第8章 四倍体或四倍型
另一说法似乎更为可能,即正在分裂中的细胞,其胞质分裂受到抑制,于是染色体数目加倍,结果产生了四倍体。这样的四倍型细胞或者形成幼小植株的全体,或者形成其中心柱,或者形成其他任何部分。
那不勒斯动物研究所的一间实验室的内部(1908年左右)。19世纪的生物学家多是研究采集来的生物标本,而那不勒斯研究所则注重活体机能的研究。
计算过染色体数目的有一千多种动物,或者也有同样多或者更多的植物。其中两三个物种只有一对染色体。在另一极端,也有物种含一百多条染色体的。不管染色体多少,每一物种的染色体数目是恒定的。
染色体有时确是不规则的分布。其中,大多数通常借某种方法自行矫正。也确有一二个例子,其染色体数目略有变化。例如Metapodius可能有一条或多条额外的小染色体,有时是几条Y染色体,有时是另一条称为M的染色体(图62)。正如Wilson所指明的,这些染色体在个体性状上,既然没有引起相应的变异,所以不妨把它们当做无关轻重的、不活动性物种看待。
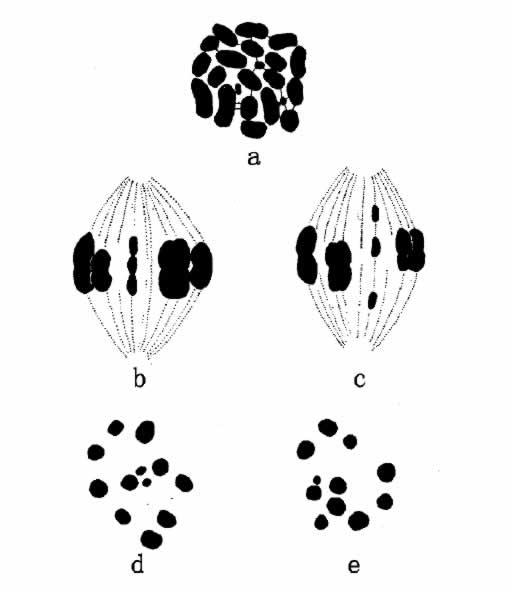
图62 示Metapodius的染色体。a.示精原细胞的染色体群,有三条m染色体;
b和c示精母细胞的侧面观;d和e示三条m染色体接合,两条趋入一极,一条趋入另一极(c的末期赤道板)(仿Wilson)
其次,我们知道,几条染色体可以彼此连接,从而减少一条或多条染色体,一条染色体又可以裂断,从而暂时增加一条(注:Hance描写过待霄草的染色体有时断裂成片。在灯蛾(Phragmatobia)和其他蛾类中,Seiler也描写过几个例子,其中某些染色体原来在精子和卵细胞内连接一起的,在胚胎细胞内却彼此分开。在蜂类的所有体细胞内,每一条染色体都被假定是断裂成两段的。在蝇类及其他动物的一些体细胞中,染色体可以分裂而细胞不分裂,从而使染色体的数目增加到二倍或四倍。),在任一种情况下,全部基因仍然被保存下来。最后,有些物种,雌者比雄者多一条染色体,另有一些物种,情形刚刚相反。所有这一切情况都经过了广泛的研究,而且是每一位研究细胞的学者所熟悉的。这类例子的存在并不使每一物种各有其特殊数目的染色体,而且染色体数目恒定不变(注:近几年来,Della Valle和Hovasse不承认在不同的组织细胞里染色体的恒定数目。这项结论是以两栖类身体细胞方面的研究为根据,但两栖类的染色体数目繁多,不容易精密辨别,所以他们的研究结果不足以推翻其他生物(甚至包括一些两栖类在内)方面的绝大多数的观察,在这些生物里,染色体的数目是能够精密决定的。)这个一般论点,归于无效。
