第八讲
一、反射运动
我们已经看到,感觉(sensation)是构成我们心理生活的大量具体过程的最终源泉。在我们的观念世界中,我们所遇到的每样东西均导源于感觉,而观念则是一切高级心理活动的原料。然而,人们可以这样提出疑问:倘若没有来自外部世界的强化(reinforcement),思维流(stream of thought)是否就不能继续呢?思维在感觉中有其源泉,感觉确实持续不断地介入以决定思维的方向,这一点至少是可以肯定的。这样一来,我们关于感觉的考察直接导致我们考虑第二个也是十分重要的心理学问题——也即从大量而又多样地进入观念的感觉元素(sensational elements)中组成观念。
在我们先前的讨论中,我们在涉及感觉本身以前考察了感觉(也即外部感觉刺激)的物理条件。现在,产生了进一步的问题:一种感觉的直接结果是什么?观察为我们提供了十分明确的答案:在具有充分强度而且未曾被相反的影响所抑制的每一种感觉之后,伴随而来的是一种肌肉运动(muscular movement),这种肌肉运动我们称之为反射运动(reflex movement),如此命名是适当的。十分清楚,在神经系统的中枢器官里发生了神经兴奋的转移,也就是从感觉纤维转向运动纤维,而且类似的刺激从这些运动纤维向它们的特定肌肉传递,可以说,刺激被抛回,也就是被反射。只要感觉神经与神经系统的中枢器官保持联系,只要感觉神经通过运动神经与肌肉保持统一,人们就可以观察到反射现象。不过,整个中枢神经系统也不一定能够实施其正常功能。脊髓可以与大脑相断离,但是脊髓内的神经仍然可以传递反射。确实,光是脊髓的很小一部分就足以传递兴奋了。
感觉神经向运动神经的刺激传递是由神经系统的基本结构提供的。如果我们借助显微镜观察大脑和脊髓,首先,我们会发现若干粗细不等的神经纤维,感觉神经干(nerve-trunk)和运动神图16经干的连续;其次,除了这些东西以外,还可发现各种大小不等的细胞——黏液结构,像大多数机体细胞一样,包含更为坚实的细胞核和若干小颗粒。这些神经细胞使神经系统的中枢器官具有特征,而且对神经系统的中枢器官来说是独特的。至于它们的功能究竟有多么重要,你可以从下述事实进行判断,即它们始终处于与传入神经纤维(afferent nerve-fibres)和传出神经纤维(efferent nerve-fibres)的联系之中。一般说来,每个神经细胞发出几根神经纤维。因此,我们可以把这些细胞部分地看做是终端器官,部分地看做是联结传入神经纤维和传出神经纤维的中介器官。为了使这种反射的机制更加清楚,我们只需设想在两根神经纤维之间插入了一个神经细胞,其中一根神经纤维(e)来自一个感觉器官,另一根神经纤维(b)则通向肌肉。于是我们便得到了反射的图式。然而,实际上,正如我们可以预料的那样,一旦我们了解了我们对中枢神经结构的复杂性该做些什么以后,神经联结要复杂得多。
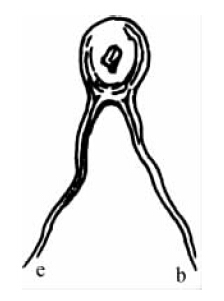
图16
对感觉刺激进行反射性反应,其运动的强度和规模在不同的情形中十分不同。一般说来,它们随着刺激强度的增强而增强。十分微弱的刺激不会激起反射运动;适度的强度引起适度的运动,适度的运动又受制于某些特定的肌肉群;随着刺激强度进一步增强,反射性反应也变得更加普遍,直到最后实际上涉及整个有机体为止。反射运动随着刺激的增加而增加的定律是始终如一的,而不顾个体差异和暂时差异可能相当大这一事实。所有这些,主要归诸于各种各样的兴奋性。感觉神经和神经中枢越是容易激发,反射便越早出现,并能更迅速地经历集中变化和扩散变化的全部过程。再者,它可以通过各种对有机体产生影响的作用而增强或减弱。切除头部或大脑能增加反射,直到死亡来临为止。许多两栖动物在被切除头部以后仍然可以生活几个月之久,在此期间,它们的反射兴奋性异常之大。还有一些化学物质,它们对中枢器官的各种组织的影响是引起反射的增强。除了某些生物碱(它们的影响并不很大)之外,我们在这里想特别提及一下马钱子碱(strychnine)。它产生这样一种过度的敏感性,使得对皮肤神经的最轻微压力(在一般情况下原本是不会引起任何运动的)也会引起反射并传向整个身体。可是,在深度睡眠或晕厥中,反射的兴奋性会减弱,正如鸦片或其他类似毒品所引起的情形那样。
