第二部分 性选择
·Sexual Selection·
我将在以下几章讨论属于各个纲的动物的第二性征,并努力把本章所阐明的原理应用于每个事例。我们用于讨论最低等动物纲的时间将很短,而对于高等动物,尤其是对于鸟类,则必须用相当的篇幅详加讨论。请注意,由于已经说明的理由,关于雄者用以寻求雌者并在寻得后牢牢把它抓住的无数构造,我只想举出少数例子用做说明。另一方面,关于雄者用以战胜其雄性对手的以及用以魅惑或刺激雌者的全部构造和本能,将予以充分讨论,因为它们在许多方面都是最有趣的。

▲巴特利特先生见过一只在求偶活动中的雄孔雀雉,并给我看了按当时姿态制作的一个标本。这种鸟的尾羽和翼羽都饰以美丽的眼斑,就像孔雀尾羽上的眼斑那样。当雄孔雀夸示自己时,便展开并竖起其尾羽,使其同躯体相横切,这是因为它站在雌鸟之前,需要同时显示其鲜蓝色的喉部和胸部。
第八章 性选择原理
第二性征——性选择——作用方式——雄者的过剩——多配性——性选择通常只使雄者发生变异——雄者求偶的热望——雄者的变异性——雌者对配偶的选择——性选择与自然选择的比较——在生命相应时期出现的遗传性,在一年相应季节出现的遗传性,限于性别的遗传性——若干遗传形式之间的关系——雌雄任何一方以及幼小动物为什么没有通过性选择而发生改变的原因——有关整个动物界雌雄两性比例的补充说明——同自然选择有关的雌雄两性比例
凡是雌雄异体的动物,雄者的生殖器官必然与雌者不同,这就是第一性征(primary sexual characters)。但雌雄的区别常常表现在亨特所谓的第二性征上(secondary sexual characters),而它们同生殖行为并无直接关系。例如,为了易于寻找或接近雌者,雄者具有某些感觉器官或运动器官,这是雌者所没有的,或比雌者的这等器官更为高度发达;又如,为了将雌者牢牢抓住,雄者具有特殊的抱握器官。这样器官的种类形形色色,无穷无尽,进级为通常被列为第一性征的那些器官,而且在某些场合中,它们同第一性征的器官几乎无法区别;在雄性昆虫腹端的复杂附属物中我们见到许多这方面的例证。因而除非我们将“第一的”(primary)这个术语的含义限于生殖腺的范畴,否则就几乎不可能决定何者是第一性征,何者是第二性征。
雌者同雄者的区别往往在于前者具有营养其后代或保护其后代的器官,譬如哺乳动物的乳腺以及有袋类的腹袋。在少数场合中,却是雄者具有类似的器官,而雌者没有,譬如某些雄鱼的储卵囊,这等器官在某些雄蛙身上也有暂时的发育。大多数蜂类的雌者具有一种采集和携带花粉的特殊工具,它们的产卵器也演变成一根蜇针,用于保卫幼虫和群体。还可以举出许多相似的事例,但与本文无关。然而,另外一些雌雄差异尽管同初级生殖器官毫无关联,却正是我们所特别注意的——譬如雄者的较大体型,力量,好斗性,用于对竞争对手进攻的武器或防御的手段,绚丽的色彩和各种装饰,鸣唱的能力以及诸如此类的其他性状。