我们也知道:在某些组织里,由于只有染色体分裂而无细胞分裂,或者由于染色体断裂为一定数目的部分,可以把染色体增加到二倍或四倍的数目。这些都是特殊的例子,不足以影响一般情况。
有一些个体突然出现,其染色体数目为该物种特有数目的两倍,这种例子,在近年来愈来愈多。这便是四倍体。又发现了其他多倍型,有的自然发生出来,有的起源于四倍体。我们总称之为多倍体。其中以四倍在许多方面最引人关注。
确实知道的四倍体动物,只有三个例子。寄生在马体内的线虫,即马蛔虫,共有两型,一型含两条染色体,另一型含四条。这两个变种彼此相似,甚至细胞的大小也相仿佛。蛔虫的染色体可以看成是一种复合体,是由许多较小的染色体(有时称之为染色粒)联合组成。在应该形成体细胞的胚胎细胞内,各染色体断裂成它的组成部分(图63a、b和c)。成分的数目恒定,或接近一个常数,二价型成分的数目约为单价型的两倍。这支持了这样的一种看法,认为二价型比单价型多一倍,而不是二价型由单价型染色体的分裂而来。
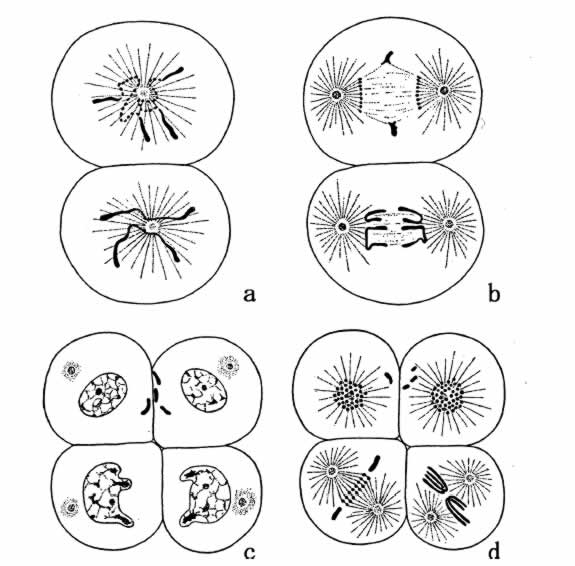
图63 单价型蛔虫的卵子含两条染色体,图示其最初两次分裂。a和b示染色体在一个细胞内的断裂作用;d示三个细胞的染色体已经断裂,第四个细胞的染色体依然完整,后者产生生殖细胞(仿Boveri)
根据Artom报道,海虾Artemia有两族的四倍体,一族有48条染色体,另一族有84条(图64)。后一族借单性生殖法繁殖。在这些情况下,不难想象,四倍体起源于原来是单性生殖的一族,因为,如果卵细胞保留一个极体,以致染色体数目加倍,或者由于在胞核第一次分裂后,染色体未能分离开来,以致数目增加一倍;这种双倍状态都是可能继续存在的。
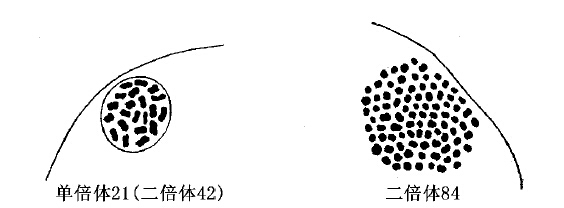
图64 Artemia salina二倍体(减数)与四倍体两族的染色体(仿Artom)
最早出现的植物四倍体中,有一种被德弗利发现,名叫巨型待霄草Oenothera gigas(图42)。最初并不知道巨型是四倍体,但德弗利看出它比杂种(拉马克待霄草)植株强壮,在其他许多细微特征上也有差异。巨型的染色体数目后来才弄明白。

图65 a.拉马克待霄草的14条二倍染色体;
b.巨型待霄草的28条二倍染色体拉马克待霄草(Oenothera Lamarkiana)有14条染色体(单倍数7)。巨型待霄草(Oenothera gigas)有28条染色体(单倍数14)。图65表示上述两种植物的染色体群。