至于神经细胞内部的化学变化是什么,马钱子碱和其他类似毒物的影响有赖于哪些神经细胞内部的化学变化,这些仍然不得而知。但是,它们的一般影响可以容易地从力量(forces)作用的普遍定律中推断出来。人们无法假设,与神经组织的成分不同的物质会创造出这般新颖的神经力量。唯一可以站得住脚的假设是,它使已经潜在地存在着的力量得以实现。也就是说,它克服了某些阻力[即将储存的能量转变为动能(kinetic energy)的阻力],从而使转化的力量更为有效。与此对照,一种相反性质的物质将会增加阻力的数量,从而也增加了使细胞从其紧张状态中得以释放的外部力量的数量。只要列举一个简单的机械例证,你们便会十分容易地理解这一点。
假定你有一只钟,它的运动随时可以停止或开始,只要在齿轮中加入或除去某种机械装置,譬如说一根弹簧。或者根据弹簧的紧或松,便能制止钟的运作或者促动钟的运作。只要收紧弹簧,那只钟转动起来的重量就对弹簧施加压力。这代表了一些应力(stress),钟的机械装置服从于这些应力。一旦弹簧松弛,这些应力便转化为运动能量。为了做到这种松弛,必须付出少量的功(work)。功的大小与弹簧弹性的恢复成比例,而且通过改变弹簧的张力(tension),这种功可以容易地增强或减弱。
在这一例证中,钟的运动相当于反射运动,弹簧的放松就是感觉刺激的运作,张力的增强或减弱代表了物质的影响,它对能量转换机制施加一种特殊的作用。正如弹簧的较大张力增加了一只钟的启动难度那样,鸦片在神经系统中产生的变化也会阻碍一种反射运动的释放。正如弹簧的较小张力能够促进钟的启动一样,马钱子碱也会促进反射运动。每只钟都能运行一段时间,到了发条松弛时,它便停转,从而需要重新拧紧发条。换言之,在钟的里面有某种潜能,它的衰竭和转化为运动都需要一定时间,然后需要更新。如果没有这种更新,钟就无法再走。这里,我们再次获得了与神经系统机制的完全类比,在中枢器官里存在一定量的潜能。也就是说,当它差不多衰竭时,它仅仅部分地得到更新(像在钟里面那样),于是睡眠期间发生了恢复的情况。但是,确实也部分地存在继续更新的过程,如果神经元素的化学合成不被如此剧烈地受到干扰,以至于使回复到正常情况成为不可能的话。因此,对潜能的过度消耗来说,其结果便是死亡。马钱子碱和类似的毒物之所以能导致死亡,原因就在于中枢器官能量的衰竭,尤其是脊髓的衰竭。身体的其他组织未被触动,甚至那些神经纤维,正如我们已经表明的那样,这些神经纤维已经与脊髓断离,却仍能处理和传递刺激。
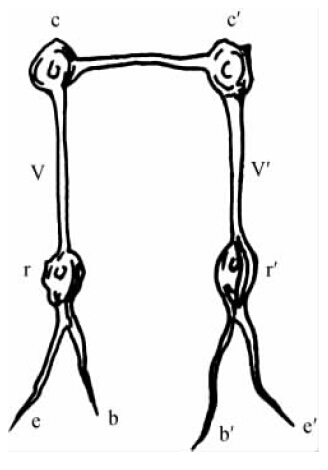
图17
现在,我们可以看到,切除大脑对反射产生的影响与这些毒药中任何一种毒药对反射产生的影响一样,有助于使反射机制运作起来。但是,很清楚,这种影响肯定以不同的方式产生。事实就是如此。对脊髓的神经细胞(将感觉纤维与运动纤维联结起来)来说,其本身是十分复杂地相互联系的,而且,发出一些通向大脑的细微神经纤维,然后在那里终止于中枢细胞丛(central cell plexus)。这些关系以图解形式在图17中呈示,在该图中,rr′是脊髓细胞,它们作为反射中枢起作用,而cc′则是中枢大脑细胞。作用于感觉神经(ee′)末端的刺激并非通过反射弧(reflex arc)的方式简单地向运动神经纤维(bb′)传递,而是传导到更高级的神经细胞(cc′)那里,然后扩散开来——它实际上可能是没有限制的。此外,图17还表明,单一的感觉纤维并不总是与单一的运动纤维相连。细胞与细胞之间的传导通路如此众多,以至于每一根感觉纤维与一些运动纤维相联结,甚至与其他感觉纤维相联结。因此,如果在e点上产生一个兴奋,我们就可以预料不仅在b点上,而且在b′和e′点上也会释放出力量来。为了对中枢器官有个大致的概念,你必须无限地增加这些图式的联结。一个感觉印象不仅导致一组肌肉运动,而且还导致身体不同部位的运动和感觉,也许还会扩展到整个肌肉群和若干感觉器官中去。
