,以及合群的秃鼻乌鸦(rooks)、寒鸦(jackdaws)和欧椋鸟,都是这样。人类对狗的强烈爱好,表现了同样的感情,狗也高兴地报答他们。大家一定都曾注意到,当马、狗、羊等离开它们的同伴时表现得多么悲惨,而以前曾在一起的两个种类至少在重聚时所显示互爱之情是何等强烈。一只狗同它的主人或其他任何家庭成员在室内可以安静地一连卧上几个小时,一点也不必去理会它;但是,让它自己待在那里,即使时间不长,它也要忧郁地吠叫;思索一下狗的这种情感是多么奇妙吧。我们将把注意力局限于高等社会性动物;至于昆虫,则略去不谈,虽然它们有些也是社会性的,而且以许多重要方式彼此互助。在高等动物中最普通的相互服务,就是利用全体的统一感觉彼此发出危险警告。正如耶格尔(Jaeger)博士 (255) 所说的,每一个猎人都知道,要想接近成群的动物是多么困难。我相信野马和野牛不发任何危险信号;但是,它们当中的任何一个最先发现敌对者时,就会用姿态来警告其他成员。兔用后腿跺地发出高声作为信号:羊和小羚羊则用前脚跺地,发出的声响好像口哨,以为信号。许多鸟类以及某些哺乳类动物都放岗哨,据说海豹一般是由雌者担当这项任务的。 (256) 一群猴的头头所作所为均如岗哨,它发出表示危险以及表示安全的叫声。 (257) 社会性动物彼此还做些小服务:马彼此互啃痒处,牛则彼此互舐痒处;猴彼此捉身上的寄生虫;布雷姆叙述,当一群灰绿长尾猴(Cercopithecus griseoviridis)冲过一片棘刺很多的林丛之后,各猴都在树枝上伸展肢体,另一只猴坐在旁边,“认真地检查它的毛皮,把每一根棘刺都拔掉”。
动物彼此服务,还有更为重要的:例如,狼以及某些其他食肉兽成群猎食,在攻击其猎物时彼此互助。鹈鹕(pelicans)捉鱼时相互协作。埃塞俄比亚鼯猴一齐翻转石头去找昆虫,等等;当遇到一块大石头时,在它周围能站多少只就站多少只,共同把它推翻,而且分享所获之物。社会性动物还彼此相助以保卫自己。北美野牛(bison)当有危险时就把母牛和牛犊赶到牛群的当中,它们在外围进行防卫。我还要在下一章举出一项记载,表明奇呤哈姆园囿中的两头小野公牛彼此协作向一头老公牛进行攻击,还有两匹公马一齐试图把另一匹公马从母马群中赶跑。布雷姆曾在埃塞俄比亚遇到过一大群狒狒,它们正在穿过一个山谷;有些已经登到对面的山上,有些还在山谷中:这时众狗向后者发动攻击,于是老雄狒狒立即从山上急驰而下,大张其口,凶猛吼叫,以致众狗吓得疾引而退。跟着众狗受到鼓动,再次进行攻击;不过所有狒狒这时已登上山顶,但还落下一只六个月左右的小狒狒,它高声呼助,爬上一块岩石,并且受到了众狗的包围。这时一只最大的雄狒狒,一位真正的英雄,又从山上下来,徐徐走近那只小狒狒,哄着它,得意洋洋地让它走开——众狗对此感到惊讶不止,以致停止了攻击。我不能不谈一谈另一个场面,这是上述同一位博物学者亲眼所见的:有一只鹰抓住了一只小长尾猴,由于它紧紧握住树枝,没有能够立即把它带走;这只小长尾猴高声呼助,在树上的这群猴的其他成员大肆喧嚣,急来相救,把那只鹰团团围住,拔掉它的羽毛如此之多,以致它不再想到捕获物,而只得考虑如何溜之大吉了。正如布雷姆所说的,这只鹰肯定永远不会再攻击猴群中的单独一只猴了。 (258)
合群的动物肯定有一种彼此相爱的感情,不合群的成年动物没有这种感情。在大多数场合中,它们对于其他动物的痛苦和快乐实际上究竟能同情到怎样程度,还是很可疑的,尤其关于快乐是如此。巴克斯顿(Buxton)先生掌握了极好的观察方法 (259) ,然而他写道,他在诺福克(Norfo1k)自由放养的金刚鹦鹉(macaws)对一对有巢的同类非常有兴趣;每当那只雌鸟离巢的时候,就被群鸟围住,呜呜地狂叫,以表尊敬。动物对其同类其他成员的痛苦是否抱有什么感情,常常是难以判断的。当众牛环绕并且目不转睛地注视其将死的或死去的同伴时,谁能说出它们有何种感觉呢;然而,如赫祖所说,它们显然并无怜悯之情。动物有时完全没有同情感,是非常确实的;因为,它们把受伤的动物赶出群外,或者把它们抵死,要不就把它们咬死。这几乎是博物学中一个最黑暗的事实,除非对这个事实所提出的解释是正确的,即,它们的本能或理性导致它们把一个受伤的同伴赶出群外,免得食肉兽——包括人类在内——被引诱去追猎全群。在这种情况下,它们的行为并不比北美印第安人的更坏,后者把病弱的亲密同伴丢在荒原之上任其死亡;或者,也不比斐济人(Fijians)的行为更坏,他们把年老的或患病的父母活活埋掉。 (260)
然而,许多动物肯定彼此同情对方的苦痛或危险。甚至鸟类亦复如此。斯坦斯伯里(Stansbury)船长 (261) 在犹他(Utah)的一个盐湖上发现一只完全瞎了的老鹈鹕,但它很肥,一定曾经长期由其同伴给予很好的喂养。布赖茨先生告诉我说,他看见过印度的母牛喂养两三头瞎牛;我曾听说过一个近似的事例,是关于家养雄鸡的。如果我们喜欢把这等行为认为是本能的,那也可以;不过对于任何特殊本能的发展来说,这等例子实在是太少了。 (262) 我亲自见到一只狗,是一只猫的伟大朋友,当这只猫卧病在篮中时,那只狗每次经过那里,总要用舌头把猫舐几下,这是狗表示亲善感情的最可靠信号。
一只勇敢的狗当其主人受到任何人的攻击时,它一定向他们猛扑上去,引导狗这样行动的,一定可以叫做同情心。我曾看到一个人假装去打一位妇女,在她的膝上正好有一条胆怯的小狗,而且以前从未做过这样的试验;这个小东西立刻跳下来跑开了,但当假装的殴打完了之后,它是多么固执地要舐女主人的脸,对她进行安慰,看到这种情景的确使人感动。布雷姆 (263) 陈述,当对一只圈养的狒狒实行惩罚时,其他狒狒就努力保护它。在上述场合中,导致狒狒和长尾猴去保护它们幼小的亲密同伴不受狗和鹰侵害的,一定是同情心。我再举另外一个有关同情的和英雄的行为的事例,这是关于小美洲猴的。几年之前伦敦动物园的一位饲养员叫我看他颈背上一条刚刚愈合的深伤痕,那是他跪在地板上时被一只凶猛的狒狒弄伤的。有一只小美洲猴,是这位饲养员的亲密朋友,它同那只大个狒狒居住在同一大间猴室内,而且对狒狒怕得要命。尽管如此,小美洲猴一看到它的朋友处于危险之中,还是立即猛冲来救,狂叫乱咬,把那只狒狒弄得晕头转向,饲养员才得以跑开,事后外科医生认为他逃脱了一次生命的大危险。
除去爱和同情之外,动物还表现有同社会本能有关系的其他属性,这在人类来说可以称为道德;我同意阿加西斯(Agassiz) (264) 的看法,他认为狗也具有某种很像良心那样的品质。
狗有某种自制的能力,看来这并不完全是恐惧的结果。布劳巴哈说,狗当主人不在时会抑制自己不偷吃东西。 (265) 长期以来大家都承认狗是忠诚和顺从的真正模范。但象同样也是很忠于驾象人或饲养人的,可能把他们视为象群的领袖。胡克(Hooker)博士告诉我说,他在印度骑的一头象有一次陷入泥沼中如此之深,以致到次日都无法自拔,后来还是用绳索把它从泥沼中拉出来的。在这种情况下,象总是用鼻子卷住任何东西,不管是活的还是死的,把它们放在膝下,以免在泥沼中陷得更深;这时驾象人深怕胡克博士被捉到,被踩死。但胡克博士有把握地说,驾象人自己那时不会有这种危险。这样沉重的动物在如此可怕的危急中所表现的自制,乃是其高尚忠诚品质的惊人证明。 (266)
所有合群生活的并且彼此协同保卫自己或攻击敌对者的动物,在某种程度上一定是彼此忠实的;而那些追随一个领袖的动物,在某种程度上一定是服从的。在埃塞俄比亚,当一群狒狒劫掠果园时,它们毫不做声地追随着领头的狒狒;如果有一只冒失的小狒狒竟然喧闹,别的狒狒就会给它一掌,教它安静和服从。 (267) 高尔顿先生有极好的机会去观察南非的半野生牛,他说,它们甚至片刻也不离开牛群。 (268) 它们本质上是奴性的,接受公共的决定;如果被任何一头有足够自信心担任领导的公牛去领导它们,那就是碰上了最好的运气。训练这等牛作为使役之用的人们孜孜不倦地注视着那些离群吃草而表现有自信心的牛,并且把这样的牛作为带头牛进行训练。高尔顿先生接着又说,这样的牛是罕见的而且是值钱的;如果生下来的牛很多是这样的话,它们很快就要被消灭掉了,因为狮子总是注意那些离群徘徊的个体。
关于引导某些动物联合在一起并且以多种方式彼此互助的冲动,我们可以推论,在大多数场合中是由实行其他本能活动时所体验到的同样满足感或快乐感来推动的;要不就是当其他本能活动受到抑制时,由同样的不满足感来推动的。我们在无数事例中看到这种情形;而且由我们家养动物后天获得的本能以显著的方式给予了阐明;例如,一只年幼的牧羊狗(shepherd dog)以驱赶和驰绕羊群为乐,但并不咬它们;一只年幼的猎狐狗以猎狐为乐,而有些其他种类的狗,如我亲眼所见,却完全不理会狐。一定有一种非常强烈的内在满足感推动着一只充满活动力的鸟日复一日地去孵卵。候鸟如被阻止不能迁徙,是会十分痛苦的;也许它们会享受开始长途飞行的乐趣;奥杜邦(Audubon)描写一些可怜的不会飞的鹅(goose)到了一定时期也要开始徒步跋涉约1000英里以上,很难相信它们对此会感到什么乐趣。有些本能完全是由痛苦感情、如恐惧所决定的,恐惧会导致自我保存,并且在某些场合中是指向特种敌对者的。我设想,没有人能够分析快乐的或痛苦的感觉。然而,在许多事例中大概是,仅仅由于遗传的力量,本能就会固执地发生,而无须快乐或痛苦的刺激。一只年幼的向导猎狗(pointer)当第一次嗅出猎物时,显然不会不把头指向猎物。笼中松鼠轻轻拍打那些它不能吃掉的坚果,好像要把它们埋人地下,简直无法想象它们这样做是由于快乐,还是由于痛苦。因此,通常假定人们的每一个行为一定都是由快乐的或痛苦的经验所推动,可能是错误的。虽然遵从一种习性可能是盲动的和含蓄不明的,而且那时既不感到快乐,也不感到痛苦,但是,如果它突然地受到强有力的抑制,一般就会体验到一种不满足的模糊感觉。
常有这样假设:动物原本就是社会性的,其结果便是它们在彼此离散之后感到不舒适,而群居在一起则感到舒适;但可能更合理的观点是,这等感觉的最初发展,乃是为了诱使那些可以从社会生活中获益的动物彼此生活在一起,其方式正如最初获得饥饿的感觉和饮食的愉快无疑是为了诱使动物去吃食。来自社会的愉快情感大概是亲与子爱情的延伸,因为社会本能的发展似乎是由于幼儿同双亲长期逗留在一起所致;这种延伸局部地可归因于习性,但主要地还应归因于自然选择。就那些在生活中密切联系而获得利益的动物而言;最喜欢群居的个体将会最好地躲避各种危险,而那些最不照顾同伙而独居生活的个体将会较大数量地死亡。亲与子的爱情起源,显然是以社会本能为基础的,我们还不知道它们是经过怎样的步骤而被获得的;但我们可以推论,在很大程度上是通过自然选择。关于最近亲属之间的异常而相反的憎恨感情,几乎肯定也是如此,如工蜂弄死其雄蜂兄弟以及后蜂弄死其女儿皆是;在这样场合中毁灭其最近亲属的欲望对群体是有利的。双亲之爱,或者代替它的某种感情,在某些极端低等的动物,如海星(starfish)和蜘蛛中也有所发展。在动物的整个类群中间或只有少数成员表现有这种感情,如球螋属(Forficula)或蠼螋即是。
最重要的同情感和爱是有区别的。母亲热爱她的熟睡而默从的婴儿,但简直不能说她在那样时刻是对婴儿同情。人对狗的爱是和同情有区别的,狗对其主人的爱亦复如此。亚当·史密斯(Adam Smith)以前曾辩说,最近贝恩先生也这样辩说:同情感的基础是建筑在我们强烈保持着以往痛苦或快乐的状态之上的。因此,当看到另一个人饥饿、寒冷、疲劳时,就会唤起我们对这等情况的回忆,“甚至在观念中也要使人痛苦”。这样,我们就被推动着去解脱他人的痛苦,为了我们自己的痛苦感情同时也可得到解脱。我们以相似的方式去分享他人的快乐。 (269) 但我无法理解这个观点如何解释下面的事实,即,由被爱的人比被不关心的人所激起的同情,其程度之强烈要大至不可估量。仅仅看到同爱无关的痛苦,就足可以唤起我们鲜明的回忆和联想。其解释可能在于如下的事实:在所有动物中,同情是专门指向同群的诸成员的,所以是指向相识的以及多少相爱的诸成员的,而不是指向同一物种的所有个体。这一事实并不比许多动物专门畏惧特殊的动物更令人惊奇。非社会性的物种,如狮和虎,对于自己的幼兽痛苦无疑感到同情,而对于任何其他动物的幼兽并不如此。正如贝恩阐明的,关于人类,在同情能力中大概还可加入自私、经验和模仿;因为我们对他人同情的友好行为,是希望在报答中得到好处所致;而且同情由于习性而大大被加强了。不管这种感情的起源多么复杂,由于对所有那些彼此帮助、相互保卫的动物来说,同情乃是最重要的感情之一,所以它将通过自然选择而被增强;这是因为包含最大数量的最富同情的成员的那些群体将最繁盛,而且会养育最大数量的后代。
然而,在许多场合中不可能决定某些社会本能究竟是通过自然选择获得的,还是其他本能和官能如同情、理性、经验以及模仿倾向的间接结果;或者,它们是否为习性长期连续实行的单纯结果。像设置岗哨向其同群发出危险警告那样的一种如此显著的本能,几乎也不会是任何这等官能的间接结果,所以它一定是被直接获得的。另一方面,某些社会性动物的雄者所遵循的保卫群体的习性,以及协同攻击敌对者或猎物的习性,也许起源于相互同情;但勇气以及在许多场合中的力气,一定是以前获得的,这大概要通过自然选择。
在各种本能和习性中,有些比另外一些要强得多;或者大概同等重要的是,它们通过遗传会更加持久地被遵循,而不激起任何快乐或痛苦的特殊感情。我们会自觉到,自己有些习性远比另外一些习性难于矫正或改变。因此,可以常常观察到在动物中不同本能之间的以及一种本能和某种习性之间的斗争;例如,当一只狗追逐一只兔而被制止时,它踌躇不前,再起追逐,或羞愧地回到主人身旁;又如,一只母狗对其狗仔的爱和对其主人的爱之间的斗争——当这母狗鬼鬼祟祟地溜到狗仔那里时,好像没有能够陪伴主人而感到有点羞愧。但是,关于一种本能战胜另一种本能,我所知道的一个最奇妙的事例是,候鸟迁徙的本能胜过了母性的本能。前一种本能之强令人吃惊;到了迁徙季节,被拘禁的鸟就会以胸部撞击鸟笼的铁丝,直到把毛撞光和流血为止。这种本能还致使年幼的鲑鱼(salmon)跳出它们本可在其中继续生存的淡水之外,这样就无意识地自杀了。每一个人都知道,母性本能是何等之强,它甚至可以导致怯懦的鸟类为了保护幼鸟去面对巨大的危险,虽不免有些踌躇,而且它同自我保存的本能正好背道而驰。尽管如此,候鸟迁徙的本能还是如此强有力,以致燕子、家燕和东亚雨燕到了晚秋季节往往丢弃它们的弱小幼鸟,而进行迁徙,任幼鸟在巢中悲惨地死去。 (270)
我们可以理解,如果一种本能的冲动无论在什么方面都比另外某种本能或相对立的本能更有利于一个物种的话,那么它就会通过自然选择在二者之中成为更强有力的;因为这种本能最强烈发达的诸个体将会较大数量地生存下来。然而,关于候鸟迁徙本能和母性本能的比较,情况是否如此,尚属疑问。在一年的某些季节中迁徙本能整天整日所表现的这种巨大固执性或稳定活动,可能暂时给予它以重大力量。
人类是一种社会性动物
任何人都会承认人类是一种社会性动物。从人类不喜欢孤独以及要求自己家庭之外的社会生活,我们可以看出这一点。单身监禁是人所受的最严厉惩罚之一。有些作者设想人类原本是营单独家庭生活的;但时至今日,虽然单独家庭,或仅二三家庭相集,漫游于野蛮荒凉之地,就我所能发现的来说,他们总是同居住在同一地区的其他家庭保持着友好的关系。这等家庭不时集会协商,团结起来共同防卫。居住相邻地区的部落彼此几乎争战不绝,但这不能作为反对未开化人是一种社会性动物的论据;因为社会本能从来不会延伸到同一物种的一切个体。从大多数四手类的相似性来判断,人类的早期类猿祖先很可能同样也是社会性的;不过这对我们并没有多大重要性。虽然像现今生存的人类那样,仅有少数特殊的本能,并且失去了其早期祖先可能有的任何本能,但这并不能作为理由来说明人类为什么不应从远古时代起就对其同伴保持某种程度的本能之爱和同情。我们每一个人一定都会意识到我们确有这种同情感; (271) 但我们的意识没有告诉我们,这种感情是否为本能的,就像低于人类的动物那样起源于很久以前,或者,它们是否为我们每一个人在其生命早期所获得的。由于人类是一种社会性动物,几乎可以肯定他将遗传这样一种倾向,即:对他的同伙忠实,并对他的部落领袖服从;因为这等属性是大多数社会性动物所共有的。结果他将具有一定的自制能力。他由于一种遗传的倾向,甘心情愿同其他人协力保卫他的同胞;如果不过多地同其自身利益或其自身强烈欲望相抵触,他将乐于以任何方式对其同胞进行帮助。
最低等的社会性动物对其同群诸成员所给予的帮助,几乎完全受特殊本能所支配,而较高等的社会性动物所给予的这种帮助则大部分受特殊本能所支配,同时部分地还被互爱和同情所推动,此外还有相当的理性帮助。虽然人类像刚才所说的那样,并没有特殊本能告诉他去如何帮助其同胞,但他仍然有这种冲动,并且由于他有进步的智力,在这方面自然要大大被理性和经验所支配。本能的同情还会使他高度评价同伴们的称赞;因为,正如贝恩先生所明确阐述的,对受表扬的喜爱,对荣誉的强烈感觉,以及还要更加强烈地对蔑视和臭名的恐惧感,乃是“由于同情的作用”。 (272) 因而人类就要最高度地被其同胞用姿态和语言表达出来的愿望、称赞以及谴责所影响。这样,社会本能一定是当人类还处于很原始状态时就获得的,而且很可能甚至人类的早期类猿祖先就已经获得了社会本能,人类那时的这种本能仍然产生冲动以实行某些最良好的行为;不过人类的行为在较大程度上是由其同胞所表示的愿望和裁判来决定的,不幸的是,还常常由他自己的强烈自私欲望来决定。但是,由于爱、同情以及自制通过习性而被加强,而且由于推理的能力日益变得清晰,所以人类能够合理地评价同伴们的评判,他将感到自己必须撇开暂时的快乐或痛苦,被迫遵从一定的行为路线。于是他可能宣告——任何野蛮人或未开化人都不会有这样想法——我是我自己行为的至高无上的裁判者,用康德的话来说,我不愿亲自侵犯人类的尊严。
比较持久的社会本能征服比较不持久的本能
然而,关于按照我们现今观点来看的整个道德观念问题的主要之点,迄今尚未论及。为什么一个人会感到他应该服从某一本能的欲望,而不是服从另一欲望?如果一个人屈服于强烈的自我保存感,而没有冒生命的危险去挽救同伴的生命,为什么他会痛苦地后悔不已?为什么由于饥饿而曾偷窃食物也会使他后悔?
首先,本能的冲动在人类中显然具有不同程度的力量:一个未开化人会冒生命的危险去挽救一个同群成员的生命,而对一个陌生人就完全漠不关心了;一位怯弱的年轻母亲在母性本能的推动之下,为她自己的婴儿会毫不踌躇地去冒最大的危险,而对于其同群的人就不会这样做。尽管如此,许多文明人,甚至一个少年,虽然以前未曾为他人冒过生命危险,但还充满了勇气和同情,无视自我保存的本能,立刻投入急流之中去挽救一个溺水的人,即使这是一个素不相识的人。在这种场合中,推动人类这样去做的本能的动机,和上述致使英雄的小美洲猴为了挽救其饲养员而去攻击可怕的大狒狒的那种本能的动机是一样的。上面这等行为似乎是社会本能或母性本能的力量大于任何其他本能或动机的力量的简单结果;因为那是瞬间决定实行的,以致当时没有工夫去考虑或感到快乐和痛苦;但如果受到任何原因的阻止,还会感到苦恼甚至悲痛。另一方面,对于一个胆怯的人来说,他的自我保存的本能可能非常强烈,以致他不能迫使自己去冒任何这种危险,甚至对他自己的小孩恐怕也会如此。
我知道有些人主张上述那些起于冲动的行为不受道德观念的支配,因而不能称为道德。他们把这一名词限于那些战胜相反欲望后而审慎实行的行为,或者那些在某种崇高动机的激励下而审慎实行的行为。但是,要想划出这种区别的明显界线 (273) ,似乎很少可能。就崇高动机来说,曾经记载过许多关于未开化人的事例,他们对人类缺少任何博爱的感情,而且不受任何宗教动机的支配,却宁愿作为俘虏而从容就义 (274) ,也不背叛他们的同伙;他们这种行为确可视为道德。就审慎以及战胜相反动机来说,我们可以看到当动物从危险中拯救其后代或同伙时在相反的本能之间所表现的迟疑不决;然而它们的行为虽然是为了其他动物的利益而实行的,却不能称为道德。再者,任何事情只要我们经常去做,最终就会不经过深思熟虑或毫不踌躇地去做;于是这同本能就无法加以区别了;然而肯定没有人会妄称这样一种行为并不是道德。恰恰相反,除非一种行为的完成系出于冲动,没有经过深思熟虑或努力,正如一个人需要有内在品质才能做到的那样,否则我们莫不感到这种行为不能被视作完善的或者是以最高尚方式来完成的。然而,一个人在完成一种行为之前,被迫去克服他的恐惧或缺少同情心,从某方面来看,将比一个不经过努力而由内在倾向引导着去完成一种良好行为的人,将会受到更高的称赞。由于我们无法对不同动机之间加以区别,所以我们只好把某一类的一切行为都纳人道德的范畴,如果这是由一种有道德的生物所完成的话。所谓有道德的生物乃是这样一种生物,它能对过去的和未来的行为或动机进行比较,而且能赞成哪些或反对哪些。我们没有理由来假定任何低于人类的动物具有这种能力;所以,一条纽芬兰狗(Newfoundland dog)拖出一个落水的小孩,一只猴面对危险去营救它的同伙或抚养一只失去母猴的幼猴,我们都不把这种行为称为道德的。但是,毫无疑问只有人类才能被纳入有道德的生物的地位,在人类的场合中某一类行为,不论是经过与相反动机的斗争后而深思熟虑地完成的,还是出于本能的冲动,或者是由于缓慢获得的习性的效果,都可称为道德的。
现在回头来讨论一下我们更直接的问题。虽然某些本能比另外一些本能更加强有力,而且由此导致了相应行为的发生,但是,要说人类的社会本能(包括喜爱称赞和惧怕谴责)比自我保存、饥饿、色欲、报复等本能具有更大的力量,或者说通过长期的习性获得了更大的力量,还是站不住脚的。那么,为什么人类会对他遵从了某一自然冲动而没有遵从另一自然冲动而感到遗憾,纵使他想排除这种遗憾而不可得?而且,为什么他会进一步感到他应该对他的行为有所遗憾?关于这一点,人类同低于人类的动物有深刻的差别。不过,我想我们在某种程度上还能清晰地理解这种差别的原因。
人类,由于他的心理官能的活动,无法不进行思考:过去的印象和意象不断地而且清晰地在他头脑中通过。关于那些永久在一块儿生活的动物,其社会本能是永远存在的,而且是持续的。这等动物总是随时发出危险的信号,保卫群体;并且按照它们的习性对其同伴提供援助;它们不论何时对其同伴都感到某种程度的爱和同情,而无须任何特殊的激情或欲望;它们如果长期和其同伴分离就会不愉快,如果和其同伴重聚就会高兴。而我们自己亦复如此。甚至当我们十分孤独的时候,我们还常常想到别人对自己的评价——想象中的他们对自己的褒贬;所有这一切都来自同情,而同情乃是社会本能的基本要素。连这等本能一点痕迹都没有的人大概是一个反常的怪物。另一方面,满足饥饿的欲望,或者像报复那样的任何激情,在其性质上都是暂时的,所以能够暂时地得到充分满足。完全逼真地唤起像饥饿那样的感觉是不容易的,也许几乎是不可能的;正如常常提到的,任何痛苦的感觉确实都是如此。除非在有危险的情况下,不会感到自我保存的本能;许多懦夫非面逢仇敌不会感到自己的勇气。占有别人产业的希图也许是可以举出的最固执的一种欲望;即使在这一场合中,实际占有得到满足后的感情一般也比占有的欲望为弱:许多贼,如果不是惯贼,在偷窃既遂之后,也不免对他为什么要偷东西感到惊讶。 (275)
一个人无法阻止过去的印象重新通过他的头脑;这样,他就要把过去的饥饿、报复,牺牲别人以避免危险等印象与几乎永远存在的同情的本能加以比较,而且还要与他对他人所给予的褒贬的早期认识加以比较。这种认识无法从他的头脑中排除,并且由于本能的同情,它还要受到高度的评价。于是在遵从现在的本能或习性时,他将会感到好像畏缩不前,这对所有动物来说,都会引起不满足甚至痛苦。
上述有关燕子的例子虽然具有相反的性质,但它阐明了一个暂时的,但眼下是强烈固执的本能征服了平时凌驾一切之上的另一种本能。到了适当季节,这等鸟似乎终日为迁徙的欲望所迫;它们的习性改变了;它们变得惶惶不安,喧噪而群集于一处,当母鸟饲喂它的雏鸟或孵卵时,母性本能大概大于迁徙本能;但是,更为固执的本能获得了胜利,最后,当她看不见群雏的那一刹那,便马上起飞而遗弃了它们。当到达她的长途旅程终点并且迁徙本能停止活动时,如果她赋有巨大的心理活动力,而无法阻止有关她的幼雏在凄凉的北方死于饥寒交迫之中的意象不断地通过她的头脑,那么她将会感到由悔恨而引起多么强烈的痛苦。
在人类有所行为的当时,无疑他将易于遵从较强的冲动;这种冲动虽然有时会促使他取得最高尚的业绩,但更加普通的是引导他牺牲别人以满足自己的欲望。不过,他的欲望一经得到满足之后,如果过去的和较弱的印象受到永恒的社会本能的评判并且还要受到敬重同伴们善良公意的评判,那么内心的惩罚肯定将会来临。这时他将感到后悔、遗憾或羞耻;然而羞耻这种感情几乎完全与别人的评判有关。结果他将有多大程度地决定将来不再有这种行为了;这就是良心;因为良心鉴于既往而指导将来。
被我们称为遗憾、羞耻、后悔或悔恨的那些感情,其性质和力量不仅决定于受到侵犯的本能的力量,而且局部地决定于诱惑的力量,往往还要更多地决定于我们同伴们的评判。每个人对别人的称赞重视到什么程度,决定于其内在的或后天获得的同情感;而且还决定于对其行为的遥远后果的理解能力。另一个要素虽不是必然的,却极重要,即每个人对其所信仰的神或鬼的崇敬或畏惧:在悔恨的场合中尤其如此。有几位评论家持有反对意见,他们认为,有些轻微的遗憾或后悔虽然可以用本章所提出的观点来解释,但这样去解释那种震动灵魂的悔恨感情却是不可能的。但我看不出这种反对意见有多大力量。这些评论家们并没有对他们所谓的悔恨下过什么定义,我以为最合适的定义就是,悔恨乃为占有压倒之势的后悔感。悔恨同后悔的关系恰如狂怒同怒或者极度痛苦同痛苦的关系一样。一种非常强烈而且非常受到普遍称赞的本能,如母爱,如果没有被遵从的话,那么引起这种未被遵从的过去印象一旦有所减弱,就会引起最深刻的悲痛,这一点也不奇怪。甚至一种行为同任何特殊本能并不相反,仅仅由于知道朋友们和地位相等的人们鄙视自己,也足可以招致巨大的悲痛。由于恐惧而拒绝决斗曾使许多人感到羞耻的极度痛苦,谁还能对此有所怀疑呢?据说,许多印度教徒由于吃了不洁净的食物,其灵魂深处都要激动起来。这里还有另外一个事例,我以为一定可以称为悔恨。兰多尔(Landor)博士曾是澳大利亚西部的地方行政官,说道,在他的农庄内,“有一个土著居民,其众妻之一因病死去之后,他来说,他将到一个远方部落用矛刺杀一个妇人,以满足对他妻子的义务感。我告诉他说,如果他这样干,我就要把他送去终身监禁。他在农庄又待了几个月之后,显得异常消瘦,并且抱怨说,他无法睡眠,也不能吃东西,他的妻子的幽灵总是缠绕着他,因为他没有为亡妻取来一条生命之故。我坚决不为他所动,并且使他确信,如果他这样干,什么也不能挽救他”。 (276) 尽管如此,这个人还是失踪了一年多,然后意气昂扬地回来了;他的另一个妻子告诉兰多尔博士说,她的丈夫从一个远方部落取来了一个妇人的生命;但是关于他的行为不可能得到法律的证据。可见一个部落所视为神圣的准则如被违反,就会引起极深刻的感情——而这种感情同社会本能完全无关,除非这种准则是以同群的评判为基础的。全世界许多奇异迷信是怎样起源的,我们还不知道,我们也无法说出最低等的未开化人为什么憎恶某些真正的重大罪恶,如乱伦(然而这并不十分普遍)。甚至可以怀疑,有些部落是否认为乱伦比同姓的,但没有亲属关系的男女结婚更可嫌忌。“澳洲人认为违犯这一法律就是罪恶,他们最憎恶这种罪恶;北美的某些部落也完全如此。无论在上述任何一个地方问道,杀死一个远方部落的妇女和娶一个本族的女子这两件事,哪一件更坏,他们将会给予正和我们相反的答复”。 (277) 因此,我们可以否定某些作者最近坚持的那种信念,即认为对乱伦的憎恶乃是由于我们具有一种特殊的、由上帝植入的良心。总之,一个人被教导去相信作为一种赎罪应该自行投案要求审判,可以理解导致他有这样行为的乃是由于他受到了如此强有力的一种思想感情,如悔恨所推动,虽然悔恨有上述那样的起因。
受到良心驱使的人通过长期的习性将获得完全的自制,这样,他的欲望和情欲最终就会不经斗争而直接屈服于他的社会同情心和社会本能,其中也包括他对同伴评判的感觉。依然饥饿的或依然充满仇恨的人将不会想到偷窃食物或实行报复。就像我们以后将看到的那样,自制的习性正如其他习性,可能、甚至很可能是遗传的。这样,通过后天获得的以及也许遗传的习性,人类最终会感到,对他来说最好是遵从他的比较固执的冲动。“应该”这个专横的词似乎仅仅是针对意识到行为准则的存在而言,不论这种意识是如何发生的。以前一定常常热烈地主张,一位有身份的人如果受到侮辱,就应该进行决斗。我们甚至说,向导猎狗应该用头指向猎物,拾物猎狗应该衔回被击中的猎物。如果它们没有这样做,那就是它们没有尽到义务,而且行为失误。
如果导致违犯他人利益的任何欲望或本能仍然出现,而且当在头脑中回忆及此时,其强烈程度同社会本能相等,或者还要超过后者,那么这个人对于曾经遵从这种欲望或本能就不会感到深刻的遗憾;但他会意识到,如果他的行为被他的同伴们知道,就要受到谴责;倘发生这种情形而不感到不安,像这样缺乏同情心的人还是很少。如果他没有这种同情心,导致这种坏行为的欲望很强,而且当回忆时也没有被社会本能以及他人的评判所克服,那么他本质上就是一个坏人 (278) ;剩下来的唯一抑制的动机就是对惩罚的畏惧;以及深信为了自私的利益从长远看与其注重自己的利益莫如注重他人的利益。
如果一个人的欲望并没有侵犯他的社会本能,这就是说没有侵犯他人的利益,显然他可以问心无愧地满足他的欲望;但是,为了完全不受自责,至少不受忧虑不安的影响,那么避免同胞们的谴责——不论合理与否,对他来说几乎还是必要的。他还一定不会打破他的生活习惯,特别是这等生活习惯合乎情理时,尤其如此;因为,他如果这样做了,肯定要感到不满足。按照他的知识或迷信,可能信仰一个上帝或多神,因此他还一定要避免上帝或多神的摒弃,不过在这种场合中,对神罚的恐惧常常伴随发生。
最初受到重视的仅为严格的社会美德
上述关于道德观念——它告诉我们应该做的是什么——的起源及其性质的观点,以及关于良心——如果我们违背它就要受到谴责——的起源及其性质的观点,同我们看到的人类这种官能的早期不发达状态很一致。原始人类的美德至少是普遍实行的,所以他们才能联成一体,那些美德至今仍被认为是最重要的。但是,这些美德几乎专门施于同一部落的人,而与此相反行为如果施于其他部落的人则不视为罪恶。如果凶杀、抢劫、叛变等盛行,任何部落都无法团结一致,因而这等罪恶在同一部落的范围内就要“被打上千古臭名的烙印”; (279) 但超出这等范围之外,就不会激起这种思想感情了。北美印第安人如能剥取其他部落一个人的头皮,自己就会感到十分高兴,而且还会得到别人的尊敬;达雅克人(Dyak) (280) 割掉一个无辜人的头,并把它晾干作为战利品。杀婴以极大规模通行于全世界, (281) 并没有受到谴责;杀婴、特别杀女婴曾被认为对部落有好处,至少没有害处。自杀在以往时代里并没有被普遍视为一种罪恶 (282) ,且由于显示了勇气,反被视为一种光荣的行为;有些半开化民族以及未开化民族至今仍然实行自杀而不受到谴责,显然这种行为同部落的其他人并无利害关系。曾经记载,印第安的萨哥人(Thug)对于他自己抢劫和勒死过往行人没有能够像以前他父亲干的那样多,从良心上感到遗憾。在原始的文明状态下,抢劫陌生人诚然被视为光荣。
奴隶制度在古代虽然有某些方面的益处, (283) 却是一种大罪恶;然而在最近以前并不这样认为,甚至最文明的民族也是如此。由于奴隶一般属于和其主人不相同的种族,情况就尤其是那样了。因为野蛮人不重视妇女的意见,所以普遍对待妻子就像对待奴隶一样。大多数未开化人对于陌生人所遭受的痛苦完全漠不关心,甚至以目睹此事为乐。众所熟知,北美印第安人的妇女和儿童在对敌人施行严刑拷打时,也从旁相助。有些未开化人以虐待动物作为消遣 (284) ,这种行为令人发指,但对他们来说,人性还是一种未知的美德。尽管如此,除了家族的感情之外,同一部落诸成员之间的友好行为还是普遍的,尤其在有人患病期间更加如此,这种友好行为有时会扩展到这等范围以外。芒戈·帕克(Mungo Park)关于非洲腹地黑人妇女对其友好行为的动人记载,是众所熟知的。未开化人彼此高尚地忠诚相待,但对陌生人并不如此,关于这一点可以举出许多事例;普通经验证实了西班牙人的一句格言:“万万不可信任印第安人。”无诚实则无忠诚;诚实这一基本美德在同一部落诸成员之间并非罕见。例如,芒戈·帕克曾听到黑人妇女教育她们的孩子们要热爱诚实。再者,这是头脑中如此根深蒂固的美德之一,以致未开化人有时甚至不惜重大代价而施此美德于陌生人;但是,向敌人说谎却很少被认为是一种罪过,近代外交史非常明显地展示了这一点。部落一旦有了一个公认的领袖,不服从就会成为一种罪恶,而且,甚至卑鄙的屈服也被视为神圣的美德。
在原始时代,一个人如果缺少勇气就不会有益于或忠实于他的部落,所以这一品质普遍被列入最高的等级;在文明国度里,一个善良而怯懦的人可能远比一个勇敢的人对群体更为有益,但我们还是禁不住本能地尊敬后者,不管懦夫多么乐善好施都是一样。另一方面,同他人福利无关的慎重,虽为一种很有益的美德,却从来没有受到高度的尊重。如果不能自我牺牲、不能自制以及没有忍耐力,就无法实行为部落福利所必需的那些美德,所以对于这等品质无论何时都高度地而且公正地给予了评价。美洲未开化人甘受最可怕的酷刑而不发一点呻吟,以证明和增强他的毅力和勇气;我们对他不得不加以称赞,甚至对印第安的法基尔人(Indian Fakir),由于一种宗教动机而把铁钩插入肉中悬空摆动,我们也要加以称赞。
另一种所谓自重的美德,对部落福利的影响虽不明显,但确实存在,未开化人从来不尊重这种美德,而现今却受到文明民族的高度欣赏。未开化人并不谴责最无节制的放纵生活。极度的淫荡生活以及鸡奸流行之广,已达到使人震惊的程度。 (285) 然而,一夫多妻或一夫一妻的婚姻一旦普及之后,嫉妒就会导致妇女美德的反复灌输,这种美德受到尊重后,就倾向于扩大到未婚妇女。而它扩大到男性却非常缓慢,我们在今天还可以看到这种情形。贞洁显著地需要自制;所以在文明人的道德史中,自古以来它就受到了尊重。其结果便是,毫无意义的独身生活自古以来就被列为一种美德。 (286) 对下流猥亵的憎恶,在我们看来是如此自然,以致被认为是天生的,它对贞洁是一种多么可贵的帮助,这是一种近代的美德,正如斯汤顿(G.Staunton)爵士 (287) 所指出的,它专属于文明生活。这从各个不同民族的古代宗教仪式,从庞贝(Pompeii)古都的壁画,以及从许多未开化人的习俗,都可以得到阐明。
于是我们可以知道,未开化人认为,很可能原始人类也认为,行为是好或是坏,显然仅仅看它们对部落福利的影响如何,并不考虑它们对种族以及对部落的个体成员有何影响。这一结论同以下的信念十分符合,即,所谓道德观念原本发生于社会本能,因为二者在最初都只与群体有关。
如果按照我们的标准去衡量,未开化人道德低下的主要原因为:第一,同情仅限于同一部落。第二,其推理能力不足,不能认识许多美德、特别是自重美德同部落一般福利的关系。例如,未开化人无从探知大量罪恶是由缺少节制、贞洁等所引起的。第三,自制力薄弱;因为这种能力没有通过长期连续的,也许是遗传的习性,更没有通过教育和宗教而被加强。
我之所以对未开化人的不道德 (288) 进行如上的详细讨论,是因为有些作者最近高度估量了他们的道德本性,或者把他们的大部分罪恶归因于仁慈的误用。 (289) 这些作者的结论似乎是依据未开化人所具有的那些美德对家族和部落的生存都是有益的,甚至是必需的——无疑他们确有这等品质,而且往往达到高度水平。
结 语
有一个学派认为道德是派生的(derivative school of morals),这一学派的哲学家们以前假定,道德的基础系建筑在利己之上的;但最近“最大幸福原则”(Greatest happiness principle)被突出地提出来了。 (290) 然而,把后一原则作为行为的标准,而不是作为行为的动机,是比较正确的说法。不过,我查阅过一些著作,所有这些作者们,除去少数例外, (291) 皆谓每一种行为一定都有一个特殊的动机,而且这个动机一定都同某种愉快或不愉快相关联。但是,人类的行为似乎常常出于冲动,这就是说,出于本能或长期的习性,却没有感到愉快的任何意识,其方式很可能恰如一只蜜蜂或一只蚁盲目地遵从其本能时所做的那样。在像火灾那样极端危险的情况下,当一个人毫无片刻踌躇、竭力去救他的同伙时,他简直不能感到什么愉快;而且他更没有时间去考虑如果他不这样干,以后可能会感到不满足。如果此后他回想起自己的行为,他大概会感到有一种冲动的力量存在于他自身之中,而这种力量同追求愉快或幸福大不相同;这似乎就是根深蒂固的社会本能。
在低于人类的动物场合中,把它们社会本能的发展说成是为了物种的一般幸福,莫如说是为了物种的一般利益更加恰当得多。我们可以给一般利益这个术语下这样一个定义,即:在它们所隶属的外界条件下,把最大数量的个体养育得充满活力和十分健壮,而且使其一切能力均臻完善。由于无论人类的或低于人类的动物的社会本能;都是以差不多一样的步骤发展的,所以在这两种场合中,采用同一个定义,并且以群体的一般利益或福利、而不以一般幸福作为道德的标准,如果行得通,还是适当的;但是,由于政治的伦理学的关系,对这个定义也许需要某种限制。