Gates测量过各种组织的细胞。巨型药囊的表皮细胞几乎为普通型体积的四倍,柱头的表皮细胞为三倍,花瓣表皮细胞为两倍,花粉母细胞比普通型约长一倍半。花粉母细胞的胞核为普通型体积的两倍。两型的细胞在外形上也有时有显著的差别。各种待霄草大多产生三叶的盘状花粉,而巨型的一些花粉则为四叶状。
Gates、Davis、Cleland和Boedijn研究过花粉母细胞的成熟过程。据Gates报道,拉马克待霄草的巨型通常有14对二价染色体(gemini),在第一次成熟分裂时,每条二价染色的两半分别走入一个子细胞。在第二次成熟分裂时,每一条染色体纵裂为两条,于是花粉粒各得14条染色体。据推想,在胚珠成熟中也发生了同样过程。Davis讲到当拉马克待霄草的染色体从联会混乱状态中出现时,染色体参差不齐地互相集结,而不是严格平行联合。随后,染色体分别趋入一极,完成了减数分裂。Cleland近来也讲过另一个二倍型物种Oenothera franciscana的染色体,在进入成熟纺锤体时,相互间端与端相连(图66)。在Davis早期发表的图内,有一些也略有端与端相连合的情况。

图66 Oenothera franciscana花粉母细胞的成熟过程(仿Cleland)
近年来在其他雌雄同株的显花植物中,也找到了一些四倍体。因为这类植株同时产生卵子和花粉,所以显然应该比雌雄异株的植物形成更多的四倍体。因此,如果植株一开始便是四倍体,那么,它会产生有着二倍数目的染色体的卵细胞和花粉细胞。经过自体受精,势必产生四倍体。相反,在雌雄异株的动物和植物中,一个个体的卵子必须同另一个体的精子受精。如果发生了一个雌性四倍体,其成熟卵子有二倍数目的染色体,卵子一般会同普通雄体的单倍型精子受精,结果只能产生三倍体。要从三倍体恢复到四倍体,机会是很少的。
谱系培养中所产生的四倍体,在说明四倍体的起源上,比偶然发现的四倍体,提供了更精确的知识。事实上,四倍体在人工控制下发生的例子,已经有了一些记录。Gregory发现了两种巨型的报春花primula sinensis:一种出现在两株二倍体植株的杂交里。因为亲株含有已知的遗传因子,所以Gregory能够研究四倍体中各种性状的遗传过程。根据他的研究结果,他无法决定:在四条相似的染色体中,某一条染色体究竟是同其他三条中的另一条特殊染色体联合,或者是同其中任何一条染色体联合。Muller分析同一数据,指出后一结论更为可能。
Winkler通过嫁接的媒介作用,得到一株巨型龙葵,Solanum nigrum和一株巨型番茄(Solanum lycopersicum)。就我们所知道的,嫁接作用本身对于四倍体的产生是没有直接关系的。