确实,关于反射性感觉(reflected sensations)的存在——即感觉不仅由它们本身的感觉神经引起,还可以由其他一些感觉神经引起——是毫无疑问的。一般说来,在正常情况下,这种反射性感觉是十分微弱的;而且,只有在病理性增加兴奋的状态下它们才达到更大的强度。因此,看来它们与反射运动有着根本的不同,在反射运动中,它们从来没有像直接受到刺激的感觉那样集中,而且,一般说来,只有在十分注意的情况下方能感知到。
问题的另一方面更为重要——由于感觉提供的冲动传递到身体的各个部分,因而扩展的反射运动不断地远离它的源点。提高刺激强度将会产生相当大的运动扩散,但是,相当普遍的反应只能在异常兴奋的情况下才能获得,例如,在马钱子碱的影响下获得。这里,对反射性反应的扩展没有规定明确的界限,实际上,所有的肌肉群在感觉刺激作用期间是强烈震动的。
但是,撇开这些异常的情况不谈,我们可以为反射的扩散界定一些明确的规则。那就是反射的扩散通常有赖于外部刺激的强度。当刺激恰好强到足以引起一个反射性反应时,所产生的运动始终限于一组与感受区域直接相连的肌肉群。如果受到刺激的部分是视网膜,那么由此产生的运动仅仅是眼睛的运动;如果四肢之一的皮肤受到刺激,那么,由此产生的运动仅仅限于该肢体的运动;如果头部或躯干的皮肤的某个部分受到刺激,那么,在最邻近的肌肉群里便会产生运动,而且,一般说来,在与刺激点最具密切关系的那个肢体上也会产生运动。于是,作用于左面颊的一个微弱刺激导致脸部左侧的歪曲,并导致左臂的运动。四肢与眼睛一起组成了人体上最能动的一些部分,这些最能动的部分明显地倾向于由皮肤刺激引起运动。
如果刺激的强度增加,那么反射运动就会更为广泛地扩散,但是在一定时期内仍然受到被刺激器官邻近部分肌肉的制约。这样,反射运动便可从一条腿或手臂扩展到其他的腿或手臂。随着强度进一步增加,反应变得越来越普遍;在强度达到最高点时,四肢都会普遍运动起来。起初,这样的运动是一种弯曲运动,但是在强度达到最高点时,弯曲运动变成伸展运动。一种影响所有运动纤维的刺激同样也产生一种伸展运动——这一事实似乎表明,引起伸肌的神经并不与引起屈肌的感觉神经纤维密切地联系着。除非刺激变得十分强烈,致使伸展的神经受到影响,否则的话,尽管反射性反应达到最高限度,伸展也无法比弯曲更占上风。
二、反射的目的性
在这两种主要形式中,反射具有目的性(purposiveness)特征。如果运动仅限于肌肉群,或者直接毗邻的受刺激点,那么,其结果便是使受影响的部分摆脱刺激的影响。如果反应更加扩散,那么肌肉的作用主要是帮助摆脱运动;而且,只有在一些极端的例子中(在那里,整个肌肉群被投入到伸展运动中去),人们才难以发现目的性证据。
关于这种目的性特征的主要理由是,反射运动通常指向某个目的(end),而且一旦达到目的,反射运动便会停止。这个目的是与受刺激的那个部分相联系的。如果你用一根尖针刺激无头蛙躯干的背部,那么,无头蛙的其中一条腿就会朝着受刺激的那个点剧烈地运动。这种触及运动看来就是整个运动的目的;而且,触及是以最简单的方式受影响的——通过一个肢体,或通过肌肉(它们可以用最少的力气发挥中介作用)。