当一个人冒着生命危险去救一个同伙的生命时,我们说他的这种行为是为了人类的幸福,莫如说是为了人类的利益,似乎也是更为正确的。毫无疑问,个人的利益和个人的幸福通常是一致的;一个满足的、幸福的部落将比一个不满足的、不幸福的部落繁荣兴旺。我们已经知道,甚至在人类历史的早期阶段,群体的明确愿望将会自然地在很大程度上影响每一个成员的行为;因为所有成员都希望幸福,所以,“最大幸福原则”便成为最重要的第二位的指针和目的了;然而,社会本能以及同情心(它引导我们重视他人的褒贬)则为第一位的冲动和指针。这样,对于把我们本性最高尚的部分建筑在利己原理的基础之上所进行的指责就会被消除;诚然,除非每一种动物当遵从其固有本能时所感到的满足,以及当这种本能受到制止时所感到的不满足被称为利己,那就另当别论了。
同群诸成员最初由口头,其后由文字表示出来的愿望和意见,或者单独形成我们行为的指针,或者大大加强社会本能;然而,这等意见不时有直接反对社会本能的倾向。“荣誉律”(Law of Honour)对后述这一事实提供了很好的例证,这就是由地位相同的人的意见、而非由所有同胞们的意见形成的一项律条。违反这一律条,甚至当知道这种违反是同真实道德严格符合时,也会致使许多人感到比真正犯罪时更大的极度痛苦。我们在下述那样的感觉中可以辨认出同样的影响,即:如果偶然地违反了一种细小的,但是确定的礼节,当我们回忆及此时,即使事隔多年,大多数人还会有一种炽烈的羞愧感。从长远观点看,对所有成员来说什么是最好的,群体对此所做的评判一般要受到某种幼稚经验支配;但是,由于愚昧无知以及推理方的薄弱,这种评判陷于错误者并不罕见。因此,同人类的真正利益和幸福完全相反的最奇怪的风俗和迷信在全世界便成为威力无穷的了。在打破其社会等级的印度教徒所感到的恐怖以及许多其他这样的事例中,我们看到了上述这种情形。一个印度教徒被诱惑吃了不洁净的食物后所感到的悔恨同他犯了偷窃后所感到的悔恨有何不同,是难以区别的;不过前者很可能要更剧烈些。
我们不知道,如此众多的荒谬行为准则以及如此众多的荒谬宗教信仰是怎样发生的,我们也不知道,它们在世界各地怎么会如此深入人心;但值得一提的是,一种信仰如果在生命早期当脑筋易受影响时受到不断反复的灌输,那么这种信仰似乎就会获得一种差不多本能的性质;一种本能的本质就在于它的被遵从并不依靠理性。我们无法说,为什么某些可称赞的美德,如热爱诚实,在某些部落远比在另外一些部落受到更高的欣赏 (292) ;我们也无法说,甚至在文明民族之间也普遍有同样的差别。既然知道许多奇怪的风俗和迷信已经多么稳固地固定下来,那么我们对下面的情况就不必感到惊奇了,即受到理性支持的自重美德,虽然在人类早期状态下没有得到重视,但现今在我们看来它是如此自然,以致被认为是天生的。
尽管有许多疑惑根源,我们还是能够一般地而且容易地区别高级的和低级的道德准则。高级道德准则是建筑在社会本能之上的,而且同别人的福利有关。它们受到我们同伙称赞的以理性的支持。有些低级道德准则当含有自我牺牲的意思时,虽然不应称其为低级的,但它们主要同自我有关,而且系由舆论所引起,并由经验和教养使其成熟;因为野蛮部落不实行之。
当人类文明有所进步,并且小部落联合成较大的群体时,最简单的理性将告诉每一个人,他应该把他的社会本能和同情扩大到同一民族的一切成员,虽然在个人方面他们并不相识。这一点一旦达到之后,阻止其同情扩大于所有民族和所有种族的人,就只有一种人为的障碍了。诚然,如果这等人们由于容貌和习惯的巨大差异而被区分开,经验不幸地向我们阐明,在我们把他们视为同胞之前,不知要经过多么悠久的岁月。超越人类范围以外的同情,即对低于人类的动物施以人道,似乎还是最近获得的道德之一。未开化人除了对其玩赏动物外,显然没有这种感觉。古罗马人可恶的人兽格斗表演,阐明了他们对人道所懂得的是何等之少。就我所能看到的来说,潘帕大草原(Pampa) (293) 上的大多数高卓人(Gauchos) (294) 还不知道真正的人道概念。这是人类被赋予的最高尚美德之一,它似乎是我们的同情变得愈益亲切而且愈益广施的附产物,直到把同情扩大到一切有知觉的生物。这种美德一旦受到少数人的尊重并实行之,它就会通过教育和榜样传播于青年之间,最终便成为舆论的一部分。
道德修养的可能的最高阶段是,我们认识到应该控制自己的思想,“甚至在内心深处的思想中也不再去想过去使我们感到非常快活的那些罪恶”。 (295) 无论什么坏行为,只要为心理所熟悉,就容易实行得多。正如罗马皇帝奥瑞利亚斯说过的:“汝之习以为常之思想为何,汝之心理特性亦为何,盖灵魂被思想之色所染也。” (296)
英国大哲学家斯宾塞最近说明了他对道德观念的观点。他说:“我相信,通过人类种族一切过去世代所组织起来并且巩固下来的功利经验,已产生了相应的变异,这等变异由于连续的遗传和积累便成为我们道德直觉的一定能力——道德直觉乃是对正确行为和错误行为反应的一定情绪,而这等行为在个人功利经验方面,并没有明显的基础。” (297) 美德的倾向或多或少都是遗传的,在我看来,这并无固有的不可能性。因为,且不谈许多我们的家养动物将其各种性情和习性传递给后代,我曾听到一些可靠的事例表明,偷窃的欲望和说谎的倾向看来在一些上层家庭中也有所蔓延,因为偷窃在富有阶级中是一种罕见的犯罪,所以如果同一家庭的两三个成员都有这种倾向,简直就不能用偶然的巧合来加以解释了。如果坏倾向是遗传的,那么好倾向很可能也同样是遗传的。身体状态由于可以影响脑部,所以对道德倾向也会发生重大影响,大多数患有慢性胃病和肝病的人都明白这一点。“道德观念的堕落或毁灭往往是精神错乱的最早症状之一”, (298) 这也阐明了同样的事实,疯狂常常被遗传,乃是众所周知的。除非根据道德倾向的遗传原理,我们就无法理解据信存在于人类各个种族之间的这方面差异。
美德的倾向即使部分地遗传,也会对直接或间接来自社会本能的第一位冲动给予莫大帮助。只要承认美德倾向是遗传的话,那么似乎很可能是,至少在像贞洁、自我克制、对动物施行人道等那样的场合中,美德倾向通过在同一家族中连续若干代的习性、教育和榜样而最初印记在精神机构中;并且通过具有这等美德而在生存斗争中获得最大成功的个体,而最初印记在精神机构中,不过后者的程度是十分次要的,或者根本没有作用。关于任何这样的遗传,我的主要疑问是,无感觉的风俗、迷信和嗜好,如印度教徒对不洁净食物的恐惧,是否应该按照同一原理而传递下去。我还没有遇到过任何证据可以支持迷信的风俗和无感觉的习性之遗传,虽然实质上这比下述情况的可能性不见得更小,即:动物可以获得对某些食物种类的遗传的嗜好或对某些敌对者的遗传的恐惧。
总之,人类无疑就像低于人类的动物那样,为了群体利益而获得的社会本能,从最初起就会使他有某种帮助同伴的愿望,某种同情感;以及强迫他重视同伴们的褒与贬。这等冲动在很早时期就作为他的原始的是非准则。但是,由于人类智力逐渐进步,并且能够探知其行为的比较遥远的后果;由于他获得了充分的知识以抵制有害的风俗和迷信;由于他不仅重视其同胞们的利益,而且日益重视其幸福;由于有遵从有益的经验、教育和榜样的习性,他的同情变得愈益亲切而且广施于人,以至扩大到一切种族的人、低能儿、残废人以及社会上其他无用的人,最终扩大到低于人类的动物——所以他的道德标准步步升高。派生学派的道德学者们以及直观学派的学者们都承认,自从人类早期历史以来道德标准就升高了。 (299)
由于不时可以看到在低于人类的动物的各种本能之间进行着一种斗争,所以在人类的社会本能以及由此派生出来的美德和他的低级的、虽然暂时比较强烈的冲动和欲望之间也应该有一种斗争,就不足为奇了。正如高尔顿先生 (300) 所说的,人类是在相当近的时期内才脱离野蛮状态的,所以上述就愈益不足为奇了。当屈服于某种诱惑之后,我们就要感到不满足、羞愧、后悔或悔恨,这同其他强有力的本能或欲望没有得到满足或受到压抑时所引起的那种感觉是相似的。我们把对过去受到诱惑的薄弱印象同永久存在的社会本能进行比较,或者同幼年时期获得的而在一生中增强的、直到差不多像本能那样强烈的习性进行比较。如果在我们面前依然有这种诱惑,而我们不为所动,那是因为社会本能或某种风俗习惯当时占有优势,要不就是因为我们已经懂得社会本能或某种风俗习惯今后如与对受到诱惑的薄弱印象相比较,前者似乎更加强烈,而且违背它,就要招来痛苦。展望未来诸代,没有理由惧怕社会本能将会变弱,我们可以预料美德的习性将会变强,也许通过遗传而固定下来。在这种场合中,在我们高级冲动和低级冲动之间所进行的斗争将比较不剧烈,而且美德终将胜利。
以上两章提要
毫无疑问,最低等动物和最高等动物之间的心理差异是巨大的。一个类人猿如果能够不带偏见地观察他自己的情形,他大概会承认,虽然他能作出狡诈的计划去抢掠一个田园,虽然他能用石头去打仗或者砸开坚果,但把石头制成一种工具的思想却完全在其范围之外。他大概会承认,关于进行一系列形而上学的推理,或者解答一个数学题,或者对上帝的思考,或者对庄严的自然景色的赞美,他所能做的就更少了。然而,有些猿类很可能宣称,他们能够赞美而且的确赞美过其对象在结婚期间所表现的皮毛颜色之美。他们大概还会承认,虽然他们能用叫声使其他猿理解其某些知觉和比较简单的需要,但用一定声音去表达一定意思的概念,决不会通过他们的头脑。他们大概要坚决主张,他们乐于以许多方式去帮助同群的伙伴,为了伙伴不惜冒生命的危险,并且对孤儿给予照顾;但他们将被迫承认;对所有生物的无私之爱——人类的最高尚品质,却完全超出其理解力之外。
尽管人类和高等动物之间的心理差异是巨大的,然而这种差异只是程度上的,并非种类上的。我们已经看到,人类所自夸的感觉和直觉,各种情感和心理能力,如爱、记忆、注意、好奇、模仿、推理等等,在低于人类的动物中都处于一种萌芽状态,有时甚至处于一种十分发达的状态。这等情感和心理能力像我们在家狗和狼或豺的比较中所看到的那样,也能通过遗传而有某种进步。如果能够证明一般概念的形成,自我意识等那样的某些高等心理能力绝对为人类所特有(这似乎是极其可疑的),那么,这等属性很可能仅仅是其他高度进步的智能的附带结果,而智能的高度进步主要是一种完善语言连续使用的结果。新生的婴儿到什么年龄才会有抽象的能力或自我意识并且可以考虑到其本身的存在?我们还无法作出回答;关于上升到怎样的生物等级才能有上述心理能力,我们也同样无法作出回答。语言的半人为、半本能的状况仍然带有其逐渐进化的标志。那种对上帝的崇高信仰,并非人类普遍具有的;而对精灵作用的信仰都是其他心理能力所自然产生的结果。道德观念在人类和低于人类的动物之间,也许提供了一个最好的和最高级的界限;但是关于这个问题,我不必多说什么,因为晚近我曾力图阐明社会本能——人类道德构成的首要原则 (301) ——在活跃的智力以及习性的效果帮助下,自然会引出一项金科玉律:“汝等所欲人之施于己者,即应以此施于人”;而这正是建立在道德的基础之上的。
在下一章中我将略述人类的几种心理官能和道德官能逐渐进化所经过的可能步骤的方式。这种进化至少是可能的,无可否认的,因为我们日常在每一个婴儿身上都可以看到这等官能的发展;而且我们还可以从比低于人类的动物的心理官能还要低的完全白痴,到一个像牛顿那样的伟人追踪出一系列完整的心理等级。
第五章 智能和道德官能在原始时代和文明时代的发展
智力通过自然选择的进步——模仿的重要性——社会的官能和道德的官能——它们的发展限于同一部落的范围之内——自然选择对文明民族的影响——关于文明民族一度曾是野蛮民族的证据。
本章所讨论的这个问题是极其有趣的,但我处理的方法并不完善,而且是片断式的。华莱士先生在上述曾经提及的那篇可称赞的论文 (302) 中争辩说,人类自从局部地获得那些智能和道德官能以区别于低于人类的动物之后,他就很少可能通过自然选择或其他方法发生身体变异。这是因为人类能够通过他的心理官能“使一个不变的身体同正在变化着的世界保持和谐一致”。人类有巨大能力使其习性适应于新的生活条件。他发明武器、工具以及获得食物和保卫自己的各种策略。当他迁徙到比较寒冷的气候中时,他穿衣裳,建棚屋,而且生火;他用火烧煮非如此不能消化的食物。他用各种方式对他的同胞们进行帮助,并且预测未来的事变,甚至在远古时代,他就实行了某种分工。
另一方面,低于人类的动物必须在身体构造上发生变异,才能在大大变化了的生活条件下生存下去。它们必须变得更加强壮,或者获得更加有效的牙或爪,以抵御新的敌对者;要不它们就必须缩小,以逃避发觉和危险。当它们迁徙到比较寒冷的气候中时,它们的皮毛必须变厚,或者体质发生改变。它们如果不能这样变异,就要灭亡。
然而,正如华莱士先生所正确坚持的,关于人类的智能和道德官能,情况就大不相同了。这等官能是易于变异的;我们有各种理由可以相信,这等变异有遗传的倾向。因此,它们如果以往对原始人类及其类猿的祖先有高度重要性的话,那么它们大概就要通过自然选择而有所完善或进步。智能的高度重要性,不容置疑,因为人类在世界上之所以能够取得优越地位主要应归功于他的智能。我们知道,在最原始状态的社会中,那些最精明的、发明和使用最优良的武器和陷阱的并且能够最好地保卫自己的个人,将养育最大数量的后代。部落如果包含最大数量的赋有这等智能的人,这些部落的人数就要增加,而且会取代其他部落。人口数量首先决定于生活资料,而生活资料则部分地决定于一个地方的自然性质,但在非常大的程度上决定于那里所实行的技术。当一个部落增大了而且胜利了的时候,它往往通过同化其他部落而进一步增大。 (303) 一个部落的人们的身材和体力对于它的成功同样也有某种重要性,而身材和体力则部分地决定于他们所能得到的食物的性质和数量。在欧洲,青铜时代的人被一个更加强有力的种族所代替,根据他们的刀柄来判断,后者的双手是比较大的; (304) 不过他们的成功,更多地还是由于他们在技术方面的优越性。
所有我们知道的有关未开化人的情况,或者从他们的传说和古代碑石——其历史已完全为现代居民所遗忘——推论出来的情况,都阐明了自极其遥远的古代以来成功的部落就曾取代其他部落。在整个地球上的文明地方,在美洲的辽阔平原上,并且在太平洋的孤岛上,都曾发现过绝灭的或被遗忘的部落废墟。今天文明民族到处取代野蛮民族,除非那里的气候设置了致命的障碍,他们的成功主要是,纵使不完全是,通过他们的技术获得的,而技术则是智能的产物。因此高度可能的是,人类的智能主要是通过自然选择而逐渐达到完善的;这一结论就可以充分满足我们的意图了。当然,从低于人类的动物的智能状态到人类的智能状态追踪出各个独立智能的发展无疑是有趣味的,但我的能力和知识都不容我做这样的尝试。
值得注意的是,一旦人类的祖先成为社会性的(这很可能发生于很早的时期),模仿、理性以及经验的原则就会在某种程度上增大并大大改变其智力,现今我们还可以在低于人类的动物中看到这等智力的仅有痕迹。猿类像最低等的未开化人那样,很喜欢模仿;以前提到的一个简单事实表明,经过一段时间后,在同一地方用同一种类的陷阱就不能捉住任何动物,这阐明了动物会从经验中得到教训,而且可以模仿其他动物的谨慎。且说,如果在一个部落中,有某一个人比其他人更精明,发明一种捕捉动物的新圈套或一种新武器或其他攻守工具,那么,最明显的自身利益就会鼓舞其他成员去模仿他,而无须很大推理力的帮助;而所有成员都会因此受益。各种新技术的经常实践一定也在某种微小程度上可以使智力加强。如果新发明是一项重要的发明,这个部落的人口数量就会增加,广为散布,并取代其他部落。一个部落的人口如果因此而愈益增多,那么降生另外优秀的和富有发明才能的人,始终有更多的机会。如果这样的人留下来的孩子们继承了其心理上的优越性,那么降生越发机灵的成员的机会,大概多少要多些,而在一个很小的部落中决定的要多些。甚至他们没有留下孩子,部落依然包含有其血缘关系的亲属;农业学者们现已查明, (305) 当一头动物被屠宰后,如果发现它是有价值的,那么用这头动物的家系进行保存和繁育就可以获得我们所需要的性状。
现在转来谈谈社会的官能力和道德的官能。原始人类或人类的类猿祖先要成为社会性的,就必须获得那些迫使其他动物进行合群生活的同样本能情感;而且毫无疑问,他们显示了同样的一般倾向。当他们离开他们的伙伴时就会感到心神不安,他们对伙伴们大概会感到某种程度的爱;他们在遇到危险时将彼此发出警告,而且在进攻或防御中彼此进行帮助。所有这一切都意味着某种程度的同情、忠诚和勇气。这等社会属性对低于人类的动物的高度重要性已是无可争辩的了,毫无疑问,人类祖先也是以相似的方式,即在遗传的习性帮助下通过自然选择获得这等属性的。当生活在同一地方的两个原始人类的部落进行竞争时,如果(其他条件相等)某一个部落包含有大量勇敢的、富有同情心的并且忠实的成员,他们时刻准备彼此发出危险警告,相互帮助,相互防卫,那么这个部落就要获得较大的成功而征服其他部落。让我们记住,在未开化人的永无休止的战争中,忠诚和勇气是多么重要。受过训练的军人之所以优于没有受过训练的乌合之众,主要在于每个人对其同伙所感到的信赖。正如巴奇霍特 (306) 所很好阐明的,服从具有最高的价值,因为任何形式的政府都比没有政府好。自私的和好争论的人们不会团结一致,而没有团结一致,什么也不能完成。一个部落如果富有上述那些属性,就会广为分布,战胜其他部落:但是,根据过去的历史来判断,经过一定的时间,这个部落又会被另一个禀赋更高的部落所征服。这样,社会的和道德的属性就倾向于徐徐进步,而普及于全世界。
但可以这样问:大量成员在同一部落的范围内最初怎样赋有这等社会的属性和道德的属性呢?美德的标准又是怎样提高的呢?比较富有同情心的和仁慈的双亲所生育的后代,或者对其伙伴比较忠诚的双亲所生育的后代,其数量是否会比同一部落的自私而奸诈的双亲所生育的后代更多,是极其可疑的。一个人宁愿牺牲自己的生命,就像许多未开化人所做的那样,也不背叛他的伙伴,他大概常常不会留下后代以继承其高尚本性的。最勇敢的人们在战争中永远心甘情愿奔向前方,而且慷慨地为他人献出自己的生命;这样的人平均要比其他人死的多。因此,赋有这等美德的人们的数量或他们的美德标准不能通过自然选择,即最适者生存。而被提高,似乎是很可能的;因为我们在这里所谈的并非是某一部落战胜另一部落的问题。
导致赋有这等美德的人们在同一部落内增加其数量的情况虽然过于复杂,而无法清楚地把它探究到底,但我们还能够追踪出某些可能的步骤。首先,当部落成员的推理力和预见力有所进步时,每一个人很快就会懂得,如果他帮助同伙,通常也会得到作为回报的帮助。从这个低等动机出发他大概可以获得帮助其同伙的习性;行使仁慈行为的习性肯定要加强同情感,而对仁慈行为的最初冲动则是同情感给予的。加之,在许多世代中被遵从的习性很可能有遗传的倾向。
但是对社会美德发展的另一个更加强有力的刺激则是由我们同伙的褒贬所提供的。正如我们已经看到的,我们经常对他人加以赞扬或给予谴责主要是由于同情本能,如果这是施于我们自己,我们当然爱赞扬而怕谴责;这种本能无疑像所有其他社会本能那样,最初也是通过自然选择而获得的。在多么早的一个时期,人类祖先在其发展进程中变得能够感觉到其同伙的赞扬和谴责,并被它们所激励,我们当然无法说出。不过,甚至狗似乎也懂得鼓励、赞扬和谴责。最原始的未开化人也有光荣感,如他们保存那些英勇获得的战利品,他们有过分自夸的习性,他们甚至极端注意其个人容貌和装饰,这就明确地阐明了上述感觉,因为,除非他们重视其伙伴们的意见,否则这等习性就没有什么意义了。
如果违反他们的某些次要准则,他们肯定也要感到羞愧,而且显然要感到悔恨,例如,那个澳洲土人由于没有能够及时谋杀另一个妇女以安慰其亡妻之灵而日益憔悴和心神不安,就是一个说明。我虽然没有遇到过任何其他见于记载的事例,但下述事例足以说明一个未开化人宁愿牺牲自己的生命而不背叛他的部落,宁愿坐牢也不违反他的誓言, (307) 像这样的人当没有完成他视为神圣的义务时,而不在灵魂深处感到悔恨,简直是令人不可相信的。
因此我们可以断言,在很遥远的古代,原始人类已经受到了其同伴赞扬和谴责的影响。显然,同一部落的成员对那些在他们看来具有普遍利益的行为将会表示赞成,而对那些看来是有害的行为则会予以谴责。为他人谋利益——汝如何施于人,人亦将如何施于汝——乃是道德的基础。因此,关于原始时代中爱赞扬,怕谴责的重要性,我们简直无法把其重要性再予以夸大了。一个人如果为了他人的利益而牺牲自己的生命,并非被任何深刻的本能情感所推动,而是被一种荣誉感所激起,那么他就会以他的榜样唤起其他人要求荣誉的愿望,而且还会以实行这种行为来加强对其称赞的高尚情感。这样,他给部落带来的好处远比他留下一些倾向于承继其自己那样高尚品格的后代还要多得多。
人类的经验和理性增长了,就可以察觉出其行为的更加遥远的后果;而自重的美德,如自我克制、贞洁等,即将受到高度的尊重,甚至被视为神圣的,可是这等美德,像我们以前看到的那样,在早期却完全不受重视。然而,我没有必要再重复我在第四章中关于这个问题的叙述。我们的道德观念或良心终于成为一种高度复杂的思想感情——它起源于社会本能,大大被我们同胞们的称赞所指导,还受到理性和自我利益而且晚近又受到深厚的宗教情感的支配,更被教育和习性所巩固。
一定不要忘记,对任何人及其子孙胜过同部落的其他人来说,道德的高标准虽然仅有很少一点利益,或者根本没有利益,但禀赋优良的人在数量上的增加以及道德标准的进步,对某一个部落胜过另一个部落来说,肯定有巨大的利益。一个部落如果包含有许多这样的成员:他们由于高度具有爱国精神、忠诚、服从、勇敢以及同情而永远彼此相助,并为公共利益不惜牺牲自己,那么这个部落就会战胜大多数其他部落;这大概就是自然选择。某些部落取代了其他部落,遍及全世界,无论何时都是如此;因为道德是他们成功的一个重要因素,所以道德标准和禀赋优良的人们的数量这样就会到处有提高和增加的倾向。
为什么某一个特殊的部落,而不是另一个部落获得成功并且在文化等级上有所提高呢,对此很难形成任何判断。许多未开化人现今所处的状态同几世纪前他们最初被发现时的状态没有两样。正如巴戈霍特先生所说的,我们容易把人类社会的进步视为正常之事;但历史反驳了这一点。古代人甚至没有这种进步观念,东方民族迄今还是如此。按照另一位大权威亨利·梅因爵士 (308) 的说法,“人类的大部分对其文明制度的改进从来没有显示过一点愿望”。进步似乎决定于许多同时发生的有利条件,不过这太复杂了,以致无法查明其究竟。不过常常这样说,凉爽的气候可以导致勤奋和许多技术的发生,所以这曾是高度有利的。爱斯基摩人为艰难的需要所迫,虽成功地完成了许多精巧的发明,但他们的气候太严酷了,以致不能继续进步。游牧生活的习性,无论是在辽阔的平原上,还是穿过热带的密林,或是沿着海岸,都是高度有害的。当我对火地的野蛮居民进行观察时,给我留下深刻印象的是,拥有某种财产,一个固定的住所,许多家庭在一个首领下的联合,都是文明所不可缺少的必要条件。这等习性几乎需要土地耕作;正如我在别处所阐明的, (309) 耕作的第一步很可能是这样一种偶然事件的结果,即一棵果树的种子偶然落在垃圾堆上,然后产生了一个异常优良的变种。然而未开化人最初如何向着文明进步的问题迄今还是非常难以解决的。
自然选择对文明民族的影响
迄今为止,我仅考虑了人类从半人类状态进步到近代未开化人状态。关于自然选择对文明民族的作用还值得再谈一谈。格雷格(W.R.Greg)先生 (310) 对这个问题进行了富有才华的讨论,以前华莱士先生和高尔顿 (311) 先生也讨论过这个问题。我的论述均来自这三位作者。关于未开化人,无论身体或精神,只要衰弱,很快就会被淘汰;凡生存者普遍都显示了精力充沛的健壮状态。另一方面,我们文明人竭尽全力以抑制这种淘汰作用;我们建造救济院来收容低能儿、残废者以及病人;我们制定恤贫法令(poor-1aws);我们的医务人员以其医术尽最大努力去挽救每一个人的生命直到最后一刻。我们有理由相信,种痘保存了成千上万人的生命,而以前由于体质虚弱死于天花者真是成千上万。这样,文明社会的衰弱成员也可繁殖其种类。凡是注意过家养动物繁育的人不会怀疑这对人类种族一定是高度有害的。缺少注意或管理错误导致家养族退化之迅速,足以惊人;除非在人类本身的场合中,谁也不会愚蠢到允许他的最坏的动物去繁育。
我们感到被迫给予不能自助的人们以帮助,乃是来自同情本能的附带结果,同情本能最初是作为社会本能的一部分而获得的,但如以上所指出的,其后却变得愈益亲切而推及愈广。即使在坚强的理性迫使下,如果我们本性的最高尚部分没有堕落,我们也无法抑制我们的同情。外科医生当施行手术时可能无动于衷,因为他知道他所做的是为了病人好;但是,如果我们故意忽视弱者和不能自助的人,这只能是为了毫无把握的利益,而给现在带来的弊害却是无穷的。因此,我们必须承担弱者生存并繁殖其种类的毫无疑义的恶劣后果;但是,似乎至少有一种抑制作用在稳定地进行着,即:社会的衰弱成员和低劣成员不会像强健成员那样自由地结婚;由于身体或心理衰弱的人不能结婚,这种抑制作用可能无限地增强,虽然这只是可望而不可求的事。
在保持一支大规模常备军的每一个国家里,最优秀的青年都要被招募或被征集。这样,在战争期间就有早死之虞,而且常常被诱人腐化堕落之途:在青春时代不能结婚。另一方面,体质不良的比较矮小而衰弱的人们却留在家中,因而结婚以及繁殖其种类的机会就要好得多。 (312)
人积聚财产,并把它传给孩子,因此富家子弟在成功的竞争中,就比贫家子弟占有优势,而这同身体和智力的优越性却无关。另一方面,短寿的父母,其健康和精力平均都差,他们的孩子却比另外的孩子继承财产较早,而且结婚很可能较早,于是留下的遗传其低劣体质的后代数量也较多。但是财产继承本身远非一种坏事;因为没有资本的积累,技术就不能进步;文明种族主要是通过技术的力量扩大了而且今天到处扩大着它们的范围,以取代比较低劣的种族。财富的适度积累并不妨碍自然选择的进程。当一个穷人有了中等财产的时候,他的孩子们就会进入竞争相当剧烈的商业或其他职业,所以身体和心理都健壮的人可得到最大的成功。有一批受到良好教育的人,不必为每日的面包去劳动,是非常重要的,对其重要程度给予怎样估量也不会过分;因为所有高等智力工作都是由他们进行的,而所有种类的物质进步主要都是决定于这种工作,更不要不谈其更高级的利益了。无疑地当财富过多时,就倾向于把人们变成无用的寄生虫;这里就会发生某种程度的淘汰,因为我们天天看到那些愚蠢的或生活放荡的富人把财产挥霍精光。
长子财产继承权是一种更加直接的弊害,虽然它以前对形成一个统治阶级可能有巨大好处,因为任何政府都比没有政府好。大多数长子虽然身体或心理可能都衰弱,却可以结婚,而幼子即使其身体或心理都优越,一般也不能结婚。况且承继遗产的长子即使无能,也不会把财产挥霍精光。但这里和别处一样,文明生活的亲戚关系是如此复杂,以致有某种补偿的抑制作用介入其中。富人通过长子继承便可以逐代选娶比较美丽而媚人的妇女,而这等妇女一般必定是身体健康和心理灵敏的。连续保存同一血统而不经过任何选择所应有的恶劣后果,为贵族永远希图增加其财富和权力所抑制;他们是以娶女继承人来实现这一愿望的。如高尔顿先生所阐明的, (313) 只生单性小孩的父母的女儿,其本身有不生育的倾向;这样,贵族家庭的直系就要经常被切断,而他们的财富流入旁支;不幸的是,旁支并不是以任何种类的优越性来决定的。
这样,虽然文明在许多方面对自然选择的作用有所抑制,但自然选择显然还是偏袒那些靠着良好食物和没有偶然困苦而身体发育较好的人。从下述情况可以推论这一点,即:在任何地方都可以发现文明人的身体比未开化人的身体强壮。 (314) 他们的耐力似乎也相等,这在许多次探险考察中已得到了证明。甚至富人的穷奢极欲也没有多大害处;因为英国贵族男女在一切年龄范围内的估计寿命比低等阶级的健壮英国人的寿命短不了多少。 (315)
我们现在来看看智能。在社会的每一个阶级中,如果把其成员分为相等的两群,一群的成员智能优越,一群的成员智能低劣,几乎无可怀疑的是,前者在所有职业中都能获得较大的成功,并且生育较大数量的孩子。即使在最低等的阶层中,有技艺和有才智的人一定也占有某种优势;但许多行业已经实行很细的分工,这一优势并不很大。因此,在文明民族中无论智能的数量或标准都有增加的倾向。但是,我不愿断言这种倾向不会在其他方面受到抵消而有余,如挥霍乱用和不顾将来所起的抵消作用即是;即使如此,有才智的人还会占有某种优势。
上述那样的观点常常遭到反对,即:历来最卓越的人士都没有留下遗传其伟大才智的后代。高尔顿先生说道:“我遗憾,我不能解决一个简单的问题:具有非凡天才的男人或女人是否不生育,并且不生育到怎样程度。然而,我曾阐明卓越的人士决非不生育。” (316) 伟大的制定法典者、仁慈的宗教奠基者、伟大的哲学家和科学发明家以他们的工作对人类进步所给予的帮助,其程度远比留下为数众多的后代要高得多。就身体构造来说,禀赋稍好的个体的被选择以及禀赋稍差的个体的被淘汰,并不是强烈显著而罕见的畸形的被保存,就会导致一个物种的进步。 (317) 关于智能,也是如此。因为在每一个社会阶层中,才智多少高些的人就比才智差些的人能够获得较大的成功,因而在其他方面如果没有受到抑制就可增加其数量。在任何民族中,当智力的标准以及智力优越的人士的数量提高了的时候,正如高尔顿先生所阐明的,根据平均离差的法则我们可以预料,非凡的天才将比以前似乎多少要更加常常出现。
关于道德属性,对于最恶劣性情的淘汰一直在进行着,即使在最文明的民族中也是如此。犯罪者被处死或长期监禁,所以他们不能自由地传递其恶劣属性。忧郁病患者和精神病患者受到隔离或自杀。凶暴的人和好争吵的人难免流血的结局。不安静的人不会从事任何固定的职业——这种野蛮状态的遗风是文明的最大障碍 (318) ——而他们迁移到新殖民地,却证明是有用的拓荒者。酗酒是高度有害的,例如,酗酒者从30岁算起,其估计寿命仅为13.8年;而英国农工从同一年龄算起,其估计寿命则为40.59年。 (319) 荒淫的女人生孩子很少,荒淫的男人则很少结婚;二者都因此得病。在家养动物的繁育中,淘汰那些有任何低劣性质的个体,即使为数不多,在走向成功方面也绝不是一个不重要的因素。关于那些通过返祖有重现倾向的有害性状尤其如此,如绵羊的重现黑色即是;关于人类,某些最恶劣的性情,没有任何可指出的原因,间或出现于一些家族中,这也许是归返一种野蛮状态,而这等野蛮状态正是在我们很多世代中没有被消除掉的。不错,用普通语言来说,这种观点似乎承认了那些人就是家族中的黑色绵羊。
关于文明民族,就道德的先进标准以及优秀人士的数量增加而言,自然选择所起的作用显然是不大的,虽然说基本的社会本能最初是通过自然选择而获得的。但是,当我讨论较低等种族时,对于导致道德进步的一些原因已经作了足够的叙述,这些原因就是:我们同胞所给予的称赞——我们的同情通过习性得到加强——榜样和模仿——理性——经验,甚至自我利益——幼年时代的教育以及宗教感情。
格雷格先生和高尔顿先生 (320) 曾强烈主张,在文明国家中,对于优秀阶级人士数量的增加有一个重要的障碍,那就是,很贫穷的人和不顾一切而乱来的人往往因恶行而堕落,他们几乎一定早结婚,而谨慎的、俭朴的人一般在其他方面也是有道德的,他们结婚都晚,所以能够维持自己和孩子们的舒适生活。早婚的人在一定时期内不仅产生的世代数较多,而且如邓肯(Duncan) (321) 博士所阐明的,他们生的孩子也较多。再者,母亲在壮年时期生的孩子比在其他时期生的孩子要重些和大些,所以很可能精力也充沛些。这样,社会上那些不顾一切乱来的、堕落的而且往往是邪恶的人比节俭的而且一般是有道德的人,其增加速度要快些。或者,像格雷格先生所说的那种情形:“满不在乎的、肮脏的、不求上进的爱尔人增殖的像兔子那样快;俭朴的、有远见的、自尊的、有雄心壮志的苏格兰人,其道德是严格的,其信仰是高尚的,其智力是精明的而且是训练有素的,却在斗争和独身生活之中度过其风华正茂的岁月,他们结婚晚,留下的子女很少。设有一地,最初居住着1000个撒克逊人(Saxons) (322) 和1000个凯尔特人(Celts) (323) ——经过12代以后,人口的5/6将为凯尔特人,而5/6的产业、权力以及才智则属于存留下来的1/6撒克逊人。在永恒的‘生存斗争中,低劣的和天赋较差的种族曾占有优势——他们占有优势并不是凭借其优良品质,而是凭借其缺点’。”
然而对于这种向下的倾向,则有某些抑制之道。我们已经看到,酗酒者的死亡率高、过度荒淫者留下的后代很少。最贫穷的阶级涌入城镇,斯塔克(Stark)博士根据苏格兰的10年统计, (324) 证明了城镇的死亡率在所有年龄中都比农村的高,“在生活的最初五年期间,城镇的死亡率差不多正好是农村的两倍”。由于这些统计既包括富人也包括穷人,所以要保持城镇赤贫居民和农村居民的人口比例不动,其降生的数量无疑需要提高两倍以上。对妇女来说,如果结婚太早,那是高度有害的;因为在法国发现“20岁以下,已婚妇女的死亡率为未婚妇女的两倍”。20岁以下的已婚男子的死亡率也是“非常高”的, (325) 但其原因是什么,似乎还无法确定。最后,如果男子在能建立一个舒适家庭之前,谨慎地推迟结婚,那么,像他们常常做的那样,将会选择壮年的妇女,这样,优等阶级人口增长率的减少只是微乎其微而已。
根据1853年所做的大量统计,证明全法国年龄在20~80岁之间的未婚男子比已婚男子的死亡率高得多,例如:每一千个年龄在20~30岁之间的未婚男子中,每年死亡者为11.3人,而已婚男子死亡者仅为6.5人。 (326) 相似的规律被证明也适用于1863和1864年苏格兰20岁以上的男子人口普查,例如:每一千个年龄在20~30岁之间的未婚男子中,每年死亡者为14.97人,而已婚男子死亡者仅为7.24人,这就是说,比一半还少。 (327) 斯塔克博士关于这一点说道,“独身比最有害健康的行业或者比居住在最有害健康的房屋或地方——那里对改善环境卫生从来没有过最长远的打算——对生活更加有害”。他认为死亡率的降低乃是“结婚以及比较有规律的家庭生活习惯”的直接结果。然而他承认酗酒、荒淫以及犯罪的人,寿命不长,普遍都不结婚;还必须承认,体质衰弱的、健康不良的、身体或心理有任何重病的人们往往都不愿结婚,或者人家拒绝同他们结婚。斯塔克博士似乎得出这样一个结论,即结婚本身为延长益寿的一个主要原因,因为他发现已婚老人在这两点上仍然胜过同样高龄的未婚者;但每个人一定都知道有些人的事例;他们在幼年时期不健康,没有结婚;虽然他们终生衰弱因而寿命或结婚的机会一直在缩小,但仍然活到高龄。还有另一个值得注意的情况似乎可以支持斯塔克博士的结论,即:在法国,寡妇和鳏夫同已婚者相比,前者的死亡率要高得多;不过法尔(Farr)博士把这种情形归因于由家庭破坏而引起的贫穷和恶习,并且归因于遭到不幸后的悲痛。总之,我们同意法尔的说法,可以作出这样的结论:已婚者比未婚者的死亡率低,似乎是一般的法则,这“主要是由于对不完善类型的经常淘汰,以及对最优秀个体在连续世代中的巧妙选择”;这仅仅是和婚姻情况有关的选择,而且这种选择对身体的、智力的以及道德的所有属性都发生作用。 (328) 因此,我们可以推论,健康的和善良的人们出于谨慎而暂时不结婚,其死亡率也不会高。
上述两节所举的各种抑制因素,也许还有其他抑制因素,如果不能制止社会上那些不顾一切乱来的、邪恶的以及其他方面低劣的分子的增长速度快于优等阶层的人们,那么这个民族就要退化,这在世界历史中已屡见不鲜了。我们必须记住,进步并非是永恒不变的规律。为什么某一个文明民族兴起了,比另一个民族更强大,而且分布得更广;或者,为什么同一个民族在某一个时期比在另一个时期进步较快,对此很难有所说明。我们只能说,这是决定于人口实际数量的增加,决定于赋有高度智能和道德官能的人们的数量,同时还决定于他们的美德标准。身体构造似乎也有一点小影响,不过只是在旺盛的身体活力导致旺盛的心理活力的情况下才如此。
有几位作者极力主张,高度的智力既有利于一个民族,如果自然选择的力量是真实的话, (329) 那么在智力方面高出于曾经存在的任何种族的古希腊人就应该愈益提高其智力,增加其人口数量,而遍布于整个欧洲。这里有一个不言而喻的假设,这是常常对身体构造作出的,即:心理和身体的连续发展有某种内在的倾向。但是,所有种类的发展都决定于许多共存的有利环境条件。自然选择的作用只是试探性的。个人或种族可能获得了某些无可争辩的优势,然而由于其他特性不好,也不免于灭亡。古希腊人之所以衰退,可能由于许多小邦之间缺少团结,可能由于整个国土不大,可能由于实行奴隶制,也可能由于极度耽于声色口腹之乐;因为直到“他们削弱和腐败到极点” (330) 然后才败亡。现今欧洲西部民族超越其以往野蛮祖先的程度是不可估量的,他们站在文明的顶峰,虽然他们受惠于古希腊人的著作至多,但其优越性来自这个非凡民族的直接遗传都很少,或者全无。
谁能肯定地说出一度如此占有优势的西班牙民族为什么在竞争中被远远甩在后面了。自从中世纪黑暗时代以来,欧洲诸民族的觉醒是一个更加错综复杂的问题。正如高尔顿先生所说的,在古代那一时期,几乎所有本性高尚的人,要想沉思冥想或进行精神修养,除了投入必须严守独身生活的教会的之外, (331) 简直没有任何隐身之所,这几乎不可避免地要对相继的各代发生退化的影响。在这同一时期,宗教法庭极意搜捕思想最自由和行动最勇敢的人们,把他们烧死或囚禁起来。仅在西班牙,最优秀的人士——他们遇事持怀疑态度并且提出问题,而没有怀疑就不能有进步——在3个世纪内每年被消灭的数以千计。尽管如此,欧洲还是以无比的速度前进了。