产生龙葵四倍体的方法如下。取番茄幼株的一段,嫁接在龙葵的幼茎上面,摘除龙葵所有的腋芽。10天后,沿嫁接平面横切(图67)。从切面的愈伤组织中,长出了一些不定芽。由此长成的植株中,有一株是嵌合体,也就是一部分组织为龙葵,一部分组织为番茄的植株。把嵌合体取下,并让其繁殖。新植株的腋芽中,有些由番茄的表皮和龙葵的中心柱组成。把这些枝条取下,另行栽培。这些幼小植株同其他确是二倍体的嵌合体有所区别,使人怀疑这种新型也许有一个四倍型的中心柱。检查结果,证实了这一点。截除嵌合体的顶梢,并且摘除下半部的腋芽。从愈伤组织的不定芽里,得到了全身都是四倍型的幼株。图68右侧示巨型龙葵,左侧为普通型(二倍体)或亲型。图69的右上示巨型的花,左侧为亲型的花。同图的左上示巨型和新型的苗木。
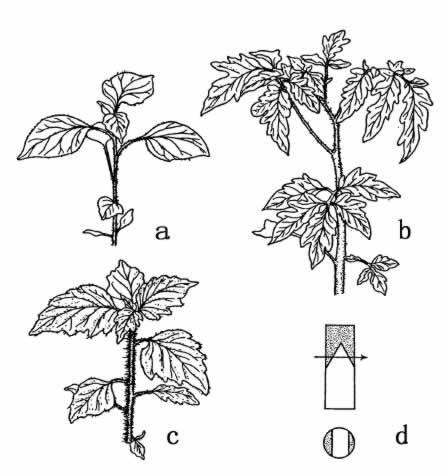
图67 a.龙葵的苗;b.番茄的苗;c.Solanum tubingense.的嫁接杂种;d.嫁接方法(仿Winkler)
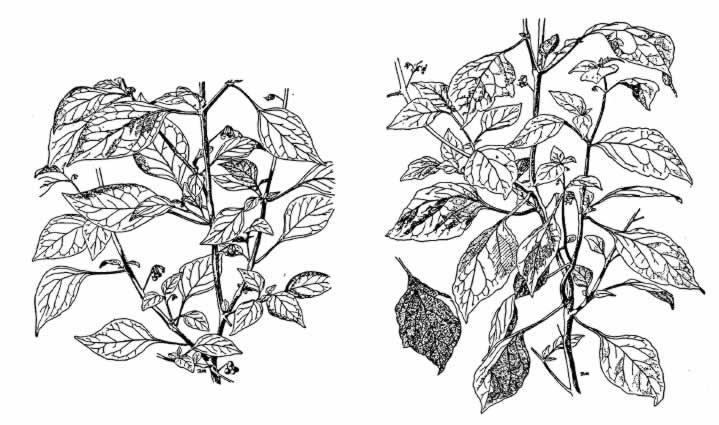
图68 左侧为亲型龙葵的普通二倍体;右侧为四倍体(仿Winkler)
图69下半部示若干组织细胞的差异。左侧示巨型叶和亲型叶的栅状细胞;右侧示两型气孔的保护细胞;其下则是两型的毛;巨型的髓细胞也比普通型大些。图下半部的中央左边为亲型花粉粒,右边为巨型花粉粒。
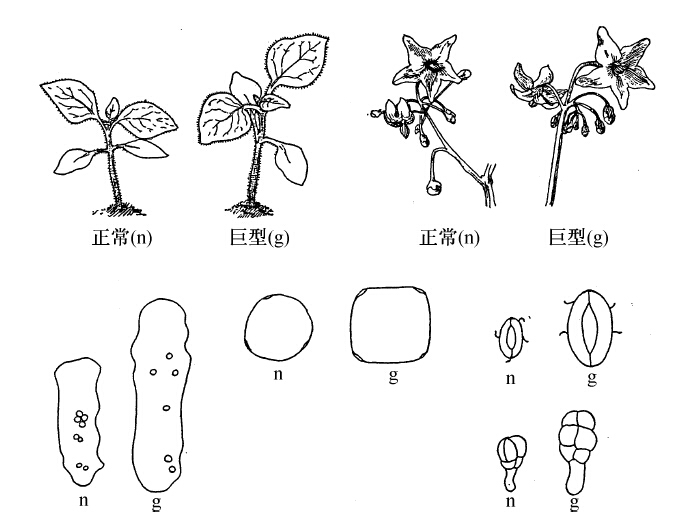
图69 上部示龙葵的二倍体和四倍体的苗木和花,下部示它们的组织。左上,苗木;右上,花;左下,栅状细胞;中,花粉粒;右下,上为气孔,下为毛(仿Winkler)
用以下方法也得到了一株四倍体番茄。取番茄幼株的一段,按照常用的方法嫁接在龙葵砧木的上面(图67)。待其完全联合以后,横切两种植物联合的部分,并且摘除砧木上的腋芽。从愈伤组织里长出了幼芽。移植这些幼芽。其中有一株由龙葵的细胞构成表皮,由番茄的细胞构成中心柱,进一步检查,发现表皮细胞为二倍体,中心柱细胞则为四倍体。为了从嵌合体造出全身都是四倍体的植株,于是横切嵌合体的茎,并摘除切面以下的腋芽。切面上发生新的不定芽,芽体内外大部分由番茄组织构成。巨型番茄植株和其亲型植株间的差异,同巨型和亲型龙葵间的差异,是一样的。
二倍体龙葵含24条染色体,其单倍数为12;四倍体含48条染色体,其单倍数为24。二倍体番茄含72条染色体(单倍数为36);四倍体含144条染色体(单倍数为72)。图70和图71表示各型染色体。