对眼睛进行刺激会产生与皮肤刺激中获得的结果相类似的结果。如果你注视新生儿的一对眼睛,你一定会看到新生儿双眼凝视的固定性。事实上,眼睛在运动,尤其是当光刺激作用于眼睛时;但是,这种眼动是不规则的,仿佛在眼动和视觉印象的位置之间没有明确的联系。这种联系是逐步建立起来的。如果你将一盏灯放入出生才几天或几星期的婴儿视野之内,你就会发现婴儿将其眼睛转向于你,并凝视着灯光。如果你同时引入两盏灯或者两盏以上的灯,婴儿通常在一盏灯和其他的灯之间交替地凝视。但是,婴儿的凝视集中在灯上;他的眼睛以一种机械的必然性固定在灯光上面,而且只有在视觉印象由于疲劳而变弱时,或者在其他某个刺激物取代该灯光而出现时,眼睛才会离开原来的灯光。因此,可以这样说,我们面临着与反射运动中触及皮肤上受刺激的部分相类似的现象。当一种光刺激出现在视野之内时,眼睛便转向这种光刺激,正如手朝着皮肤上受刺激的部位移动一样。
然而,我们必须注意,眼的反射机制是双重的。一方面,在光觉(light-sensation)和使瞳孔收缩与眼睑闭合的肌肉之间存在一种联结;另一方面,在光觉和使眼球转动的肌肉之间也存在一种联结。如果一双眼睛处于黑暗环境中的话,那么眼睑反射可由相当微弱的光刺激引起。因此,我们不难理解,当新生儿第一次睁开眼睛接触到光线时,伴随而来的便是激烈的和痉挛性的眼睑闭合反射。但是,眼睛这一器官很快对光线有了适应;于是,光觉和眼球肌肉之间的联结便产生了。正如我们已经见到的那样,起初,当光刺激进入视野时,仅仅产生运动;我们不能说这种运动具有控制性或导向性。它仅仅是对光线的一种不确定的探索而已。但是,在婴儿出生后的第二个星期和第四个星期之间,已经可以观察到某种规律性。婴儿开始凝视;出现在婴儿视野内的每种光刺激引发一种对它凝视的倾向。据假设,凝视取决于眼睛所处的位置,在该位置中,一种明确的光印象在视网膜的最敏感部位形成一个图像。这个点位于整个视网膜表面的中心,在该点旁边不远处,视神经进入眼睛,并从那里辐射开来布满视网膜。从解剖学角度讲,它具有这样的特征,即存在一组十分接近的视网膜元素来处理光的刺激,并带有黄色色彩。后者就是通常所谓的“黄斑”(yellow spot)。
眼睛反射的发展方式是这样的:首先,光刺激仅仅激起一些不规则的运动。接着,这些运动表现出一种明确的形式,并服务于明确的目的。眼睛以这样的方式运动,结果使刺激性的光图像落到了黄斑上。如果在这个阶段一种光在视野范围内来回运动,那么眼睛便以相等的连续运动跟随着它。
可是,通过哪些步骤使这种规律性从原先的无规律性中产生呢?很显然,黄斑与反射运动的确定性关系无法用制约这些反射运动的神经元素的相互联结形式来提供。如果情况果真如此的话,那么投射到视网膜上的第一条光线将会释放出同样特性的反射运动,正如我们后来观察到的特性那样。现在,除了反对这种观点的证据以外(这种观点是由经验的事实提供的),没有任何东西可以用这样的方式来提示它,在该方式中,释放运动的力量是在中枢器官内传递的。从感觉神经纤维向运动神经纤维传递的力量,就强度和扩散而言,有赖于刺激的强度,以及促进其传递的器官的临时状态。因此,没有理由说,为什么黄斑附近的一个集中刺激只能激起一种很轻微的眼动,而视网膜周边的微弱刺激却激起了强烈的眼动。不!在逐步使它产生这种情况的感觉发展期间,显然有一些影响在起作用,而一种反射运动的释放是受到物理机制影响的,它的规模和方向完全有赖于受刺激部位的地点;所以,器官感受性越强,或者刺激的强度越增加,只能从更强的能量和更快的运动中得到反映。为了对这一切究竟是如何发生的问题获得一个完整的解释,我们必须密切地注意感觉器官的结构。
三、触觉反射和视觉反射的发展