除了上述的第一性差异与第二性差异以外,某些动物雌雄二者的构造差异是和不同的生活习性有关,而完全不是或仅是间接地和生殖功能有关。像某些蝇类(蚊科,Culici dae和虻科Tabanidae)的雌者都是吸血虫,而雄者则以花为生,其口器缺少上颚。 (1) 某些蛾类和一些甲壳类(异足虫Tanais)的雄者具有不完备而封闭的口器,不能取食。某些蔓足类的补雄(complemental males)就像附生植物那样,或依雌者为生,或依两性体为生,它们既没有口器也没有抱握肢。在这等场合中,是雄者发生变异并失去雌者所具有的某些重要器官。在另外一些场合中,则是雌者失去了这等器官;例如,雌萤火虫无翅,许多种雌蛾也如此,其中有些甚至永远没有脱茧而出。许多寄生甲壳类的雌者已失去它们用于游泳的后肢。有些像甲科(Curculionidae)的象甲虫,其雌雄二者的喙长有巨大差异, (2) 但这种性差异以及许多与此相似的差异的意义何在,尚属不明。与不同生活习性有关的雌雄二者之间的构造差异,一般只限于低于人类的动物;但关于少数一些鸟类的喙,雄者和雌者的却不相同。新西兰辉雅鸟(Huia)的这种差异非常之大,我听布勒博士说, (3) 那种雄鸟用坚固的喙从腐朽树木中凿取昆虫的幼虫,而雌鸟则用长得多的而且非常柔软易弯的喙在树木较柔软的部位探求之,它们就这样彼此互助。在大多数场合中,两性之间的构造差异多少都同种的繁殖有直接关系,因而一个雌者为了给大量的卵供应养分,需要比雄者更多的食物,所以需要特殊的取食手段。一只生命很短的雄性动物的取食器官由于不使用而消失,并不会带来什么损害;但要保持完善状态的运动器官,以便接近雌者。反之,在雌者方面,如果逐渐获得一些习性使飞翔、游泳或行走等能力成为无用的话,也可能失去这等运动器官,而不会不安全。
然而,我们在这里所涉及的仅是性选择。性选择是以某些个体专在繁殖方面比同一性别和同一物种的其他个体占有优势为前提的。如上所述,当雌雄二者因生活习性不同而引起构造上的差异时,它们无疑是通过自然选择而发生变异,并靠遗传作用使该变异限于同一性别。除此之外,第一性器官以及养育或保护幼小动物的那些器官也都处于同样的影响之下;因为最善于繁殖和养育其后代的那些个体,在其他条件相等的情况下,将会留下最大量的后代,以继承它们的优越性;而不善于繁殖和养育其后代的那些个体就会留下少量的后代,以继承其弱小的能力。当雄者势必寻找雌者时,他就需要感觉的和运动的器官,但如果这些器官也是其他生活用途所需要的,它们将像在一般场合中那样,通过自然选择而得到发展。当雄者找到了雌者时,雄者有时绝对需要抱握的器官以便把雌者抓牢;因此华莱士博士告诉我说,某些蛾类雄者的跗节(即脚)如果破裂,它们就不能同雌者结合。许多海洋甲壳类的雄者一旦达到成年,它们的足和触角就会以一种异常的方式发生改变,以便抱握雌者;从这一点我们可以设想,这是因为这等动物被大海的浪涛冲向四方,因此它们为了繁殖其种类,它们就需要有这些器官,如果是这样的话,这些器官的发展就是正常选择即自然选择的结果。有些等级极低的动物也为了同样的目的而发生了变异,因此某些寄生虫的雄者,一旦充分成长,其躯体末端的下表面就变得像一把粗锉刀那样粗糙,它们借此把雌虫盘绕起来并持久地抱握住雌者。 (4)
倘若雌雄二者都遵循完全相同的生活习性,而且雄者的感觉器官或运动器官比雌者的更为高度发达,那么这些器官的完善化可能就是由于雄者为了寻求雌者所必不可少的;但这些器官在绝大多数情况下,仅仅是给某个雄性个体提供一种优势以胜过另一个雄性个体,因为,那些禀赋较差的雄者只要有充裕的时间也会成功地同雌者交配;再从雌者的构造来判断,它们在其他一切方面对于正常的生活习性也都适应得一样好。在这等场合中,既然雄者获得其现有构造并非由于要在生存斗争中更好地适于生存,而是由于获得了一种优势以胜过其他雄者,还由于把这种优势仅仅传给了其雄性后代,所以在这里性选择一定起了作用。