像上面所提到的,直到现在我们还不知道嫁接对于愈伤组织内发生四倍型细胞,有什么明显的关系。这些细胞究竟怎样发生,也不明确,也可能像Winkler一度相信的那样,是由于愈伤组织中两个细胞的融合,但另一说法似乎更为可能,即正在分裂中的细胞,其胞质分裂受到抑制,于是染色体数目加倍,结果产生了四倍体。这样的四倍型细胞或者形成幼小植株的全体,或者形成其中心柱,或者形成其他任何部分。
Blakeslee、Belling和Farham在常见的曼陀罗(Datura stramonium)里发现了一种四倍体(图72下部)。就外形看来,它被描述为在几个方面同二倍型有差别。图73示二倍体(第二行)和四倍体(第四行)的蒴果、花以及雄蕊方面的区别。
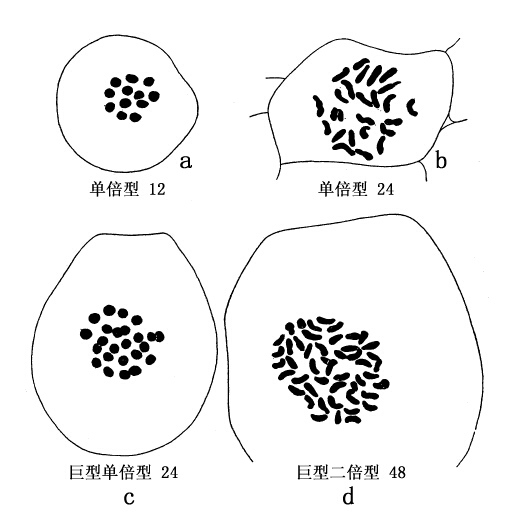
图70 a和b示龙葵的单倍型和二倍型细胞及染色体,c和d示四倍体龙葵的单倍型和二倍型细胞及染色体(仿Winkler)
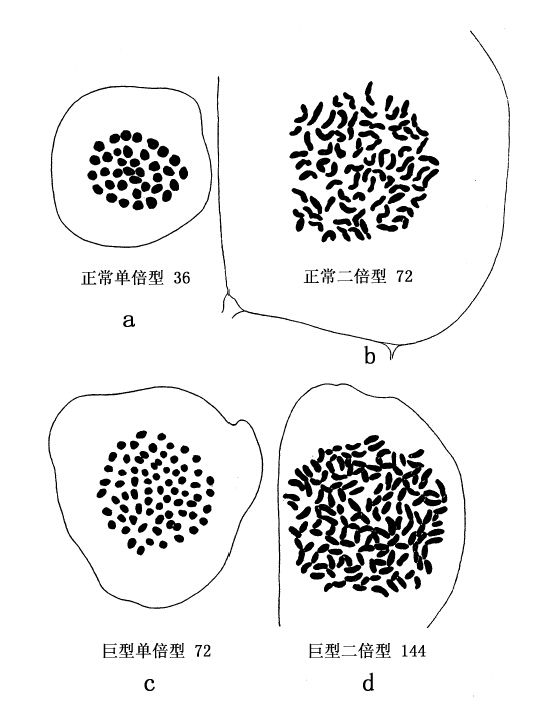
图71 a和b示番茄的单倍型和二倍型细胞及染色体,c和d示四倍体番茄的单倍型和二倍型细胞及染色体(仿Winkler)

图72 上行示曼陀罗的二倍体植株;下行示其四倍体(仿Blakeslee)

图73 曼陀罗单倍体、二倍体、三倍体和四倍体巨型的蒴果、花和雄蕊(仿Blakeslee,载在《遗传学》杂志上)
二倍体植株含12对染色体(24条染色体),按照Belling和Blakeslee的意见,这些染色体依体积大小可以安排为六型(图74),即大号(Ll)、中号(Mm)、小号(Ss)或2(L+4l+3M+2m+S+s)。单倍染色体群的方程式为L+4l+3M+2m+S+s。当这些染色体将进行第一次成熟分裂时(前期),它们形成一对一对的环状结构,或者以一端相连接(图75第二列)。以后在每一对中,一条接合体走到一极,另一条则趋向另一极。到第二次成熟分裂之前,每条染色体中缢,从而产生了图74a(注:原文为图74b,但根据作者的叙述,应该是图74a,译文中已予修正。——译者注。所示的形式。一半中缢后的染色体进入纺锤体的一极,另一半则进入另一极。每个子细胞各得12条染色体。

图74 a.曼陀罗二倍染色体群的第二次成熟分裂中期;b.曼陀罗四倍体(含24条染色体)相应的染色体群(仿Belling和Blakeslee)
