藓类的性别决定
在上述显花植物性染色体被发现的几年以前,两位Marchals证明:在雌雄异体的藓类植物里(配子体分雌雄两种(注:苔藓和蕨类的单倍体世代(或称配子体世代)分为雌雄两种,其二倍体(或称孢子体)世代无雌雄之别,或者说是中性。显花植物本身相当于藓类的孢子体。其配子体世代好像是深藏在雌蕊和雄蕊里面。所以雌雄二词,用于藓类方面,则指单倍体世代而言,用于显花植物方面则指二倍体世代而言。两者的意义互相矛盾。但是这个矛盾不是二倍体和单倍体问题(因为甚至在某些动物如蜜蜂、轮虫等的同一世代里,也碰到这个矛盾),而是有性世代和无性世代同用了雌雄这两个词。不过,有了这样的理解,以后采用这种习惯用法就不会发生严重困难了。)),由同一孢子母细胞所产生的四粒孢子中,有两粒发育成雌配子体,另两粒发育成雄配子体。
稍后,Allen在亲缘关系相近的苔类植物里,(图124)发现了单倍体的雌原叶体(配子体)有八条染色体,其中最长的一条为X;单倍体的雄原叶体(配子体)也有八条染色体,其中最短的一条为Y(图124b′)。这样,每个卵子有一条X,每条精子也有一条Y。由受精卵发育而成的孢子体,有16体染色体(包括X和Y各一条)。减数分裂发生在孢子形成的过程里,X和Y分开。半数的单倍型孢子各有一条X,以后发育成雌原叶体,另一半各有一条Y,以后发育成雄原叶体。
最近,Wettstein用雌雄异株的藓类植物进行若干精密实验,作出了进一步的分析。他袭用Marchals所发现的一种方法,造成了具备雌雄两群染色体的配子体(图125左侧)。例如,他仿照Marchals的方法,截取一段载有孢子的柄部(细胞为二倍型)。由该段发育而成的配子体也是二倍体。这样获得了雌雄兼备(FM)(注:F代表雌性,M代表雄性。——译者注。)的配子体。
Wettstein用另一种方法造成二倍体的雌藓和雄藓,成为双重雌性(FF)或双重雄性(MM)。方法如下:Wettstein用水合氯醛和其他药剂处理原丝体,使一个个别细胞的胞质,在染色体分裂以后,受到抑制。这样,他就能够在这些雌雄异株的植物里造出二倍型的巨大细胞,各有二重的雌性要素或雄性要素,例如染色体。从这样的二倍型细胞,又造成了几种新组合:其中有些是三倍体,有些是四倍体。图125右侧表示最有趣的几种组合。