正是这种区别的重要性,引导我把这种选择的形式命名为性选择。再者如果抱握器官对雄者的主要用途是在于当其他雄者到来之前或受到其他雄者的攻击时,防止雌者逃脱,那么这些器官将会通过性选择,也就是凭借某些个体所获得的胜于其竞争对手的优势而完善起来。然而在大多数这类场合中,要把自然选择的效果和性选择的效果区别开来是不可能的。关于雌雄二者的感觉、运动和抱握器官的种种差异细节可以连篇累牍地加以叙述。然而,由于这些构造并不比那些能适应正常生活用途的其他构造更为有趣,所以我将几乎完全略而不谈,而只在各个动物纲之下列举少数例子。
有许多其他构造和本能必定是通过性选择而发展起来的——诸如雄者用于同其竞争对手进行战斗并把它们赶走的进攻武器和防御手段——雄者的勇敢和好斗性——它们形形色色的装饰物——它们用来发生声乐或器乐的装置——以及它们那散发气味的腺体,后面这些构造中的多数仅仅是为引诱或刺激雌者服务的。显然,这些性状是性选择而不是自然选择的结果,因为要是没有禀赋较好的雄者在场,那些没有武装,没有装饰或是没有魅力的雄者也会同样成功地在生存斗争中留下众多的后代。我们可以推论,情况正是如此,因为雌者既没有武装又没有装饰,也仍然能够生存下来并繁殖其种类。我们刚刚涉及的这类次级性征,由于在许多方面都饶有兴趣,特别是由于这类性征有赖于任何一种性别的诸个体的意志,选择和竞争,所以将在以下几章详加讨论。当我们见到两个雄者为了占有雌者而战斗或是一些雄鸟在一群雌鸟面前展示它们华丽的羽衣并作出奇特滑稽的表演时,我们毫不怀疑它们这样做虽然是由于本能所引致,但显然懂得它们的所作所为是为了什么,而且是有意识地发挥其心理的和肉体的能力。
正如人们能从斗鸡场上选择优胜者来改进其斗鸡品种一样,在自然界里看来也是那些最强壮的、精力最充沛的或具有最佳武器的雄者占有优势,并导致自然界里的品种或种的改进。一种轻微程度的变异性可导致某种优势,无论这种变异性多么轻微,但在反复的生死争夺中,对于完成性选择的过程,这已经够用了;可以肯定第二性征是显著容易变异的。正如人类能按照自己的审美标准使雄性家禽产生美色,或更确切地说,人类能对原来由亲种所获得的美色加以改变,从而能使塞勃赖特短脚鸡(Sebright bantam)产生漂亮的羽衣和一种直立而独特的姿势——在自然状况下,看来雌鸟同样经过对那些魅力较强的雄鸟进行长期选择,曾使后者增添美色以及其他吸引雌者的属性。毫无疑问,这意味着雌者方面具有鉴别和审美的能力,乍一看,这似乎是极其不可能的;但根据以后所提出的事实,我希望能够阐明雌者确有这等能力。然而,当我们说到低于人类的动物具有美的感觉时,绝不可设想它可以同一个具有多种多样复杂联想的文明人的这种感觉相 比拟。把动物的审美力同最低等未开化人的审美力加以比较,是较为恰当的,这等未开化人赞美任何灿烂发光的或奇特的东西并用来装饰自己。
由于我们对若干之点还是无知无识,所以有关性选择作用的确切方式多少有点无法肯定。尽管如此,如果那些已经相信物种可变性的博物学者们读到以下章节,我想他们还会同意我的意见,即性选择对有机界的历史起了一种重要作用。可以肯定的是,在几乎所有的动物中,存在着雄者之间为了占有雌者的斗争。这个事实如此为大家所熟知,以致再举例说明就成为多余的了。假定雌者的心理能力足够用来选择对象,那么雌者就有从若干雄者当中选中一个对象的机会。在许多场合中,特定的环境条件使雄者之间的斗争特别剧烈。这样,英国的雄性候鸟一般都先于雌鸟抵达繁殖地,因此许多雄鸟早就做好了争夺每只雌鸟的准备。詹纳·韦尔(Jenner Weir)先生告诉我,那些捕鸟人断言夜莺(歌鸲nightingale)和莺(blackcap)永远都是如此,而关于后者,詹纳·韦尔先生自己就可以证实这种说法。