四倍体有24对或48条染色体。在它们进入第一次成熟分裂纺锤体以前,它们每四条集合一起(图76和图75)。从两图中可以看到四价群内染色体联合的各种不同方式。染色体大致在这种状态下进入第一次成熟纺锤体。在第一次成熟分裂时,每个四价染色体的二条染色体趋入一极,另两条趋入另一极(图75)。每颗花粉粒有24条染色体。不过偶尔也可能有三条染色体同入一极,另两条趋入另一极(图75)。每颗花粉粒有24条染色体。不过偶尔也可能有三条染色体同入一极,另一条则趋入另一极。
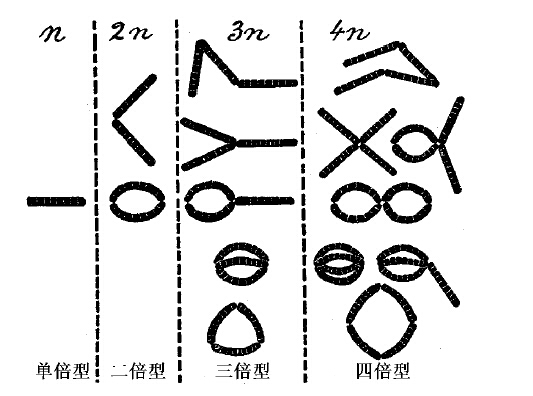
图75 曼陀罗二倍体、三倍体、四倍体各型染色体的接合方法(仿Belling和Blakeslee)

图76 曼陀罗四倍体的染色体互相接合,四条相同的染色体联合组成一个群(仿Belling和Blakeslee)
图75示第二次成熟分裂时四倍体的24条染色体。这些染色体和同一时期中二倍体的染色体相仿佛。每条的一半趋入一极,另一半则趋入对极。据Belling记载,有规则地分布的,即每极各得24条染色体(24+24)的,占68%;一极23条,另一极25条的(23+25),占30%;一极22条,另一极26条的,占2%。分布为21—27的,有一个例子。以上结果表示,在四倍体曼陀罗中,染色体不规则的分布,并不是少见的。这一点可用四倍体自体受精来进一步检验。让这样产生的子代生长成熟,并且计算其生殖细胞内染色体的数目。有55株植物,各含48条染色体,5株各含49条,一株含47条,另一株含48条(?)。如果染色体在卵细胞内像在花粉细胞内一样分布,结果含24条染色体的生殖细胞最可能生存下来,并且有作用。含48条以上的染色体的一些植株,由于增加了额外染色体,也许会产生染色体分布更不规则的新型。
De Mol发表了一种四倍体水仙(Narcissus)。其二倍体物种有14条染色体(七对),另有栽培的两个变种则各有28条染色体。De Mol指出,直到1885年为止,主要栽培的是矮小的二倍体变种。以后才出现了较大的三倍体;最后,约在1899年,又得到了第一株四倍体。
据Longley报道:墨西哥大刍草中,多年生teosinte所含染色体,为一年生teosinte染色体的两倍。多年生型有40条染色体(n=20)(图77a)。一年生型有20条(n=10)(图77c)。

图77 a.多年生墨西哥大刍草(Euchlaena)第一次成熟分裂前期;有19条二价染色体和2条单染色体;a1 .第一次分裂中期;a2 .第一次分裂后期。b.玉蜀黍第一次成熟分裂前期,有10条二价染色体;c.墨西哥Euchlaena mexicana第一次成熟分裂前期,有10条染色体;ab.多年生Euchlaena同玉蜀黍所生的杂交一代的杂种,第一次成熟分裂前期,有3条三价染色体,8条二价染色体和5条单染色体;ab1 ,同上,第一次成熟分裂后期的晚期(仿Longley)