同其他欧洲民族相比,英国人在殖民方面获得了惊人的成功,这曾被归因于他们的“果敢和不挠的精力”;把英国血统的加拿大人和法国血统的加拿大人的进步做一比较,就会很好地说明其结果;但是,谁能说出英国人是怎样得到其精力的呢?有人相信美国的惊人进步及其人民的特性乃是自然选择的结果,这是非常正确的。因为,精力较强的,勤劳勇敢的人们在最近10~12代期间从欧洲各地迁移到这片大陆,而且在那里获得了最大的成功。 (332) 从遥远的未来来看,我并不认为津克(Zincke)以下的观点是夸大的,他说, (333) “所有其他一系列事件——如希腊精神文明所产生的事件和罗马帝国所产生的事件——只有同盎格鲁撒克逊人的巨大西移潮流这一事件相联系,毋宁说作为它的次要事件来看,似乎才有意义和价值。”文化进步的问题固然还是模糊不清,但我们至少能够看出,一个民族如果在长年累月中不断产生最大数量的高智力的、精力旺盛的、勇敢的、爱国的以及仁慈的人,一般就会比天赋较差的民族占有较大的优势。
自然选择来自生存斗争;而生存斗争则来自人口的迅速增加。对于人类的增加速度,我们不能不痛苦地感到遗憾,这是否明智,则是另一个问题;因为,这在野蛮部落中导致杀婴以及许多其他弊害,在文明民族中导致赤贫、独身以及谨慎小心的人们实行晚婚。但是,由于人类蒙受到的身体弊害同低于人类的动物一样,所以他没有权力期望去避免由生存斗争所引起的弊害。如果人类在原始时代未曾受自然选择所支配,那么他决不会达到现在这样的地位。因为我们在世界许多部分看到还有土壤最肥沃的广阔区域能够维持大量的快乐家庭,但只有少数游牧的未开化人生活于其间,因此,可以这样辩说,生存斗争并没有达到足够的剧烈程度以迫使人类向上发展到最高的标准。根据我们所知道的人类以及低于人类的动物的全部情况来判断,他们的智能和道德官能对于通过自然选择而发生的稳定进步永远有足够的变异性。毫无疑问,这种进步需要许多共存的有利环境条件;不过,如果没有人口的迅速增加以及由此引起的极其剧烈的生存斗争,最有利的环境条件是否会发生足够的作用,还是完全可以怀疑的。例如,根据我们在南美一些地方所看到的情况来说,甚至一种可以称为文明的民族,如西班牙殖民者,看来当生活条件很安逸的时候,就容易变得懒惰而致倒退。关于高度文明的民族,其不断进步在次要程度上还决定于自然选择;因为,这等民族并不像野蛮部落那样,彼此取代而被消灭之。尽管如此,从长远观点来看,同一群体内智力较高的成员比智力较低的成员将会获得较大的成功,留下较多的后代,这就是自然选择的一种形式。进步更加有效的原因似乎在于:当幼年期间头脑易受影响时施以良好教育,由最有才能和最优秀的人士反复灌输高标准的美德,体现民族的法律、风俗和传统,并且由舆论进行强制。然而,应该记住,舆论的强制性决定于我们能够鉴别他人的称赞和谴责;这种鉴别是以我们的同情为基础的,而同情作为社会本能的一个最重要因素最初通过自然选择而得到发展,简直是无可怀疑的。 (334)
关于所有民族一度曾为野蛮民族的证据
这个问题已由卢伯克爵士 (335) 、泰勒先生、伦南先生等人进行了充分的和可称赞的讨论,我在这里只是叙述一下他们所得结果的最简短提要而已。最近阿盖尔公爵 (336) 提出的和以前沃特利(Whately)大主教提出的论点支持了这样一种信念:认为人类本来是作为一种文明者进入这个世界的,所有野蛮人是由于此后发生了退化,在我看来,这种论点同另一方所提出的论点相比似乎就显得虚弱了。许多民族无疑都曾与文明背道而驰,有些可能堕入完全野蛮的状态,虽然我还没有遇到过关于后面这一点的证据。火地人大概为其他胜利的游牧民族所迫,定居在现今那块荒凉的地方,结果他们可能变得有点更加退化了;但很难证明他们已经降到博托克多人(Botocudos)以下,而博托克多人却是在巴西的最好地方居住的。
所有文明民族都是野蛮人的后裔,其证据在于:一方面,在现今依然存在的风俗、信仰、语言等等之中,还有他们以往低等状态的明显痕迹;另一方面,已证明未开化人能够独立地在文明等级上提高少数几步,而且他们确曾这样提高过。有关第一方面的证据是极其奇妙的,我还不能在这里举出:我谈到的这等例子是关于计数技术的,正如泰勒所明确阐述的,这同现今在某些地方依然使用的字有关,计数发源于手算,最初用一只手,然后用两只手,最后连脚趾也用上了。在我们自己使用的十进制以及在罗马数字上都有这种痕迹,罗马数字的V料想为一只人手的简形,在V之后为Ⅵ等等,当时无疑两只手都用上了。再者,“当我们说三个20加10时,我们是用二十进制计算的,每个20在概念上代表一个整人,如墨西哥人或加勒比人(Carib)所云”。 (337) 按照一个日益扩大的学派的语言学者们的意见,每一种语言都有其缓慢而逐渐进化的痕迹。书法亦复如此,因为字母就是图形代表的痕迹。凡是读过伦南的著作 (338) 的人,简直不能不承认几乎所有文明民族至今仍然保持着用暴力抢婚那样的粗野习俗。同一位作者问道,能够举出什么古代民族原本就实行一夫一妻制吗?正义的原始概念,如仍然保留其痕迹的战争法以及其他风俗所阐明的,同样也是最粗野的。许多现存的迷信正是以往虚假宗教信仰的残余。宗教的最高形态——上帝憎罪恶而爱正义的崇高概念——在原始时代是不知道的。
转来谈谈另一类证据:卢伯克爵士曾阐明,最近有些未开化人在某些技艺方面稍有进步。他所做的非常奇妙的叙述表明,世界各地未开化人使用的武器、工具以及技艺差不多都是独立发明的,也许取火的技术除外。 (339) 澳洲土人的飞镖(boomerang) (340) 是这种独立发明的一个良好事例。塔希提人(Tahitians) (341) 当最初被发现时,在许多方面就比其他波利尼西亚诸岛上大多数居民进步。关于秘鲁士著居民和墨西哥土著居民的高度文化是由国外传来的信念 (342) ,并没有充分的根据,那里栽培着许多土著植物并饲养少数土著动物。从大多数传教士所发生的影响很小来判断,我们应该记住,来自某一半文明地方的一群漂流者如果被冲到美洲海岸,若非当地土著居民已经多少有点进步的话,这群漂流者对他们是不会发生任何显著影响的。看看世界历史的远古时代,用卢伯克爵士的著名术语来说,我们就可以发现一个旧石器时代和新石器时代,没有人会妄称磨制粗陋燧石器的技术是从外边传来的。在欧洲的所有地方,一直东到希腊,在巴勒斯坦、印度、日本、新西兰以及包括埃及在内的非洲,都曾发现过大量的燧石器;而现今居民都没有保持使用它们的任何传统。关于中国人和古代犹太人以前都使用过石器,也有间接的证据。因此,差不多包括全部文明世界的这等地方的居民一度都处于野蛮状态,这简直是无可怀疑的了。认为人类原本是文明的,其后在许多区域发生了完全退化的那种信念,乃是可怜而又可鄙地低看了人类的本性。而认为进步远比退步更加普遍,并且认为人类虽然经过缓慢而中断过的步骤却由低等状态上升到今天那样的知识、道德和宗教的最高标准,显然是一种更加真实、更加令人振奋的观点。
第六章 人类的亲缘和系谱
人类在动物系列中的位置——系谱的自然分类法——价值微小的适应性状——人类同四手类在各个微细之点上的类似——人类在自然分类中的等级——人类的发源地及其古老性——作为连接环节的化石的缺少——第一根据人类亲缘、第二根据人类构造推论出来的人类系谱较低诸阶段——脊椎动物的早期雌雄同体状态——结论
纵使承认人类同其关系最近的同源动物之间在身体构造方面的差异大到像某些自然学者们所主张的那样,而且纵使我们必须承认他们之间在心理能力方面的差异也是巨大的,但上述各章所列举的事实看来还以最明显的方式表明了人类是从某一较低等类型传下来的;尽管连接的环节迄今尚未被发现,亦复如此。
人类容易发生众多的、微小的和各式各样的变异,这等变异就像在低于人类的动物中那样,是由同样的一般原因所引起的,并且受到同样的一般规律的支配而遗传下去。人类增殖得如此之快,以致他必然要处于生存斗争之中,因而要受到自然选择。人类产生了许多种族,其中有些种族彼此差异如此之大,以致他们常常被自然学者们列为不同的种(species)。他的身体是按照同其他哺乳动物一样的同源图案构成的。他通过同样的胚胎发育阶段,他保持着许多残迹的和无用的构造,毫无疑问,这些构造以前一度是有用的。性状不时在其身上重现,我们有理由相信他的早期祖先曾经具有这等性状。如果人类的起源完全不同于一切其他动物,则上述种种表现只能是一种空洞的欺骗;但承认这一点乃是令人难以相信的。相反,如果人类和其他哺乳动物都是某一未知的、较低等类型的共同后裔,则上述表现就是可以理解的了,至少在很大程度上是可以理解的。
有些博物学者们由于对人类的心理和精神动力有深刻的印象,所以把整个有机界分为三个领域,即:人类、动物界、植物界,这样就把人类立为单独的一界(kingdom) (343) 。博物学者无法对精神能力进行比较或加以分类:但他可以像我曾经做的那样,尽力阐明人类和低于人类的动物的心理官能虽在程度上有巨大差异,但在种类上并无不同。一种差异的程度不论多么大,也不能证明我们把人类列为独特的一界是正当的,把两种昆虫,即无疑属于同纲(class)的胭脂虫(coccus)和蚂蚁的心理能力加以比较,也许会对这一点作出最好的说明。在这里,二者心理能力的差异大于人类和最高等哺乳动物之间心理能力的差异,虽然其种类多少有点不同。雌胭脂虫当幼小时用喙附着在一种植物上,吸其液汁,此后决不再移动;于是受精产卵;这就是它的全部生活史。另一方面,描述工蚁的习性及其心理能力,像胡伯尔所做的那样,则需要巨卷著作;但我将简略地列举少数几点。蚁类肯定会彼此互通消息,若干蚁联合起来进行同一项工作,或者在一起游戏。分离数月之后,还能认识它们的同群伙伴,而且彼此会感到同情。它们建筑大厦,保持清洁,晚间关闭门户,并设警卫。它们修筑道路以及在河床下面修筑隧道,架设临时桥梁以连接在一起。它们为群体聚集食物,如运回窝中的东西太大而不能进门时,它们就把门开大,然后修复原状。它们贮存子实,防止它们发芽。如果受潮,就把子实运到地面上进行干燥。它们畜养蚜虫和其他昆虫作为奶牛。它们以整齐的队列出发征战,并且为了公共福利从容地牺牲自己的生命;它们按照事先预定的计划进行迁徙;它们俘获奴隶。它们把蚜虫的卵和自己的卵、茧运到窝中暖和的部分,以便它们尽快孵化;还可以举出无数相似的事实。 (344) 总之,蚂蚁和胭脂虫之间在心理动力方面的差异是巨大的;但从来没有人梦想过把这两种昆虫放人不同的纲,更不用说放人高得多的不同的界了。这两种昆虫之间的差异无疑可以由其他昆虫衔接起来,但人类和高等猿类之间的差异就不是这样了。不过我们有各种理由可以相信,这一系列的中断仅仅是许多类型已经绝灭的结果。
欧文教授主要根据脑的构造把哺乳动物分为四个亚纲(sub-class)。他把人类专门列为一个亚纲,又把有袋类和单孔类合并列为另一个亚纲;所以他把人类从其他哺乳动物区分出来正如把有袋类和单孔类合并起来一样。就我所知道的来说,凡是能够作出独立判断的博物学者,都不同意这一观点,因而无须在这里给予进一步讨论。
我们能够理解,为什么以任何单一性状或器官为根据的分类法——即使这种器官异常复杂而重要得像脑那样——或以心理官能的高度发达为根据的分类法,几乎肯定被证明都是不能令人满意的。这一原则确曾对膜翅类昆虫试用过;但是,当以它们的习性或本能进行这样分类时,便证明这种排列法是彻底人为的了。 (345) 当然,无论根据什么性状、如身体大小、颜色或居住的自然条件进行分类都可以;但博物学者们长期以来就深信有一种自然分类法。这种分类法现已得到普遍承认,它必须尽可能地按照谱系进行排列,——这就是说,同一类型的共同后裔必须纳入一个类群中,而同任何其他一个类型的后裔分开;但是,如果亲本类型彼此有关系,那么它们的后裔也要如此,并且两个类群合在一起就会形成一个更大的类群。几个类群之间的差异量——这就是各个类群、所发生的变异量——则由属(genera)、科(families)、目(orders)、纲(classes)这样专门名词表示之。因为关于生物由来的系统,我们没有记录,所以只能根据对被分类的生物之间的类似程度所做的观察,才能发现谱系。为了这个目的,多数的类似之点要远比在少数几点的相似量或不相似量重要得多。如果两种语言在大量的单词和构造上彼此类似,它们就会被认为是从一个共同的根源发生的,尽管它们在某些少数单词或构造上有重大差异,也是如此。但对生物来说,类似之点的形成必须不是由于对相似生活习性的适应;例如,两种动物由于在水中生活,其全部身躯可能都发生变异,然而在自然分类中却不会因此把它们放得更近一点。因此,我们可以知道,在若干不重要的构造上,在无用的和线迹的器官上、即在现今已无功能作用或处于胚胎状态下的器官上,彼此的类似性何以对分类是最重要的;因为这等类似性几乎不能是在晚近期间由于适应而形成的;这样,它们就揭示了远古的生物由来的系统、即真正的亲缘。
我们还能进一步知道,某一种性状的巨大变异量为什么不应引导我们把任何两种生物分得很远。一个部分如果和亲缘相近类型的同一部分已经大不相同,那么按照进化学说而言,这个部分已经发生了重大变异;因而它就容易进一步发生同一种类的变异;这等变异如果是有利的,大概会保存下来,并由此而不断地扩大。在许多场合中,一个部分的不断发展,例如鸟喙或哺乳动物牙齿的不断发展,对这个物种获得食或达到任何其他目的都不会有什么帮助;但关于人类,我们还看不出脑和心理官能的不断发展,仅就利益而言,有什么一定界限。因此,在自然分类、即谱系分类中决定人类的位置时,不应认为极度发达的人脑其重要性超过其他较不重要的或完全不重要的大量彼此类似之点。
大多数博物学者当考察了人类的全部构造及其心理官能之后,每依布鲁曼巴哈和居维叶之说,把人类放在单独的一目(order),名为双手目(Bimana),因此同四手目(Quadrumana) (346) 和食肉目等处于相等地位。最近我们许多最优秀的博物学者们又重新遵循如此富有洞察力的林纳最先提出来的观点,他们把人类和四手类放在同一个目,名为灵长类(Primates)。这一结论的正确将会得到承认:因为,第一,我们必须记住,人脑的高度发达对分类来说在比较上并没有什么重要意义,而人类和四手类的头骨之间的强烈显著差异(比肖夫、阿比以及其他人士的最近主张)显然由于它们脑的不同发达所致。第二,我们必须记住,人类和四手类之间的几乎一切其他更加重要的差异显然是由于对它们本性的适应而发生的,而且主要同人类的直立姿势有关;如人类的手、足、骨盆的构造,脊骨的弯曲以及头部的位置,都是如此。关于适应的性状对分类不很重要,海豹科(family of Seals)提供了良好例证。这等动物在其身体形状上、在其四肢构造上同所有其他食肉类的差异,远远大于人类和高等猿类在这方面的差异;然而在大多数分类法中,从居维叶分类法一直到最近的弗劳尔(Flower)分类法, (347) 只不过把海豹列为食肉目的一个科。如果人类不是他自己的分类者,大概不会想到为了容纳自己而设置一个单独的目。
把人类和其他灵长类动物在构造上的无数一致之点列举出来,并不在我的讨论范围之内,而且也完全不是我的知识所能及的。我们伟大的解剖学家和哲学家赫胥黎教授已对这个问题做过充分讨论, (348) 他的结论是:人类在其体制的一切部分上与高等猿类的差异,小于猿类与同一类群较低等成员的差异。因而“把人类列为一个独特的目,是不正确的”。
在本书的前一部分,我曾列举各种事实以阐明人类在体质上同高等动物是多么密切一致;这种一致性决定于我们在微小构造和化学成分上的密切相似。我曾举出一些事例来说明,我们有感染同样疾病的倾向,而且有受到相似寄生虫侵袭的倾向;我们对同样的兴奋剂有共同的嗜好,而且这等兴奋剂以及各种药物对我们会产生同样的效果,还有其他诸如此类的事实。
因为人类和四手类之间的微小而不重要的类似之点,在分类学著作中普遍没有受到重视,并且因为当这等类似之点为数众多时就揭示了我们之间的亲缘关系,所以我将列举少数几点加以说明。人类和四手类的面貌上的相应部位是显著相同的;各种情绪是由肌肉和皮肤——主要是眉的上部以及口的周围的肌肉和皮肤——的差不多相似的运动所表现出来的。某些少数表情的确是差不多一样的,例如某些猴的种类的哭泣,以及其他一些种类的嘈杂大笑,在这样时候,它们的嘴角向后扯,而且眼睑起皱。彼此的外耳异常相似。人类的鼻远远高出大多数猴类的鼻;但我们可以从白眉长臂猿(hoolock gibbon)的鹰钩鼻查出猴类高鼻的开端,至天狗猴(Semnopithecus nasica) (349) ,它的鼻就大到可笑的极点了。
许多猴类的面部装饰有下巴胡子、连鬓胡子或上唇胡子。天狗猴属(Semnopithecus)一些物种的头发非常之长 (350) ;帽猴(Macacus radiatus)的头发自头顶的一点散出,向下至中部而分开。普遍都说前额使人有了高贵而智慧的面貌;但是,帽猴的浓密头发向下骤然终止,接下去的毛如此短而细,以致前额除去眉毛之外,还有一小段看来好像是完全无毛的。有人错误地断言,任何猴都没有眉毛。刚才提到的那个物种的前额无毛,其程度在不同个体有所不同;埃舍里希特说 (351) ,我们小孩的有发头皮和无毛前额之间的界限有时并不十分明显;所以我们在这里似乎找到了一个有关返祖的微小事例,人类祖先的前额那时还没有完全无毛。
众所周知,我们手臂上的毛由上下两方趋向肘的一点。这种奇异的排列同大多数低等哺乳动物的都不相似,却同大猩猩、黑猩猩、猩猩、长臂猿的某些物种、甚至某些少数美洲猴类的臂毛排列相同。但是,敏捷长臂猴(Hylobates agilis)前臂上的毛以正常方式向下趋向腕部;白手长臂猿(H.lar)前臂上的毛差不多是直立的,稍微向前倾斜而已;所以在后一物种中臂毛的趋向正处于一种过渡状态。大多数哺乳动物背部的厚毛及其趋向适应于雨水流下,简直是无可怀疑的;甚至狗的前腿上横向的毛,当它卷曲起来睡觉的时候,也可用于这个目的。华莱士先生曾仔细地研究过猩猩的习性,他说,猩猩的臂毛趋向肘部可以解释为便于雨水流下,因为这种动物在阴雨天弯臂而坐,用双手环握树枝或放在头部之上。按照利文斯通(Livingstone)的说法,大猩猩也是“在倾盆大雨中把双手置于头部之上坐在那里”。 (352) 如果上述解释是正确的话,看来似乎很可能如此,则人类臂毛的趋向提供了一个有关我们往昔状态的奇妙记录;因为谁也不会假定现今在便于雨水流下方面它还有任何用处;而且在我们现今直立的状况下,它也不适合这种目的了。
然而,关于人类及其早期祖先的臂毛趋向,不要轻率地过分相信适应的原理;因为,凡是研究过埃舍里希特所绘制的人类胎儿身上毛的排列图(成人也是一样),不可能不同意这位最优秀的观察家所说的还有其他更加复杂的原因介入其中。毛的趋向各点似乎同胚胎最后停止发育各点有某种关联。看来四肢上毛的排列似乎还同髓动脉(medullary arteries)的走向有某种关联。 (353)
千万不要假定,人类和某些猿类在上述各点以及许多其他诸点——例如前额无毛和头部的长发束等等——的类似,一定全是从一个共同祖先继续不断遗传的或后来返祖的结果。许多这等类似更可能是由于相似变异;如我在他处试图阐明的那样, (354) 相似变异的发生是由于共同起源的生物具有相似的体质,并且被诱发相似改变的相同原因所作用。关于人类和某些猴类前臂毛的相似走向,因为这一性状几乎为一切类人猿所共有,大概可以把它归因于遗传,但也并非肯定如此,因为某些亲缘很远的美洲猴类也具有这样性状。
我们已经看到,人类虽然没有正当权利为了容纳自己而设立一个单独的目,但他或者可以要求一个独特的亚目(sub-order)或科(family)。赫胥黎教授在晚近的著作中 (355) 把灵长类分为三个亚目,即:人亚目(Anthropidae),只包含人类;猴亚目(Simiadae),包括所有种类的猴;狐猴亚目(Lemuridae),包括狐猴的多种多样的属。就构造某些重要之点的差异而言,人类无疑可以合理地要求一个亚目的等级;如果我们所注意的主要是他的心理官能,那么这一等级就太低了。尽管如此,从系谱的观点来看,这个等级好像又太高了,人类只应形成一个科,可能甚至仅仅是一个亚科。如果我们想象从一个共同祖先发出的三条系统线,那么完全可以料想到,其中有两条经过长年累月之后所发生的变化如此微小,以致依然保持同属的物种地位,而第三条所发生的改变却如此重大,因而可以列为一个独特亚科、一个科甚至一个目。但在这种情况下,几乎可以肯定,第三条线通过遗传依然会保持类似于另外两条线的众多微小之点。于是,这里发生了迄今不好解决的一个难题,即在我们的分类中,对于差异强烈显著的少数各点——这就是说,对于已经发生的变异量应该给予多大注重;而对于那些表示系统线或系谱的众多不重要各点的密切类似,又应该给予多大注重。虽然许多微小的类似各点作为显示真正的自然分类来说,对其给予重大注意,看来是比较正确的,但对于少数而强烈的差异多予注重,却是最明显的而且恐怕是最稳妥的道路。
当对人类这一问题下一判断时,我们必须看一看猴科的分类。几乎所有博物学者都把这一科分为狭鼻猴群(Catarrhine group)、即旧世界猴类和阔鼻猴群(Platyrrhine group)、即新世界猴类。所有前者正如它的名称所表示的,都以鼻孔的特殊构造以及上下颚具有四个前臼齿为特征;所有后者(包括两个很特殊的亚群)却以不同构造的鼻孔以及上下颚具有六个前臼齿为特征。此外还有一些微小差异。那么,毫无疑问,人类在其齿系方面,在其鼻孔构造方面,以及在其他方面,是属于狭鼻猴类、即旧世界猴类的;除了少数不十分重要而且显然是一种适应性的性状以外,人类同狭鼻猴类的类似均比同阔鼻猴类的类似更为密切。所以,要说某些新世界物种以往发生了变异,并且产生了具有旧世界猴类所固有的一切独特性状的类人动物,同时失去了它自己所有独特的性状,乃是完全不可能的。因而人类是旧世界猴类系统的一个分支,并且从谱系观点来看,必须把他划为狭鼻猴的同类,几乎是无可怀疑的。 (356)
大多数博物学者都把类人猿、即大猩猩、黑猩猩、猩猩和长臂猿作为一个独特的亚群,同其他旧世界猴类分开。我知道葛拉条雷根据脑的构造不承认这一亚群的存在,而且无疑它是一个中断的亚群。例如,米伐特先生说,“可以看到猩猩是这一目中最特殊而脱离常轨的类型之一”。 (357) 有些博物学者还把其余不是类人的旧世界猴类分为两三个更小的亚群;具有特殊囊状胃的天狗猴属就是这等亚群的一个典型。但是,根据高德利(Gaudry)在古希腊雅典城邦(Attica)的惊人发现,那里在中新世(Miocene period)期间曾经存在过一个连接天狗猴属和猕猴属的类型,这大概证明了其他较高等的诸类群一度混合在一起的方式。
如果承认类人猿形成一个自然的亚群,那么,因为人类同他们的一致,不仅表现在人类同狭鼻猴群所共有的一切性状上,而且表现在无尾、无胼胝那些特殊性状上,同时还表现在一般面貌上,所以我们可以推论,那个类人亚群的某一古代成员产生了人类。通过相似变异的法则,任何一个其他较低等亚群的成员大概不可能产生在许多方面都同较高等类人猿相类似的类人动物。人类同其大多数亲缘相近者比较起来,曾经发生了非常大的变异量,这主要是人类脑部及其直立姿势巨大发展的结果;尽管如此,我们还应该记住,他“不过是灵长目的几个例外类型之一而已”。 (358)
凡是相信进化原理的每一位博物学者都会同意猴科的两个主要部分、即狭鼻猴类和阔鼻猴类及其亚群全是出自某一极古的祖先。这一祖先的早期后裔在其彼此分歧到相当程度之前,大概依然形成一个单一的自然群;但有某些物种,即初生的属大概已经开始以其分歧的性状表明了狭鼻猴类和阔鼻猴类的未来独特标志了。因此,这一假定的古代类型成员在其齿系或其鼻孔构造上,一方面既不像现存的狭鼻猴类、另一方面也不像阔鼻猴类那样的非常一致,而在这一点上却同亲缘相近的狐猴科相类似,后者在其鼻口部的形状上彼此差别重大, (359) 而其齿系的差别程度就非常之大了。
狭鼻猴类和阔鼻猴类毫无问题完全属于同一个目,这阐明了它们的很多性状是彼此一致的。它们所共有的那些性状简直不能由如此众多的物种那里分别获得的;因此,这等性状一定是遗传的。但是,一个古代类型如果具有狭鼻猴类和阔鼻猴类所共有的许多性状、其他处于中间状态的性状而且恐怕还有少数不同于这两个类群的性状,那么一个博物学者无疑会把它分类为一种猿或一种猴的。从系统的观点来看,由于人类属于狭鼻猴类,即旧世界的猴类系统,所以我们必须作出结论说,人类的早期祖先也应该这样称呼才是适当的,不管这个结论多么有损于人类的骄傲, (360) 都必须如此做。但我们千万不要犯这样的错误:假定包括人类在内的整个猴类系统的早期祖先同任何现存的猿或猴是完全一致的,或者即使是密切类似的。
人类的诞生地及其古老性
自然我们要被引导着去追问,当我们的祖先从狭鼻猴类系统分歧出来的时候,人类在那一进化阶段的诞生地是在哪里呢?他们属于这一系统的事实明确地指出,他们那时是栖居于旧世界的;但不是澳洲,也不是任何海洋岛,从地理分布的法则可以推论出这一点。在世界各个大区内,现存哺乳动物和同区绝灭物种是密切关联的。所以同大猩猩和黑猩猩关系密切的绝灭猿类以前很可能栖居于非洲;而且由于这两个物种现今同人类的亲缘关系最近,所以人类的早期祖先曾经生活于非洲大陆,而不是别处地方,似乎就更加可能了。但是,对这个问题进行推测是无益的;这是因为有两三种类人猿,其中之一为拉脱特命名的森林古猿(Dryopithecus) (361) ,同人差不多一样大,而且同长臂猿的亲缘关系密切,曾在中新世生存于欧洲;再者,还因为地球从一个如此遥远的时代以来,肯定发生过许多重大变迁,并且对极大规模的移居会有充分时间。
无论何时,也无论何地,当人类最初失去其覆毛的时候,很可能是栖居于一处炎热地方的;根据类推来判断,人类那时以果实为生,所以那里大概是适于这种情况的一种环境。我们远远不知道,人类在多久以前才最初从狭鼻猴系统派生出来,但可能是发生于始新世那样远古的时代;因为森林古猿的存在阐明,早在后期中新世高等猿类就从低等猿类派生出来了。我们完全不晓得,生物——无论在等级上多高或多低——在适宜的环境条件下可能以怎样的速度发生变异;然而我们知道,有些生物经过漫长的时间还保持了同样的形态。根据我们所看到的在家养下发生的情况,我们知道,同一物种的共同后裔在同一期间内,有些可能完全不变化,有些可能稍微变化,这些则可能大大地变化。因此,人类也可能是这样情形,同高等猿类比较起来,人类在某些性状上曾发生过大量变异。
在生物链上人类同其最近亲缘种之间的巨大断裂是无法由绝灭的或现存的物种连接起来的,这常常被提出来作为一种重大理由来反对人类起源于某种低等类型的信念;但对那些根据一般理由相信一般进化原理的人们来说,这种反对理由看来并没有多大分量。在生物系列的所有部分常常出现断裂,有些是广阔的、突然的和明确的,其他断裂则较此为差,程度有种种不同;例如,猩猩和其最近亲缘种之间——跗猴(Tarsius)和其他狐猿科动物之间——象和所有其他哺乳动物之间,都是如此,而鸭嘴兽(Ornithorhynchus)或针鼹(Echidna)和其他哺乳动物之间的情况就更加显著。但是,这种断裂仅仅被绝灭了的亲缘类型数量所决定。在将来的某一时期,以世纪来衡量这一时期不会很远,人类的文明种族几乎肯定要在全世界内消灭和取代野蛮种族。同时,譬如沙夫豪森教授所说的, (362) 类人猿无疑也将被消灭。那时人类和其最近亲缘种之间的断裂将更加广阔,因为现在的断裂在于黑人或澳洲土人和大猩猩之间,而那时的断裂,将在文明状态甚至高于白种人的人类——如我们所期望的——和低于狒狒的某种猿类之间。
关于连接人类和其似猿祖先之间的化石遗骸的缺乏,凡是读过莱伊尔爵士论述 (363) 的人,谁都不会过分注重这一事实,他指出在所有脊椎动物纲中发现化石遗骸乃是一个很缓慢而偶然的过程。我们也不应忘记,地质学者们迄今还没有探查到那样的地区,在那里最可能提供一些连接人类和某种绝灭的似猿动物之间的遗骸。
人类系谱的较低诸阶段
我们已经知道,人类看来是从狭鼻猴类、即旧世界猴类派生出来的,而后者在此之先又是从新世界猴类派生出来的。现在我将努力追溯一下人类系谱的古远遗迹,这主要依据各个纲之间和各个目之间的相互亲缘关系,也要稍微涉及他们相继出现于地球之上的可以确定的时期。狐猴科接近猴科,而位于其下,组成了灵长类中一个很独特的科,或者,根据赫克尔和其他人士的看法,组成了一个独特的目。这一类群的歧异和断裂已达到异常程度,其中包括许多畸变类型。所以,它很可能大量绝灭了。大部分残存者都生活在像马达加斯加和马来群岛那样的岛屿上,在那里它们所面临的竞争并不像在生物繁多的诸大陆上那样剧烈。同样地,这一类群也呈现许多等级,这些等级之多,就像赫胥黎所述说的,“从动物界最高顶峰的生物缓慢地同下直到最低等的哺乳动物——同那些胎盘哺乳动物中最下属的、最小的而且智力最低的生物看来仅仅只差一步”。 (364) 从这种种考察看来,猴科很可能原本就是从现存的狐猴科祖先发展而来的;而狐猴科又是从哺乳动物系列中最低等类型发展而来的。
有袋类在许多重要性状上都低于胎盘哺乳动物。有袋类是在较早的地质时期出现的,它们以往的分布范围要比现在广阔得多。因此,一般假定胎盘类(Placentata)起源于无胎盘类(Implacentata)、即有袋类;但不是起源于密切类似现存有袋类的类型,而是起源于有袋类的早期祖先。单孔类同有袋类的亲缘关系显然密切,前者在哺乳动物的大系列中形成了第三个还要更低的部门。今天它们仅以鸭嘴兽和针鼹为其代表;而这两个类型可以安全地被视为更加大得多的类群的残遗,其代表种在澳洲由于某些共同起作用的环境条件而被保存下来了。单孔类是显著有趣的,因为它把若干重要的构造特点引向爬行动物纲。
当我们在哺乳动物、因而在人类的系列中向下追踪其系谱时,将会愈来愈大地卷入暧昧不明之中;但是,正如一位最有才能的评论家派克(Parker)先生论述的,我们有良好的理由可以相信,在动物由来的直接系统中,并不存在真正的鸟或爬行动物。凡是想知道才智和学识能起多大作用的人,不妨请教一下赫克尔教授的著作 (365) 。我很愿意举出少量的一般论述。每一位进化论者都会承认,五个大的脊椎动物纲,即哺乳类、鸟类、爬行类、两栖类、都是从某一个生物原型传下来的;因为它们有许多部分是共同的,尤其在胚胎状态下是如此。由于鱼类的构造是最低等的,而且出现于其他几类之前,因此我们可以断言,脊椎动物界的一切成员都是从某一种似鱼的动物派生出来的。如果相信性质如此截然不同的动物,如一种猴、一种象、一种蜂鸟、一种蛇、一种蛙和一种鱼等等,完全来源于相同的双亲,那么对那些没有注意到博物学晚近进步的人们来说,这一信念就显得荒谬绝伦了。其所以如此,是因为这一信念意味着以前曾经存在过一些环节把所有这等现今完全不相像的类型紧密连接在一起。
尽管如此,肯定有些类群的动物曾经存在过,或者现在仍然存在着,多少紧密地把几个大的脊椎动物纲连接在一起了。我们已经看到,鸭嘴兽逐渐向着爬行类变化;赫胥黎教授发现,并为科普(Cope)及其他人士所证实,恐龙类(Dinosaurians)在许多重要性状上,介于某些爬行类和某些鸟类之间——这里提到的鸟类是指驼鸟族(它本身显然是一个较大类群的广为分布的残余)和始祖鸟(Archeopteryx),这种奇异的次级鸟具有一条蜥蜴那样的长尾。再者,按照欧文教授的说法 (366) ,鱼龙类(Ichthyosaurians)——具有鳍状肢的大型海蜥蜴——同鱼类表现有许多亲缘关系,根据赫胥黎的意见,更确切地是同两栖类有许多亲缘关系;两栖动物纲在其最高部分包含着蛙类和蟾蜍类,它显然同硬鳞鱼类(Ganoid fishes)密切近似。光鳞鱼类在较古的地质时期非常繁盛而且其构造是所谓一般的基本模式,这就是说,它同其他生物类群表现有各式各样的亲缘关系。南美肺鱼(Lepidosiren)同两栖类和鱼类也非常密切近似,以致博物学者们长期以来都在争论应该把它分类在哪一纲;肺鱼类,还有某些少数硬鳞鱼类由于栖居在作为避难所的河湾,免于完全绝灭,而被保存下来了,这些河湾同大洋的关系正如岛屿同大陆的关系一样。
最后,巨大而变化多端的鱼纲还有一个独一无二的成员,叫做文昌鱼(1ancelet),它和其他鱼类如此不同,以致赫克尔主张它在脊椎动物界中应该形成一个独特的纲。这种鱼以其反面的性状而著称;简直不能说它有脑、脊柱或心脏等等,所以先前的博物学者们曾把它分类在蠕虫中。许多年以前,古德瑟(Goodsir)教授发现文昌鱼同海鞘类(Ascidians)表现有某种亲缘关系,而海鞘类乃是无脊椎的、雌雄同体的水生动物,永久附着在一个支持物上。它们简直不像动物,体部为一种简单的、粗糙的、坚韧的囊,具有两个突出的小孔。它们属于由赫胥黎命名的拟软体动物门(Mulluscoida)——位于软体动物(Mollusca)大界的较低部分;但是,最近有些博物学者把它放在蠕形动物中。它们的幼体在形状上同蝌蚪多少类似 (367) ,具有自由游动的能力。柯瓦列夫斯基(M.Kovalevsky) (368) 最近观察到海鞘类的幼体在其发育方式上,在神经系统的相对位置上,而且在一种同脊椎动物的脊索(chorda dorsalis)密切相似的构造上,都同脊椎动物相关联;库弗尔(Kupffer)教授后来证实了这一点。柯瓦列夫斯基教授从那不勒斯(Naples)写信给我说,他现在对此正做进一步观察,如果他的结果充分地得到证实,那将是一项价值极大的发现。这样,要是我们可以依据胚胎学——从来就是分类的最稳妥的指南,看来我们最终就会得到一条追踪脊椎动物起源的线索。 (369) 于是,可以证明我们持有如下的信念是有道理的,即,在极其遥远的时代,曾有一动物类群存在过,它们在许多方面都同现今的海鞘类幼体相类似,海鞘类曾分为两大枝——一枝在发育上退化了,产生海鞘类现在这一纲,另一枝产生了脊椎动物,因而上升到动物界的顶峰。
在其相互亲缘关系的帮助下,我们对脊椎动物的系谱,就大略地追踪至此。现在我们来看看现存的人类;我想,我们能够部分地恢复人类早期祖先在相继时期内的构造,但不是按照适当的时间顺序。根据人类依然保持的残迹器官,根据通过返祖在人类中时而出现的一些性状,并且在形态学和胚胎学的帮助下,我们是能够做到上述那一点的。我在这里将要提到的各种事实曾在以上各章叙述过。
人类的早期祖先一定曾经一度全身被毛,男女都长胡须;他们的耳朵大概是尖形的,并且能够活动;体部有尾,有适当的肌肉。那时他们的四肢和体部还有许多对其起作用的肌肉,现在只是偶尔重现,但在四手类中却是正常存在的。在这一时期或某一更早时期,上膊骨的大动脉和神经是由上颗孔穿过的。盲肠要比现在的大得多。从胎儿的大拇趾来判断,那时的脚是能抓握的;我们的祖先无疑有树栖的习性,并出没于温暖的、覆盖着森林的地方。男性生有巨大的犬齿,用做锐利的武器。在更加早得多的时期,子宫是双重的;粪便由泄殖腔(cloaca)排泄出来;而且有第三眼睑、即瞬膜来保护眼睛。在还要更早的时期,人类的祖先一定具有水生的习性;因为形态学明显地告诉我们,人类的肺是由一种改变了的鳔(swim-bladder)构成的,后者一度作为浮囊之用。人类胎儿颈部的裂隙表明那里曾经一度有鳃存在。在我们每月或每周定期运转的机能中,还清楚地保有我们原始诞生地的痕迹,那里曾是潮水冲击的滨岸。大约在与此同样早的时期,真肾是由伍夫氏体(Corpora Wolffiana,中肾)来代替的。心脏仅仅是一种简单的搏动器;脊索代替了脊柱。在朦胧时代的遥远过去,如此看来,这等人类早期祖先的构造一定像文昌鱼那样简单,或者,甚至比文昌鱼的构造还要简单。
还有另外一点更加值得充分注意。长期以来就知道,在脊椎动物界中,某一性别生有属于生殖系统的各种附属部分的残迹物,这等部分本来是属于另一性别的;现在已经确定,在很早的胚胎时期,雌雄两性都有真正的雄性腺和雌性腺。因此,整个脊椎动物界的某一遥远的祖先看来曾经是雌雄同体的。 (370) 但在这里我们遇到了一个特别的难题。在哺乳纲中,雄性在其前列腺囊(Vesiculae prostaticae)中具有子宫及其连接管道的残迹;它们还有乳房的残迹,而且某些有袋类的雄者有袋囊。 (371) 还可以再举出另外一些与此近似的事实。那么,某种极其古代的哺乳动物在获得这一纲的主要特征之后,因而在它从脊椎动物界的较低诸纲分出来之后,我们还能假定它继续是雌雄同体的吗?这似乎是很不可能的,因为我们势必指望在鱼类——脊椎动物界中最低的一纲里去寻找依然是雌雄同体的类型。 (372) 每一性别所固有的附属部分,如果在相反性别处于残迹状态,对此可做如下解释,即,这等器官逐渐由某一性别获得,然后以多少不完全的状态遗传给另一性。当我们讨论性选择时,我们将遇到无数这样遗传的事例,——如雄鸟为了战斗或装饰获得了距、羽毛以及耀眼的色泽,这等性状则以一种不完善的或残迹的状态遗传给雌鸟。
雄性哺乳动物具有功能不完善的乳房器官,从某些方面看是特别奇妙的。单孔类动物有一种正常泌乳的腺和孔口,但没有乳头;由于这等动物在哺乳动物系列中位于最底层,所以哺乳纲的祖先很可能也是只有泌乳腺,而没有乳头。已经知道它们的发育方式支持了这一结论,因为特纳(Turner)教授根据克利克尔(K lliker)和朗格尔(Langer)的权威材料告诉我说,在胚胎中,当乳头一点也看不到之前,就可以明显地查出乳腺;而个体的相继诸部分的发育一般代表着同一个由来系统的相继诸生物的发展,二者正好一致。有袋类同单孔类的差别在于前者有乳头;所以很可能是有袋类在从单孔类分出来并高于其上之后,最先获得了这等器官,然后传给了有胎盘的哺乳动物。 (373) 在有袋类大致获得了它们现今这样的构造以后,没有人会假定它们依然保存着雌雄同体状态。那么,我们又如何解释雄性哺乳动物还有乳房呢?可能是乳房先在雌者得到发展,然后传给了雄者,但从下述情况看来,这简直是不可能的。