雌原丝体的一个二倍型细胞发育成二倍体植株(FF),后者又产生二倍型卵细胞。一个二倍型雄原丝体的细胞也同样发育成MM植株。一个FF卵子同一条MM精子相遇,结果产生了一株四倍型孢子体(FFMM)。
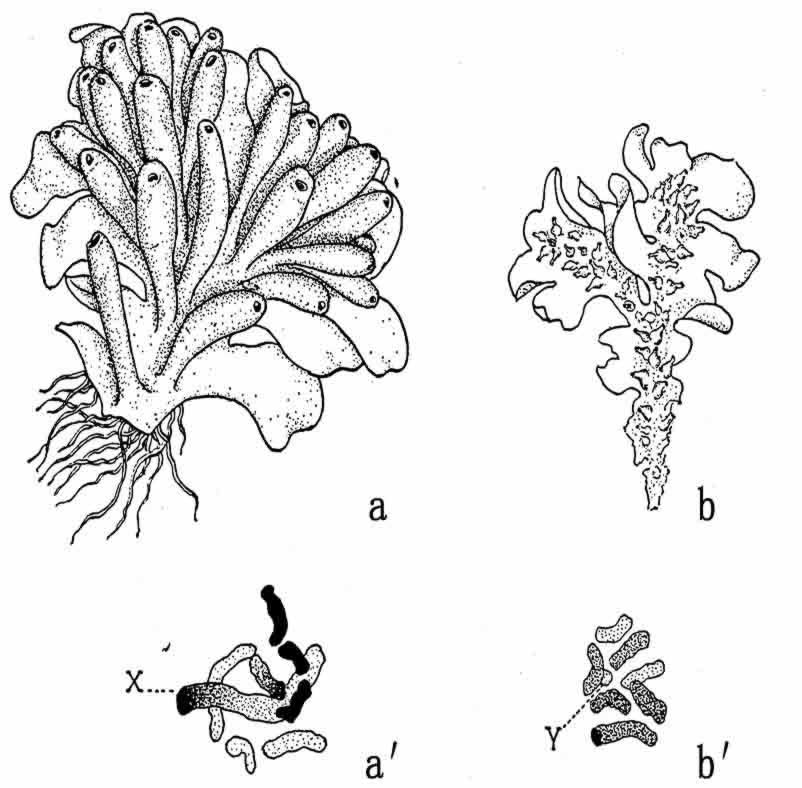
图124 a.苔类的雌原叶体;b.雄原叶体,a′.雌性有一条大的X染色体;b′.雄性有一条小的Y染色体(仿Allen)
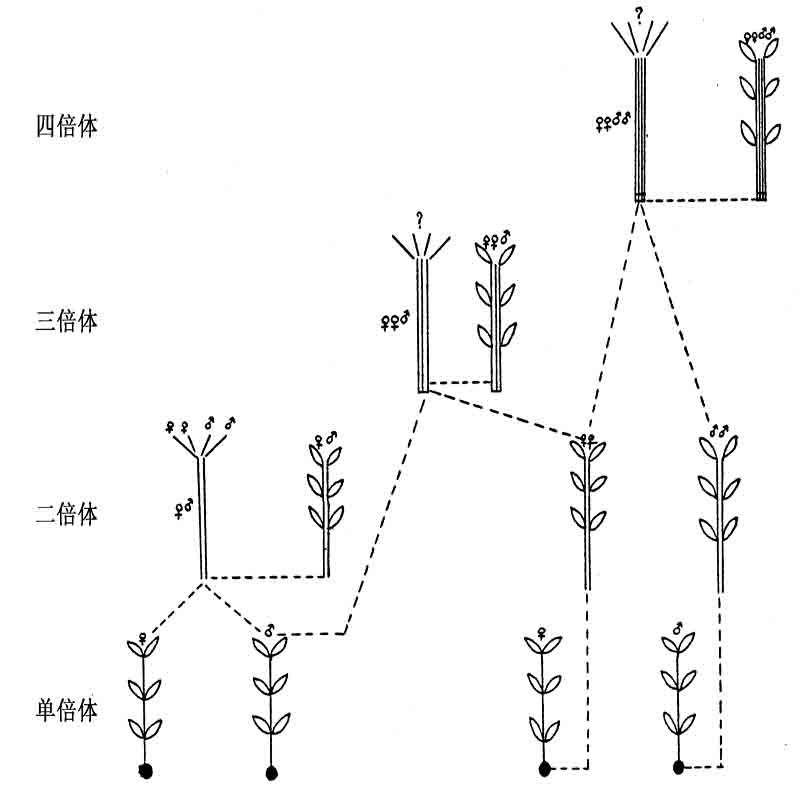
图125 示藓类二倍体和三倍体的各种不同组合(仿Wettstein)
一个FF胚珠同一条正常雄精子(M)受精,结果产生了三倍型植株(FFM),如下表:
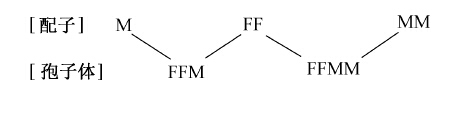
从FFM或FFMM的孢子体又能够再生出配子体。这些配子体都能够发生雌雄两种要素,也都能够产生卵子和精细胞;不过雌性器官(颈卵器)和雄性器官(精子器)的多少及其出现的迟早,则表现出特殊的差别。
上面谈到两位Marchals,在Wettstein所用的同一物种里,得到了二倍型的FM配子体,并且证明了该配子体产生雌雄两性的器官。Wettstein证实了这个事实,并且报道雄性器官发生在雌性器官之前。
比较一下FM、FFM、FFMM三型,是有意义的。FM植物的雄性器官成熟极早。开始时精子器比颈卵器多多了。颈卵器发生较晚。
像Wettstein所说的,FFMM植物雄性器官的早熟性,比FM型植物的要强两倍。开始时只出现精子器。在这一年很晚的时候,衰老的精子器已经凋落,这时才有少数颈卵器出现;有一些植株根本不发生颈卵器。更晚一些,雌性器官才开始发育旺盛。
三倍型植物中,雌性器官首先成熟。至少是当四倍体只有雄性器官的时候(七月间),三倍体还只有雌性器官,以后(九月间)才具备雌雄两种器官。
有趣的是:这些实验表示了原来是雌雄异体的植物,经过雌雄两种要素的联合,可以成为人工的雌雄同体的植物。这些结果也表示了性器官发生的先后,决定于植物的年龄。更重要的是,两种性器官发生时间上的关系,由于遗传组合循相反方向改变而颠倒起来。