Longley把两种teosintes同含20条染色体(n=10)的玉蜀黍(图77b)分别杂交。一年生teosinte和玉蜀黍所生的杂种有20条染色体。新种的花粉母细胞成熟时,有10条二价染色体,二价染色体各自分裂,分别趋入两极,没有停滞在中途的。这意味着从teosinte得来的10条染色体,同从玉蜀黍得来的10条染色体,互相接合了。又多年生teosinte同玉蜀黍杂交的杂种有30条染色体。杂种花粉母细胞成熟时,其染色体互相接合,有两条一起的,有三条一起的;也有孤立无偶的(图77ab)。由此引起了后来分裂中的错乱情况(图77ab)。
当雌雄同株的植物性别决定问题不涉及分化性的性染色体时,它的四倍体可说是又平衡、又稳定。所谓平衡,就是说:它的基因之间的数字关系,同二倍体或普通型基因的数字关系,是一样的。所谓稳定,就是说:成熟机制能够使该型一旦建立以后会继续存在(注:Blakeslee对于这些词汇有不同的解释。)。
早在1907年,Elie和Emile Marchal便已用人工方法产生了四倍体藓类。每株藓有两个世代:一个是产生卵子和精子的单倍型原丝体时期(配子体),和一个产生无性孢子的二倍体时期(孢子体)(图78)。
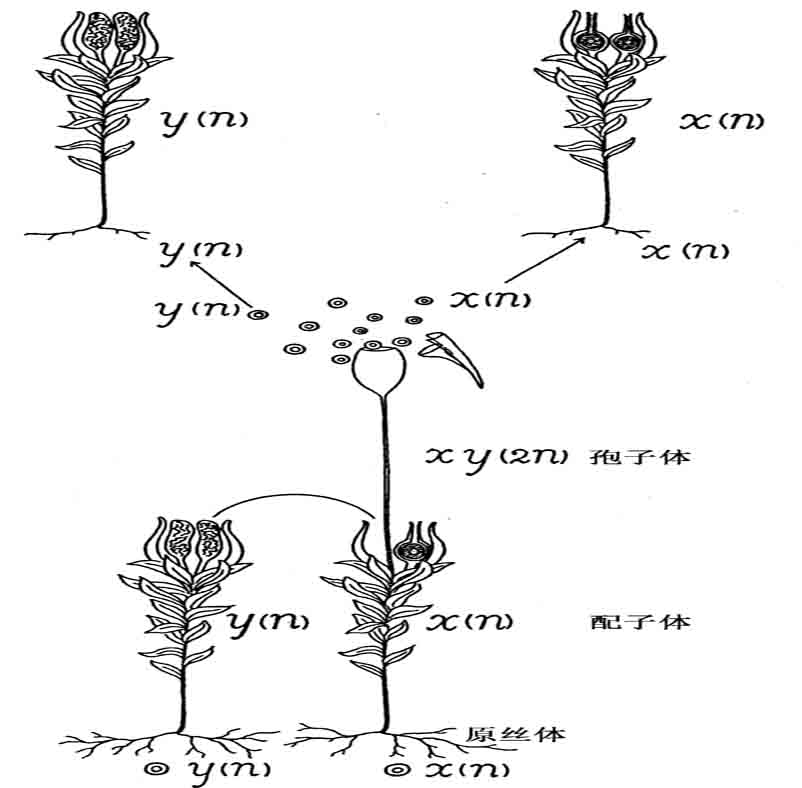
图78 雌雄同株的藓类的正常生活史
孢子体的一段在潮湿情况下,会产生由二倍型细胞组成的细丝。细丝发育成真正的原丝体,后者迟早会产生二倍型卵子和二倍型精细胞。两种生殖细胞联合,于是形成了四倍型孢子体(图79)。这里,正常单倍型被二倍型原丝体和藓的植株重复一次;二倍型孢子体又被四倍型孢子体重复一次。
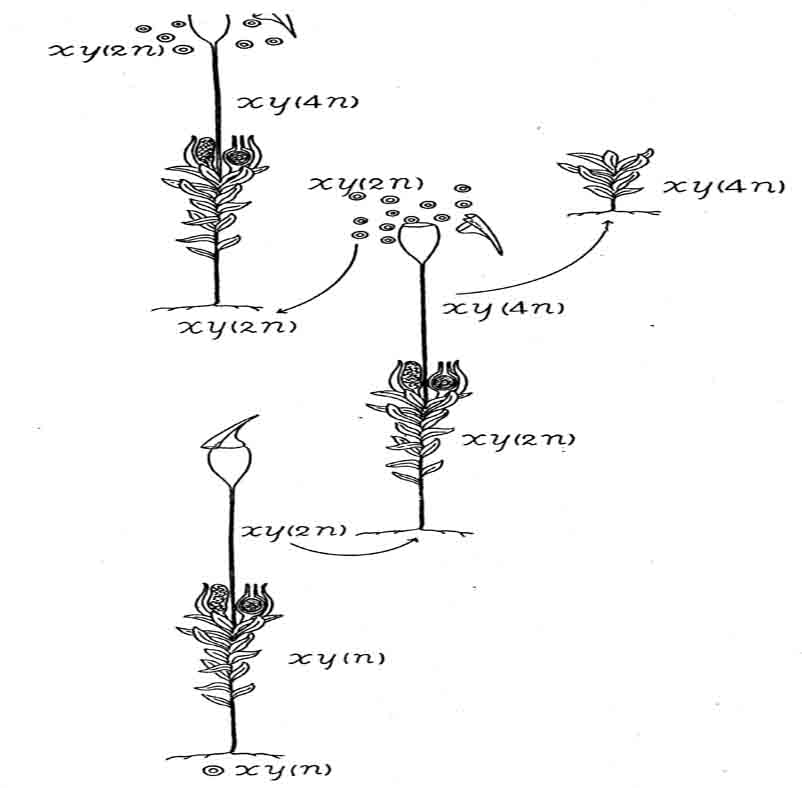
图79 正常雌雄异株的藓类,一段2n的胞子体再生,形成二倍型原丝体(2n),2n配子体通过自交,产生了四倍体或4n孢子体,四倍体再生,产生了四倍型配子体(仿Winkler)
两位Marchals对普通型和四倍体两者的细胞体积,作了比较测量。正常花被细胞的体积与四倍体同样细胞的体积有三种比例:1∶2.3,1∶1.8和1∶2。两型的精子器细胞体积比为1∶1.8,胞核体积比为1∶2。卵细胞体积比为1∶1.9。测量精子器(内藏精子)和颈卵器(内藏卵子),也表示在所有例子里,四倍体比正常型长些、宽些。显然,四倍体体积之所以增大,是由于有了较大的细胞,细胞之所以增大,又由于有了较大的胞核,而且像其他证据所已经证明过的,四倍体大核内的染色体又为正常型的二倍。这自然是在意料中的,因为四倍体是由正常孢子体再生出来的。