lliker)和朗格尔(Langer)的权威材料告诉我说,在胚胎中,当乳头一点也看不到之前,就可以明显地查出乳腺;而个体的相继诸部分的发育一般代表着同一个由来系统的相继诸生物的发展,二者正好一致。有袋类同单孔类的差别在于前者有乳头;所以很可能是有袋类在从单孔类分出来并高于其上之后,最先获得了这等器官,然后传给了有胎盘的哺乳动物。 (373) 在有袋类大致获得了它们现今这样的构造以后,没有人会假定它们依然保存着雌雄同体状态。那么,我们又如何解释雄性哺乳动物还有乳房呢?可能是乳房先在雌者得到发展,然后传给了雄者,但从下述情况看来,这简直是不可能的。
如果根据另一种观点,也可提出如下的看法,即,在整个哺乳纲的祖先久已停止雌雄同体以后,雌雄两性还都泌乳,这样来养育它们的幼仔;在有袋类的场合中,雌雄两性都有养育幼仔的育儿袋。看来这好像是并非完全不可能的,如果我们考虑到下述情形:现存的海龙类(syngnathous fishes)的雄鱼把雌鱼的卵放在它们的腹囊内,进行孵化,如有些人所相信的,此后还在其中养育幼鱼 (374) ——某些其他种类的雄鱼在口中或鳃腔中孵卵;——某些雄蟾蜍从雌者那里取来卵环,放在自己的大腿周围,使其风干,一直到把它们孵化成蝌蚪为止,——某些雄鸟完全担负起孵卵的任务,还有,雄鸽以及雌鸽都用嗉囊中的分泌物来饲喂雏鸽。但是,我最初想到上述看法,是由雄性哺乳动物的乳腺所引起的,其乳腺要比其他附属生殖部分的残迹物发达得多,这等附属生殖部分虽然为某一性别所固有,却见于另一性别。像现在雄性哺乳动物所有的乳腺和乳头实际上简直不能称为残迹的;它们只是没有充分发育而且机能活动力不强而已。在某些疾病的影响下,它们可以变得像雌者的同类器官那样地合用。它们常常在出生时或在青春期泌出少数几滴乳汁:这一事实曾在以前提到的一个奇妙事例中发生过,这一事例就是一个男性青年具有两对乳房。在男人和某些其他雄性哺乳动物中,这等器官据知有时变得如此充分发育,以致可以泌出丰富的乳汁。于是,如果我们假定,在先前一个长期内,雄性哺乳动物帮助雌者去哺育后代, (375) 以后由于某种原因(如由于产仔数量的减少),雄者不再提供这种帮助,那么器官在成熟期的不使用将会导致它们变得不活动;而且根据两项众所熟知的遗传原理,这种不活动状态很可能在相应的成熟期中传递给雄者。但是,在一个较早的时期,这等器官大概没有受到影响,所以在雌雄两性的幼仔中它们差不多是同等充分发育的。
结 论
冯·贝尔(von Baer)解释生物等级的提高或增进比其他任何人都好,他的解释是以一种生物的几个部分的分化程度和特化程度为依据的,——我愿补充一点,即这等部分是达到成熟期的。那么,由于生物通过自然选择缓慢地对多种多样的生活方式变得适应了,它们的一些部分由于从生理分工得到利益,也会在各种功能上变得越来越分化和特化了。同一部分好像常常最初是为了一个目的而改变了,于是经过长期以后又为了另一个完全不同的目的发生了改变;这样,所有部分都变得越来越复杂了。但是每一种生物依然保持着其最初祖先的一般构造模式。按照这种观点,如果我们转向地质学的证据,全体生物似乎在整个世界上都以缓慢而中断的步骤向前进了。在脊椎动物这个大界中,到人类便达到顶点。然而,千万不要假定,一旦生物类群产生其他更加完善的类群之后,它们就永远被取代而消失。更加完善的类群虽然胜过它们的先辈,但可能不会变得更好地适应于自然组成中的一切地方。有些古老类型由于栖居在有保护的处所,看来还会生存下来,在那里它们没有遇到很剧烈的竞争;这等类型可以使我们对既往消失的种群得到一个合理概念,于是在构成人类的系谱方面对我们有所帮助。但我们千万不要犯这样的错误:认为任何体制低等类群的现存成员都是它们古代先辈的完全代表。
我们对脊椎动物界中的最古老祖先虽然只能有一种模糊认识,但它们显然是由一个同现存海鞘类幼体相类似的水生动物类群组成的。 (376) 这等动物很可能产生了像文昌鱼那样低等体制的鱼的类群;从此一定又发展出硬鳞鱼类以及像肺鱼那样的其他鱼类。从这种鱼再向前做很小的迈进,就会把我们带到两栖类。我们已经看到,鸟类和爬行类一度是紧密连接在一起的;而且单孔类现在已经轻微地把哺乳类和爬行类连接起来了。但是,今天谁也无法说出三个比较高等而关联的纲、即哺乳类、鸟类和爬行类怎么通过生物由来的系统从两个较低等的纲、即两栖类和鱼类派生出来的。在哺乳纲中,从古代的单孔类到吉代的有袋类所经过的步骤,再从此到胎盘哺乳类的步骤,是不难想象的。这样,我们便可以向上追溯到狐猿科;再从此到猴科,其间隔并不很广阔。于是猴科分为两大支,一为新世界猴类,一为旧世界猴类;在一个遥远的过去时期,人类——宇宙的奇迹和光荣——从旧世界猴类产生出来了。
如上所述,我们曾指出人类有一个非常悠久的系谱,但或者可以说,他并不具有高尚的品质。人们常常说,这个世界为了人类的到来好像作了长期的准备:在某种意义上,这是完全正确的,因为他的诞生要归功于祖先的悠久系统。这条链索的任何一个环节如果从来没有存在过,人类大概就不会同现在完全一样。除非我们故意闭上双眼,那么根据我们现有的知识,我们大致可以认识我们的来历,我们无须为此感到羞耻。最低等的生物也远比我们脚下的无机尘土高出许多;一个人如果不持偏见,研究任何生物,无论其低等到何等地步,也不会不被它的奇异构造和性质所深深打动。
第七章 论人类种族
物种性状的性质及其价值——对人类种族的应用——支持和反对把所谓人类种族分类为独特的物种——亚种——一元发生论者和多元发生论者——性状的趋同——最不相同的人类种族之间在身体和心理上的无数类似之点——人类最初分布于全地球之上时的状况——每一个种族都不是来源于单独一对配偶——种族的绝灭——种族的形成——杂交的效果——生活条件直接作用的轻微效果——自然选择的轻微作用或没有作用——性选择
我无意在这里描述几个所谓的人类种族;但我要在分类学的观点下,对于什么是种族之间的差异价值以及它们是怎样起源的加以探索。在决定两个或两个以上近缘类型是否应该分类为物种或变种,博物学者们实际上是以下述事项为指针的;即,它们之间差异量,这等差异是否同少数或许多构造之点有关系,而且这等差异是否具有生理上的重要性;但更为重要的是,它们是否稳定。博物学者们所重视的和追求的主要是性状的稳定性。无论何时,只要能够阐明问题中的类型长期保持其独特性,或者很可能如此,这就可以成为一个很有分量的论点,把它们分类为物种。任何两个类型当第一次杂交时,哪怕有轻微程度的不育性,或者其后代如此,那么一般就视为这是一个决定性的证据,用来鉴别它们物种的独特性;如果它们在同一区域内继续持久的不相混合,通常就把这种情况作为某种程度的不育性的充分证据,或者,在动物的场合中就把这种情况作为某种程度的相互拒绝交配的充分证据。
撇开由于杂交而混合的情况不谈,在一个经过充分研究的地区里,如果完全缺少一些变种来联结任何两个亲缘密切的类型,那么这大概就是一个最重要的准则、用来鉴别它们物种的独特性;单单从性状的稳定性来看,这多少是一种不同的考虑,因为两个类型可能是高度易于变异的,而且还没有产生中间变种。地理分布的作用常常是无意识的,有时是有意识的;因此,生活在距离辽远的两个区域内的类型,——在那里大多数其他生物如果都是独特的物种——其本身通常也会被视为独特的物种;其实这对认识地理宗和所谓好的或真正的物种之间的区别并无助益。
现在,让我们把这等一般公认的原理应于人类的种族,以博物学者观察任何其他动物的同样精神来观察人类。关于种族之间的差异量,我们必须承认,我们从观察自己的长期习惯中得到了良好的识别能力。在印度,正如埃尔芬斯通(Elphinstone)所论述的,一个新到的欧洲人虽然不能识别各种不同的土著种族,但他很快就会发现他们是极不相似的; (377) 印度人最初也不能看出几个欧洲民族之间有任何差别。甚至特性最明确的人类种族在形态上的彼此非常相似,也远远超出了我们最初所能设想的以外;罗尔夫斯(Rohlfs)博士写信告诉我,我也曾亲眼看到,有些黑人部落具有高加索人的面貌,但某些部落必须除外。在巴黎博物馆人类学部的搜集品中,有一些法国人拍摄的各个不同种族的照片,它们充分阐明了人类种族的一般相似性,我曾把这些照片给许多人看过,他们都认为其中大多数可以冒充高加索人。尽管如此,如果我们看到这些真人,他们无疑还会显得特性很明确,所以单是毛发色和肤色,面貌的轻微差别以及表情声调,显然都会大大影响我们的判断。
然而,经过仔细的比较和测量,毫无疑问各个不同民族彼此差别甚大,——如毛发的组织、身体所有部分的相对比例 (378) 、肺的容量、头颅的形状和容量,甚至脑旋圈 (379) ,都是如此。但是,要列举无数的差异之点乃是一项无尽无休的工作。各个种族在体质上、适应气候上以及感染某些疾病上都有差别。他们的心理特性同样的也很不相同;这主要表现在他们的表情上,部分地也表现在他们的智能上。凡是有机会进行这种比较的人,一定都会被沉默寡言的、甚至是忧郁的南美土著居民和无忧无虑的、健谈的黑人之间的鲜明对照所打动。马来人和巴布亚人(Papuans)之间差不多也有相似的对照, (380) 他们生活在同样的自然条件之下,彼此仅仅被一条狭窄的海域所分开。
现在我们先对那些支持把人类诸种族分类为独特物种的论据加以考察,然后再对反面的论据加以考察。一个博物学者以前从来没有见过黑人、霍屯督人(Hottentot) (381) 、澳洲土人或蒙古人,如果对他们加以比较,他将立刻觉察到他们的许多性状是有差别的,其中有些性状是微不足道的,有些性状则是相当重要的。经过调查,他会发现他们适于在广泛不同的气候下生活,而且他们在体质上或心理倾向上多少有点差别。如果告诉他说,从同一地方可以找来数百种相似的标本,那么他会有把握地宣称,他们是不折不扣的人种,同他习惯地授以种名的那许多人种一样。一旦他确定了这等类型许多世纪以来全都保持同样的性状,而且至少在四千年前生活的黑人显然同现存的黑人完全一样, (382) 那么上述结论就会大大得到加强。根据卓越的观察家伦德(Lund)博士的权威材料, (383) 他还会知道,在巴西洞窟内和许多绝灭动物埋藏在一起的人类头骨,同现今遍布于美洲大陆者属于同一模式。
于是,这位博物学者也许要转而注意到地理分布,他很可能宣称,那些类型一定是独特的人种,他们不仅在外貌上有差别,而且有些适于炎热的地方,有些适于潮湿的或干燥的地方,还有些适于北极地区。他也许要诉诸下列事实,即在次于人类的类群——四手类中没有一个物种能够抵御低温或气候的重大变化;而且同人类关系最近的物种甚至在欧洲的温和气候下也决不会被养育到成熟。亚加西斯(Agassiz) (384) 最初注意到的下述事实将会给他留下深刻的印象,即分布于全世界的人类诸种族所栖居的动物地理区,正是哺乳动物的确实独特的物种和属所栖居的那些动物地理区。澳洲土人、蒙古人以及黑人的诸种族显然如是;霍屯督人较不显著;但是,巴布亚人和马来人明显是这样的,正如华莱士先生所阐明的,把他们分开的那条线差不多就是划分马来和澳洲二大动物地理区的那条线。美洲土著居民分布于整个大陆,乍一看这种情形好像同上述规律相反,因为南半大陆和北半大陆的大多数生物大不相同:然而少数某些现存类型,如负鼠(opossum),也分布于南北大陆,巨大的贫齿目(Edentata)中有些成员以往就是如此。爱斯基摩人就像北极动物那样,环布于整个北极地区。应该看到,几个动物地理区的哺乳动物之间的差异量同这等动物地理区的隔离程度并不一致;所以简直不应把下述情形视为反常现象,即,如果以非洲大陆的和美洲大陆的哺乳动物与其他地区的哺乳动物之间的差别相比,黑人同其他人类种族之间的差别较大,而美洲土人同其他人类种族之间的差别则较小。还可以附带提一下,看来人类原本不是栖居在任何海洋岛上的;关于这一点,他同哺乳纲的其他成员是相类似的。
要决定同一种类家养动物的假定变种是否应如此分类,或应分类为独特物种,这就是说,它们之中是否有来源于独特的野生物种的,每一位博物学者都要十分强调它们的外部寄生虫是否为独特物种这一事实。当这是一种例外情形时,就要更加强调这一事实;因为丹尼(Denny)先生告诉我说,在英国,种类大不相同的狗、鸡和鸽的身上寄生的虱子(Pediculi)是同种的。默里(A.Murray)先生曾仔细检查过从不同地方搜集来的不同人类种族的虱子; (385) 他发现它们不仅在颜色上有差别,而且爪和脚的构造也不一样。不论采集多少标本,这种差异都是固定不变的。太平洋捕鲸船船医向我保证说,有些挤在船上的桑威奇群岛居民身上的虱子传给英国水手之后,不出三四天就要死去。这等虱子的颜色较黑,看来同南美奇洛埃(Chiloe)土人身上固有的虱子有差别,他曾给过我后者的标本。再者,这等虱子比欧洲虱子的个儿大而且软得多。默里先生从非洲得到四种虱子,即两种得自非洲东海岸和西海岸的黑人,一种得自霍屯督人,一种得自卡菲尔人(Kaffirs);他还从澳洲土著居民那里得到两种;又从北美和南美各得两种。在后述这些场合中,可以推定那些虱子是来自不同地区的土著居民。昆虫只要有轻微的构造差异,如果固定不变,一般就会被估定有物种的价值,而人类诸种族身上的寄生虫如果属于独特的物种,这大概可以作为一个重要的论据来说明人类诸种族本身也应分类为独特的物种。
这位假想的博物学者的研究进行到这里时,下一步他也许要追查人类诸种族在杂交时是否有任何程度的不育性。这时他大概要请教慎重的、有哲人态度的观察家布罗卡教授的著作 (386) ,他在这部著作中将找到良好的证据来说明有些种族相交是十分能育的,不过关于其他种族也有相反性质的证据。例如,已经断定澳洲土著妇女和塔斯马尼亚土著妇女同欧洲男人很少生孩子;然而关于这方面的证据现在已被阐明几乎没有什么价值了。混血儿要被纯粹的黑人杀死:最近报道,有11个混血婴儿同时被谋杀而且被烧掉,他们的遗骸曾被警察发现 (387) 。还有,常常有人说,当黑白混血儿彼此通婚时,他们生的孩子很少;另一方面,查尔斯顿(Charleston)的巴克曼(Bachman)博士 (388) 肯定地断言,他知道一些黑白混血儿的家庭已经彼此通婚达数代之久,而他们的平均生育力与纯粹白人或纯粹黑人无异。莱伊尔爵士以前就这个问题做过调查,他告诉我说,他得到了同样的结论 (389) 。根据巴克曼博士的材料,美国1854年的人口调查表明有黑白混血儿405751人;就所有情况来考察,这个数字似乎偏小;但这可以由下述情况得到部分解释:这个阶级的地位低下而且反常,同时他们的妇女淫乱。一定数量的黑白混血儿经常被黑人所同化;这就导致了前者数量的明显减少。在一部可信赖的著作 (390) 中,黑白混血儿的低弱生命力被说成是一种众所熟知的现象;这一点虽然同其能育性的变小有所不同,但或者可以作为一个证据来证明其亲代种族乃是独特的人种。毫无疑问,无论动物杂种或植物杂种,如果是由极独特的物种产生的,都有夭亡的倾向;但黑白混血儿的双亲并不属于极为独特的人种范畴。普通的骡子多么以其长命和精力强盛而闻名于世,而它又是多么不育,这阐明了在杂种中变小的能育性和生命力之间并没有多大的必然关联;还可以举出另外一些与此相似的例子。
正如今后可以证实的那样,即使所有人类种族彼此相交而完全能育,根据其他理由把人类分类为独特物种的人也许会正当地主张能育性和不育性并不是区别物种的安全标准。我们知道,这等特性容易受生活条件变化或密切近亲交配所影响,而且受高度复杂的法则所支配,例如,同样两个物种之间正交和反交的能育性就是不相等的。关于必须分类为确实物种的那些类型,从杂交绝对不育到差不多或完全能育有一个完整的系列。不育性的程度同双亲在外部构造或生活习性上的差别程度并不严格一致。人类在许多方面可以同那些长期家养的动物相比拟,可以提出大量证据来支持帕拉斯学说(Pa1 lasian doctrine), (391) 他认为家养有消除不育性的倾向,而不育性乃是自然状况下物种间杂交的非常普遍的结果。根据这几点考察可以正当地主张,人类种族杂交的能育性,即使得到证实,也不致绝对阻碍我们不把它们分类为独特的人种。
撇开能育性不谈,杂交后代所表现的性状曾被认为可以表明亲代类型是否应该分类为物种或变种;但经过仔细研究证据以后,我得出这样的结论:没有一个这类的普遍规律可以信赖。正常的杂交结果会产生混合的或中间的类型;但在某些场合中,有些后代酷似这一亲代类型,有些却酷似另一亲代类型。如果双亲在最初表现为突然变异或畸形 (392) 的那些性状上有所差别,上述情形就特别容易发生。我之所以提到这一点,是因为罗尔夫斯博士告诉我说,他在非洲屡屡见到,黑人同其他种族的人交配之后,所生的后代不是完全黑的,就是完全白的,很少是混合颜色的。另一方面,众所熟知,在美洲,黑白混血儿普遍都呈现中间的外貌。
现在我们已经看到,这位博物学者也许以为他把人类诸种族分类为一些独特的人种是正确的。因为,他已经发现,他们是以许多构造上的和体质上的差异被区分的,其中有些是重要的。这等差异还很长期地几乎保持不变。这位博物学者在某种程度上将会被人类巨大分布范围所影响;如果把人类看为一个单独的物种,人类的分布范围在哺乳纲中,就会成为一个重大的反常现象。若干所谓人类种族的分布同其他哺乳动物的无疑是独特物种的分布彼此一致的情形将会把他打动。最后,他可能主张所有人之间的能育性迄今还没有得到证实,即使得到证实,这也不能作为绝对的证据来说明人类诸种族是同一个物种。
现在再从问题的另一个侧面来看,如果我们这位假想的博物学者去追查人类诸类型,当在同一处地方大量混合在一起时,是否像普通物种那样地保持其独特性,那么他大概会立刻发现,情况绝非如此。在巴西,他会看到大量的黑人和葡萄牙人的混血居民;在智利和南美的其他部分,他会看到整个人口是由印第安人和西班牙人的不同程度的混血儿构成的。 (393) 在同一大陆的许多部分,他会遇到黑人、印第安人和欧洲人最复杂的混血情况;根据植物界来判断,这种三重杂交对亲代类型的相互能育性提供了最严格的检验。在太平洋的某一个岛上,他会找到波利尼西亚人和英国人混血的少数居民;在斐济群岛上,有波利尼西亚人和矮小黑人(Negritos) (394) 的各种混血程度的居民。还可以再提出许多与此相似的事例,比如在非洲就是这样。因此,人类居住在同一地方不会不融合而充分保持其独特性;而不融合对于物种的独特性乃是一项普通的和最好的检验。
当这位博物学者看到所有种族赖以区别的性状都是易于变异的,他同样会感到极大困惑。凡是最初看到从非洲各地输入到巴西的黑人奴隶,都要被上述这一事实所打动。波利尼西亚人以及许多其他种族也是如此。能否举出一个种族的任何性状是独特的而且是固定不变的,尚属疑问。正如常常所断定的那样,甚至在同一部落范围内的未开化人,其性状差不多也不是一致的。霍屯都妇女有某些特征,远比其他种族的那些特征显著,但是,据知这等特征也不是固定不变的。在几个美洲部落里,肤色和毛发的差别相当大;非洲黑人的肤色有一定程度的差别,他们面貌形状的差别就相当大了。有些种族的头骨形状变异很大, (395) 各种其他性状也是如此。所有博物学者根据高价买来的经验现在都晓得,试图以不固定的性状来规定物种,则是何等轻率。
但是,一切论据中,反对人类种族是独特物种的最有分量的论据,乃是他们可以彼此逐渐进级;根据我们所能判断的来说,在许多场合中这与他们的相互杂交并无关系。对人类的研究要比对其他动物的研究来得仔细,而富有才华的鉴定家们的意见还是分歧至大,有的认为应该把人类分类为单独1个种(species)或种族(race),有的认为应该分类为2个种(维瑞,Virey),分类为3个种(贾奎诺特,Jacquinot),分类为4个种(坎特,Kant),分类为5个种(布鲁曼巴哈),分类为6个种(布丰,Buffon),分类为7个种(亨特,Hunter),分类为8个种(阿加西斯),分类为11个种(皮克林,Pickering),分类为15个种(圣文森特,B.st.Vincent),分类为16个种(德斯摩林,Desmoulins),分类为22个种(莫顿,Morton),分类为60个种(克劳弗德,Crawfurd),或者按照伯克(Burke)的意见 (396) ,分类为63个种。这样判断的分歧,并非证明人类诸种族不应被分类为种,而是阐明他们彼此逐渐级进,还阐明在他们之间几乎不可能发现明显的独特性状。
凡是不幸对一个高度变异的生物类群做过描述的博物学者,都会遇到过这种同人类恰相类似的情况(我是根据经验这样说的);如果他的性情谨慎,最终他会把彼此逐渐等级的一切类型都放在单独一个物种之下;因为他要对自己说,他没有任何权利对他无法确定的对象授以名称。这类事例见于包括人类在内的“目”,即见于某些猴属;然而在另外的属、如长尾猴属(Cercopithecus)中,大多数物种都可以确定无疑地被决定下来。在美洲的卷尾猴属(Cebus)中,种种类型被一些博物学家分类为物种,却被另外一些人士仅仅分类为地理族。于是,如果从南美各地采集来大量的所有卷尾猴标本,而且现今看来是独特物种的那些类型被发现的紧密的阶梯彼此逐渐进级,那么通常就会把它们仅仅分类分变种或种族;关于人类种族,大多数博物学者也都遵循这种方针行事。尽管如此,还必须承认有些类型不得不被命名为物种,但它们却被无数等级联结在一起,而与杂交无关,至少在植物界中是如此。 (397)
有些博物学者最近使用“亚种”(sub-species)这一术语来标示那些具有真正物种的许多特性的类型,不过简直不值得给它们这样高的等级。现在,如果我们回想一下上面列举的那些有力论据,一方面要把人类诸种族抬高到物种的高位,另一方面在决定此事上又有不可克服的困难,那么,在这里使用“亚种”这一术语恐怕还是得体的。但是,由于长久以来的习惯,“种族”这一术语也许要一直沿用下去。术语的选择只有在所用的术语尽可能地合乎表达同一程度的差异时才是重要的。不幸的是,很少能做到这一点:这是因为较大的属一般包括亲缘密切的类型。它们只能极其困难地被区别开,而同一科内较小的属所包括的类型则是完全区别分明的;然而所有这些都必须同等地分类为物种。还有,同一大属内的物种彼此相似的程度决不一样:相反,其中有些物种一般可以作为环绕其他物种的小类群加以安排,就像卫星环绕行星那样。 (398)
人类究竟是由一个人种或几个人种组成的,这是近几年来人类学者们广泛讨论的问题,他们分为两个学派,一是一元论者,一是多元论者。那些不承认进化原理的人们一定把人种视为分别创造的,或者在某种方式上把他们视为区别分明的实体;而且他们必须按照把其他生物分类为物种所通常使用的相似方法,来决定人类的什么类型应该被视为人种。但要想决定这一点,乃是一种无望的努力,除非“物种”这一术语的某种定义能够普遍得到公认;而且这个定义还必须不包括像创造作用那样性质不明的成分。在没有任何定义的情况下,或许我们也能试着决定一定数量的房屋,是否可以被称为村、镇或城市。北美的和欧洲的许多亲缘密切的哺乳类、鸟类、昆虫类以及植物,彼此相互代表,它们究竟应分类为物种或分类为地理族,还存在着无尽的疑点,这是一个难以作出决定的实例;距离大陆极近的许多岛屿上的生物也是如此。
另一方面,那些承认进化原理的博物学者们会认为所有人类种族,无疑都是来源于单独一个原始祖先,并且这一原理现在已为大多数青年所承认了;不论他们是否认为应该把人类种族命名为区别分明的人种,以表达他们的差异量,都会这样认为的。 (399) 关于我们的家养动物,各式各样的族是否起源于一个或一个以上的族,多少就是另外一个问题了。虽然可以承认所有的族以及同属的所有自然物种都发生于同一个原始祖先,但下述情况还是一个适于讨论的题目:例如所有狗的家养族是否从某一个物种最初被人类家养以来就获得了它们现今的差异量;或者,它们的某些性状是否从一个独特物种遗传而来,而这个物种在自然状况下已经发生了分化。关于人类,不会有这样的问题发生,因为不能说人类在任何特定阶段内曾被家养过。
在人类诸种族从一个共同祖先分化出来的早期阶段,种族之间以及他们的人数之间的差异一定很小;因此,就其有区别的性状而言,那时的人类种族比现存的所谓人类种族,更没有资格被分类为人种。尽管如此,人种这一术语是如此任意使用的,以致人类的这些早期种族恐怕还会被某些博物学者们分类为独特的物种;它们的差异,即使是极其轻微的,如果比它们现在的差异更为稳定或者没有彼此渐进为一,那些博物学者们就会这样进行分类的。
人类早期祖先的性状以往也许非常分歧,直到他们彼此之间的不相似比任何现存种族之间不相似更甚;可是,如福格特所提出的, (400) 此后它们的性状又趋同了,这种情形并非是不可能的,但决不一定如此。当人类为了同一个目的选择两个独特物种的后代时,仅就一般外貌而言,他时常会引起相当的趋同量。正如冯·纳图西亚斯(von Nathusius)所阐明的, (401) 从两个独特种传下来的猪的改良品种就是如此;牛的改良品种也是如此,但比较不显著。伟大的解剖学者格拉条雷(Gratiolet)主张,类人猿并不能形成一个自然的亚群(sub-group);但猩猩却是高度发展了的长臂猿、即森诺猴,黑猩猩是高度发展了的猕猴,大猩猩是高度发展了的西非狒狒。这一结论几乎完全是以脑的性状为依据的,如果是可以被承认的话,那么这至少是一个外部性状趋同的例子,因为类人猿在许多方面的彼此相似肯定比它们同其他猿类的相似更甚。所有相近的类似性,如鲸和鱼的类似,的确都可以说成是趋同的例子;但这个术语决不能应用于表面的和适应的类似性。区别甚大的生物留下改变了的后裔,它们有许多构造上的性状是密切相似的,可是,要把这种相似性也归因于趋同,那就未免过于轻率了。一个结晶体的形态完全是由分子力来决定的,而且,不相似的物质有时呈现同样的形态,并不足为奇。不过关于生物,我们应该记住,每一种生物的形态都决定于无数的复杂关系,即,决定于变异,而引起变异的原因又如此复杂,以致无法进行追查——决定于被保存下来的变异的性质,这等变异的性质又决定于自然条件,更加决定于彼此竞争的周围生物——最后,决定于来自无数祖先的遗传(遗传本身就是一个变动无常的因素),而所有这些祖先的形态又是通过与上述同等复杂的关系来决定的。如果两种生物的改变了的后裔,彼此差别显著,那么,要说它们此后又趋同到如此密切的地步,以致它们的整个体制都几乎相等,看来那就是令人不可相信的了。关于上述猪族趋同的例子,按照纳图西亚斯的说法,它们来源于两个原始祖先的证据,依然明显地保存在它们的某些头骨之中。就像某些博物学者所假定的,如果人类诸种族来源于2个或2个以上的人种,而这等人种彼此差别之大就像或差不多像猩猩同大猩猩那样,那么几乎无可怀疑的是,某些骨在构造上的显著差异,依然还可以在现存人类中发现。
虽然现存的人类诸种族在许多方面,如在肤色、毛发、头骨形状、身体比例等等方面有差别,但是,如果从他们的整个构造来考虑,可以看出他们在众多之点上还是彼此密切类似的。许多这等类似点的性质是如此不重要而且如此奇特,以致它们非常不可能是从原本独特的物种或族那里分别获得的。对于最独特的人类种族之间的无数心理上的相似点,这一意见同样可以应用或者可以更加有力地应用。任何可以举出的三个种族之间的心理差别也不会像美洲土著居民、黑人和欧洲人之间的差别那样大,然而,我在贝格尔号舰上和那几位火地人一起生活的情形时常使我激动不已,许多微小的特性阐明他们的心理状态和我们是多么相似;有一位纯种黑人也是如此,我一度和他来往很密切。
凡是读过泰勒先生和卢伯克爵士的有趣著作 (402) 的人,简直不能不对所有人类种族在嗜好、性情和习惯上的密切相似留下深刻印象。下述情形阐明了这一点:所有他们都喜欢跳舞、原始的音乐、演戏、绘画、文身或其他装饰自己的方法;他们还喜欢用姿势语言来达到相互理解,当由于同样情感而激动时,他们的面貌有同样的表情,而且发出同样无音节的呼喊。如果同猴的独特物种所做的不同表情和所发出不同叫声相对照,上述的相似性,毋宁说一致性,就引人注目了。有良好的证据可以证明,用弓射箭的技术并不是从任何共同的人类祖先传下来的,然而正如韦斯楚卜和奈尔逊(Nilsson)所论述 (403) 的,从世界最遥远地方带来的以及在最远古时期制造的石箭头差不多都是一样的;这一事实只有根据各个不同种族具有相似的发明能力、即心理能力才可以得到解释。关于某些广泛流行的装饰品,如“之”字形饰物等等,并且关于各种不同的朴素信仰和风俗,如把死人葬于巨石建筑之下,考古学者们看到过同样的情况。 (404) 我记得在南美曾看到,在那里,就像在世界许多其他地方一样,人们一般选择巍峨的高山之顶,聚石成堆,以纪念某一异常事件,或埋葬他们的死者。 (405)
那么,如果博物学者们在两个或两个以上的家养族之间,或在亲缘接近的自然类型之间,观察到习性、嗜好以及性情上的许多细微之点是密切一致的话,那么他们就会利用这一事实作为论据来说明它们来源于一个具有同样禀赋的共同祖先;因而所有它们都应分类在同一个物种之下。同样的论据可以更加有力地应用于人类种族。
由于若干人类种族之间在身体构造和心理能力上(我这里所说的不涉及相似的风俗)众多的、不重要的类似点不可能全都是分别获得的,所以这些类似点一定是从一个具有同样特性的祖先那里遗传而来的。这样,我们就能洞察人类在其逐步分布于地球整个表面之前的早期状态。人类分布于被海洋隔离得很远的各地之前,若干种族的性状无疑不会有任何大量的分歧;如果不是这样,我们就应在不同大陆上时常遇到同样的种族,但情况决非如此。卢伯克爵士在比较了世界各地未开化人现今所熟悉的技艺之一后,详细列举了当人类最初离开其原始诞生地时所不能知道的那些技艺,因为,如果一旦学会这些技艺,他们就永远不会忘记。 (406) 于是他指出,“矛不过是小刀尖端的发展,棍棒不过是槌的延长,留下来的东西仅此两件而已”。然而他承认取火之术很可能早已发现,因为所有现存的种族均能为之,而且欧洲古代洞窟居民也通晓此术。制造原始独木舟或木筏的技术恐怕也早已为人所知,但是,在遥远的古代就有人类存在,那时许多地方的陆地高度和今天的很不相同,所以人类不借助于独木舟大概也能广为分布。卢伯克爵士进一步论述,“鉴于如此众多的现存种族不会计数到四以上”所以我们的最早期祖先很不可能“计数到十”。尽管如此,即使在那样早的时期,人类的智力和合群力也几乎不会极端劣于今天最低等未开化人所具有的这等能力;否则,原始人类就不能在生存斗争中获得如此显著的成功,他在早期的广泛散布证明了这一点。
根据某些语言之间的基本差别,有些语言学者推论当人类最初广为散布时,他还不是一种有语言的动物;但可以猜测那时所用的语言并不像今天的语言那样完善,而且还要助以姿势,然而在此后更高度发达的语言中并没有留下它的痕迹。如果没有语言的使甩,不论它多么不完善,人类的智力能否升高到他早期支配地位所暗示的那样水平,尚属疑问。
原始人类只掌握少数几样技艺,而且是最粗陋的,他的语言能力也极不完善,这样,他们是否值得叫做人类必须决定于我们所采用的定义如何。由某一类猿动物徐徐地进级到现存人类,在这一系列的类型中,要想在某一固定之点上使用“人类”这一术语大概是不可能的。但这是一桩重要性很小的事情。再者,所谓人类种族不论是否这样被命名,或者被分类为人种或亚种,几乎都是无关紧要的事,不过后一名称似乎比较恰如其分。最后,我们可以断言,当进化原理普遍被接受的时候,肯定不久以后就会如此,一元论者和多元论者之间的论争将会在不声不响和不知不觉中消失。
还有一个另外的问题不应忽略而不予以注意,即人类的各个亚种或族是否出自单独一对祖先,就像时常所假设的那样。关于我们的家养动物,小心地使单独一对配偶的变异着的后代进行交配,就能够容易地形成一个新族,甚至使具有某种新性状的单独一个个体的变异着的后代进行交配,也能如此;但是,大多数家养族的形成,并不是由于有意识地选择一对配偶而来的,而是由于以某种有用的或合乎人意的方式发生变异的许多个体被无意识地保存下来的缘故。如果在某一地方习惯上喜好强壮的、重型的马,而在另一地方习惯上喜好轻型的、快速的马,那么我们可以肯定,在这两处地方并不需要挑选出一对马并使它们繁育,经过一定时间就会产生出两个区别分明的亚品种。许多家养族都是这样形成的,而且它们的形成方式同自然物种的形成方式是密切相似的。我们还知道,运到福克兰诸岛的马连续经过几代之后,就变得小而弱,而那些潘帕草原(Pampas)上的野生马则获得了较大的、接近原始形状的头;这种变化显然不是由于任何一对配偶而发生的,而是由于所有个体都处于相同的条件之下,也许还有返祖原理的助力。在这等场合中,新的亚品种并非起源于任何单独一对配,而是起源于以不同程度,但按同样一般方式发生变异的许多个体;我们可以断言,人类种族的产生也与此相似,其变异或为暴露在不同条件下的直接结果,或为某种类型的选择的间接结果。不过以后我们还要讨论后一问题。
关于人类种族的绝灭
许多人类种族或亚种族的部分绝灭或完全绝灭,乃是历史上已知的事情。洪堡(Humboldt)在南美看见过一只鹦鹉,它是能够说出一个消亡部落的语言中一个单词的唯一活生物。在世界所有地方都曾发现过古代遗迹和石器,并且在现代居民中对此并没有保留任何传说,它们暗示着大量的绝灭。有些破碎的小部落作为以往种族的残余,依然生存在隔绝的、一般是山岳的地区。按照沙夫豪森的说法,欧洲的古代种族“在等级上全都比最粗野的现存未开化人为低”; (407) 所以他们一定在某种程度上不同于现存的种族。布罗卡曾对莱埃季斯(Les Eyzies)的出土遗骸做过描述,虽然这些遗骸不幸是属于单独一个家族,却还表明了一个种族具有低等特征(即猴类的)和高等特征的奇异结合。这个种族“完全不同于我们听到过的任何其他古代的和近代的种族”。 (408) 所以它同比利时的第四纪洞穴种族也是有差别的。
人类能够长期忍耐那些看来极其不利于其生存的外界条件 (409) 。他曾长期生活在极北地区,没有木料可以制造独木舟或器具,仅以鲸油为燃料,而且溶雪为饮。在美洲的极南端,火地人没有衣服,没有任何可以称为茅舍的建筑,以资保护自己,但他们还是活下来了。南非的土著居民漫游于干旱的平原之上,那里充满了危险的野兽。人类能够顶得住喜马拉雅山脚下瘴疠的致命影响,还能顶得住热带非洲海岸流行的瘟疫。
绝灭主要是由部落与部落、种族与种族的竞争所引起的。各种抑制经常在起作用,——如周期发生的饥馑,流浪习性以及由此引起的婴儿死亡,哺乳的延长,战争,意外事故,疾病,淫乱、窃取妇女,杀婴,特别是生育力的降低——各个未开化部落的人数因此而受到压缩。如果这等抑制中的任何一种增加其力量,哪怕是微小的,也会使受到影响的部落倾向于减少人数;当两个邻接部落中的一个变得人数较少、力量较弱时,双方之间的争夺问题很快就会被战争、屠杀、吃人习俗、奴隶制以及吞并所解决。即使一个较弱的部落没有这样一扫而光,如果它的人数一旦开始减少,一般就要继续不断地减少,直至灭亡 (410) 。
当文明民族同野蛮人接触之后,斗争是短暂的,除非那里的严酷气候有助于士著种族。导致文明民族胜利的原因,有些是清楚而简单的,有些是复杂而暧昧不明的。我们可以知道,土地的耕种在许多方面都是决定未开化人命运的问题,因为他们不能或者不愿改变他们的习性。新的疾病和恶习在某些场合中被证明是有高度破坏性的;一种新疾病看来时常会造成很大的死亡,直到那些最易感受这种破坏影响的人逐渐被清除掉为止 (411) ;好酒贪杯的恶劣影响以及如此众多的未开化人的这种难改的强烈嗜好,亦复如此。还有一个更加不可思议的事实,即区别分明而彼此相隔的人民初次相遇好像会引起疾病的发生 (412) 。在温哥华岛密切注意过绝灭问题的斯波罗特(Sproat)先生相信,由于欧洲人到来而引起的生活习惯变化导致了严重的健康恶劣。他还十分强调一种显然是微小的原因,即,土著居民“被围绕他们的新生活弄得迷惑而迟钝了;他们失去努力的动机,又没有找到新动机以代之”。 (413)
竞争诸民族的文明水平似乎是他们取得成功的最重要因素。几个世纪以前,欧洲害怕东方野蛮人的侵略,现在任何这种恐怖大概都是荒谬可笑的了。有一个更加奇妙的事实,像贝哥霍特所说的,往昔未开化人在古代文明民族之前并不像他们现今在近代文明民族之前那样地衰亡下去;他们倘真如此,古代道德学家大概要对此事加以深思,但是那个时期的任何作者都未曾悲痛地记载过野蛮人的覆灭。 (414) 在许多场合中,造成绝灭的最有力的原因似乎是生育力的降低和健康的恶劣,孩子们的健康恶劣尤其会如此,这等情况是由生活条件的变化所引起的,尽管新的生活条件可能对他们本身无害,也是如此。我非常感激豪沃思(H.H.Howorth)先生,他使我注意到这个问题,并向我提供了有关资料。我曾搜集到下述事例。
当塔斯马尼亚最初殖民时,粗略估计,有些人认为那里的土著居民为7000人,其他人认为是20000人。他们的人数很快就大大减少了,这主要是由于同英国人进行战争,他们彼此也相互打仗。在全体殖民者进行了那次著名的所谓狩猎的大屠杀之后,当时向官厅自首的残余土著居民仅有120人 (415) ,这些人在1832年被运到弗林德斯(Flinders)岛。这个岛位于塔斯马尼亚和澳大利亚之间,40英里长,12~18英里宽:那里看来适于生活,而且运到那里的土人得到了良好待遇。尽管如此,他们的健康还是受到损害。到1834年,他们的成年男子为47人,成年妇女为48人,儿童16入,共计:111人。到1835年,仅余下100人。由于他们的人数继续迅速地减少,而且由于他们自己认为如果在别处,死亡当不至于如此之快,所以1847年又把他们迁移到位于塔斯马尼亚南部的蚝湾(Oyster Cove)。那时他们有男子14人,妇女22人,儿童10人(1847年12月20日)。 (416) 但是住地的变化并没有带来好处。疾病和死亡依然纠缠着他们,1864年只有1个男子(1869年死去)和3个年长的妇女活着。妇女的不孕比全部人都易患健康不良和死亡甚至是一个更加值得注意的事实。在蚝湾仅剩下9个妇女的时候,她们告诉邦威克(Bonwick)先生说(386页),她们之中只有2人生育过,而且这两个妇女一共只生过3个孩子!