覆盖在身体表面上的皮肤对刺激十分敏感。整个视网膜也同样敏感,唯一的例外是视神经的入口处,在那里没有任何特定的有益于视觉的终端器官。但是,由皮肤各部分产生的感觉或由视网膜各部分产生的感觉并不完全相似。就皮肤而言,你们可以十分简单地使自己确信这一点。如果你用手指触摸自己的面颊和手掌,小心翼翼地在每种情形里施加同样的压力,你就可以感觉到这两种感觉明显地不同。如果你将手掌与手背相比,颈部与后颈相比,或者将胸部与背部相比,或者将相隔一定距离的皮肤的两个部分相比,你可以同样感觉到相同的情况,即两种感觉明显地不同。不仅如此,如果你观察得仔细一点,你还会发现来自相邻两部分的感觉特性也有相当的不同。随着你从敏感表面的一点转向另一点,你便会在触觉上经历逐步的和连续的变化过程,尽管外部压力的性质仍保持完全不变。即便是身体两半对应的部位,虽然它们很相似,但是它们的感觉还是不同的。如果你先触摸一只手的手背,然后再触摸另一只手的手背,你将会发现这两种感觉之间是有细微差别的。
在视网膜内也可表明存在这种类似的变化。凝视一张拿在手里的红纸,然后缓慢地将红纸移开,不许你的眼睛跟踪红纸直到红纸消失。当然,红纸的图像首先落在黄斑上面;然后,穿过视网膜朝周边扩散。你将会观察到,在物体的横向运动中,红色的感觉经历了逐渐的变化:色调变得更深,似乎带点蓝色,最后完全变成纯粹的黑色。你选择的任何一种颜色,白色的也一样,都会显示出类似的变化。感觉的最后阶段往往是黑色。
对这种现象的明显解释是,在视网膜的不同部位,我们有着不同的感觉;随着物体印象从中央向外围区域移动,感觉也逐渐发生变化。就我们所能讲的而言,不管运动的方向如何,变化是以相同的方式发生的;但是——而且这是值得注意的——这种变化在不同的方向上是以不同的速度发生的。如果物体从中央向外或向上移动,那么颜色系列要比物体向内或向下移动经历更为迅速的变化;结果,位于视网膜外周或上部的物体图像显得发黑,而如果物体图像落在视网膜内部或下部的相应部位,它看起来仍然是彩色的。
当你进行色彩变化的实验,即将一些小物体置于视网膜外区以便它们被看到时,你会惊奇地向自己提问,为什么你并未注意到这些差异;也就是说,蓝色的天空或房子的红墙并不总是镶上黑边。确实,我们应该自然地预料到,如果一张蓝色的纸或者一张红色的纸逐渐变黑,这是由于它趋向视网膜外周部分的缘故,天空的蓝色和砖头的红色随着向视野的周边移动,也会显示出某种变化。如果我们在讨论感觉问题时,未能熟悉大量的事实(这些事实指出了解决该问题的途径),那么,我们在回答这个问题时就会发生相当大的困难。我们必须记住,一种感觉并非确定的或不变的某种东西,而是一种比较的产物,或者,更确切地说,是与其他感觉发生联想关系的结果,它不由单一刺激的特性所形成,而是由同时产生的和先前产生的印象的特性所形成。它并不单独存在,而是被带入到关系之中。如果这些协作的决定因素如此强烈,以至于使我们在某些情境里看到蓝红色和红蓝色——你们记得色彩对比(colour-contrast)的现象——则它们也许会使我们忽视色调的差异,这种差异是由物体在视网膜不同部位的移动造成的。
我们有理由假设,我们从注视大型的彩色表面和注视均匀的彩色表面中获得的感觉本身是完全一致的。这是因为,如果我们移动眼睛,连续凝视彩色表面的不同部位——也即使它们有序地落在黄斑上——则我们便在每一种情形里精确地接收到同一种颜色印象。原先的感觉提供了一些差异,这是可以肯定的;但是,在成千上万次的经验中,也即将它们直接归之于印象的空间分配,我们已经把这些差异消除了,这样的联想是以机器般的确定性和精确性加以贯彻的。因此,当我们正在注视大而连续的彩色表面时,我们由于印象的地点之原因而并未注意到这些差异:感觉已从差异中解脱出来。