在孢子体世代中,2n孢子母细胞同4n孢子母细胞两者的体积比约为1∶2。
藓类的两次成熟分裂,即紧接着染色体接合后的两次分裂,发生在孢子体内孢子形成的时候,每个孢子母细胞产生四个孢子,藓类的染色体上如果带有基因,预料四倍体所有的加倍的染色体将会产生不同于正常型的比率。虽然Wettstein在几种藓类杂交中找到过遗传的显明证据,Allen在亲缘相近的几群藓类中也得到了配子体两种性状的遗传学证据,但是这方面的研究仍然是很少的。
在雌雄异体的藓类以及在某些苔类里,Marchals、Allen、Schmidt和Wettstein已经分别证明:在孢子形成的时候,与性别决定有关的要素分离开来。他们的观察和实验将于性别一章中讨论。
有许多涉及四倍体细胞体积方面的重要问题,是胚胎学上的问题,而不是遗传学上的问题。四倍体的细胞一般较大,往往大到两倍,不过组织不同,细胞大小也颇悬殊。
四倍体整个植株的大小以及其他一些特性,似乎都是由于细胞增大的缘故。如果这种解释是对的,这便意味着,这些特性属于发育范畴,而不属于遗传范畴。四倍体发生的方法,已经有了一些讨论。以上所提出的关于四倍体细胞内如何增加胞质的方法,尚有待于进一步的研究。
假设同一物种的两个细胞彼此融合,两个胞核也迟早联合,也许由此会产生四倍型细胞。假设四倍型细胞在生长期内,继续维持其两倍大的体积,预料会得出比正常型体积大两倍的卵子。大型胚胎的细胞同正常胚胎特有的细胞,在数目上预料是一样多的。
另一个可能的解释,即四倍型生殖细胞的体积,在二倍型母株的生殖细胞内,或者不能增加到两倍。这样,卵子也将不会大于正常卵子,但却带有两倍多的染色体。从这种卵子发育而成的胚胎,在达到能够从外界获得食物的胚后期或幼虫期以前,也许不能摄取足够的营养来增加它的细胞体积。至于在这样晚的时期,每个细胞内的双组染色体是否会扩大各该细胞的胞质,还是不确定的。不过下一代的卵子在母体内开始发育时,便有了四倍染色体,在这种情况下,卵子在分裂前增加到两倍的体积,也是可以想象的。
如果说在受精以后成熟卵子中,随着染色体数目的加倍,胞质也会立刻增加起来,这或者是不可期望的。在动物胚胎开始形成器官以前,它经历了相当一定次数的细胞分裂。如果胚胎一开始便是一个体积正常而染色体加倍的卵子,如果由于染色体的双倍数目以致卵裂时期比正常卵子提早结束,从而开始了器官形成,那么,这样的四倍体胚胎的细胞将会比正常胚胎的细胞大两倍,但数目却只有一半。
在显花植物里,胚囊空阔,并富于养料,也许为卵子发生大量的胞质提供了更有利的机会。