关于这种异常事态的原因,斯托里(Story)博士述说,死亡是由试图使土著居民文明化引起的。“如果让他们自己像以往那样不受干涉地到处漫游,他们大概会养育更多的孩子,而且死亡率也会较小”。另一位对土著居民仔细进行过观察的戴维斯(Davis)先生述说,“出生的少,而死亡的却极多。这在很大程度上可能是由于他们的生活和食物的变化;更重要的是由于他们从范迪门领地(van Diemen's land)的大陆上被驱逐出来,而引起他们意气消沉”。
在澳洲两处大不相同的地方也曾观察到相似的事实。著名的探险家格雷戈里(Gregory)先生告诉邦威克先生说,在昆士兰,“黑人已经感到生殖的低落,并且已经显出衰亡的倾向”。有13个土著居民从沙克湾(Shark Bay)迁移到穆尔其逊河(Murchison River)流域,其中12人在三个月之内都死于肺结核病。 (417)
范东(Fenton)先生曾对新西兰的毛利人(Maories)的减少详细进行过研究,他写过一篇令人钦佩的“报告书”,下面的叙述除了一个例外都引自这份“报告书”。 (418) 自1830年以后人口数量的减少已为每一位人士所承认,包括土著居民本身在内;而且这一减少仍在稳定地进行着。虽然迄今为止已经看到要想对土著居民进行一次实际的人口调查是不可能的,但许多地方的居民已对他们的人数进行过谨慎的估计。其结果似乎是可信的,表明1858年前的十四年中人数减少了19.42%。受到这样调查的部落,有些相距一百哩以上,其中有些在海岸,有些在内地,而且他们的生活手段和生活习惯在一定程度上都有所不同(28页)。据信在1858年人口总数为53700人,在1872年,即在第二个14年期间,做过另一次人口调查,总数只有36359人,表明减少了32.29%! (419) 普通解释这一异常减少的原因为新疾病,妇女的淫乱,酗酒,战争等等,但范东详细阐明了这些原因不足之处以后,非常有根据地断定,这一减少主要决定于妇女的不生育以及婴儿特别高的死亡率(31、34页)。为了证明这一点,他指出(33页),1844年成年人与未成年人之比为2.57∶1;到1858年,成年人与未成年人之比仅为3.27∶1。成年人的死亡率也是非常高的。他提出人口减少的又一个原因为男女人数的不相等;因为生下来的男比女多。关于后一点,其原因恐怕大不相同,我在后面一章还要谈到。范东先生把新西兰的人口减少和爱尔兰的人口增长加以对比之后,感到惊讶;两地气候并非很不相似,而且两地居民的现今习惯差不多是一样的。毛利人自己(35页)“认为他们的衰落在某种程度上是由于新的食物和衣着的引入以及伴随而来的习惯的变化”;当我们考虑到变化了的外界条件对能育性的影响时,可以知道他们的看法很可能是正确的。其人口的减少始于1830—1840年之间;范东先生指出(40页),约在1830年,发明了在水中长期浸泡玉米使其发酵的精制技术,这证明了,当移住新西兰的欧洲人还很少时,土著居民的习惯就开始了变化。1835年我访问群岛湾(Bay of Islands)的时候,那里居民的衣服和食物已经发生了很大改变:他们种马铃薯、玉米以及其他农作物,并且用这些农产品交换英国的工业品和烟叶。
从帕特森(Patteson)主教传记 (420) 中的许多记载可以明显看出,新赫布里底群岛(New Hebrides) (421) 及其毗连岛屿上的美拉尼西亚人(Melanesians)当被迁移到新西兰、诺福克(Norfolk)岛以便对他们进行传教士的教育时,他们的健康大受损害而且大量死亡。
桑威奇群岛(Sandwich Islands) (422) 土著居民人口的减少有如新西兰的情形,也是众所熟知的。根据最有才能的判断者的约略估计,当库克(Cook)于1779年发现该群岛时,那里的人口大约为300000人。按照1823年不严格的人口调查,那时的人口约为142050人。1832年以及此后数年,官方进行了精确的人口调查,但我所能得到的只是下列统计表。
** 根据复算,这一栏的数字按顺序应为:4.17,2.03,0.79,2.07,2.05。——译者注
从上表可以看出,从1832年至1872年这四十年期间人口的减少竟不下于68%! (423) 大多数作者把这一情况归因于妇女的淫乱、以往血腥的战争、加给被征服部落的剧烈劳动,以及若干次起过极端破坏作用的新引入的疾病。毫无疑问,这些原因和其他这样的原因曾经是高度有效的,而且可以用来解释从1832—1836年人口锐减的情形;但其中最有力的一个原因却是能育性的降低。从1835到1837年美国海军的鲁申贝格尔(Ruschenberger)医生曾经游历过这些岛屿,按照他的说法,在夏威夷的某一大岛,每1134个男子中仅有25人的家庭有3个孩子,在另一地方每637个男子中仅有10人的家庭有3个孩子。在80个已婚妇女中,仅有39人生过孩子,而且“官方报告指出,全岛每一对夫妇平均只有0.5个孩子”。这一平均值同蚝湾塔斯马尼亚人的几乎完全一样。贾维斯(Jarves)于1843年发表的自传中说道“有3个孩子的家庭可以免去一切赋税,有3个以上的孩子赏以土地并可得到其他奖励”。由政府颁布的这一空前的法令,充分阐明了那里的种族已经变得多么不育。1839年毕晓普(A.Bishop)牧师在夏威夷的《旁观者》杂志上写道,大量的儿童都早期夭亡,斯特利(Staley)主教告诉我说,现今的情况依然如此,在新西兰恰好也是这样。这种情形常被归因于妇女对儿童的照顾不周,但很可能是大部分由于儿童体质的先天衰弱,而这同他们双亲能育性的降低又有关系。再者,同新西兰情况相似的还有如下事实,即男婴远远超过女婴,1872年的人口调查表明;一切年龄的男和女之比为31650人对25247人,即每100名妇女对125.36名男子;而在所有文明的地方,女性却超过男性。妇女的淫乱无疑可以用来部分地解释其能育性的衰弱;但他们生活习惯的变化则是一个远为可能的原因,而且同时还可以解释死亡率的增大,特别是儿童死亡率的增大。1779年库克、1794年范库弗(Vancouver)访问了夏威夷群岛,此后捕鲸船也常常来此访问。1819年传教士到达,发现崇拜偶像在那里已被废除,而且国王完成了另外一些改革。在这一时期以后,土著居民的差不多一切生活习惯都迅速发生了变化,不久他们就成为“太平洋岛民中的最文明者”。向我提供资料的一位人士寇恩(Coan)先生出生于该群岛,他说那里土著居民50年间发生的变化比英国人1000年间发生的变化还要大。根据斯特利主教给我的资料,那里比较贫穷的阶级食物方面的变化似乎并不很大,虽然许多新种类的水果已被引进,而且甘蔗已普遍食用。由于他们热心地仿效欧洲人,所以在早期他们就改变了服装的样式,而且饮酒普遍盛行。虽然这等变化看来无足轻重,但我根据所知道的动物情形,充分相信它们足可以降低土著居民的能育性了。 (424)
最后,麦克纳马拉(Macnamara)先生 (425) 述说,在孟加拉湾东侧的安达曼群岛(Andaman Islands)上的退化的低等居民“对气候的任何变化都显著敏感:事实上如果把他们迁移到海岛家乡之外,几乎肯定就要死亡,而且这同食物和外在影响并无关联”。他进一步述说,夏季极其炎热的尼泊尔山谷中的居民,还有印度各地的山岳部落,当在平原居住时常受痢疾和热病的危害;如果他们试图全年都在那里度过,就会死去。
由此我们知道,许多人类比较野蛮的种族当遇到外界条件或生活习惯发生变化时,其健康就容易受到严重的危害,如果迁移到新的气候条件下,也会如此。仅仅是习惯的改变,看来对其本身并无害处,似乎也会产生这种相同的效果;而且在若干场合中,儿童特别容易受害。像麦克纳马拉先生那样,往往有人说,人类能够泰然地抵抗多种多样的气候以及其他变化;但这只是对文明种族来说才是如此。处于原始状态的人类在这方面似乎像其亲缘关系最近的类人猿那样,差不多具有同样的敏感性,当类人猿离开其本土时,决不会活得很久。
由于条件变化而引起能育性的降低,如塔斯马尼亚人、毛利人、桑威奇群岛居民、显然还有澳大利亚土著居民的情形,比他们容易健康恶劣和死亡更为重要;因为,甚至是轻微程度的不育性,如果同那些可以抑制人口增长的其他原因结合起来,也会迟早导致绝灭。能育性的降低在某些场合中可以由妇女的淫乱(如塔希提人 (426) 晚近的情形)得到解释,但范东先生指出,这一解释对新西兰人来说并不是充分的,对塔斯马尼亚人也是如此。
在上面引用的论文中,麦克纳马拉先生举出理由使我们相信,流行疟疾地区的居民容易不育;但这对上述几个事例不能应用。有些作者提出,海岛土著居民由于常期不断的近亲繁殖使能育性和健康都受到损害;但在上述场合中,不育性同欧洲人的到达如此密切符合,以致我们无法承认这种解释。今天我们也没有任何理由可以相信,人类对近亲繁殖的恶劣效果是高度敏感的,特别是新西兰的地域如此广阔,桑威奇群岛的位置如此变化多端,更不致如此。相反,我们知道诺福克的现在居民差不多全是从堂兄弟姐妹或近亲,就像印度的图达人(Todas) (427) 和苏格兰的某些西方海岛居民那样;但他们的能育性似乎没有因此受到损害。 (428)
根据从低于人类的动物来类推,可以提出一个远为可能的观点。生殖系统对变化了的生活条件的极度敏感是能够阐明的(我们还不知道为什么);这种敏感性导致了有利的或有害的结果。关于这个问题,我搜集了大量事实,见《动物和植物在家养下的变异》第2卷,第18章,在这里我只能举出极其简略的提要;凡是对这个问题有兴趣的人可以参考上述著作。很轻微的变化可以使大多数或所有生物增进健康并提高其活力和能育性,而另外的变化据知可以使大量动物成为不育的。最为大家所熟知的一个例子是,印度的驯象不生育;但阿瓦(Ava) (429) 的象就常常生育,在那里,允许雌象在某种范围内漫游于森林之中,这样它们就被置于更加自然的条件之下了。如种美洲猴的雄者和雌者在其原产地多年来都养在一起,但它们很少或者从来不生育,这是一个更加适当的事例,因为美洲猴同人类的关系很近。值得注意的是,条件多么轻微的一种变化常常会致使被捕获的野生动物发生不育性;而更加奇怪的是,所有我们的家养动物都比它们在自然状况下更为能育;其中有些家养动物还能抵抗最不自然的外界条件,但其能育性并不降低。 (430) 动物的某些类群远比另外一些类群更加容易受到拘禁的影响;而同一类群的所有物种一般都是按照同一方式受到影响。但是,有时只是一个类群中的单独一个物种成为不育的,而另外一些物种并不如此;另一方面,可能只有单独一个物种保持其能育性,而大多其他物种都不生育。某些物种的雄者和雌者在原产地如果受到拘禁,或者,如果不允许它们完全自由生活,而只是有限地自由生活,那么它们从不交配;处于这样环境条件下的其他物种虽然常常交配,但从不产生后代;还有一些物种产生后代,但比在自然状况下产生的为少;同人类的上述事例联系起来看,注意儿童易于衰弱多病或畸形以及早期夭亡是重要的。
鉴于生殖系统对生活条件变化的敏感是多么普遍的一项法则,而且这一法则也适用于人类的最近亲属四手类动物,所以我简直不能怀疑,同样它也可以应用于原始状态的人类。因此,如果任何种族的未开化人的生活习性突然发生变化,他们就会或多或少地变得不育,而且他们孩子的健康也要受到损害,这同印度的象和猎豹(hunting-leopard) (431) 、美洲的许多种类的猴以及所有种类的多数动物当被移出其自然条件时所发生的情况是一样的,而且其原因也是相同的。
我们可以知道,长期生活在海岛上的、而且一定是长期处于差不多一致条件下的土著居民为什么特别容易感受生活习性任何变化的影响;事实似乎就是如此。文明种族在抵抗所有种类的变化方面肯定远比未开化人为优;关于这一点,他们同家养动物相类似,因为后者的健康有时虽然受到损害(例如欧洲狗在印度的情况),但他们极少是不育的,不过有少数这样的事例曾被记载过。 (432) 文明种族和家养动物没有受到这种影响,很可能是由于他们比大多数野生动物曾在更大范围内受到多种多样的、即变化着的外界条件的支配、因而就多少更习惯于这样条件;还由于他们以往曾到处迁徙或到处被运送;而且还由于不同家族或不同亚种族之间曾相互杂交。土著居民同文明种族只要进行一次杂交,前者似乎就可以立即免除由条件变化所引起的恶劣结果。例如,塔希提人和英国人的杂交后代当移居在皮特凯恩岛(Pitcairn Island) (433) 后,他们增加得如此之快,以致该岛很快就人满之患了;1856年6月他们又被移到诺福克岛。那时他们的人数中已婚者为60人,儿童134人,共计194人。同样的,他们在那里也增加得非常之快,虽然1859年有16人返归皮特凯恩岛,但到1868年正月仍增加到300人,男女正好各半。这个事例同塔斯马尼亚人的情况是多么明显的一个对照;诺福克岛民仅在12.5年期间内由194人增至300人;而塔斯马尼亚人在15年期间内由120人减至46人,其中只有10个儿童 (434) 。
再者,根据1866—1872年这一期间的人口调查,桑威奇群岛的纯血土著居民减少了8081人,而那些被认为健康较好的混血儿却增加了847人。但我不清楚后一数字是否包括混血儿的后代,或者仅仅是第一代混血儿。
我这里所举的例子全是关于土著居民由于文明人移入而遇到新条件的情况。如果未开化人受到某种原因所迫,例如征服部落的侵入,背井离乡以及改变习惯,大概也会引起不生育和不健康。有一个有趣的情况:野生动物变为家养的主要抑制,在于它们最初被捕获时的自由繁育能力,而野蛮人初与文明人接触,能否形成一个文明种族而生存下来的主要抑制,也是一样的,即在于由生活条件变化而引起的不育性。
最后,人类诸种族的逐渐减少而终至绝灭,虽然是一个由许多原因所决定的高度复杂问题,同时这等原因又随时随的有所不同;但这个问题同高等动物之一,例如化石马的绝灭问题还是一样的,马在南美消亡之后不久,就在同一地区内被无数的西班牙马群取而代之了。新西兰人似乎已经意识到这种相似现象,因为他把自己的将来命运比做当地的鼠,现在后者差不多已被欧洲鼠消灭了。如果我们要确定其真实原因及其作用的方式,在我们想象中这虽然是困难的,而且的确是困难的,但对我们的推理来说,并不应该这样困难,只要我们切记各个物种和各个种族的增加会不断地受到种种方面的抑制就可以了;所以,如果增添了任何新的抑制,哪怕是一种轻微的抑制,这个种族的数量肯定也会减少;数量的减少迟早要导致绝灭:在大多数场合中,其结局将由征服部落的侵入而迅速决定之。
论人类种族的形成
在某些场合中,独特种族的杂交曾导致新种族的形成。欧洲人和印度人都属于雅利安(Aryan)人血统,所用的语言基本一致,但他们的面貌却大不相同,而欧洲人和犹太人的面貌虽差别不大,但后者却为闪米特人(Semitic)血统,并且使用一种完全不同的语言,布罗加 (435) 对这一奇特事实提出如下解释:某些雅利安人的分支在其广泛散布的期间,曾同土著部落的人大量进行了杂交。当两个种族密切接触而进行杂交后,其最初的结果乃是一种异质的(heterogeneous)混合:这样,亨特先生在描述一个印度山地部落桑塔利人(Santali)时说道:“由黑色的、矮胖的山地部落到具有智慧之额、平静之眼和高而狭之头的高个子橄榄色的婆罗门 (436) ”可以查出成百上千的级进;所以在法庭上有必要询问证人,他是桑塔利人还是印度人。 (437) 一个异质的种族,如波利尼西亚群岛上的一些居民,系由两个独特种族形成的,只留下少数或者没有留下纯种的成员,他们是否会成为同质的,还没有直接的证据足以说明。但是,关于我们的家养动物,经过少数几代细心选择的过程, (438) 肯定能够形成一个固定的和一致的杂交种,我们于是可以推论,一个异质的混血种族在长期传衍中的自由杂交大概可以代替选择的作用,而胜过任何返祖的倾向;所以,杂交种族虽然没有同等程度的具有双亲种族的性状,它最终还会成为同质的。
关于人类种族之间的差异,皮肤的颜色最惹人注目而且最为显著。以往认为,这种差异可以由长期暴露于不同气候中得到解释;但帕拉斯阐明,这种解释是站不住脚的,以后差不多所有人类学者都追随他的主张。 (439) 这一观点之所以遭到驳斥,主要因为各种不同皮肤颜色的种族一定长期居住在他们现今的故乡,而他们的分布同气候的相应差异并不符合。根据最优秀的权威意见, (440) 我们听说有些荷兰人的家族在南非居住了300年之后,其皮肤颜色丝毫也没有发生改变,对这等事例似乎多少应该给予一点重视。属于这方面的论据还有一个,即世界各地的吉卜赛人和犹太人的面貌都是一致的,不过犹太人面貌的一致性多少被夸大了。 (441) 一种很潮湿的或很干燥的空气曾被假定比单纯的炎热对于改变皮肤的颜色更有影响;但多比尼(D'Orbigny)在南美和利文斯顿(Livingstone)在非洲关于潮湿和干燥所得出的结论正好相反,对这个问题的任何结论都必须看为很可疑的。 (442)
我在别处所举的各种事实,证明皮肤和毛发的颜色时常同完全避免某些植物毒害的作用以及某些寄生虫的侵袭以一种可惊的方式彼此相关。因此我想到,黑人和其他黑皮肤的种族也许由于较黑的个体在一长列的世代中逃脱了他们家乡瘴疠的致命影响而获得了黑的色泽。
以后我发现韦尔斯(Wells)博士 (443) 长期以来就持有同样见解。黄热病在热带美洲是一种毁灭性的病 (444) ,而黑人、甚至黑白混血儿几乎可以完全避免这种病,这种情形早为世人所知。他们在很大程度上还能避免致命的疟疾,这种病流行于非洲沿岸至少达2600英里之遥,白人殖民者每年有1/5死于此病,还有1/5因患此病被送回家乡。 (445) 黑人的这种免疫性似乎部分是先天的,这决定于某种未知的体质特性,部分乃是水土适应的结果。普歇说,由埃及总督借来参加墨西哥战争的黑人团队是在苏丹(Soudan)附近招募的,他们同那些原本来自非洲各地并且习惯于西印度群岛(West Indies)的黑人,差不多同样能够避免黄热病。 (446) 有许多事例表明,黑人在较寒冷的气候下居住一些时候之后, (447) 就变得多少容易感染热带的热病,这一情况阐明了水土适应是有一定作用的。白人曾经久居其下的气候性质同样对他们也有某种影响;因为,1837年在德梅拉拉(Demerara) (448) 流行可怕的黄热病期间,布莱尔(Blair)博士发现,移民的死亡率同其家乡的纬度是成比例的。黑人的免疫性作为水土适应的结果而言,这意味着需要非常悠久的时间;因为,热带美洲的土著居民自远古以来就在那里居住,还不能避免黄热病;特里斯特拉姆(H.B.Tristram)牧师说,在北非有些地方,虽然黑人能够安然无恙地住,而土著居民却要被迫年年从那里离去。
黑人的免疫性同其皮肤颜色的任何程度的相关不过是一种推测而已:这种免疫性也许同其血液、神经系统或其他组织的某种差异相关。尽管如此,根据上述事实,根据面色同肺病之间显然存在的某种关联,这种推测在我看来并非是不近理的。因此,我曾试图确定这种推测究竟有多大可靠性,但没有获得很大成果。 (449) 已故的丹尼尔(Daniell)博士曾长期在非洲西海岸居住,他告诉我说,他不相信有任何这种关联。他本身的皮肤异常之白,却惊人地经受了那里的气候。当他在少年来到非洲西海岸的时候,一位年老的有经验的黑人酋长根据他的面貌预言他能如此。安提瓜(Antigua) (450) 的尼科尔森(Nicholson)博士研究了这个问题之后给我写信说,暗色皮肤的欧洲人避免黄热病的,比浅色皮肤的欧洲人为多。哈里斯(J.M.Harris)先生完全否认深色毛发的欧洲人比其他人能够更好地经受炎热的气候;相反,经验教导他,当挑选在非洲海岸服务的人员时,要找那些红发的人。 (451) 有一种假说认为皮肤黑色的产生,是由于日益变黑的个体曾经长期在引起发热的瘴疠中更好地生存下来的缘故,然而仅就以上那些迹象而言,这种假说似乎就没有什么根据了。
夏普(Sharpe)博士述说, (452) 热带的太阳可以把白人的皮肤晒伤而起水疱,但对黑人的皮肤却一点也不损害;他又说,这并非由于个体的习性所致,因为6个月或8个月的黑人小孩常常被裸体抱出,并不受影响。一位医务人员使我确信,几年之前每到夏季,他的双手就出现淡褐色的斑块,同雀斑相似但较大,一到冬季即行消失,这些斑块决不受日灼的影响,而其皮肤的白色部分有几次却受到严重的日灼而起水疱。在低于人类的动物中,复被白毛的皮肤部分和其他部分对太阳作用的反应也有一种体质差异。 (453) 皮肤不被这样灼伤对说明人类通过自然选择逐渐获得暗色是否具有足够的重要性,我还不能作出判断。果真如此,我们就应假定热带美洲的土著居民生活在那里的时间远比黑人在非洲生活的时间或巴布亚人在马来群岛南部生活的时间为短,正如浅色皮肤的印度人居住在印度的时间比这个半岛中部和南部的深色皮肤的土著居民居住在那里的时间为短。
以我们现在的知识来看,虽然还不能解释人类种族肤色的差异是由于因此获得任何利益,或是由于气候的直接作用所致;但我们千万不要对后一种作用完全加以漠视,因为有充足的理由可以相信某些遗传的效果可以由此产生。 (454)
我们在本书第二章中已经看到,生活条件以一种直接的方式影响身体构造的发育,而这种效果是遗传的。这样,正如众所公认的,在美国,欧洲殖民者的面貌发生了轻微的、但异常迅速的变化。他们的体部和四肢都变长了;我听伯尼斯(Bernys)上校说,在晚近的美国战争期间,提供了有关这一事实的良好证据,即德国军队穿上为美国市场缝制的现成服装时,样子显得滑稽可笑,这等服装的各种尺寸对德国人都太长了。还有大量的证据可以阐明,美国南部诸州的第三代家内奴隶在面貌上同田间奴隶已有显著不同。 (455)
然而,如果注意看一看分布于全世界的人类种族,我们就一定会推论他们的特性差异不能由不同生活条件的直接作用得到解释,即使受到这种作用的时间非常悠久,也是如此。爱斯基摩人完全以动物食品为生,他们穿着厚皮衣,暴露在酷寒和长期黑暗之中;中国南方的居民完全以植物食品为生,几乎裸体,暴露在炎热和阳光耀眼的气候之中,但二者之间并没有任何极度的差异。不穿衣服的火地人以荒凉海岸的水产品为生;巴西的波托鸠斗人(Botocudos) (456) 漫游于腹地的炎热森林之中,主要以植物性食品为生;然而这等部族如此密切相似,以致有些巴西人误把“贝格尔舰”上的火地人当做波托鸠斗人。再者,波托鸠斗人以及热带美洲的其他居民同大西洋彼岸的黑人完全不同,但他们暴露在差不多相似的气候之下时遵循几乎一样的生活习惯。
人类种族的差异也不能由身体各部分的增加使用或减少使用的遗传效果得到解释,即便能如此,也只能起到十分轻微的作用。惯常在独木舟生活的人们,腿多少有点短;居住在高原地区的人们,胸部可能增大;不断使用某些感觉器官的人们,容纳这等器官的腔可能多少有些增大,因而他们的容貌也会稍有改变。关于诸文明民族,颚部的缩小是由于减少使用——为了表达不同的感情,惯常运转不同的肌肉——脑的增大是由于智力活动的增强——当同未开化人比较时,所有这些都对文明民族的一般面貌产生了相当的影响。 (457) 身材增大,而脑的大小不相应增加,可能使某些种族的头骨加长,而成为长头型(由上述家兔的例子可以推断)。
最后,所知甚少的相关发育原理有时也会发生作用,如肌肉的非常发达同眶上脊的强烈突出的相关就是如此。皮肤颜色同毛发颜色显然相关,如北美曼丹人(Mandans) (458) 的毛发组织同其颜色就是相关的。 (459) 皮肤颜色同它发出的气味同样也有一定的关系。关于绵羊品种,一定面积上的羊毛数量同分泌孔的数量有关联 (460) 。如果我们可以根据家养动物进行类推的话,则人类构造的许多变异大概也在相关发育这一原理的支配之下。
现在我们已经看到,人类外部特征的差异不能由生活条件的直接作用,也不能由身体诸部分的连续使用、同时还不能由相关原理得到令人满意的解释。所以这就引导我们去追问,人类显著容易发生的轻微个体差异是否不会在一长列的世代中通过自然选择而被保存下来并有所扩大。但在这里我们立刻会遇到这样的障碍,即只有有利的变异才能这样被保存下来;就我们所能判断的来说,虽然对于这一问题的判断往往容易陷于错误,人类种族之间的差异对他并没有任何直接的或特别的用处。当然,智慧的、道德的或社会的能力一定不在此论之内。人类种族之间外在差异的巨大变异性,同样地表明了它们并不具有多大重要性;如果是重要的,它们很久以前或者被固定而保存下来,或者被消除掉。在这方面,人类同那些被博物学者们称为变化多端的、即多态的类型相类似,这等类型极其容易变异,这似乎是由于这等变异具有无关紧要的性质,而且由于它们因此逃避了自然选择的作用。
在我们解释人类种族之间的差异的所有试图中,遇到了上述这么多的阻碍;但还剩下一个重要的力量即性选择,看来曾对人类而且也对许多其他动物发生过强有力的作用。我的意思并非断言根据性选择可以解释人类种族之间的一切差异。还留有不可解释的一点,由于我们对此是无知的,我们只能说,因为一些个体生下来,比如,就具有稍微圆一些或狭一些的头以及稍微长一些或短一些的鼻子,所以,如果诱发这等轻微差异的未知力量比较经常不断地发生作用,它们就会变得固定而一致。在本书第二章提到这等变异时,是把它们纳入暂时的那一类的,由于还缺少较好的术语,常常把它们叫做自发性的(spontaneous)。我也并非妄说性选择的效果能够以科学的正确性被表示出来;但可以阐明,看来曾对无数动物发生过强有力作用的这种力量,如果没有使人类改变,那么这就是一个无法解释的事实了。进一步可以阐明,人类种族之间的差异,如肤色、毛发、面型等等,预料是处于性选择影响之下的一类差异。但是,为了恰当地处理这个问题,我发现有必要对整个动物界加以回顾。因此,我在本书的第二部分对此进行讨论。最后,我还要回到人类上来,在努力阐明人类通过性选择发生了怎样程度的改变之后,再对第一部分的各章做一个简短的提要。
附录 人类和猿类的脑部在构造和发育上的异同
英国皇家学会会员 赫胥黎教授 著
关于人类和猿类脑部构造的差异性质和差异程度,早在15年前已发生了争论,直到现在还没有停止,虽然现在所争论的主要问题同以往已经完全不同了。最初有人一再异常顽固地断言,一切猿类的脑,甚至最高等猿类的脑,都和人类的脑有所不同,因其缺少诸如大脑半球的后叶以及这等后叶中所包含的侧室后角和小海马体(hippocampus minor)等那样的显著构造,而这些构造在人类中都是非常明显存在的。
问题中的这三种构造在猿脑中发育之良好,同人类无异,或者说甚至更好;而且具有良好发育的这三部分乃是一切灵长类的特征(如果狐猴类除外),这一真实情况的基础之稳固有如比较解剖学中的任何命题。再者,一长系列的解剖学家,凡是近年来特别注意到人类和高等猿类大脑半球表面上复杂的脑沟(sulci)和脑回(gyri)的排列者,无不承认它们在人类和猿类中的配置形式都是完全一样的。黑猩猩脑的每一个主要脑沟和脑回都明显地代表着人脑的这等部分,所以应用于人脑的专门术语完全可以应用于猿脑。关于这一点,已没有任何不同的意见了。数年前比肖夫教授曾就人类和猿类的脑旋圈发表过一篇论文; (461) 我的这位博学同事的目的肯定不在于降低猿类和人类关于这方面的差异价值,所以我愿引述该文如下:
“猿类尤其是猩猩、黑猩猩和大猩猩在其体制上同人类很接近,比同任何其他动物都更加接近得多,这是众所熟知的一个事实,已无所争论。单以体制的观点来看事物,大概没有人再对林奈(Linnaeus)的观点进行争论,即,人类仅仅作为一个特殊的物种,也应被置于哺乳动物以及猿类之首。人类和猿类的一切器官表明了它们的亲缘关系如此之近,以致为了证实它们之间确有差异存在,还需要进行极精确的解剖研究。对于脑部也要如此。人类、猩猩、黑猩猩以及大猩猩的脑尽管有非常重要的差异,但彼此还很接近”(原著101页)。
至于猿脑和人脑在基本性状上的相似已无争论余地了;甚至黑猩猩、猩猩和人类的大脑半球上脑沟和脑回的排列细节也表现了惊人的密切相似性,对此亦无任何争论的余地。高等猿类的脑和人脑之间在差异性质和差异程度上也不存在任何严重的问题。众所周知,人类的大脑半球绝对的和相对的大于猩猩和黑猩猩的这一部分;其前叶由于眶脊向上隆起,因而凹入较少;人类的脑回和脑沟的配置通常都对称较差,并且呈现了较大数量的次级褶。而且,众所共认,人类的颞颥后头裂(temporo-occipital)、即“外垂直”裂通常不甚显著,而这是猿脑的一个强烈显著的特征。但这等差异显然并不构成人脑和猿脑之间的明确界限。关于葛拉条雷所谓的外垂直裂,例如人脑的,特纳教授有如下记述 (462) :
“在一些人脑中,它简单地表现为大脑半球边缘的一种齿痕,但在另外一些人脑中,它却伸长到一定距离,多少横向外出。我曾见到,它在一只女脑右半球上向外逾出二英寸以上;在另一个标本上,也是右半球,它向外逾出十分之四英寸,然后向下延伸,直达半球外表面的较低边缘。在大多数四手类动物中,这种脑裂沟是有显著特征的,相比之下,大部人脑的这种裂沟就不那样完全明确了,其所以如此,乃是由于人脑具有某种表面的、十分显著的次级旋圈,以沟通诸裂沟,并把颅顶叶(parietal lobe)和后头叶(occipital lobe)连接在一起了。这等第一级的起沟通作用的脑回位置同纵向沟裂愈近,则颅顶后头的外在沟裂就愈短”(原著12页)。
因此,葛拉条雷所谓的外垂直裂的消失,并不是人脑的一种固定特性。另一方面,它的充分发育也不是高等猿类脑的一种固定特性。因为,在黑猩猩脑的这一侧或那一侧外垂直脑回由于起沟通作用的脑旋圈而或多或少强烈消失的情况,已由罗尔斯顿(Rolleston)教授、马歇尔先生、布罗卡以及特纳教授一再予以记载。特纳教授在一篇有关这个问题的专门论文中写道: (463)
“葛拉条雷曾试图形成一种概念,把第一级起连接作用的脑旋圈的完全缺如以及次级脑旋圈的隐蔽作为黑猩猩脑的基本特征,但刚才描述的这种动物脑的三个标本证明了这一概念决不能普遍应用。只有一个标本的脑在这等特点上符合葛拉条雷所表明的法则。关于起沟通作用的上部脑旋圈,我以为它是存在于一个脑半球之上的,至少迄今为人所绘出的或所描述的多数黑猩猩脑是如此。起沟通作用次级脑旋圈的表面位置显然较不常见,我相信,到目前为止只见于这份报告中所记载的A脑。两个脑半球上的旋圈排列是不对称的,以往的观察家们对此已有所描述,在这些标本中也得到了清楚的图示。”(8,9页)
颞颥后头脑沟或外垂直脑沟的存在,即便说是高等猿类和人类之间的一个区别标志,但阔鼻猴类脑的构造使这种作为区别的性状的价值很可怀疑了。事实上,尽管颞颥后头脑沟为狭鼻猴类、即旧世界猴类的最固定的一种性状,但在新世界猴类中,它的发育从来不很强烈;在较小的阔鼻猴类中根本没有这种性状;在僧面猴(Pithecia)中它的发育是残迹的; (464) 在蛛猴(Ateles)中它已经被起沟通作用的脑旋圈或多或少地所消除。
在单独一个类群范围内如此容易变异的一种性状不会有任何重大的分类价值。
已经进一步证实,人脑两侧旋圈的不对称程度有很大个体变异;而且在经过检查的布什门(Bushman)族的那些个体中,其两半球上的脑回和脑沟远不如欧洲人的复杂,但比他们的对称,然而,在黑猩猩的一些个体中,脑回和脑沟的复杂性和不对称性却是值得注意的。布罗卡所绘制的一只幼小雄猩猩的脑,其情况尤其如此(《灵长目》,L'ordre des Primates,165页,图11)。
再者,就脑的绝对体积问题来说,已经证明最大的和最小的健康人脑之间的差异,比最小的健康人脑同最大的黑猩猩脑或猩猩脑之间的差异为大。
还有一种情况使猩猩脑和黑猩猩脑同人脑相类似,而同低等猿类的脑有所区别,这就是前者具有两个乳头体(corpora candicantia)——而犬猿这一类(Cynomorpha)只有一个。
鉴于这等事实,在这1874年我毫不踌躇地重复并坚持1863年我提出的主张: (465)
“就脑部构造来说,显然,人类同黑猩猩或猩猩之间的差异,甚至小于后者同猴类之间的差异,而且,黑猩猩脑同人脑之间的差异,如黑猩猩脑同狐猴脑之间的差异相比,就几乎微不足道了。”
在我以前引用的那篇论文里,比肖夫教授并不否认这一叙述的第二部分,但第一,他文不对题地说,如果一个猩猩脑和一个狐猴脑很不相同,那是毫不足怪的;第二,他继续断言,“如果我们连续地以人脑同猩猩脑相比较,以猩猩脑同黑猩猩脑相比较,以黑猩猩脑同大猩猩脑相比较,以次及于长臂猿(Hylobates)、天狗猴(Semnopithecus)、鼯猴(Cynocephalus)、长尾猴(Cercopithecus)、猕猴(Macacus)、卷尾猴(Cebus)、绢毛猴(Callithrix)、狐猴(Lemur)、懒猴(Stenops)和狨(Hapale),我们在脑旋圈发育程度上所看到的间隙并不比人脑同猩猩脑或黑猩猩脑之间的间隙更大,甚至相等。”
对此我回答如下:第一,不论这一主张的真伪如何,都同《人类在自然界的位置》一书所提出的主张毫无关系,该书所讨论的并非仅仅是脑旋圈的发育,而是脑的整个构造。如果比肖夫教授不厌其烦地阅读一下他所批评的该书第96页,实际上他将会看到下文:“值得注意的一个情况是,就我们现有知识所能达到的来说,在猿猴类脑的一系列类型中存在着一种真正的构造上的间隙,而这种间隙并不存在于人类和类人猿之间,而是存在于较低等的和最低等的猿猴类之间,换句话说,即存在于旧世界的和新世界的猿类、猴类和狐猴类之间。每一种已经被检查过的狐猴的小脑实际上都是可以部分地从上面看得见的;它的后叶及其所包含的后角(posterior cornu)和小海马体或多或少都是残迹的。相反,每一种狨、美洲猴、旧世界猴、狒狒或类人猿的小脑在其后方为大脑叶所遮蔽,都是完全隐匿不露的,并且有一个大型后角以及一个十分发育的小海马体。”
这一叙述是根据当时已知的严格准确的有关记载作出的;而且在我看来,此后虽然发现合趾猿(siamang)和吼猴(howling monkey)的小脑后叶相对地不甚发育,但这一叙述显然并未因此而减弱其力量。尽管这两个物种的小脑后叶例外地短小,还没有任何人会以此为借口说,它们的脑以最轻微的程度接近狐猴类的脑。如果不把狨类置于它的自然位置之外,像比肖夫教授最令人不解地所做的那样,则可把他所选用的那些动物系列排写如下:人类,猩猩,大猩猩,长臂猿,天狗猴,鼯猴,长尾猴,猕猴,卷尾猴,绢毛猴,狨,狐猴,懒猴,我敢再次重申,在这一系列中,狨和狐猴之间的间隙最大,这一间隙比这一系列中任何其他二类之间的间隙都大得多。比肖夫教授忽视了在他撰写该文很久以前葛拉条雷就提出的一个事实,即,完全根据大脑的性状就可以把狐猴类分出于灵长目之外;弗劳尔教授在描述爪哇懒猴的过程中曾做过如下观察: (466)
“特别值得注意的是,有些猴类、即阔鼻猴类群的较低成员,普通被认为在后叶以外的其他方面都同大脑半球态短的狐猴类相接近,就后叶的发育来说,其中没有一种猴类是同狐猴类接近的。”
过去十多年来如此众多的研究者所做的研究大大地增进了我们的知识,就成熟的脑的构造而言,这些知识充分证明了我在1863年所做的叙述。据说,即使承认人类和猿类的成熟的脑是彼此相似的,但其实它们之间的差异还是重大的,这是因为它们在发育方式上表现了根本的差异。如果这等发育的根本差异确实存在的话,恐怕我比任何人都乐意承认这一论点的力量。但我否认确有这等根本差异存在。相反,人类和猿类的脑在发育上却是根本一致的。
引起葛拉条雷作出有关人类和猿类的脑在发育上存在根本差异的论述;在于他认为:在猿类中,脑沟最先出现在大脑半球的后区,而在人类胎儿中,脑沟最初出现在脑的前叶 (467) 。
这一论述总的是以两种观察为基础的,其一,有一只快要生产的长臂猿,其后部脑回十分发达,而前叶的脑回则“几乎看不见” (468) (原著,39页);其二,有一个怀孕22个或23个星期的人类胎儿,它的脑岛还没有被覆盖起来,尽管如此,“脑前叶仍有齿痕,一种不很深的裂沟显示着脑后叶的分离,其沟甚浅,与其发育期相应。而脑之其余表面则处于完全平滑状态”。