这个事实用来表明在感知觉的范畴内普遍的规律是什么,而且也用来说明我们将来经常有机会涉及的感知觉。我们完全忽略了一个特定感觉的大量不同特征,正是因为它们与相应知觉的客观内容并不直接相关。如果我们正在考虑由某个外部物体产生的一种颜色,我们就尽力在它的特性方面力求正确。但是,当色调中存在某种特殊性时(这种色调与外部印象没有关系),我们只能通过特定仪器的帮助才能感知它,或者通过注意力的异常集中才能感知它。因此,我们几乎可以毫不怀疑地认为,皮肤表面不同区域的感觉差异或视网膜不同区域的感觉差异,实际上远比它们在我们的实验中表现出来的情况大得多。尽管我们可能有所准备,我们仍然倾向于仅仅注意那些由外部印象的本质所决定的感觉特征;即便把我们自己从一种规则(这种规则在整个感觉发展过程中一直存在着,而且对于这种规则我们也必然地和无意识地加以遵奉)中解放出来,单单凭此意图还不足以消除它。相应地,我们不应自我陶醉于以下事实,即一种感觉特性(它有赖于受刺激的感觉器官部位的位置)只能用粗略的方式或者一般的方式来论证。但是,我们可以肯定地假设,这类差异是存在着的,而且在下列情形中颇为见效,在这些情形中,我们的观察方式之缺陷阻止了我们对它们的认识。这是因为,上述事实表明,如果感觉差异被理解为感觉的主观差异,而不仅仅归之于客观刺激的部位差异(local difference),则它一定会变得格外的大。
那么,造成感觉中这些特定差异的原因是什么呢?很显然,原因在于部位;因此,必须在感觉器官的结构中去寻找。我们将音调和色彩的差异最终归结为耳朵和眼睛终端器官的差异。因此,这些深入一步的质的差异(对视觉来说特别如此)必须归之于结构中的微小变化或视网膜终端器官的化学成分。这个假设不是没有事实根据的。观察表明,尤其是对红光的感受性在视网膜边缘趋于减弱。这一事实是有趣的,因为最为常见的部分色盲的类型是红色盲,正如我们前面说过的那样。因此,一般的红色盲可能意味着视网膜边缘部分的正常感受性扩展到了中央部分。
在皮肤方面也存在着许多部位差异,它们可能有助于解释感觉的质的变化与受刺激的器官的部位有关。那些呈灯泡状的、专门处理触觉刺激的终端器官以不同的数量分布着,像视网膜的组成成分一样。但是,它们的数量要多得多,例如,在敏感的指尖部分,比起相对而言不灵敏的背部或大腿来说,专门处理触觉刺激的终端器官要多得多。而且,在表皮(epidermis)的厚度方面也存在进一步的差异;在相邻的皮肤区域,其神经供应方面也存在进一步的差异,它们可能使之产生这样的情况,即在身体表面的不同部位,对同一种印象可以有不同的感受。
现在,我们已经确立了一个事实,它可能有助于我们回答刚才提出的问题。我们已经提出了一些条件,在这些条件下,一种起初完全不规则的反射系统可以产生规则的和一致的运动反射系统。我们已经发现,皮肤和眼睛(它们的刺激是反射运动的主要诱因)呈现出意指感觉中明确的部位差异的结构特性。那么,关于感觉我们该做怎样的推论呢?很显然,其结果将与一种颜色被认做同样的颜色或一种音调被认做同一个音调时我们具有的结果相一致。每一种特定的感觉将根据有赖于印象地点的这种属性被辨认出来;当我们一旦具有感觉位置的经验时,我们便能从属性中辨认出感觉本身的位置。
于是,整组事实引导我们得出个单一的结论:我们具有反射目的性发展的主要条件。该发展由下述内容组成——原先无目的的一种运动开始有了明确的目标,目标就是受外部印象刺激发出反射的感受点。该点可由反射运动来发现,对它来说,在每一种特定的情形里被辨认出来是很有必要的。正如颜色和音调因其感觉可以辨别或不可以辨别而被理解为相似或不同一样,一种印象的位置也只能依靠明确的感觉特征(专门有赖于该位置的特征)进行辨认。我们已经表明,确实存在这些特征。借此证据,我们可以提供反射的规律性和一致性的第一条件:它必须在器官本身的结构中去寻找。与此同时,我们关于反射的发展尚未提供完整的和令人满意的描述。我们发现运动始终以最短和最简单的路径通向其目的,对此,感官的结构当然无法解释。它只能告诉我们反射如何具有目的,而不是反射如何达到目的。关于这方面的进一步解释,可在运动本身中发现。因此,我们必须继续询问,是否能够容许任何一种针对性的解释,以及它的条件是什么。