该书第二图版的1、2、3图只示明了这个大脑半球的上面、侧面和下面,而未示明其内面。值得注意的是,该图决不能证明葛拉条雷的描述,因为大脑半球后半部表面上的裂沟(前颞)比前半部所模糊显示的任何裂沟都更显著。如果该图是正确的话,则决不能证明葛拉条雷的结论是正确的。他的结论是:“在美发猴和长臂猿的脑同人类胎儿的脑之间,存在着根本的差异,即人类的颞颥脑沟未出现之前,额部脑沟久已存在。”
然而,自从葛拉条雷时代以来,关于脑回和脑沟的发育,已由施密特(Schmidt)、比肖夫、潘施(Pansch) (469) 等人重新进行了研究,尤其埃克尔的研究 (470) 不仅是最近完成的,而且是迄今最完善的。
他们研究的最后结果可总结如下:
1.人类胎儿的西耳维厄斯氏裂(Sylvian fissure)是在怀孕的第三个月内形成的。在第三个月和第四个月,大脑半球平滑而圆(西耳维厄斯氏凹除外),远远向后突出于小脑之外。
2.所谓真正的脑沟是在胎儿的第四个月末到第六个月初这段时间内才开始出现,但埃克尔慎重地指出,不仅它们的出现时期,而且它们的出现次序都有相当的个体变异。然而,无论额部脑沟或颞颥脑沟都不是最早出现的。
事实上最早出现的是在大脑半球的内面(无疑,葛拉条雷似乎没有检查过胎儿的内面,所以忽略了这一点),无论内垂直脑沟(即顶枕脑沟)或小海马脑沟都是这样,此二者密切接近,终于合二而一。通常顶枕脑沟(occipito-parietal)在二者之中较先出现。
3.在上述期间较晚阶段,另一种脑沟,即“顶后脑沟”(posterio-parietal)或罗兰德氏裂(Fissure of Rolando)发育了,继此,至胎儿的六个月期间内,其他额叶、颅顶叶、颞颥叶和枕叶的重要脑沟发育了。然而,还没有明显的证据可以证明,其中某一种脑沟永远出现在另一种脑沟之前;而且值得注意的是,埃克尔所描述的和绘制的这一时期的脑(见原著212—213页,第二图版,1、2、3、4图)示明,作为猿脑的显著特征的前颞颥脑沟(平行裂Scissure parallèle),其发育如果不比罗兰德氏裂更好,至少也是一样的,前颞颥脑沟比正常的额部脑沟要显著得多。
根据现今已知的事实,我以为人类胎儿的脑沟和脑回的出现次序,同进化的一般原理以及人类是从某种与猿相似的类型进化而来的观点完全符合;虽然那种与猿相似的类型在许多方面同现今生存的灵长类任何成员都不相同。
50年前,冯·贝尔教导我们说,亲缘关系相近的诸动物在其发育过程中最初带有它们所属的那较大类群的性状,以后逐渐地呈现它们那一科、属和种所专有的那些性状;同时他也证明了,一种高等动物的任何发育阶段都不会同任何低等动物的成熟状态确切相似。可以十分正确地说,一只蛙曾经通过一条鱼的状态,因为在其生命的某一时期,蝌蚪具有一条鱼的所有性状,如果它不再进一步发育,势必要被列入鱼类之中。但同等正确的是,一个蝌蚪同任何已知的鱼都很不相同。
同样地,五个月人类胎儿的脑可以被正确地说成不仅是一种猿的脑,而且可以说成是一种钩爪类(Arctopithecine) (471) 的或一种同狨相似的猿的脑;这是因为它的大脑半球具有大的后叶,而且除了西耳维厄斯氏脑沟和小海马脑沟外,并无其他任何脑沟,这种特性只有在灵长目的钩爪类中才能找到。但同等正确的是,正如葛拉条雷所说的,在其宽阔的西耳维厄斯氏裂方面,它同任何真正的狨都不相同。毫无疑问,它同一只狨的令期较大的胎儿的脑就要相似得多。但我们对狨类脑的发育情况却一无所知。关于阔鼻猴类(Platyrhini),我所知道的唯一观察是由潘施做的,他发现一只卷尾猴(Cebus apella)胎儿的脑除了西耳维厄斯氏裂和深的小海马裂之外,仅有一个很浅的前颞颥裂(即葛拉条雷所谓的“平行裂”)。
像松鼠猴(Saimiri)那样的阔鼻猴类具有前颞颥脑沟,它们仅在大脑半球的外部前一半表现有脑沟的残迹,或根本全无,这一情况以及上述事实毫无疑问为葛拉条雷的假说提供了良好的证据,即,他认为阔鼻猴类的后脑沟出现在前脑沟之前。但是,决不能因此就把适用于阔鼻猴类的规律扩充到狭鼻猴类。关于犬猿类的脑,我们还没有掌握任何材料;关于类人猿,除了上述有关即以诞生的长臂猿的脑以外,并无其他记载。现在,没有一点证据可以阐明黑猩猩的或猩猩的脑沟的出现次序和人类的不同。
葛拉条雷以如下格言开始了他的序文:“在科学上急于下结论,极为危险。”我恐怕他在其著作中讨论人类和猿类的差异时一定忘记了这一正确的格言。毫无疑问,这位优秀的作家对于正确理解哺乳动物的脑还是作出了前所未有的最卓越贡献,如果他活到今天,从这方面研究的进展得到益处,他大概会首先承认他的研究数据是不够充分的。不幸的是,他的结论被那些不懂得其基本原则的人们用来作为支持愚昧主义的论据了 (472) 。
但重要的是应该指出,葛拉条雷在其关于颞颥脑沟或额部脑沟相对出现次序的假说中无论是对还是错,事实仍然是存在的,即,在颞颥脑沟或额部脑沟出现之前,人类胎儿的脑所呈现的性状只有在灵长目的最低等类群中(狐猴类除外)才能找到;如果人类从某一类型逐渐变化而成,而这一祖先类型同其他灵长类所来自的类型正好相同,那恰恰是我们所期待的确应如此的情况。

————————————————————
(1) 《人类大脑的裂纹》(Grosshirnwindungen des Menschen),1868年,96页。这位作者以及格拉条雷(Gratiolet)和艾比(Aeby)的关于脑的结论,均经赫胥黎教授讨论过,见本版序言中提及的那篇附录。
(2) 《生理学讲义》(Lec.sur la Phys),1866年,890页,达利(M.Dally)在《灵长目与进化论》(L'Ordre des Primates et le Transformisme,1868年,29页)中加以引用。
(3) 即狂犬病。——译者注
(4) 林赛(W.L.Lindsay)博士在《心理学杂志》(Journal of Mental Science),1871年7月;以及《爱丁堡兽医评论》(Edinburgh Veterinary.Review),1858年7月,相当详细地讨论了这个问题。
(5) 一位评论家非常激烈而轻蔑地批评了我在这里所说的[《不列颠每季评论》(British Quarterly Review),1871年10月1日,472页],但我并没有用“相等”这个字眼,我看不出我有多大错误。在我看来,两种不同动物因感染同样疾病而产生同样的或密切相似的结果,与对两种不同液体用同样化学试剂所进行的测定,是非常近似的。
(6) 《巴拉圭哺乳动物志》(Naturgeschichteder S ugethiere von Paraguay),1830年,50页。
ugethiere von Paraguay),1830年,50页。
(7) 有些在等级上低得多的动物也有同样的嗜好。尼科尔斯(A.Nicols)先生告诉我说,他在澳大利亚的昆士兰养了三只灰色袋兔(Phaseolarctus cinereus),一点也没有教过它们,就对朗姆酒和吸烟有强烈的嗜好。
(8) 布雷姆,《动物生活》(Thierleben),第1卷,1864年,75,86页。关于美洲蛛猴(Ateles),参阅105页。关于其他相似记载,见25,107页。
(9) 林赛博士,《爱丁堡兽医评论》,1858年7月,13页。
(10) 关于昆虫,参阅莱科克(Laycock)博士的《生命周期性的一般法则》,原载《不列颠学会会报》,1842年。麦卡洛克(Macculloeh)曾见到一只狗患间日疟,见《西利曼北美科学杂志》(Silliman's North American Journal of Science),第17卷,305页。以后我还要重谈这个问题。
(11) 关于这一点,我曾在我著的《动物和植物在家养下的变异》(Variation of Animals and Plants under Domestication)第2卷,15页举出过证据,此外还可补充更多的证据。
(12) 许多种类的雄性猿猴确能分别人类的男女,最初凭嗅觉,其次凭外貌。尤雅特(Youatt)先生在伦敦动物园长期从事兽医工作,是最细心、最敏锐的观察家,他曾为我证实此事,动物园的其他饲养员和管理员的说法也相同。安德鲁·史密斯和布雷姆说,狒狒也能如此。权威人士居维叶也多次谈过此事。我以为人类和猿类所共有的现象,其恶劣程度莫有过于此者。有人说,狒狒见到妇女就发狂,但并非见到所有妇女都如此,它能从众人中识别年幼的妇女,以奇特的声音和容态召唤之。
(13) 这是小圣·伊莱尔(Isidore Geoffroy St.Hilaire)和弗·居维叶(F.Cuvier)对犬面狒狒(鼯猴)和类人猿所做的论述,见《哺乳动物志》(Hist.Nat.des Mammifères),第1卷,1824年。
(14) 赫胥黎,《人类在自然界的位置》(Man's Place in Nature),1863年,34页。
(15) 《人类在自然界的位置》,1863年,67页。
(16) 人的胚胎图(图1)采自埃克(Ecker)的《生理图解》(Icones Phys.),1851—1859年,表ⅩⅩⅩ,图2。这个胚胎的长度为0.833英寸,所以绘图时放大很多。狗的胚胎图采自比肖夫的《狗卵发育史》(Entwicklungsgeschichte des Hunde-Eies),1845年,表Ⅺ,图42B。这幅图放大五倍,胎龄25天。两幅图中内脏皆略去,子宫附属物亦从略。我所以刊登这两幅图,是受赫胥黎教授的《人类在自然界的位置》一书所启发。赫克尔在他的《自然创造史》中也登过相似的图。
(17) 怀曼(Wyman)教授,《美国科学院院报》(Proc.of American Acad.of Science),第4卷,1860年,17页。
(18) 欧文(Owen),《脊椎动物解剖学》(Anatomy of Vertebrates),第1卷,533页。
(19) 《关于人类的大部分脑旋图》(DieGrosshirnwindungen des Menschen),1868年,95页。
(20) 《脊椎动物解剖学》,第2卷,553页。
(21) 《博物学会会刊》(Proc.Soc.Nat.Hist.),波士顿,第9卷,1863年,185页。
(22) 《人类在自然界的位置》,65页。
(23) 当我读到卡内斯垂尼(Canestrini)的一篇有价值的论文——《原始人类的残迹器官的特征》(Caratteri rudimentali in ordine all'origine dell'uomo),原载《摩德纳自然科学协会年报》(Annuario della Soc.d.Nat.,Modena),1867年,81页——之前,我已经写完了这一章的草稿;我从此文中受益颇多。赫克尔在《普通形态学》和《自然创造史》两书中以“无目的论”(Dysteleology)的标题对这个问题进行了可称赞的全面讨论。
(24) 莫利(Murie)和米伐特(Mivart)两位先生对这个问题做过一些很好的评论,见《动物学会学报》(Transact.Zoolog.Soc.),第7卷,1869年,92页。
(25) 《动物和植物在家养下的变异》,第2卷,317,397页;《物种起源》,第五版,535页。
(26) 例如,理查德(M.Richard)描述和图示他称为“手上足肌”(muscle pédieux de la main)的残迹状态,他说它有时是“非常微细”的。叫做“后胫肌”(1e tibial postérieur)的肌肉一般在手中完全缺无,但它却时时以或多或少的残迹状态出现。
(27) 特纳教授,《爱丁堡皇家学会会刊》(Proc.RoyalSoc.Edinburgh),1866—1867年,65页。
(28) 参阅《人类和动物的表情》(Expression of the Emotions in Man and Animals),1872年,144页。
(29) 卡内斯垂尼引用海塔尔(Hyrtl)的材料,表明有同样情况,见《摩德纳自然科学协会会报》,1867年,97页。
(30) 皇家学会会员托因比,《耳病》(The Diseases oftheEar),1860年,12页;卓越的生理学家普瑞尔(Preyer)教授告诉我说,他最近对外耳的机能作了试验,得出与此相似的结论。
(31) 麦卡利斯特(A.Macalister)教授,《博物学年刊杂志》(Annals and Mag.of Nat.History),第7卷,1871年,342页。
(32) 米伐特,《基础解剖学》(Elementary Anatomy),1873年,396页。
(33) 参阅莫利和米伐特两位先生的优秀论文,其中附有狐猴亚目(Lemuroidea)的耳的绘图,见《动物学会会报》,第7卷,1869年,6,90页。
(34) 《关于达尔文所论的尖耳》(Ueber das Darwin'sche Spitzohr),见《解剖学和生理学文献集》(Archiv für Path.Anat.und Phys.),1871年,485页。
(35) 《人类和动物的表情》,136页。
(36) 参阅米勒(Müller)的《生理学原理》(Elements of Physiology),英译本,第2卷,1842年,1117页。欧文(Owen),《脊椎动物解剖学》,第3卷,260页。关于海象,《动物学会会报》,1854年11月8日。再参阅诺克斯(Knox)的《伟大的艺术家和解剖学家》(Great Artists and Anatomists),106页。这一残迹物在黑人和大洋洲人中比在欧洲人中显然多少大一点,参阅卡尔·沃格特的《人类讲义》(Lectures on Man),英译本,129页。
(37) 洪堡(Humboldt)关于南美土人所具有的嗅觉能力的记载是众所周知的,而且已被其他人士所证实。乌泽(M.Houzeau)断言,黑人和印第安人能在黑暗中根据气味来认人,见《心理能力的研究》(Etudessur les FacultésMentales),第1卷,1872年,91页。奥格尔(W.Ogle)博士就嗅觉与嗅区(Olfactory region)黏膜的以及皮肤的色素物质之间的关系作了一些奇妙的观察,据此我才谈到黑色人种的嗅觉优于白色人种,参阅他的论文,《外科学学报》(Medico Chirurgical Transactions),伦敦,第53卷,1870年,276页。
(38) 《心理的生理学和病理学》(The Physiology and Pathology of Mind),第2版,1868年,134页。
(39) 埃舍里希特(Eschricht),《论人类身体的无毛》(Ueber die Richtung der Haare am menschlichen K rper),见米勒的《解剖学和生理学文献集》(Miiller's Archiv für Anat.und Phys.),1837年,47页。以后我将常常引用这篇非常奇妙的论文。
rper),见米勒的《解剖学和生理学文献集》(Miiller's Archiv für Anat.und Phys.),1837年,47页。以后我将常常引用这篇非常奇妙的论文。
(40) 佩吉特,《外科病理学讲义》(Lectures on Surgical Pathology),第1卷,1853年,71页。
(42) 参阅《动物和植物在家养下的变异》,第2卷,327页。勃兰特教授最近寄给我另一件事例:有一父及其子,生于俄国,具有这等特性。我从巴黎收到这两个人的画像。
(43) 韦布(Webb)博士,《人类和类人猿的齿》(Teeth in Man and the Anthropoid Apes),卡特·布莱克(Carter Blake)博士曾予引用,见《人类学评论》(Anthro pological Review),1867年7月,299页。
(44) 欧文,《脊椎动物解剖学》,第3卷,320,321,325页。
(45) 《关于头骨的原始形态》(On the Primitive Form of the Skull),英译本,见《人类学评论》,1868年10月,426页。
(46) 蒙特加沙(Montegazza)从佛罗伦萨写信告诉我,他最近对不同人类种族的最后一个臼齿进行了研究,他得出同本书一样的结论,即,在高等的或文明的诸种族中,这个臼齿正走向萎缩和消灭的途中。
(47) 树袋熊为一种貌似小熊的无尾动物,即Phascolarctos cinereus,栖于树上,澳大利亚产。——译者注
(48) 欧文,《脊椎动物解剖学》,第3卷,416,434,441页。
(49) 《摩德纳自然科学协会年报》,1867年,94页。
(50) 马丁斯(M.C.Martins),《论生物界的一致性》(Del'Unité Organique),见《两个世界评论》(Revue des Deux Mondes),1862年6月15日,16页。赫克尔,《普通形态学》,第2卷,278页,他们二人都曾谈到这一残迹物有时会引起死亡的奇特事实。
(51) 关于它的遗传性,参阅斯特拉瑟斯博士的论文,见Lancet医学杂志,1873年2月15日,以及另一篇重要论文,见同杂志,1863年1月24日,83页。我听说诺克斯博士是注意人类这一特殊构造的第一位解剖学者,参阅他的《伟大的艺术家和解剖学家》,63页。再参阅格鲁勃(Gruber)博士的有关这一骨突的重要论文,见《圣彼得堡皇家学会会刊》(Bullelinde l'Acad.Imp.de St.Pétersbourg),第12卷,1867年,448页。
(52) 米伐特先生,《科学协会会报》(Transact.Phil.Soc.),1867年,310页。
(53) 《关于直布罗陀的洞穴》(On the Caves of Gibraltar),见《史前考古学国际会议报告书》(Transact.Internat.Congress of Prehist.Arch.),第三届会议,1869年,159页。怀曼最近阐明,来自美国西部和佛罗里达古代坟墩中的人类遗骸具有此孔的,占31%。黑人常具此孔。
(54) 古石器时代的后半。——译者注
(55) 考垂费什(Quatrefages)最近搜集了有关这个问题的证据,见《科学报告评论》(Revue des Cours Scientifiques),1867—1868年,625页。弗莱施曼(Fleischmann)在1840年曾展出过一个人类婴儿的标本,具有一条包含一些椎体的尾,这种情形并不多见;这条尾曾被出席埃尔兰根(Erlangen)自然科学者大会的许多解剖学者们严密地检查过,见马歇尔(Marshall),《荷兰的动物学文献》(Niederl ndischen Archiv für Zoologie),1871年12月。
ndischen Archiv für Zoologie),1871年12月。
(56) 欧文,《关于四肢的性质》(On the Nature of Limbs),1849年,114页。
(57) 洛伊卡特,见托德编的《解剖学全书》(Todd's Cyclop.ofAnat.)。男人的这一器官只有三至六赖因(1ines,十二赖因为一英寸)长,但像如此众多的其他残迹部分那样,它在发育上以及其他性状上都是容易变异的。
(58) 关于这个问题,参阅欧文的著作,《脊椎动物解剖学》,第3卷,675,676,706页。
(59) 比昂科尼(Bianconi)在最近发表的一部附有精美雕版图的著作[《达尔文学说与生物独立创造论》(La Théorie Darwinienne et la création dite indépendante),1874年]中,力图阐明上述诸例以及其他各例中的同源构造可以依据与其用途相一致的机械原理得到充分的解释。他对这等构造如何美妙地适应其终极目的所做的阐明,实为他人所莫及;但我相信,这种适应性可以通过自然选择得到解释。在讨论蝙蝠的翼时,我以为他所提出的仅仅是一种形而上学的原理(借用奥古斯特·孔德的用语),即“保全此动物的哺乳性质”。他只在少数几个事例中讨论过残迹器官,而所说的仅是那些不完全呈残迹状态的部分,如猪和牛的小蹄,而这与实质性问题根本无关,他明确指出这等残迹器官对动物还有作用。不幸的是,他没有考虑过以下事例,如牛的永不穿出牙龈的小牙,雄四足动物的乳房,存在于密闭翅盖之下的某些甲虫的翅,各式各样花的雄蕊和雌蕊的痕迹,以及其他许许多多这等事例。虽然我十分赞扬比昂科尼教授的著作,但大多数自然科学者今天所持的信念,在我看来,是不可动摇的,即,仅仅根据适应原理,残迹构造是不能得到解释的。
(60) 古尔德(B.A.Gould)著,《关于美国士兵的军事学和人类学的统计之研究》(Investigations in Military and Anthropolog.Statistics of American Soldiers),1869年,256页。
(61) 关于美洲土著居民的头颅类型,参阅艾特肯·梅格斯(Aitken Meigs)的文章,见《费城科学院院报》(Proc.Acad.Nat.Sci.,Philadelphia),1868年5月。关于澳洲人,参阅赫胥黎的叙述,见莱伊尔的《人类的古远性》,1863年,87页。关于桑威奇群岛(即夏威夷群岛。——译者注)的居民,参阅怀曼(Wyman)教授的《头颅观察》(Observations on Crania),波士顿,1868年,18页。
(62) 奎因(R.Quain)著,《动脉解剖学》(Anatomy of the Arteries),前言,第1卷,1844年。
(63) 《爱丁堡皇家学会会报》(Transact.RoyalSoc.Edinburgh),第24卷,175,189页。
(64) 《皇家学会会报》(Proc.RoyalSoc.),1867年,544页;1868年,483,524页,以及以前的文章,1866年,229页。
(65) 《爱尔兰皇家科学院院报》(Proc.R.Irish.Academy),第10卷,1868年,141页。
(66) 《圣彼得堡科学院院报》(Act.Acad.St.Petersburg),第二部分,1778年,217页。
(67) 指美洲大陆。——译者注
(68) 布雷姆,《动物生活》(Thierleben),第1卷,58,87页。伦格尔,《巴拉圭的哺乳动物》(S ugethiere von Paraguay),57页。
ugethiere von Paraguay),57页。
(69) 《动物和植物在家养下的变异》,第2卷,第十二章。
(70) 《遗传的天才:关于它的法则及其推论结果的探究》(Hereditary Genius:an Inquiry into its Laws and Consequences),1869年。
(71) 贝茨(Bates)先生说[《亚马孙河上的博物学者》(The Naturalist on the Amazons),第2卷,1863年,159页],关于同一个南美部落的印第安人,“其头部形状,没有两个人是完全一样的;其中一人面形椭圆,相貌美好,而另一人则完全是蒙古人的样子,面颊宽阔而突起,鼻孔掀张,两眼斜视”。
(72) 布鲁曼巴哈(Blumenbach),《关于人类学的论文》(Treatises on Antropolog.),英译本,1865年,205页。
(73) 米特福德(Mitford)的《希腊史》(History of Greece),第1卷,282页。色诺芬(Xenophon,希腊哲学家、历史学家,公元前434?一前355。——译者注)所著的《回忆录》(Memorabilia)第2卷第四章中有一段也谈到,男人择妻应以孩子们的健壮和精力旺盛为目的,这是希腊人所公认的原则(赫尔牧师使我注意到这一段)。希腊诗人色奥格尼斯(Theognis)生活于公元前550年,他明显地看出选择如被谨慎地应用,对人类的改进是何等重要。他还看出财富往往会抑制性选择的适当作用,因而乃作诗如下:
克氏(Kurnus)养马牛,凡事依规则,
选种贵血统,避免劣与弱,
为了增收益,成本所不恤。
吾人婚配中,钱却为一切,
为了金钱故,男人娶其妻;
复为金钱故,女人嫁其夫,
恶棍与流氓,亦愿随君去;
财源大茂盛,子女择配时,
门当须户对,夸富其门族,
万事皆混杂,贵贱已无殊!
外貌与精神,退化斑驳多,
劝君莫惊异,起因至明白!
后果徒悲叹,吾种已低劣。
(74) 戈德隆,《论物种》(De l'Espèce),第2卷,第3册,1859年。夸垂费什,《人种的一致性》(Unité de l'Espèce Humaine),1861年。他的有关人类学的讲义,载于《科学报告评论》,1866—1868年。
(75) 《畸形组织志及其分类》(Hist.Gén.et Part.des Anomalies de l'Organisation),三卷本,第1卷,1832年。
(76) 在我的《动物和植物在家养下的变异》第2卷,第二十二章、二十三章中,对这些法则曾进行过充分讨论。杜兰德(M.J.P.Durand)最近(1868年)发表过一篇有价值的论文,《论环境的影响》(De l'In fluence des Milieux)。关于植物,他非常强调土壤的性质。
(77) 古尔德,同前书,1869年,93,107,126,131,134页。
(78) 关于波利尼西亚人,参阅普里查德(Prichard)的《人类体格史》(Physical Hist.of Mankind),第5卷,1847年,145,283页。再参阅戈德隆的《论物种》,第2卷,289页。居住在上恒河(Upper Ganges)和孟加拉(Bengal)的印度人在外貌上也有显著差异;参阅埃尔芬斯通(Elphinstone)的《印度史》,第1卷,324页。
(79) 《人类学会纪要》(Memoirs,Anthropolog.Soc.),第3卷,1867—1869年,561,565,567页。
(80) 布雷肯里奇(Brakenridge)博士,《特异素质理论》(Theory of Diathesis),见《医学时报》(Medical Times),1869年6月19日,7月17日。
(81) 在我的《动物和植物在家养下的变异》第2卷,297—300页中,为某些陈述给以根据;耶格尔(Jaeger)博士,《论骨的伸长生长》(Ueber das L ngenwachshthum der Knochen),《耶拿学报》(Jenaischen Zeitschrift),B.V,Heft.i。
ngenwachshthum der Knochen),《耶拿学报》(Jenaischen Zeitschrift),B.V,Heft.i。
(82) 古尔德,同前书,1869年,288页。
(83) 《巴拉圭的哺乳动物》(S ugethiere von Paraguay),1830年;4页。
ugethiere von Paraguay),1830年;4页。
(84) 《格陵兰史》(History of Greenland),英译本,第1卷,1767年,230页。
(85) 《近族通婚》(Intermarriage),亚历山大·沃克著(Alex.Walker),1838年,377页。
(86) 《动物和植物在家养下的变异》,第1卷,173页。
(87) 《生物学原理》(Principles of Biology),第1卷,455页。
(88) 佩吉特,《外科病理学讲义》,第2卷,1853年,209页。
(89) 海军士兵的视力不及陆军士军,是一个奇特而料想不到的事实。古尔德博士证明确系如此,《美国南北战争中的公共卫生报告》(Sanitary Memoirs of the War of the Rebellion,1869年,530页);他以海军士兵的视野“受到船身长度和桅杆高度的限制”来解释这种情形。
(90) 《动物和植物在家养下的变异》,第1卷,8页。
(91) 《巴拉圭的哺乳动物》,8,10页。我曾有良好的机会观察火地人的异常视力。再参阅劳伦斯(Lawrence)关于这个问题的意见,见《生理学讲义》,1822年,404页。吉拉德-托伊仑(M.Giraud-Teulon)最近搜集了大量有价值的证据来证明近视的原因是由于眼睛的过度疲劳。
(92) 普里查德,《人类体格史》,布鲁曼巴哈之说见该书第1卷,1851年,311页;帕拉斯的述说见该书第4卷,1844年,407页。
(93) 普里查德引用,见上书,第5卷,463页。
(94) 福布斯先生的有价值的论文现发表于《伦敦人种学会杂志》(Journal of the Ethnological Soc.of London),新辑,第2卷,1870年,193页。
(95) 维尔肯斯(Wilckens)博士最近发表一篇有趣味的论文,阐明生活于山岳地区的家养动物如何在其骨架上发生变异,见《农学周报》(Landwirthschaft.Wochenblatt),第十期,1869年。
(96) 《关于畸形小头的研究报告》(Mémoire sur les Microcéphales),1867年,50,125,169,171,184—198页。
(97) 莱科克博士总结了畜生般的白痴的特性,称他们为“野兽般的白痴”(theroid),见《心理科学杂志》(Journal of Mental Science),1863年7月。斯科特(Scott)博士常常观察到低能儿嗅闻食物,见《聋与哑》(The Deaf and Dumb)第2版,1870年,10页。关于这个问题以及白痴的多毛性,参阅莫兹利博士的《躯体和精神》(Body and Mind),1870年,46—51页。关于白痴的多毛性,皮内尔(Pine1)也曾举出过一个显著事例。
(98) 在《动物和植物在家养下的变异》(第2卷,57页)中,我把并非很罕见的妇女额外乳房的事例归因于返祖。由于这等附加的乳房一般都对称地位于胸部,乃使我认为这大概是一个正确的结论;特别是使我有此想法的还有一个事例,即,一个单独的有效乳房发生在一位妇女的腹股沟区(inguinal region),她是另一位具有多余乳房的妇女的女儿。但是,现在我还发现在其他部位发生的异位乳房(mammae erraticae)(参阅普瑞尔教授的《论生存斗争》,Prof.Preyer,Der Kampf um des Dasein,1869年,45页),如在背上、腋下和股上的这等乳房分泌的乳汁如此之多,以致可以把小孩养育起来。这样,附加乳房是由于返祖的可能性就大大削弱了;尽管如此,我依然认为这大概是正确的,因为两对乳房常常对称地位于胸部;关于这一点,我曾收到过几个事例的报告。众所周知,狐猴类正常有两对乳房位于胸部。关于男人生有一对以上的乳房(当然是痕迹的),曾经记载过五个事例;参阅《解剖学和生理杂志》(Journal of Anat,and Physiology),1872年,56页,其中有汉迪赛德(Handyside)博士举出的一个事例说,有两兄弟显示了这种特性;再参阅巴特尔斯(Bartels)博士的一篇论文,载于《里卡兹和鲍依斯-雷蒙的文献集》(Reichert'sand du Bois-Reymond,s Archiv.),1872年,304页。巴特尔斯博士所提的事例之一:一个男人生有5个乳房,其中一个居中,正好位于脐眼之上;梅克尔·冯·黑姆斯巴哈(Meckel von Hemsbach)以为上述事例可以由某些翼手类(Cheiroptera)的居中乳房得到阐明。总之,如果人类的早期祖先不具有一对以上的乳房,则人类男女决不会有附加的乳房发育起来;倘不如此,我们就要发生重大的疑问了。
在《动物和植物在家养下的变异》(第2卷,12页)中,经过很大踌躇我还把人类和各种动物常见的多趾畸形(polydactylism)归因于返祖。导致我有此想法的,部分是由于欧文教授叙述某些鱼鳍类(Ichthyopterygia)具有5个以上的趾,所以我设想它们保持了原始状态;不过格根鲍尔(Gegenbaur)对欧文的结论表示怀疑,《耶拿学报》(Jenaischen Zeischrift,第5卷,第3册,341页)。另一方面,按照冈瑟(Günther)博士晚近提出的意见:角齿鱼(Ceratodus)的鳍有一列骨为中轴,在其两侧生有分节的骨质鳍刺,通过返祖,一侧或两侧可能会重现6个或更多的趾,承认这一点似乎不致有很大困难。祖特文(Zouteveen)博士告诉我说,曾经记载过这样一个事例:一个男人生有24个手指和24个足趾!导致我作出“多余指的出现是由于返祖”的结论,主要是根据如下的事实:多余指不仅是强烈遗传的,而且如我那时所相信的,在截断后还有再生的能力,就像低等脊椎动物的正常趾在截断后的情形一样。但我在《动物和植物在家养下的变异》(第2版)那部书中曾经说明,为什么我对记载下来的这等事例很少信赖。尽管如此,值得注意的是,由于受到抑制的发育同返祖是两种关系极为密切的过程,所以处于胚胎状态或受到抑制状态的各种构造——如裂口盖、双叉子宫等,往往伴随着多指畸形。梅克尔和小圣·伊莱尔都曾极力主张这一点。但现今最安全的方针还是完全放弃以下的概念,即多余指的发育同返归人类某一低等构造的祖先有任何关系。
(99) 参阅法尔博士在《解剖学和生理学全书》(Cyclopaedia of Anatomy and Physiology),第5卷,1859年,642页。欧文,《脊椎动物解剖学》,第3卷,1868年,687页。特纳教授,《爱丁堡医学杂志》(Edinburgh Medical Journal),1865年2月。
(100) 《摩德纳自然科学协会年报》,1867年,83页。卡内斯垂尼关于这个问题曾摘录了各种权威著作。劳里拉德(Lauri11ard)说,因为他发现两片颧骨的形状、比例以及它们的接合,在几具人类尸体和某些猿类中完全相似,所以他不能把这等部分的这种安排视为偶然的。关于这同样的畸形,沙威奥提(Saviotti)博士发表过另一篇论文,载于《临床学通报》(Gazzetta delle Cliniche),都灵(Turin),1871年,他说,在成年人头骨上发现有分离痕迹的约为百分之二;他还说,突颚的头骨(并非雅利安种族的)较其他头骨更常发生这种情形。再参阅德勒伦则(G.Delorenzi)关于同一问题的著作,《颧骨畸形的三个新例》(Tre nuovi casi d'anomalia dell'osso malare,Torino)(都灵),1872年。还有,莫索利(E.Morselli)的《关于颧骨的罕见畸形》(Sopra una rara anomalia dell'osso malare,Modena)(摩德纳),1872年。关于这块骨的分离,格鲁勃最近写了一本小册子。我之所以举出这些参考书目,是因为有一位评论家没有任何根据毫无顾忌地对我的叙述表示了怀疑。
(101) 小圣·伊莱尔在他的《畸形志》(Hist.des Anomalies),第3卷,437页中举出了一系列的事例。一位评论家对我大加责备,说我没有讨论过见于记载的有关各种部分发育受到抑制的大量事例[《解剖学和生理学杂志》(Jour.of Anat.and Physio.),1871年,366页]。他说,按照我的理论,“一个器官在其发育中的每一个瞬变状态,不仅是为了达到某种目的的一种手段,而其本身也曾一度是一种目的”。在我看来,这种说法不一定对。为什么在发育早期发生的变异同返祖没有关系?而这等变异如果有任何一点用处,如缩短和简化发育过程,大概都会被保存下来并得到积累。还有,为什么有害的畸形,例如同以往生存状况没有任何关系的萎缩的或过度肥大的诸部分,不在早期以及成熟期发生?
(102) 《脊椎动物解剖学》(Anatomy of Vertebr),第3卷,1868年,323页。
(103) 《普通形态学》,1866年,第2卷,160页。
(104) 卡尔·福格特(Carl Vogt),《人类讲义》,英译本,1864年,151页。
(105) 卡特·布莱克(C.Carter Blake),《关于脑雷特人的一只颚》,见《人类学评论》,1867年,295页。沙夫豪森(Schaaffhausen),同前杂志,1868年,426页。
(106) 《表情解剖学》(The Anatomy of Expression),1844年,110,131页。
(107) 卡内斯垂尼教授引用,见《摩德纳自然科学协会年报》,1867年,90页。
(108) 任何人要是想知道人类的肌肉为何常常发生变异,而且终于变得同四手类动物的肌肉相似,都应该读一读这些论文。下列参考文献同我的著作中少数几点有关:《皇家学会会报》,第14卷,1865年,379—384页;第15卷,1866年,241,242页;第15卷,1867年,544页;第16卷,1868年,524页。我在这里可以补充一点,穆里博士(Dr.Murie)和米伐特先生在他们有关狐猴类的研究报告中曾阐明,这等动物——四手类的最低层成员——的某些肌肉非常容易变异。在狐猴类中,有一些肌肉渐次变得与等级更低的动物的构造相似。
(109) 再参阅麦卡利斯特教授的文章,见《爱尔兰皇家学会会报》(Proc.R.Irish Academy),第10卷,1868年,124页。
(110) 钱普尼斯(Champneys)先生,《解剖学和生理学杂志》(Jourhal of Anat.and Phys.),1871年11月,178页。
(111) 《解剖学和生理学杂志》,1872年5月,421页。
(112) 麦卡利斯特教授,《爱尔兰皇家学会会报》,121页。他曾把他的观察材料制成表,发现最常出现肌肉畸形的是在前臂,其次在面部,再次在足部,等等。
(113) 霍顿(Haughton)牧师举出一个有关人类拇指长屈肌(flexor pollicis longus)的显著事例之后,接着说道(《爱尔兰皇家学会会报》,1864年6月27日,715页):“这个显著的例子阐明,人类拇指以及其他手指的腱(tendon)的排列有时具有猕猴的特性,但这样一个事例究应被视为猕猴向上变为人,人向下变为猕猴,或者是一种先天的反常现象,我还不能说。”这位富有才华的解剖学者和进化论的强硬反对者竟会承认他的任何一个最初命题的可能性,听到这一点已经可以使人满足了。麦卡利斯特教授也曾描述过拇指屈肌以它们同四手类的同样肌肉的关系而引人注意(《爱尔兰皇家学会会报》,第10卷,1864年,138页)。
(114) 在本书第一版问世以后,伍德先生发表过一篇专题报告[《科学学报》(Phil.Transactions),1870年,83页],阐明人类的颈、肩、胸的肌肉变异。他在这里指出,这些肌肉是何等容易变异,而且这些肌肉又何等常常而密切地同低类动物的正常肌肉相似。他总结说:“如果我成功地阐明了人类尸体中所发生的比较重要的变异类型,以充分显著的方式显示了它们可以作为达尔文的返祖原理或遗传法则在解剖科学中的证据和例子,那就是达到我的目的了。”
(115) 这几段叙述的根据,见我的《动物和植物在家养下的变异》,第2卷,320—335页。
(116) 在我的《动物和植物在家养下的变异》,第2卷,第23章中对这整个问题进行了讨论。
(117) 参阅永远值得纪念的《人口论》(Essay on the Principle of Population),马尔萨斯牧师著,第1卷,1826年,6,517页。
(118) 《动物和植物在家养下的变异》,第2卷,111—113,163页。
(119) 塞奇威克(Sedgwick),《英国及国外外科学评论》(British and Foreign Medico-Chirurg Review),1863年7月,170页。
(120) 亨特著,《孟加拉农村年报》(The Annals of Rural Bengel),1868年,259页。
(121) 《原始婚姻》(Primitive Marriage),1865年。
(122) 一位作者在《旁观者》(Spectator,1871年3月21日,320页)对这一段进行了如下的批评:“达尔文先生被迫再倡导人类堕落的新学说。他阐明高等动物的本性远比未开化人种的习惯更为高尚,所以他所再倡导的学说实质上乃是一种正教类型的,对此他似乎并未觉察;作为一种科学假说,他提出这样的学说,即,暂时的,但长期延续的道德败坏的原因,乃是由于人类获得知识,未开化部落的腐败风俗,特别是婚姻表明了这一点。有一种犹太传说,谓人类的道德堕落是由于夺得知识,这是他的最高本性所禁止的,其说有过于此否?”
(123) 关于这种效果,参阅斯坦利·杰文斯(W.Stanley Jevons)的优秀记载,《根据达尔文学说的推论》(A Deduction from Darwin's“Theory”),载于《自然》(Nature),1869年,231页。
(124) 拉瑟姆(Latham),《人类及其迁徙》(Man and his Migrations),1851年,135页。
(125) 莫利和米伐特二位先生在其《狐猴类的解剖》(《动物学会会报》,第7卷,1869年,96—98页)一文中说道:“有些肌肉在分布上是如此不规则,以致无法恰当地把它们归入上述任何类群中。”这等肌肉甚至在同一个体的相对两侧也互不相同。
(126) 《自然选择的范围》(Limits of Natural Selection),见《北美评论》(North American Review),1870年10月,295页。
(127) 《每季评论》(Quarterly Review),1869年4月,392页。这个问题在华莱士先生的《对自然选择学说的贡献》(Contributions to the Theory of Natural Selection,1870年)中进行了更充分的讨论,本书所引用的一切论点均在该书重予发表。《人类随笔》(The Essay on Man)曾受到欧洲最著名的动物学者克拉帕雷德(Claparède)教授的巧妙批评,见《一般书目提要》(Bibliothèque Universelle),1870年6月。本书所引用的华莱士的话,将使每一个读过他的《从自然选择学说推论人种的起源》(The Origin of Human Races deduced from the Theory of Natural Selection)那篇著名论文的人感到惊异,该文最初发表于《人类学评论》(1864年5月,158页)。关于这篇论文,我不能不在这里引用卢伯克(Lubbock)爵士的最公正的评论[《史前时代》(Prehistoric Times),1865年,479页],他说,华莱士先生“以特有的无私精神把自然选择的概念无保留地归功于达尔文先生,虽然,众所周知,他独立地发现了这一概念,而且同时予以发表,即使他叙述得不如达尔文详尽”。
(128) 劳森·泰特(Lawson Tait)在他的《自然选择法则》(Law of Natural Selection)一文中引用,见《都柏林医学季刊》(Dublin Quarterly Journal of Medical Science),1869年2月。凯勒(Keller)也引述过同样的效果。
(129) “四手类”系一种旧的动物分类,除去人类以外的灵长类动物均包括在内;而两手类(Bimana)只包括人类。——译者注
(130) 欧文,《脊椎动物解剖学》,第3卷,71页。
(131) 《每季评论》,1869年4月,392页。
(132) 合趾长臂猿(Hylobates syndactylus),如这个名字所表示的,它的两个足趾固定地结合在一起;布赖茨(Blyth)先生告诉我说,敏捷长臂猿(H.agilis)、白手长臂猿(H.lar)、银灰长臂猿(H.leuciscus)的足趾有时也是如此。疣猴是严格树栖的,而且异常活泼(布雷姆,《动物生活》,第1卷,50页),但它是否比近缘属的物种更善于攀登,我还不知道。值得注意的是,世界上树栖性最强的动物——树懒(sloths)的脚与钩异常相似。
(133) 布雷姆《动物生活》,第1卷,86页。
(134) 《人手》(The Hand),见《布里奇沃特论文集》(Bridgewater Treatise),1833年,38页。
(135) 赫克尔(H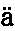 ckel)关于人类变为二足动物的步骤进行了精彩的讨论[《自然创造史》(Natiirliche Sch
ckel)关于人类变为二足动物的步骤进行了精彩的讨论[《自然创造史》(Natiirliche Sch pfungsgeschichte),1868年,507页]。比希纳博士关于人把脚用为把握器官举出了一些好例子[《达尔文学说讨论集》(Conférences sur la Theórie Darwinienne),1869年,135页];他还写过高等猿类的行进方式,我在下一节将提到;关于这个问题,再参阅欧文的《脊椎动物解剖学》,第3卷,71页。
pfungsgeschichte),1868年,507页]。比希纳博士关于人把脚用为把握器官举出了一些好例子[《达尔文学说讨论集》(Conférences sur la Theórie Darwinienne),1869年,135页];他还写过高等猿类的行进方式,我在下一节将提到;关于这个问题,再参阅欧文的《脊椎动物解剖学》,第3卷,71页。
(136) 布罗卡(Broca)教授,《尾椎的构造》(La Constitution des Vertèbres caudales),见《人类学评论》(La Revue d'Anthropologie),1872年,26页(单行本)。
(137) 《论头骨的原始形态》(On the Primitive Form of the Skull),译文见《人类学评论》(Anthropological Review),1868年10月,428页。欧文论高等猿类的乳头状突起,《脊椎动物解剖学》,第2卷,1866年,551页。
(138) 《动物界的边际;用达尔文学说进行的观察》(Die Grenzen der Thierwelt,eine Betrachtung zu Darwin's Lehre),1868年,51页。
(139) 迪雅尔丹(Dujardin),《自然科学年刊》(Annales des Sc.Nat.),第14卷,第3辑,动物部分,1850年,203页。再参阅洛恩(Lowne)先生的《一种蝇(Musca vomitoria)的解剖及其生理》,1870年,14页。我的儿子F.达尔文为我解剖了红褐林蚁(Formica rufa)的脑神经节。
(140) 《科学学报》(Philosophical Transactions),1869年,513页。
(141) 《关于选择》(Les Sélections),布罗卡,见《人类学评论》,1873年;再参阅沃格特的《人类讲义》(Lectures on Man),英译本,1864年,88,90页。普里查德,《人类体格史》(Phys.Hiot.of Mankind),第1卷,1838年,305页。
(142) 更新世晚期,旧石器时代中期的“古人”,分布在欧洲、北非、西亚一带。——译者注
(143) 在刚才提到的那篇有趣的文章中,布罗卡教授很好地谈论了:在文明民族中由于保存了相当数量的身心皆弱的个人,其头骨的平均体积一定要降低,这些人如在未开化状态下,将会立刻被淘汰。另一方面,在未开化人中这个平均数仅包括那些在极其艰苦生活条件下能够生存的富有才能的个人。于是布罗卡说明了一个没有其他方法可以说明的事实,即洛泽尔(Lozère)的史前穴居人的头骨平均容积为什么要比近代法国人的为大。
(144) 《法兰西科学报告》(Comptes-rendus des Sciences),1868年6月1日。
(145) 《动物和植物在家养下的变异》,第1卷,124—129页。
(146) 沙夫豪森所举的有关痉挛和疤痕的例子,系根据布鲁曼巴哈(Blumenbach)和布施(Busch)的材料,见《人类学评论》,1868年10月,420页。贾洛得(Jarrod)博士所举的有关头骨由于头的部位不正而发生变异的例子,系根据坎波尔(Camper)和他自己的观察,见《人类学》(Anthropologia),1808年,115,116页。他相信某些行业的人,如鞋匠,由于头部经常向前倾,前额变得较圆而且凸出。
(147) 《动物和植物在家养下的变异》,第1卷,117页,论述头骨的变长;119页,论述一只垂耳的效果。
(148) 沙夫豪森引用,见《人类学评论》,1868年10月,419页。
(149) 欧文,《脊椎动物解剖学》,第3卷,619页。
(150) 小圣·伊莱尔谈到人类的头部被有长发[《自然史通论》(Hist.Nat.Générale),第2卷,215—217页],还谈到猴类和其他哺乳动物的朝上表面比朝下表面具有较厚的毛。不同的作者同样也观察到这一点。然而热尔韦兹(P.Gervais)教授却说,大猩猩的背部却比朝下部分的毛稀,这部分是由于被摩擦掉了。
(151) 《博物学家在尼加拉瓜》(Naturalist in Nicaragua),1874年,209页。我引用的下述丹尼生(W.Denison)爵士所写的一节,是同贝尔特先生的观点一致的,“据说澳洲人有一种习惯:当蚤虱来找麻烦的时候,就用微火灼烧自己”。
(152) 圣乔治·米伐特先生,《动物学会会报》,1865年,562,583页。格雷(J.E.Gray)博士,《大英博物馆目录:骨骼部分》(Cat.Brit.Mus.)。欧文,《脊椎动物解剖学》,第2卷,517页。小圣·伊莱尔,《自然史通论》,第2卷,244页。
(153) 《人类学评论》,1872年;《尾椎的构造》(La Constitution des Vertèbres caudales)。
(154) 《动物学会会报》,1872年,2l0页。
(155) 1“赖因”为1/12英寸。——译者注
(156) 《动物学会会报》,1872年,786页。
(157) 我所指的是布朗-西奎(Brown-Séquard)对豚鼠在施行手术后所发生的癫痫症的遗传效果以及最近对切断颈部交感神经的相似效果所进行的观察。今后我还有机会提到沙尔文(Salvin)先生所举的有趣事例,即摩摩鸟(motmots)自己咬去其尾羽的遗传效果。关于这个问题的一般论述,参阅《动物和植物在家养下的变异》,第2卷,22—24页。
(158) 《动物和植物在家养下的变异》,第2卷,280,282页。
(159) 《原始人类》(Primeval Man),1869年,66页。
(160) 南非卡拉哈里沙漠地区一个游牧部族。——译者注
(161) 加里曼丹(Kalimantan)的旧称,为亚洲一大岛。——译者注
(162) 关于这几方面的证据,参阅卢伯克的《史前时代》,354页等。
(163) Henry Howard,英国诗人,1517?—1547。Thomas Clarkson,英国人,奴隶废除主义者,1760—1846。——译者注
(164) Isaac Newton,英国自然科学家,1642—1727。William Shakspeare,英国诗人,戏剧家,1564—1616。——译者注
(165) 《关于昆虫的本能》(L'Instinct chez les Insectes),见《两个世界评论》(Revue des Deux Mondes),1870年2月,690页。
(166) 《美洲河狸及其行为》(The American Beaver and His Works),1868年。
(167) 《心理学原理》(The Principles of Psychology),第2版,1870年,418—443页。
(168) 《对自然选择理论的贡献》(Contributions to the Theory of Natural Selection),1870年,212页。
(169) 河狸巧于筑巢,常在巢外筑堤为堰贮水,以防敌袭。筑巢所用的材料主要为木枝、黏土和砾石,入林采取木枝及搬运方法亦非常巧妙。——译者注
(170) 关于这个问题的证据,参阅摩格芮芝(Moggridges)先生的有趣著作,《农蚁和螲蟷》(Harvesting Ants and Trap-door Spiders),1873年,126,128页。
(171) 《蚁类习性的研究》(Recherches sur les Maeurs des Fourmis),1810年,173页。
(172) 所有以下根据这两位博物学家所做的叙述,均引自伦格尔的《巴拉圭哺乳动物志》(Naturgesch.der S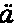 ugethiere von Paraguay),1830年,41—57页;以及布雷姆的《动物生活》,第1卷,10—87页。
ugethiere von Paraguay),1830年,41—57页;以及布雷姆的《动物生活》,第1卷,10—87页。
(173) 林赛博士(Dr.L.Lindsay)在他的《低于人类的动物精神生理学》(Physiology of Mind in the Lower Animals)一文中引用,见《心理学杂志》(Jour.of Mental Science),1871年4月,38页。
(174) 《布里奇沃特论文集》(Bridgewater Treatise),263页。
(175) 一位批评者毫无根据地对布雷姆所描述的这种行为的可能性提出质疑[《每季评论》(Quarterly Review),1871年7月,72页],这不过是为了攻击我的书而已。所以我自行试验,发现我能够容易地用我的牙把一只将近五周的小猫的小利爪咬住。
(176) 在我的《人类和动物的表情》(Expression of the Emotions)第43页简短谈到这种情形。
(177) 马丁(W.C.L.Martin),《哺乳动物志》(Nat.Hist.of Mammalia),1841年,405页。
(178) 贝特曼(Bateman)博士,《关于失语症》(On Aphasia),1870年,110页。
(179) 沃格特引用,《关于畸形小头的研究报告》,1867年,168页。
(180) 《动物和植物在家养下的变异》,第1卷,27页。
(181) 《自然科学年刊》(Annales des Sc.Nat.),第22卷,第1辑,397页。
(182) 《蚁类习性的研究》(LesMaeurs des Fourmis),1810年,150页。
(183) 莫兹利博士在其《精神的生理学和病理学》(Physiology and Pathology of Mind)一书中引用,1868年,19,220页。
(184) 杰尔登(Jerdon)博士,《印度鸟类》(Birds of India),第1卷,1862年,21页。乌泽说,他养的长尾小鹦鹉(parokeets)和金丝雀(canary-bird)会做梦:《动物的心理官能》(Facultés Mentales des Animaux),第2卷,136页。
(185) 《动物的心理官能》,第2卷,1872年,181页。
(186) 莫尔根先生的《美洲河狸》(The American Beaver)一书为这一叙述提供了一个良好例证。然而我不得不认为他过于低估了本能的能力。
(187) 《关于兽类的动作》(Die Bewegungen der Thiere),1873年,11页。
(188) 《动物的心理官能》,第2卷,1872年,265页。
(189) 赫胥黎非常清晰地分析了一个人和一只狗的心理等级,他作出的结论和我在本书中所提出的看法相似。参阅他的文章《批评达尔文先生的人们》(Mr.Darwin's Critics),见《当代评论》(Contemporary Review),1871年11月,462页,并见《评论及短论》(Critiques and Essays),1873年,279页。
(190) 贝尔特先生在他那部最有趣的著作《自然学者在尼加拉瓜》中同样也描述了一只驯服的卷尾猴的各种行为,我以为这明显地阐明了这种动物具有某种推理力。
(191) 《沼和湖》(The Moor and the Loch),45页。哈钦森上校,《狗的训练》(Dog Breaking),1850年,46页。
(192) 《个人记事》(Personal Narrative),英译本,第3卷,106页。
(193) 我高兴地看到像莱斯利·斯蒂芬(Leslie Stephen)先生那样敏锐的思想家当谈到人类和低等动物的心理之间那道假定的不可逾越的障壁时说道[见《达尔文主义和神学,自由思想论文集》(Darwinism and Divinity,Essays on Free-thinking),1873年,80页]:“诚然,划出这种区别所依据的根据在我们看来,并不比其他大量的形而上学的区别所依据的根据更好一点,这就如同说,因为你能给两种东西起不同的名字,所以它们一定有不同的性质。难于理解凡是曾经养过一只狗或者见过一头象的人,怎么还会怀疑动物实质上有可以完成推理过程的能力。”
(194) 参阅林赛博士的《动物的疯狂》(Madness in Animals),见《心理学杂志》,1871年7月。
(195) 莱伊尔(C.Lyell)爵士引用,《人类的源远流长》(Antiquity of Man),497页。
(196) 关于更多的详细证据,参阅乌泽的《论心理官能》(Les Facultés Mentales),第2卷,1872年,147页。
(197) 关于海洋岛上的鸟类,参阅我的《“贝格尔”号舰航海研究日志》(Journal of Researches during the voyage of the“Beagle”),1845年,398页。《物种起源》,第五版,260页。
(198) 《有关动物智力哲学的书信集》(Lettres Phil.surl'Intelligence des Animaux),新版,1802年,86页。
(199) 关于这个问题的证据,参阅《动物和植物在家养下的变异》,第1卷,第一章。
(200) 《动物学会会报》(Proc.Zoolog.Soc.),1864年,186页。
(201) 萨维奇(Savage)和怀曼(Wyman),《波士顿博物学杂志》(Boston Journal of Nat.Hist.),第4卷,1843—1844年,383页。
(202) 《巴拉圭哺乳动物志》,1830年,51—56页。
(203) 《印度原野》(IndianField),1871年,3月4日。
(204) 《动物生活》,第1卷,79,82页。
(205) 《马来群岛》(The Malay Archipelago),第1卷,1869年,87页。
(206) 《原始人类》(Primeval Man),1869年,145,147页。
(207) 《史前时代》(Prehistoric Times),1865年,473页等。
(208) 胡卡姆(Hookham)先生给马克斯·米勒(Max Müller)的一封信,见《伯明翰新闻》(Birmingham News),1873年5月。
(209) 一名“猛犬”,种类不少,大部分用以助猎。——译者注
(210) 《达尔文学说讨论会文集》(Conférences sur la Théorie Darwinienne),法文版,1869年,132页。
(211) 牧师麦卡恩(J.M'Cann)博士,《反对达尔文主义》(Anti-Darwinism),1869年,13页。
(212) 《人类学评论》(1864年,158页)引用。
(213) 伦格尔,同前书,45页。
(214) 参阅我写的《动物和植物在家养下的变异》,第1卷,27页。
(215) 《动物的心理能力》,第2卷,1872年,346—349页。
(216) 在泰勒(E.B.Tylor)先生的很有趣味的著作《对人类初期历史的研究》(Researches into the Early History of Mankind)中有关于这个问题的讨论,1865年,第2—4章。
(217) 关于这种效果,我曾收到几份详细报告。海军上将沙利文(B.J.Sulivan)爵士,据我所知是一位谨慎的观察家,他向我保证说,在他父亲家中长期饲养的一只非洲鹦鹉可以准确地叫出某些家人和客人的名字。在吃早饭的时候,它向每一个人说“早安”,在夜间它又向每一个离开那间屋子的人说“晚安”,从来没把这两句问候话弄颠倒过。对沙利文爵士的父亲,它惯常在“早安”之后还要加上一个短句,可是自从他父亲死后,它一次也没有重复说过这个短句。它猛烈地责骂一条从窗户蹿进屋去的生狗;它还责骂另一只鹦鹉,“你这顽皮的家伙”,当那只鹦鹉逃出鸟笼去吃厨案上的苹果的时候。关于这同样效果,再参阅乌泽的《动物的心理能力》,第2卷,309页,论鹦鹉。莫西科(A.Moschkau)博士告诉我说,他知道有一只欧椋鸟(starling)永远能够无误地用德语向来人说“早安”,向那些离去的人说:“再见,老朋友。”我还能再举出几个这样的事例。
(218) 参阅惠特尼(Whitney)教授关于这个问题的一些好意见,见他的著作《东方及其语言学的研究》(Oriental and Linguistic Studies),1873年,354页。他观察到人类彼此之间的愿望交流,乃是一种生活力,这种生活力对语言的发展“有意识地或者无意识地发生作用:就达到直接目的而言,是有意识的;就此种行为的进一步结果而言,则是无意识的”。
(219) 戴恩斯·巴林顿,《科学学报》,1773年,262页。再参阅马尔的文章,见《自然科学年刊》(Ann.des.Sc.Nat.),第10卷,第3辑,动物部分,119页。
(220) 《论语言的起源》(On the Origin of Language),韦奇伍德著,1866年。《语言问题》(Chapterson Language),法勒著,1865年。这是最有趣味的两本著作。再参阅阿尔贝·勒穆瓦纳(Albert Lemoine)著,《口头语的自然规律》(De la Phys.et de Parole),1865年,190页。已故的施莱歇尔教授关于这个问题的著作已被比克尔斯(Bikkers)博士译成英文,名为《受到语言学考验的达尔文主义》(Darwinism tested by the Science of Language),1869年。
(221) 沃格特,《关于畸形小头的研究报告》,1867年,169页。关于未开化人,我在《航海研究日志》(1869年)中举出过一些事实。
(222) 关于这个问题的明显证据,参阅经常引用的布雷姆和伦格尔的两本著作。
(223) 乌泽在他的《动物的心理能力》一书中,举出过他对这个问题所观察到的一项很奇妙的记载。
(224) 参阅莫兹利博士关于这个问题的意见,见《精神的生理学和病理学》,第2版,1868年,199页。
(225) 关于此事,曾记载过许多奇妙例子,参阅贝特曼的《关于失语症》,1870年,27,31,53,100页及其他。再参阅《关于智力的调查》(Inquires Concerning the Intellectual Powers),1838年,150页。
(226) 《动物和植物在家养下的变异》,第2卷,6页。
(227) 关于《达尔文先生的语言哲学》的讲演,1873年。
(228) 杰出的语言学家惠特尼对于这一点的评论远比我所能说的更为有力。当谈到布利克(Bleek)的观点时,他说道(见《东方及其语言学的研究》,1873年,297页),“因为语言广泛地是思想的必要辅助手段,思想赖此而发展,认识力赖此而达到清晰、丰富多彩和复杂化,以至对意识的充分掌握;所以不得不制造出没有语言就绝对不可能有思想的说法,把能力和它的工具等同起来。”他好像有道理地断言,人手如果没有工具就不能起作用。从这种教条出发,他就不能不陷入米勒的最恶劣的谬论,谓婴儿(不会说话的)不是人类,聋哑人没有学会用手指模仿说话以前不具理性。马克斯·米勒用斜体字标出下面的警句(“对于达尔文先生的语言哲学的讲演”,1873年,第三讲):“没有无语言之思想,也没有无思想之语言。”他在这里给思想这个词所下的定义是何等奇怪!
(229) 《自由思想论文集》,1873年,82页。
(230) 关于这一效果,参阅莫德斯雷的一些好议论,见《心理的生理学和病理学》,1868年,199页。
(231) 麦克吉利夫雷(Macgillivray),《大不列颠鸟类》(Hist.of British Birds),第2卷,1839年,29页。最优秀观察家布莱克瓦尔(Blackwall)说道,喜鹊(magpie)可以学会念出单字甚至短句,它们几乎比其他任何英国鸟都容易做到这一点;可是接着他又说,在长期周密地研究了它的习性之后,他从来没有发现它在自然状态下表现有任何模仿的异常能力。
(232) 莱伊尔爵士在《关于人类的源远流长的地质证据》(1863年,第二十三章)中指出,在语言发展和物种发展之间有很有趣的相似性。
(233) 关于这种效果,参阅法勒牧师在一篇题名《语言学和达尔文主义》(Philology and Darwinism)的论文中的意见,见《自然》(Nature),1870年3月24日,528页。
(234) 《自然》1870年1月6日,257页。
(235) 欧洲比利牛斯山西部地区古老居民的语言。——译者注
(236) 分布在挪威、瑞典、芬兰和苏联各国北部的拉普人的语言。——译者注
(237) 韦克(C.S.Wake)在《论人类》(Chapters on Man)101页引用。
(238) 巴克兰(Buckland),《布里奇沃特》,411页。
(239) 关于语言的简化,参阅卢伯克爵士的一些好议论,见《文化的起源》(Origin of Civilisation),1870年,278页。
(240) 《旁观者》(The Spectator),1869年12月4日,1430页。
(241) 关于这个问题,参阅法勒牧师所写的一篇最优秀的论文,见《人类学评论》,1864年8月,217页。关于进一步的事实,参阅卢伯克爵士的《史前时代》,第2版,1869年,564页;特别是《文化的起源》(1870年)有关宗教的篇章。
(242) 《对动物和植物的崇拜》(The Worship of Animals and Plants)见《双周评论》(Fortnightly Review),1869年10月1日,422页。
(243) 泰勒,《人类的早期历史》(Early History of Mankind),1865年,6页。再参阅卢伯克的《文化的起源》(1870年)中关于宗教发展那引人注目的三章。赫伯特·斯宾塞先生在《双周评论》(1870年5月1日,535页)的一篇有独创性的论文中以相似的方式说明了全世界宗教信仰的最初形式,谓人类通过梦境、形影以及其他原因的引导,把自己看成是双重的实体,即肉体的和灵魂的。由于设想死后灵魂还存在,而且富有威力,所以用各种祭品和仪式向它祈求赎罪和保佑。于是他进一步阐明,用某种动物或其他物品给一个部落的早期祖先或创始人所起的名字或绰号,经过长期以后就会被设想为代表这个部落的真实祖先:这个动物和物品自然地会被信为依然存在的灵魂,并且把它视为神圣,作为一位神而受到崇拜。尽管如此,我不能不猜想,还有一个更早的、更原始的阶段,以为那时任何显示有力量和运动的东西都被赋予了和我们自己近似的某种生命形态和心理官能。
(244) 参阅欧文·派克(L.Owen Pike)先生的一篇富有才智的文章,见《人类学评论》,1870年4月,63页。
(245) 《宗教、道德等与达尔文学说》(Religion,Moral,&c.,der Darwin'schen Art-Lehre),1869年,53页。据说(林赛博士,《心理学杂志》,1871年,43页),培根(Bacon)很久以前以及诗人伯恩斯(Burns)均持有同样见解。
(246) 《史前时代》,第2版,571页。在这部著作中,关于未开化人的变化无常的奇异风俗有最好的记载。
(247) 关于这个问题,参阅夸垂费什(Quatrefages)的《人种的一致性》,1861年,21页及其他。
(248) 《关于伦理学的论述》(Dissertation on Ethical Philosophy),1837年,231页及其他。
(249) 德国哲学家(1724—1804)。——译者注
(250) 《伦理的形而上学》(Metaphysics of Ethics),森普尔(J.W.Semple)译,爱丁堡,1836年,136页。
(251) 关于这个问题写过著作的,贝恩(Bain)先生列过一个20位英国作家的名单《心理学和道德学》(Mental and Moral Science),1868年,543—725页,他们的名字素为人人所熟悉:在这些人士中似乎还可以加入贝恩先生自己的名字,以及莱基(Lecky)先生、沙德沃思·霍奇森(Shadworth Hodgson)先生、卢伯克爵士以及另外几位的名字。
(252) 布罗代(B.Brodie)爵士在论述人类是一种社会性动物之后问道[《心理学探究》(Psychological Enquiries),1854年,192页]:“关于道德观念是否存在的问题的争论,应该由此得到解决吧?”许多人似乎都有过同样的看法,如古代的罗马皇帝兼哲学家玛卡斯·奥瑞利亚斯(Marcus Aurelius)就是其中一个。米尔(J.S.Mill)在其著名的著作《功利主义》(Utilitar ianism,1864年,45,46页)一书中说道:“社会感情是一种强有力的自然感情”而且是对功利主义道德的感情之自然基础”。他又说,“道德官能就像上述后天获得的智能那样,如果不是本性的一部分,也是从那里自然生长出来的:而且像它们那样,能够在一定微小程度上自然发生”。但是,同所有这种说法相反,他还指出:“据我所信,道德感情不是先天的,而是后天获得的,但并不因此而不是自然的。”对于如此渊博的一位思想家的看法,我大胆提出完全不同的意见,确有些踌躇,但几乎无可争辩的是,社会感情在低等动物中乃是本能的或先天的;那么,社会感情在人类中为什么不应如此呢?贝恩先生[例如,参阅《情绪与意志》(The Emotions and the Will),1865年,481页]以及其他人士相信,道德观念乃是每个人在其一生期间所获得的。根据进化的一般理论,至少这是极端不大可能的。在米尔先生的著作中对所有遗传的心理属性的忽视,我以为今后将被评价为最严重的缺点。
(253) 西奇威克(H.Sidgwick)对这个问题进行过很好的讨论(《科学院报告》,1872年,6月15日,231页。):“我们可以肯定,一只优良的蜜蜂大概渴望用比较温和的方法去解决种群数量问题。”然而,根据许多或大多数未开化的人的习惯来判断,人类是用杀害女婴、一妻多夫以及男女乱交来解决这个问题的,所以,可充分怀疑这是否为比较温和的方法。科比(Cobbe)女士对上述说法也进行过评论,说道[《达尔文主义在道德观上的应用》,Darwinism in Morals,见《神学评论》(Theological Review),1872年,4月,188—191页]:社会义务的原则将如此而被颠倒;我以为她所说的意思是,履行社会义务将危害个体;但她忽视了她必须承认的一个事实,即蜜蜂的这种本能被获得乃是为了群体的利益。她甚至说道,如果本章所提倡的伦理学原理确能被普遍接受,“我将不得不相信,其胜利之时,即为人类美德的丧钟敲响之日!”可以期望,众多人士对这个地球上人类美德永存的信念并不会如此短命。
(254) 《达尔文学说》(Die Darwin'sche Theorie),101页。
(255) 布朗(R.Brown)先生,《动物学会会报》,1868年,409页。
(256) 布雷姆,《动物生活》(Thierleben),第1卷,1864年,52,79页。关于猴彼此拔掉扎在身上的棘刺,参阅54页。关于树精狒狒翻动石头,是根据阿尔瓦雷斯(Alvarez)提出的证据(76页),布雷姆认为他的观察是十分可靠的。关于老雄狒狒攻击狗的例子,参阅79页;关于鹰的例子,56页。
(257) 贝尔特(Belt)先生举过一个尼加拉瓜的蛛猴例子,人们听到它在树林中大喊大叫差不多达两个小时之久,并且发现有一只鹰落在它的近旁。显然当它们面对面时,鹰不敢发动攻击;贝尔特先生根据他对这些猴的习性的观察,相信它们三两只聚在一起,防备鹰的攻击。《博物学者在尼加拉瓜》,1874年,118页。
(258) 《博物学年刊》(Annals of Mag.of Nat.Hist),1868年11月,382页。
(259) 卢伯克爵士,《史前时代》,第2版,446页。
(260) 莫尔根先生引用,《美洲河狸》(The Amer ican Beaver),1868年,272页。斯坦斯伯里还做过一个有趣记载:一只很小的鹈鹕被激流冲跑,有六只老鹈鹕从旁鼓励它游向岸边。
(261) 贝恩先生述说,“从适当的同情心可以产生对于一个受难者给予有效的帮助”,《心理学与道德学》,1868年,245页。
(262) 《物种的分类》(Thierleben),第1卷,85页。
(263) 《物种的分类》,1869年,97页。
(264) 《关于达尔文学说》,1869年,54页。
(265) 再参阅胡克的《喜马拉雅旅行记》(Himalayan Journals),第2卷,1854年,333页。
(266) 布雷姆,《动物生活》,第1卷,76页。
(267) 参阅他的一篇极有趣的论文:《牛类和人类的群居生活》(Gregariousness in Cattle,and in Man),见《麦克米伦杂志》(Macmillan's Mag.),1871年2月,353页。
(268) 参阅亚当·史密斯的《关于道德感的理论》(Theory of Moral Sentiments)一书的引人注目的第一章。再参阅贝恩的《心理学和道德学》,1868年,244页,以及275—282页。贝恩先生说道:“同情乃是间接地使同情者感到愉快的一个源泉”;他通过互易性(reciprocity)来解释这个问题。他又说,“受到恩惠的人或代替他的其他人,当以同情和有力的帮助作为报答以补偿对方所作出的一切牺牲。但是,同情如果严格地是一种本能——看来似乎就是如此,那么它的行使就会给人以直接愉快,其方式正如上述行使差不多每一种其他本能的情形一样。”
(269) 詹尼斯(L.Jenyns)牧师说,这一事实最初是由杰出的詹纳(Jenner)记载的,见《科学学报》(Phil.Transact.)1824年,此后又为几位观察家、特别是布莱克瓦尔所证实。后面这位细心的观察家连续两年在晚秋检查了36个鸟巢;他发现,12个鸟巢有死去的幼鸟,5个鸟巢有即将孵化的卵,3个鸟巢有接近孵化的卵。有许多鸟还未长大,难作长途飞行,同样也遭到遗弃而落在后边。参阅布莱克瓦尔的《动物学研究》(Researches in Zoology),1834年,108,118页。关于另外的证据,虽无必要,亦可参阅勒罗伊的《科学通信》(Letters Phil.),1802年,217页。关于东亚雨燕(swifts),参阅高尔得的《大不列颠鸟类导论》(Introduction to the Birds of Great Britain),1823年,5页。亚当斯(Adams)先生在加拿大观察到相似的情况,见《通俗科学评论》(Pop.ScienceReview),1873年7月,283页。
(270) 休姆(Hume)说[《关于道德原理的探讨》(An Enquiry Concerning the Principles of Morals),1751年,132页]:“似乎必须承认,他人的幸福和悲痛并非是同我们毫不相干的景象,而是看到前者……将使我们暗暗感到喜悦;而后者的出现……则会在我们的想象上投射一层忧郁的阴影。”
(271) 《心理学和道德学》,1868年,254页。
(272) 我在这里涉及的是所谓实质的和形式的道德之间的区别。我高兴地看到赫胥黎教授关于这个问题持有和我同样的观点。莱斯利·斯蒂芬先生说[《论自由思想和坦白讲话文集》(Essays on Freethinking and Plain Speaking),1873年,83页],“在实质的和形式的道德之间形而上学的区别正如其他这等区别那样,是彼此不相干的”。
(273) 我曾举过这样一个事例,即:三个巴塔戈尼亚地方的印第安人宁愿一个跟着一个地被枪毙,也不泄露其同伴的作战计划。
(274) 仇恨或敌意似乎也是一种高度持久的情感,也许比可以指出名字的任何其他情感更加持久。嫉妒的解释是,对他人的某种优点或成功感到憎恨,培根极力主张(《论文第九》)“在所有情感中,嫉妒是最缠绕不休而永续的”。狗很容易憎恨生人和生狗,尤其是它们居住靠近而又不属于同一个家族、部落或氏族时更加如此:这种情感似乎是天生的,而且肯定是最持续的一种。它同真正的社会本能似乎相辅而又相反。从我们所听到的未开化人的情况来看,似乎差不多也是这样。倘真如此,如果同一部落的任何成员对任何人有所损害或者成为他的敌人,那么后者把这等感情转而施于前者,只要再跨进一小步就可以了。一个人对敌人加以伤害,不会受到原始良心的谴责,而如果不是为自己报仇的话,那就要受到原始良心的谴责,这并非是不可能的。以德报怨,施爱于敌,乃道德之顶峰,社会本能本身是否曾导致我们如此,实属可疑。在任何这等金科玉律被想到和被遵从之前,这等本能以及同情,应该受到高度的磨炼,并且在理性、教育以及对上帝的爱和惧的帮助下而加以扩大。
(275) 安大略,《涉及法律的精神错乱》(Insanity in Relation to Law),美国,1871年,1页。
(276) 泰勒,《当代评论》(Contemporary Review),1873年4月,707页。
(277) 普罗斯佩尔·德斯平(Prosper Despine)博士在他的《天赋心理学》(Psychologie Naturelle)(第1卷,1868年,243页;第2卷,169页)一书中举出有关最恶劣罪犯的许多奇特事例,这些罪犯显然完全没有良心。
(278) 参阅一篇富有才华的论文,见《北英评论》(North British Review),1867年,395页;再参阅巴奇霍特(W.Bagehot)先生讨论服从和团结一致对原始人类的重要性的文章,见《双周评论》(Fortnightly Review),1867年,529页;1868年,457页及其他。
(279) 加里曼丹的一种原始人。——译者注
(280) 我所见过的最充分的记载是由格兰德(Gerland)作出的,见他的著作《自然民族的消亡》(Ueber dan Aussterben der Naturv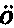 lker),1868年,但在后一章我势必还要对杀婴问题进行讨论。
lker),1868年,但在后一章我势必还要对杀婴问题进行讨论。
(281) 关于自杀的很有趣的讨论,参阅莱基(Lecky)的《欧洲道德史》(History of European Morals),第1卷,1869年,223页。关于未开化人,温伍德·里德(Winwood Reade)告诉我说,西非的黑人常常自杀。众所周知,自从被西班牙征服之后,在悲惨的南美土著居民中多么盛行自杀。关于新西兰,参阅《“诺瓦拉”航海记》(The Voyage of the“Novara”),以及关于阿留申群岛(Aleutian Islands),参阅乌泽在《论智力》(第2卷,136页)一书中引用的米勒著作。
(282) 参阅巴奇霍特的《医学与政治学》(Physics and Politics),1872年,72页。
(283) 例如,参阅汉密尔顿(Hamilton)关于卡法尔人(Kaffirs)的记载,见《人类学评论》(Anthropological Review),1870年,15页。
(284) 关于这个问题,伦南(Lennan)先生搜集了一些很好的事实,见他的著作《原始婚姻》(Primitive Marriage),1865年,176页。
(285) 莱基,《欧洲道德史》,第1卷,1869年,109页。
(286) 《出使中国记》(Embassy to China),第2卷,348页。
(287) 参阅卢伯克的《文化的起源》第七章,其中有关于这个问题的充实证据。
(288) 例如,莱基的《欧洲道德史》,第1卷。
(289) 《威斯敏特评论》(Westminster Review),1869年10月,498页,载有一篇富有才华的论文始用这一术语。关于“最大幸福原则”参阅米尔的《功利主义》(Utilitarianism),17页。
(290) 米尔(Mill)以最明晰方式承认(《逻辑体系》(System of Logic),第2卷,422页),行为可以通过习性而完成之,无须预先感到愉快。塞吉威克先生在一篇《论愉快和愿望》的文章[《当代评论》(The Contemporary Review),1872年4月,671页]中也说,总之,有一种学说谓自觉行为的冲动永远指向在我们本身产生令人愉快的感觉:与此相反,我则主张,我们到处都可以在意识中发现不受注重的冲动,这是指向某些令人不愉快的事情的:在许多场合中;这种冲动同自重如此不能和谐共存,以致二者不易在意识中同时存在。”我不能不认为,有一种模糊的感觉以为我们的冲动决非永远来自任何同时发生的或预先感到的愉快;这种模糊的感觉正是接受道德的直觉论而反对功利论或“最大幸福”论的一个主要原因。关于后一理论,行为的标准和动机无疑往往被搞乱了,实际上它们在某种程度上就是混淆不清。
(291) 华莱士先生在《科学上的意见》(Scienti fic Opinion,1869年9月15日)举出了一些好事例;在他的《对自然选择学说的贡献》(Contributions to the Theory of Natural Selection,1870年,353页)一书中有更加充分的叙述。
(292) 南美亚马孙河以南的大草原。——译者注
(293) 西班牙人和印第安人的混血种。——译者注
(294) 坦尼森(Tennyson),《国王的叙事诗》(Idylls of the King),244页。
(295) 《罗马皇帝奥瑞利亚斯·安东尼纳斯的思想》(The Thoughts of the Emperor M.Aurlius Antoninus),英译本,第2版,1869年,112页。奥瑞利亚斯生于公元121年。
(296) 斯宾塞给米尔的一书信,见贝恩先生的《心理学和道德学》,1868年,722页。
(297) 莫兹利,《躯体和精神》(Body and Mind),1870年,60页。
(298) 一位作者在《北英评论》(1869年7月,531页)中很好地作出了一个合理判断,表示强烈支持这一结论。列基先生(《道德史》,第1卷,143页)的看法似乎与此吻合。
(299) 参阅他的名著《遗传的天才》(Hereditary Genius),1869年,349页。阿盖尔(Argyll)公爵(《原始人类》,1869年,188页)关于人类本性在是非之间的斗争有过一些好议论。
(300) 《奥瑞利亚斯的思想》,139页。
(301) 《人类学评论》,1864年5月,158页。
(302) 正如亨利·梅因(Henry Maine)爵士所说的,被吸收进另一个部落中的诸成员或部落经过一段时间之后,便设想他们是同一祖先的共同后裔,见《古代法律》(Ancient Law),1861年,131页。
(303) 莫洛特(Morlot),《自然科学普及协会》(Soc.Vaud.Sc.Nat.),1860年,294页。
(304) 我在《动物和植物在家养下的变异》(第2卷,196页)举出过这方面的事例。
(305) 他以《自然科学与政治学》(Physics and Politics)为题,发表了一系列卓越的论文,见《双周评论》,1867年11月;1868年4月1日;1869年7月1日;以后印成单行本。
(306) 华莱士先生在《对自然选择学说的贡献》(1870年,354页)举出过有关事例。
(307) 《古代法律》,1861年,22页。关于巴戈霍特先生的叙述,见《双周评论》,1868年4月1日,452页。
(308) 《动物和植物在家养下的变异》,第1卷,309页。
(309) 《弗雷泽杂志》(Fraser's Magazine),1868年9月,353页。这篇文章似乎打动了许多人,由此引出两篇卓越的论文和一篇答辩,见《旁观者》,1868年10月3日及17日。在《科学季刊》(Q.Journal of Science,1869年,152页):劳森·泰特(Lawson Tait)在《都柏林医学季刊》(Dublin Q.Journal of Medical Science,1869年2月);兰克斯特先生在《长寿的比较》(Comparative Longevity,1870年,128页)均对此进行过讨论。《澳大利亚西亚人》(或可译为大洋洲人。——译者注)(Australasian,1867年7月13日)也出现过相似观点。我曾借用过其中几位作者的观念。
(310) 关于华莱士先生,参阅上面引用的《人类学评论》;关于高尔顿先生,参阅《麦克米伦杂志》,1865年8月,以及他的巨著《遗传的天才》,1870年。
(311) 菲克(H.Fick)教授关于这个问题以及其他各点做过良好叙述,见《自然科学对权力的影响》(Ein fluss der Naturwissenschaft auf das Recht),1872年。
(312) 《遗传的天才》,1870年,132—140页。
(313) 考垂费什(Quatrefages),《科学报告评论》(Revuedes Cours Scientifiques)。1867—1868年,659页。
(314) 参阅兰克斯特(Lankester)先生的《长寿的比较》一书中根据权威材料编制的表格第五栏和第六栏。
(315) 《遗传的天才》,1870年,330页。
(316) 《物种起源》,第5版,1869年,104页。
(317) 《遗传的天才》,1870年,347页。
(318) 兰克斯特,《长寿的比较》,1870年,115页。关于酗酒者的统计数字,采自尼逊(Neison)的《生命统计》(Vital Statistics)。关于荒淫生活,参阅法尔博士的《结婚生活对死亡率的影响》(Influence of Marriage on Mortality),曾在“社会科学全国促进会”(Nat.Assoc.for the Promotion of Social Science)上宣读,1858年。
(319) 《弗雷泽杂志》,1868年9月,353页。《麦克米伦杂志》(Macmillan's Magazine),1865年8月,318页。法勒(Farrar)牧师持有不同的观点(《弗雷泽杂志》,1870年8月)。
(320) 《关于妇女生育性的规律》(On the Laws of the Fertility ofWomen),见《皇家学会会刊》(Transact.Royal Soc.),爱丁堡,第24卷,287页:现以单行本出版,书名为《生殖力,生育性及不育性》(Fecundity,Fertility and Sterility),1871年。再参阅高尔顿先生的《遗传的天才》,352—357页,有对上述效果的观察材料。
(321) 五、六世纪入侵并定居于英国的日尔曼族。——译者注
(322) 公元前一千年左右居住在中欧、西欧的部落,其后裔今散布在爱尔兰、威尔士、苏格兰等地。——译者注
(323) 《苏格兰的出生与死亡情况第十次年度报告》(Tenth Annual Report of Births,Deaths&c.,in Scotland),1867年,29页。
(324) 引文系摘自关于这等问题的英国最高权威法尔博士的一篇论文:《结婚生活对法国人死亡率的影响》,此文曾在“社会科学全国促进会”宣读,1858年。
(325) 法尔博士,同上文,下述引文亦摘自同一篇著名论文。
(326) 我引用的数字是《苏格兰的出生和死亡情况第十次年度报告》(1867年)中所载的五年平均数。引用斯塔克博士的话载于《每日新闻》(Daily News),1868年10月17日,法尔博士认为此文写作严谨。
(327) 关于这个问题,邓肯博士说道(《生殖力、生育性及不育性》,1871年,334页):“在各个时期,健康而美丽者常从未婚一方走到已婚一方,于是未婚一方便充满了不幸的病弱者。”
(328) 参阅高尔顿先生关于这个问题的有独创性的最初论点,见《遗传的天才》,340—342页。
(329) 格雷格先生,《弗雷泽杂志》,1868年9月,357页。
(330) 《遗传的天才》,1870年,357—359页。法勒牧师提出过相反的论点(《弗雷泽杂志》,1870年8月,257页)。莱伊尔爵士在一段引人注目的文章中[《地质学原理》(Principles of Geology),第2卷,1868年,489页]要求人们注意宗教审判所产生的恶劣影响,通过选择它降低了欧洲的一般智力标准。
(331) 高尔顿先生,《麦克米伦杂志》,1865年8月,325页。再参阅《达尔文主义与国民生活》(On Darwinism and National Life)一文,见《自然》(Nature),1869年12月,184页。
(332) 《美国的最后冬天》(Last Winter in the United States),1868年,29页。
(333) 我非常感激约翰·莫利(John Morley)对这个问题所做的好批评:再参阅布罗卡(Broca)的《论选择》(Les Sélections),见《人类学评论》(Revued'Anthropologie),1872年。
(334) 《论文化的起源》(On the Origin of Civilisation),见《人种学会会报》(Proc.Ethnological Soc.),1867年11月26日。
(335) 《原始人类》(Primeval Man),1869年。
(336) 曾在“大不列颠皇家协会”(Royal Institution of Great Britain)宣读,1867年3月15日。还有,《对人类初期历史的研究》,1865年。
(337) 《原始婚姻》,1865年。再参阅显然是同一位作者所写的一篇优秀的论文,见《北英评论》,1869年7月。还有,莫尔根先生的《关于亲属关系的社会等级,体系的起源之推测》,见《美国科学院院报》(Proc.American Acad.of Sciences),第7卷,1868年2月。沙夫豪森博士说过“在荷马史诗和《旧约全书》中都曾记载过用人做献祭品的遗风”,见《人类学评论》,1869年10月,373页。
(338) 卢伯克爵士,《史前时代》,第2版,1869年,第十五、十六各章。再参阅泰勒的《人类的早期历史》一书中最优秀的第九章。
(339) 为澳洲土著的武器,用曲形坚木制成,打出去可飞回原处。——译者注
(340) 南太平洋塔希提岛上的土著居民。——译者注
(341) 米勒在《诺瓦拉游记:古生物学,第三部》(Reise der Novara:Anthropolog.Theil,Abtheil.Ⅲ,1868年,127页),做过一些良好的论述。
(342) 关于各个博物学者在其分类法中给人类安排的位置,小圣·伊莱尔有过详细叙述,见《博物学通论》,第2卷,1859年,170—189页。
(343) 关于蚁类的习性,贝尔特先生在其《博物学者在尼加拉瓜》(1874年)一书中,发表过一些最有趣的事实。再参阅莫格里奇先生的令人钦佩的著作《农蚁》(Harvesting Ants),1873年,以及《两个世界评论》,1870年2月,682页。
(344) 韦斯特伍德(Westwood),《昆虫的近代分类》(Modern Class of Insects),第2卷,1840年,87页。
(345) 即猿类。——译者注
(346) 《动物学会会报》(Proc.Zoolog.Soc.),1863年,4页。
(347) 《关于人类在自然界的位置的证据》(Evidence as to Man's Place in Nature),1863年,70页及其他诸页。
(348) 即Semnopithecus nasalis,多群栖于加里曼丹等处的沿河乔木上,鼻长而突出,雄性老猴者尤长,可运动自如,且如吻,故又名“长鼻猴”proboscis monkey。——译者注
(349) 若弗鲁瓦(Isid.Geoffroy),《博物史通论》(Hist.Nat.Gén.),第2卷,1859年,217页。
(350) 《论人类身体的无毛》(Ueber die Richtung der Haare),见米勒的《解剖学和生理学的历史文献》(Archiv für Anat.und Phys.),1873年,51页。
(351) 里德(Reade)引用,《非洲见闻录》(The African Sketch Book),第1卷,1873年,152页。
(352) 关于长臂猿的毛,参阅《哺乳动物志》(Nat.Hist.of Mamm.),马丁著,1841年,415页。关于美洲猴和其他种类,也可参阅若弗鲁瓦的《博物史通论》,第2卷,1859年,216,243页,埃舍里希特,同前书,46,55,61页。欧文,《脊椎动物解剖学》,第3卷,619页。华莱士,《对自然选择学说的贡献》,1870年,344页。
(353) 《物种起源》,第5版,1869年,194页。《动物和植物在家养下的变异》,第2卷,1868年,348页。
(354) 《动物分类导论》(An Introduction to the Classification of Animal),1869年,99页。
(355) 这同米伐特先生暂定的分类法差不多是一样的(《科学协会会报》,1867年,300页),他把灵长目分为狐猴科(Lemuridae)、人科(Hominidae)和猴科(Simiadae),这三者相当于狭鼻猴类、卷尾猴科(Cebidae)和狨科(Hapalidae),后两个类群则相当于阔鼻猴类。米伐特先生现仍坚持上述观点,参阅《自然》,1871年,481页。
(356) 《动物学会会报》(Transact.Zoolog.Soc.),第6卷,1867年,214页。
(357) 米伐特先生,《科学协会会报》(Transact.Phil.Soc.),1867年,410页。
(358) 莫里先生和米伐特先生论狐猴科,《动物学会会报》,第7卷,1869年,5页。
(359) 关于这一点,赫克尔作出同样的结论,参阅《论人类的发生》(Ueber die Entstehung des Menschengeschlechts),见微尔和(Virchow)的《普通学术报告》,1868年,61页。再参阅赫克尔的《自然创造史》,1868年,在该书中他详细地叙述了他的关于人类谱系的观点。
(360) 福尔西·马若尔(C.Forsyth Major)博士,《在意大利发现的猴类化石》(Sur les Singes Fossiles trouvés en Italie),见《意大利博物学会会报》,第15卷,1872年。
(361) 《人类学评论》,1867年4月,236页。
(362) 《地质学原理》,1865年,583—585页。《人类的源远流长》(Antiquity of Man),1863年,145页。
(363) 《人类在自然界的位置》,105页。
(364) 在他的《普通形态学》一书中有详细的各表阐明及此,在他的《自然创造史》(Natiirliche Sch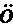 pfungsgeschichte)一书中特别论及人类。赫胥黎教授在评论后一著作时[《科学院院报》(The Academy),1869年,42页]说道,他认为赫克尔可称赞地讨论了人类由来的系统,虽然他对某些方面还持有异议。他对全书的要旨和精神给予了高度评价。
pfungsgeschichte)一书中特别论及人类。赫胥黎教授在评论后一著作时[《科学院院报》(The Academy),1869年,42页]说道,他认为赫克尔可称赞地讨论了人类由来的系统,虽然他对某些方面还持有异议。他对全书的要旨和精神给予了高度评价。
(365) 《古生物学》(Palaeontology),1860年,199页。
(366) 我在福克兰群岛于1883年4月满意地看到了一种复海鞘的能够运动的幼体,这一发现早于其他博物学者数年之久:复海鞘同Synoicum密切近似,但显然不是同属。其尾长为椭圆形头部的五倍左右,尾端为一很细的丝状体。我曾用简单的显微镜绘制过它的图,它明显地被横向不透明的部分分开,我设想这代表柯瓦列夫斯基所绘的大细胞。在发育的早期阶段,尾部紧密地缠绕在幼体的头部。
(367) 《圣彼得堡科学院研究报告》(Mémoires de l'Acad.des Sciences de St.Pétersbourg),第10卷,第15期,1866年。
(368) 但是,我理应补充一点:有些有能力的评论家们对这一结论还有争议,例如,捷得在《实验动物学文献》(1872年)中就此发表了一系列论文。尽管如此,这位博物学家还谈到(281页),“海鞘类幼虫的组织非任何假说和理论所可解释,由此可见,仅仅依靠对生活条件的适应,自然界就能使无脊椎动物产生出脊椎动物的基本形态(脊索的存在),我们虽不知这两大门动物的过渡在实际上是怎样完成的,但根据这一过渡的简单可能性,这两大门之间不可逾越的鸿沟就得以填平了。”
(369) 这是比较解剖学最高权威格根鲍尔(Gegenbaur)教授所做的结论,见《比较解剖学的主要特点》(Grundzüge der vergleich.Anat.),1870年,876页。这主要是对两栖类进行研究的结果;但是,根据沃尔戴耶(Waldeyer)的研究(《解剖学和生理学杂志》,1869年,161页),甚至“高等脊椎动物在性器官的早期状态时都是雌堆同体的”。相似的观点长期以来为某些作者所坚持,但直到最近还缺乏坚实的基础。
(370) 雄的袋狼(Thylacinus)提供了最好的事例。欧文,《脊椎动物解剖学》(Anatomy of Vertebrates),第3卷,771页。
(371) 在鮨鱼属(Serranus)的几个种中以及在某些其他鱼类中曾经观察到雌雄同体的情况,这等鱼类或是正常而对称的,或是异常而单侧的。祖特文(Zouteveen)博士给过我关于这一课题的参考资料,特别是哈尔贝茨玛(Halbertsma)教授在《荷兰科学院院报》(Transact.of the Dutch Acad.of Sciences)第16卷发表的一篇论文尤为重要。京瑟(Günther)博士怀疑这个事实,但现在有如此众多的优秀观察家们做过这方面的记录,以致没有任何争论的余地了。莱索纳(M.Lessona)博士写信告诉我说,他曾证实卡沃利尼(Cavolini)对鮨鱼所做的观察。埃科利尼(Ercolani)教授阐明鳗鲡是雌雄同体的[《波洛尼亚科学院院报》(Accad.delle Scienze,Bologna),1871年12月28日]
(372) 格根鲍尔曾阐明[《耶拿杂志》(.Jenaische Zeitschrift),第7卷,212页],在几个哺乳动物目中有两种不同模式的乳头,这二者怎么会来源于有袋类的乳头,而后者又来源于单孔类的泌乳器官,则是完全可以理解的,参阅麦克斯·赫斯(Max Huss)关于乳腺的研究报告,同前杂志,第8卷,176页。
(373) 洛克伍德(Lockwood)先生[《科学季刊》(Quart.Journal of Science),1868年4月,269页]根据对海马发育的观察,雄者的腹囊壁在某种方式上提供营养。关于雄鱼在口中孵卵,参阅怀曼教授的一篇很有趣的论文,见《波士顿博物学会会报》(Proc.Boston Soc.of Nar.Hist.),1857年9月15日;再参阅特纳教授的论文,见《解剖学和生理学杂志》,1866年,11月1日,78页。京瑟博士也描述过相似的事例。
(374) 鲁瓦耶(C.Royer)在她的《人类的起源》(Origine de l'Homme)中提出过相似的观点。
(375) 海岸生物所受潮汐的影响一定很大,无论生活在高潮线或低潮线的动物都必须每两周通过一次潮汐变化的完全循环。因此,它们的食物供给每周都要发生显著的变化。这等动物在这等条件下生活了许多世代,其生活功能几乎都要规则地按每周运转。那么,有一个难以理解的事实,即在高等的、现今为陆栖的脊椎动物以及另外一些纲中,许多正常的和异常的过程都是以一周或多周为期的:如果脊椎动物起源于同现今在潮汐中生存的海鞘类相近似的动物,上述情况就是可以理解的了。可以举出许多有关这等周期过程的事例,如哺乳动物的妊娠期、疾病的间歇热等皆是。卵的孵化也提供了一个良好的例子,因为,按照巴特利特(Bartlett)的说法[《陆与水》(Land and Water),1871年1月7日],鸽卵的孵化为两周;鸡卵的孵化为三周;鸭卵的孵化为四周;鹅卵的孵化为五周;鸵鸟卵的孵化则为七周。就我们所能判断的来说,任何一种过程或机能的循环周期,如果是在大致准确的期间内进行的,一旦获得之后,就不易再起变化:因而它将会通过几乎任何代数这样被传递下去。但是,如果机能变化了,周期势必也要变化,而且会按整个一周几乎突然地发生变化。这个结论如果正确,则是高度值得注意的:因为,每一种哺乳动物的妊娠期、每一种鸟卵的孵化期以及许多其他生命过程就这样向我们泄露了这等动物的原产地。
(376) 《印度史》(History of India),第1卷,1841年,323页。利巴神甫对中国人也做过同样记述。
(377) 古尔德著,《关于美国士兵的军事学和人类学的统计之研究》,1869年,298—358页,载有关于白人、黑人、印度人的大量测定数据。《关于肺的容量》(On the capacity of the lungs),471页。再参阅魏斯巴赫(Weisbach)博士根据舍策尔(Scherzer)博士和施瓦茨(Schwarz)博士的观察材料所举出的大量有价值的表,见《诺瓦拉游记》(Reise der Novara),1867年。
(378) 例如,参阅马歇尔先生关于一个布西门妇女的脑的记载,见《自然科学学报》,1864年,519页。
(379) 华莱士,《马来群岛》,第2卷,1869年,178页。
(380) 生活于西南非洲。——译者注
(381) 关于埃及著名的Abou-Simbel洞窟画像,普歇(M.Pouchet)说[《人类种族多源论》(The Plurality of the Human Races)英译本,1864年,50页],有些作者相信画上有12个以上民族的代表可以被辨认出来,但他都远远辨认不出来。甚至特征最显著的种族也不能被证实一致到那样的程度,就像在一些著作中关于这个问题所写的。例如诺特(Nott)和格利敦(Gliddon)两位先生说[《人类的模式》(Types of Mankind),148页],埃及国王拉米塞斯二世的面貌非常像欧洲人,而另一位坚决相信人类种族是独特物种的克诺斯(Knox)当谈到(《人类的种族》,1850年,201页)少年门南,(Memnon,伯契先生告诉我说,他就是拉米塞斯二世),却强烈地主张他的特性同安特卫普的犹太人相同。再者,当我看到阿姆诺甫(Amunoph)三世的塑像时,我同意博物馆两位职员的看法(两位都是优秀的鉴定家),即,他有特征显著的黑人面貌:但诺特和格利敦两位先生则把他描写成一个混血儿,但没有同“黑人混血”(同前书,146页,53图)。
(382) 诺特和格利敦在《人类的模式》(1854年,439页)中引用。他们还举出了确实的证据;但C.沃格特以为这个问题还需要进一步研究。
(383) 《人类种族起源的多样性》见《基督的检查员》(Christian Examiner),1850年7月。
(384) 《爱丁堡皇家学会会报》(Transact.R.Soc.of Edinburgh),第22卷,1861年,567页。
(385) 《关于人属的混血现象》(Onthe Phenomena of Hybridity in the Genus Homo),英译本,1864年。
(386) 参阅默里先生在《人类学评论》(1868年4月,53页)中发表的一封有趣的信,这封信驳斥了斯特莱斯基伯爵的如下叙述:澳洲土著妇女同白种男人生了孩子之后,再同自己种族的男人结婚就不生孩子了。夸垂费什也搜集了许多论据(《科学报告评论》,1869年3月,239页),证明澳洲土著居民和欧洲人交配,并非不育。
(387) 《对亚加西斯教授的动物界自然分布区概述的检查》,查尔斯顿,1855年,44页。
(388) 罗尔夫斯(Rohlfs)博士写信给我说,他在撒哈拉大沙漠发现一些混合种族,系来源于三个部落的阿拉伯人、栢栢尔人,以及黑人,他们特别能育。另一方面,里德先生向我说,黄金海岸的黑人虽然称赞白人和黑白混血儿,但有一句格言:黑白混血儿不应彼此结婚,因为他们生孩子少而且多病。正如里德先生所论述的,这一信念值得注意,因为白人访问和居住在黄金海岸已有四百年历史了,所以黑人有充分的时间通过经验而得到知识。
(389) 古尔德著,《关于美国士兵的军事学和人类学的统计之研究》,1869年,319页。
(390) 《动物和植物在家养下的变异》,第2卷,109页。我在这里提醒读者注意,物种杂交不育并不是一种特别获得的特性,就像某些树彼此不能嫁接在一起那样,这是由其他既获得的差异而附带发生的一种情形。这等差异的性质还不明,但它们特别同生殖系统有关系,而同外部构造或体质的正常差异的关系就少得多。物种杂交不育的一个重要因素显然在于一方或双方长期习惯于固定的条件;我们知道条件变化对生殖系统会产生特别影响,我们有良好理由相信(如上所述),多变的家养条件有消除不育性的倾向,而物种杂交不育在自然状况下则非常普遍。我在别处曾阐明(同前书,第2卷,185页,《物种起源》,第5版,317页),杂交物种的不育性并不是通过自然选择获得的:我们可以看到,两个类型如果已经成为很不育的了。那么它们的不育性几乎不可能通过保存那些日益不育的个体再把它们的不育性扩大;因为,当不育性增大之后,产生出来的后代则愈来愈少,最后仅仅在极稀疏的间隔时间内产出极少的个体而已。但是,还有较此为甚的更高级的不育性。格特纳(G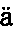 rtner)和克尔罗伊特(K
rtner)和克尔罗伊特(K lreuter)都曾证明,在包含许多物种的植物属中,从杂交后结籽愈来愈少的物种到决不结一粒种子的物种可以形成一个系列,但它们仍受其他物种的花粉的影响,从子房的膨大可以看出这一点。在这里,要想选择更加不育的个体显然是不可能的,因为这些个体已经停止结籽了:所以,如果仅是子房受到影响,极度的不育性是无法通过选择而得到的。这种极度的不育性,无疑还有另外一些等级的不育性,乃是杂交物种体质中某些未知的差异所造成的附带结果。
lreuter)都曾证明,在包含许多物种的植物属中,从杂交后结籽愈来愈少的物种到决不结一粒种子的物种可以形成一个系列,但它们仍受其他物种的花粉的影响,从子房的膨大可以看出这一点。在这里,要想选择更加不育的个体显然是不可能的,因为这些个体已经停止结籽了:所以,如果仅是子房受到影响,极度的不育性是无法通过选择而得到的。这种极度的不育性,无疑还有另外一些等级的不育性,乃是杂交物种体质中某些未知的差异所造成的附带结果。
(391) 《动物和植物在家养下的变异》,第2卷,92页。
(392) 考垂费什关于巴西的“保罗信徒”的成功和精力做过有趣的记载(《人类学评论》,1869年1月,22页),他们是葡萄牙人和印第安人多次混血的一个种族,而且还混有其他种族的血液。
(393) 分布在亚洲东南部及大洋洲。——译者注
(394) 例如美洲的和澳洲的土著居民。赫胥黎教授说(《史前人类学国际会议文献》,1868年,105页),南部德国人和南部瑞士人的头骨和“鞑靼人的一样短而阔”,等等。
(395) 关于这个问题,魏茨(Waitz)做过好的讨论,参阅《人类学概论》(Introduct.to Anthropology),英译本,1863年,198—208,227页。我曾引自塔特尔(H.Tuttle)的《人类的起源及其古远性》,波士顿,1866年,35页。
(396) 内格利(N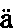 geli)教授在其《植物的中间类型》(Botanische Mittheilungen,第2卷,1866年,294—369页)一书中仔细地描述过几个显著事例,阿萨·格雷教授对北美菊科植物的一些中间类型做过同样叙述。
geli)教授在其《植物的中间类型》(Botanische Mittheilungen,第2卷,1866年,294—369页)一书中仔细地描述过几个显著事例,阿萨·格雷教授对北美菊科植物的一些中间类型做过同样叙述。
(397) 《物种起源》,第5版,68页。
(398) 关于这种作用,参阅赫胥黎教授的看法,见《双周评论》,1865年,275页。
(399) 《人类讲义》,英译本,1864年,468页。
(400) 《关于猪的族》(Die Racen des Schweines),1860年,46页。《关于猪头骨历史的预备研究》(Vorstudien für Geschichte,&c.,Schweinesch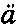 del),1864年,104页。关于牛,参阅考垂费什的《人种的同一性》(Unité de l'Espèce Humaine),1861年,119页。
del),1864年,104页。关于牛,参阅考垂费什的《人种的同一性》(Unité de l'Espèce Humaine),1861年,119页。
(401) 泰勒,《人类的早期历史》,1865年:关于姿势语言,参阅54页。卢伯克,《史前时代》,第2版,1869年。
(402) 《关于器具的相似形状》,见《人类学会纪要》,韦斯特罗普著。《斯堪的纳维亚的原始居民》(The Primitive Inhabitants of Scandinavia),英译本,卢伯克爵士编,1868年,104页。
(403) 韦斯特罗普,《关于上古遗物大石台》(On Cromlechs),见《人种学会杂志》的“科学意见”栏,1869年6月2日,3页。
(404) 《贝格尔号航海研究日志》,46页。
(405) 《史前时代》,1869年,574页。
(406) 译文见《人类学评论》,1868年10月,431页。
(407) 《史前考古学国际会议报告书》,1868年,172—175页。再参阅布罗卡的文章,其译文见《人类学评论》,1868年10月,410页。
(408) 格兰德博士,《原始民族的消亡》(Ueber das.Aussterben der Naturv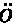 lker),1868年,82页。
lker),1868年,82页。
(409) 格兰德(同前书,12页)举出了一些事实以支持这一叙述、
(410) 关于这种影响,参阅霍兰(H.Holland)的著作《医学札记和回忆录》(Medical Notes and Reflections),1839年,390页。
(411) 我搜集过许多有关这个问题的好例子(《贝格尔号航海研究日志》,435页,再参阅格兰德的材料,同前书,8页)。波皮格(Poeppig)说“未开化人接受文明如饮毒药”。
(412) 斯波罗特,《未开化人生活的景象及其研究》(Scenes and Studies of Savage Life),1868年,284页。
(413) 巴奇霍特,《医学与政治学》(Physics and Politics),见《双周评论》,1868年4月1日,455页。
(414) 邦威克(Bonwick)著,《塔斯马尼亚人的末日》(The last of the Tasmanians),1870年,这里的叙述均引自该书。
(415) 这是塔斯马尼亚长官丹尼森(Denison)爵士的记载,《副总督生涯种种》(Varieties of Vice-Regal Life),1870年,第1卷,67页。
(416) 关于这些事例,参阅邦威克的《塔斯马尼亚人的日常生活》(Daily Life of the Tasmanians),1870年,90页,以及《塔斯马尼亚人的末日》,1870年,386页。
(417) 《对新西兰土著居民的观察》(Observations on the Aboriginal Inhabitants of New Zealand),政府出版,1859年。
(418) 肯尼迪(Alex.Kennedy)著,《新西兰》(“New Zealand”),1873年,47页。
(419) 扬格(C.M.Younge)著,《帕特森传记》(Life of J.C.Patteson),1874年,特别注意参阅第1卷,530页。
(420) 位于大洋洲。——译者注
(421) 即夏威夷群岛(Hawaiian Is.)。——译者注
(422) 根据复算,应为60.45%。——译者注
(423) 以上记述,主要引自下列著作:贾维斯(Jarves)的《夏威夷群岛的历史》,1843年,400—407页。奇弗(Cheever),《桑威奇群岛上的生活》(Life in the Sandwich Islands),1851年,277页。邦威克引用鲁申贝格的材料,见《塔斯马尼亚人的末日》,1870年,378页。贝尔彻(E.Belcher)爵士引用毕晓普的材料,见《环球航海记》(Voyage Round the World),第1卷,1843年,272页。历史人口调查的统计数字,系在尤曼斯(Youmans)博士的请求下,由寇恩先生慷慨提供的;在大多数情况下,我曾把尤曼斯的数字同上述各书的记载进行过比较,我没有用1850年的统计,因为我发现两个数据相差太远。
(424) 《印度医学公报》(The Indian Medical Gazette)。
(425) 南太平洋塔希提岛上的土著居民。——译者注
(426) 尼尔吉里(Nilgiri)山中的牧民。——译者注
(427) 关于诺福克岛民的密切亲缘关系,参阅丹尼森(Denison)爵士的《副总督生涯种种》,第1卷,1870年,410页。关于图达人,参阅马歇尔(Marshall)上校的著作,1873年,110页。关于苏格兰西方诸岛,米切尔(Mitchell)博士,《爱丁堡医学杂志》(Edinburgh Medical Journal),1865年3—6月。
(428) 缅甸中部的古城。——译者注
(429) 关于这个问题的证据,参阅《动物和植物在家养下的变异》,第2卷,111页。
(430) 即Cynaelurus jubatus Schreb.,印度人驯养之,使其猎羚羊和鹿等。——译者注
(431) 《动物和植物在家养下的变异》,第2卷,16页。
(432) 位于大洋洲。——译者注
(433) 这些数字引自《受到宽大的叛变者》(The Mutineers of the“Bounty”),贝尔契夫人著,以及英国下院1863年5月29日命令出版的《皮特凯恩岛》(Pitcairn Island)。关于桑威奇群岛的下列叙述,引自《檀香山公报》(Honolulu Gazette)以及寇恩(Coan)先生的著作。
(434) 《人类学》(On Anthropology),译文载于《人类学评论》,1868年1月,38页。
(435) 婆罗门为印度封建种姓制度的第一种姓。僧侣。——译者注
(436) 《孟加拉农村年报》(The Annals of Rural Bengal),1868年,134页。
(437) 《动物和植物在家养下的变异》,第2卷,95页。
(438) 帕拉斯(Pallas),《圣彼得堡科学院院报》(Act.Acad.St.Petersburg),第二部,1780年,69页。
(439) 安德鲁·史密斯爵士(Sir Andrew Smith),诺克斯引用,见《人类的种族》,1850年,473页。
(440) 参阅夸垂费什关于这个问题的著作,见《科学报告评论》,1868年10月17日,731页。
(441) 利文斯顿,《南非旅行调查记》(Travels and Researches in S.Africa),1857年,338,339页。多比尼和戈得隆在《论物种》中引用,第2卷,266页。
(442) 参阅1813年在皇家学会宣读的一篇论文,见1818年出版的他的《论文集》。关于韦尔斯博士的论点,我曾在《物种起源》的“历史概述”中有所说明。关于肤色同体质特性的相关,我曾在《动物和植物在家养下的变异》(第2卷,227,335页)举过种种事例。
(443) 例如,参阅诺特(Nott)和格利敦(Gliddon)合著的《人类的模式》,68页。
(444) 塔洛克(Tulloch)少校1840年4月20日在统计学会上宣读的一篇论文,载于《科学协会会刊》(Athenaeum),1840年,353页。
(445) 《人类种族的多源论》(The Plurality of the Human Race),英译本,1864年,60页。
(446) 考垂费什,《人种的一致性》,1861年,205页。魏采,《人类学概论》,英译本,第1卷,1863年,124页。利文斯顿在他的《旅行记》中举过同样事例。
(447) 在圭亚那。——译者注
(448) 1862年我曾得到陆军军医总监的许可,向海外驻军的医生发出空白表格,并附如下意见,但未获得答复,“有几个被记载下来的十分明显的事例表明,在我们的家养动物中,皮肤附属物的颜色同其体质有一定关联;众所周知,人类种族的肤色同其住地的气候也有某种有限度的关联;下述调查似乎值得注意。即,欧洲人的毛发颜色同他们感染热带地方疾病之间是否有任何关联。如果各军队的医生驻在对健康有害的热带地区,请在发病时先数一下军队中有多少人的毛发是浓色的,多少人是淡色的,多少人是中间色或不确定的颜色的;如果同一位医生对疟疾、黄热病、痢疾患者,也作出相似的统计,那么当表上有三千来个这样事例之后,很快就可以看明,在毛发颜色同感染热带病的体质之间是否存在着任何关联。也许不会发现这种关联,但这样调查还是值得一做的。倘获得正的结果,则这一结果在选用人员担负任何特殊任务时是有一定实际应用价值的。在理论上,这一结果大概也是重要的,因为它指明了自从远古以来就在对健康有害的热带气候下居住的一个人类种族,在悠久的连续世代中由于深色毛发和深色皮肤的个体更好地被保存下来而成为深色的一个途径。”
(449) 位于西印度群岛。——译者注
(450) 《人类学评论》,1866年1月,21页。夏普(Sharpe)博士也说,“在印度,生有淡色毛发和红润面色的人比生有深色毛发和青白面色的人感染热带地方病者为少;据我所知,这一意见似乎有充分的根据”(《人类是一种特殊创造物》,1873年,118页)。另一方面,塞拉利昂的赫德尔(Heddle)先生则持有直接相反的观点,“他手下的职员死于西非海岸气候者比其他人为多”(里德著,《非洲随笔》,African SketchBook,第2卷,522页),伯顿上尉持有同样见解。
(451) 《人类是一种特殊的创造物》(Man:a Special Creation),1873年,119页。
(452) 《动物和植物在家养下的变异》,第2卷,336,337页。
(453) 例如,参阅考垂费什有关在埃塞俄比亚和阿拉伯居住效果的记述(《科学报告评论》,1868年10月,724页)以及其他相似的事例。罗勒(Rolle)博士说(《人类的起源》,Der Mensch,seine Abstammung,1865年,99页),根据汉尼柯夫(Khanikof)的权威材料,大多数德国人的家族在佐治亚(Georgia)定居两代之后,头发和眼睛将会变为黑色。福布斯(Forbes)先生告诉我说,安第斯山的基切亚人(Quichuas)按照彼等所住山谷的位置,其肤色变异很大。
(454) 哈伦(Harlan),《医学研究》(Medical Researches),532页。考垂费什(《人种的一致性》,1861年,128页)曾就这个问题搜集了重大证据。
(455) 巴西的印第安人,他们在下嘴唇穿装一木塞子,叫做“botoque”,因是得名。——译者注
(456) 参阅沙夫豪森的著述,其译文见《人类学评论》,1868年10月,429页。
(457) 赛奥恩印第安人(Siouan Indians)的一个著名部落,在北达科他(Dakota)州,1837年由于天花的流行,几遭覆灭。——译者注
(458) 赛奥恩印第安人(Siouan Indians)的一个著名部落,在北达科他(Dakota)州,1837年由于天花的流行,几遭覆灭。——译者注
(459) 凯特林(Catlin)说(《北美的印第安人》,第1卷,第3版,1842年,49页),曼丹人(Mandans)的整个部落,l0个人或12个人中就有一个人的头发是明亮的银灰色的,而且这是遗传的,一切年龄的男女都是如此。现在,这种头发之粗硬如马鬃,而其他颜色的头发还是细而软的。
(460) 关于皮肤的气味,参阅戈德隆的《论物种》(Sur l'Espèce),第2卷,217页。关于皮肤上的分泌孔,参阅威尔肯斯的《家畜饲养技术的任务》(Die Au fgaben der Landwirth.Zootechnik),1869年,7页。
(461) 《关于人类的大部分脑旋圈》(Die Grosshirn-Windungen des Menschen)《巴伐利亚学院论文集》(Abhandlungen der K.Bayerischen Akademie),第10卷,1868年。
(462) 《人类大脑旋圈局部解剖学》(Convolutions of the Human Cerebrum Topographically Considered),1866年,12页。
(463) 《特别关于黑猩猩的起沟通作用的脑旋圈的记载》(Notes more especially on the bridging convolutions in the Brain of the Chimpanzee),见《爱丁堡皇家学会会报》,1865—1866年。
(464) 弗劳尔,《僧面猴的解剖》(On the Anatomy of Pithecia Monachus),见《动物学会会报》,1862年。
(465) 《人类在自然界的位置》,1862年,102页。
(466) 《动物学会学报》(Transactions of the Zoological Society),第5卷,1862年。
(467) 葛拉条雷在其《关于人类脑褶痕的研究报告》(39页,第四图版,第3图)中说道,“在一切猿类,皆为脑后叶褶痕最先发达,而前部褶痕则发达较迟,颅顶后头部裂沟在胎体中比较大。人类的前部裂沟最先出现,这是显著的例外,不过大脑前叶的普通发育同猿类均依同一规律”。
(468) 葛拉条雷说(原著39页):“此胎体的脑后部裂沟发育甚好,而脑前部的裂沟几乎不可见。”第四图版第3图中的罗兰德氏裂(Rolando fissure)和前部裂沟均甚明晰。阿利克斯(Alix)在他所写的《对葛拉条雷的人类学观点的评论》(见《巴黎人类学会会报》,1868年,32页)一文中说道:“葛拉条雷所有者为一长臂猿胎体的脑,这种猿与猩猩相近,处于生物界的很高等级,最有名的博物学家把它列入似人猿类。例如,赫胥黎力持这种看法。葛拉条雷从长臂猿的一个胎体发现脑前叶的裂沟尚未出现时,脑后叶的褶痕已有很好的发育了。这就是说,人类的褶痕的出现,由α在ω,而猿类的褶痕的发达,乃由ω在α。”
(469) 《人类和猿类大脑半球主要部分的脑沟和脑回的典型排列方式》(Ueber die typische Anordnung der Furchen und Windungen auf den Grosshirn-Hemisph ren des Menschen und der Affen),见《人类学文集》(Archiv für Anthropologie),第3卷,1868年。
ren des Menschen und der Affen),见《人类学文集》(Archiv für Anthropologie),第3卷,1868年。
(470) 《人类胎儿大脑半球主要部分的脑沟和脑回的发育过程》(Zur Entwickelungs Geschichte der Furchen und Windungen der Grosshirn-Hemisph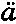 ren im Faetus des Menschen),见《人类学文集》,第3卷,1868年。
ren im Faetus des Menschen),见《人类学文集》,第3卷,1868年。
(471) 属灵长类,其性质在狭鼻猴类和阔鼻猴类之间,形小,尾长。前肢的拇指不能同其他四指对向;后肢的拇指虽有普通猿类所具有的那样扁爪,但其他趾则有钩爪而同食肉兽类相似。——译者注
(472) 例如,勒孔特(Lecomte)神甫所写的那本很糟的小册子,《达尔文主义和人类的起源》(Le Darwinisme et l'origine de I'Homme),1873年。