布赖顿的斯韦斯兰德(Swaysland)先生最近40年来有一种习惯,每当侯鸟第一次来到时他就去猎捕,而他从来没有发现任何物种的雌鸟先于雄鸟到达。在一个春天里,他打了39只雄黎氏鹡鸰(Budytes Raii)以后,还未见到一只雌鸟。丘尔德先生从解剖那些最先到达这个国家的鹬鸟中断定雄鸟先于雌鸟到达。美国绝大多数的候鸟也是如此。 (5) 从海洋溯游到英国一些河里的大多数雄鲑鱼比雌鱼先行到达,并做好了繁殖的准备。蛙类和蟾蜍类似乎也是如此。在整个昆虫这个大纲中,几乎总是雄虫先从蛹期羽化,因此一般在能见到任何雌虫之前的这段时间里到处都是雄虫。 (6) 雄者和雌者在到达期和成熟期上的这种差异,其原因是十分清楚的。那些每年最先迁徙到任何地方的雄者,在春季最先做好繁殖准备的雄者,或是最富于热情的雄者,都能留下最大量的后代,而这些后代大概都有遗传相似的本能和体质的倾向。必须记住,如果不同时干扰雌者的产仔时间,就不可能非常实质性地改变雌者性成熟的时间,而产仔时间一定是由每年的季节来决定的。总之,在几乎所有雌雄异体的动物中,雄者之间为了占有雌者要经常不断地反复进行斗争,这一点是无可怀疑的。
我们研究性选择所面临的难题,在于弄清楚战胜了其他同性对手的雄者、或者那些被证明对雌者最富有魅力的雄者怎样比被击败的、魅力较差的竞争对手留下了数量较多的后代以继承它们的优越性。除非确有上述结果发生,否则使某些雄者比其他雄者占有优势的那些性状就不会通过性选择而臻于完善和增强起来。假如雌雄二者以完全相等的数目存在,那些禀赋最差的雄者(多配性盛行的地方除外)也会最终找到雌性配偶,同那些禀赋最佳的雄者一样,留下同样多的后代,并且同样好地适应其一般的生活习性。根据各种事实和考察,我以前曾推断,关于第二性征十分发达的大多数动物,其雄者的数量大大超过了雌者,但这决不是永远如此。如果雄者同雌者是2∶1,或3∶2,甚至其比例多少更低些,那么整个情况就要简单了,因为那些武装得更好或更富有魅力的雄者将会留下最大量的后代。然而在尽可能考察了不同性别的数量比例之后,我不相信两种性别在通常情况下,其数量会悬殊太大。在大多数情况下,看来性选择的效用是通过下述方式来完成的。
让我们以任何一个物种为例,譬如说某一种鸟,把居住在某一地区的雌鸟分为相等的两群,其中一群包含的个体精力较充沛,营养状况较好,另一群包含的个体则精力较差,健康较弱。几乎没有任何疑问,前者在春季要先于其他雌鸟做好繁殖准备;这正是多年来对鸟类生活习性进行了细致观察的詹纳·韦尔先生的意见。同样无可怀疑的是,那些精力最充沛、营养状况最良好以及最早生殖的雌鸟,平均起来将会成功地养育数量最大的优良后代。 (7) 至于雄鸟,正如我们所见到的,一般先于雌鸟做好生殖准备;那些最健壮的雄鸟,以及有些物种里那些武装得最好的雄鸟,把弱者赶走之后,将会同那些精力较充沛、营养状况较良好的雌鸟进行交配,这因为它们都是最早开始生殖的。 (8) 这等精力充沛的配偶肯定会比那些发育迟缓的雌鸟养育数量较多的后代,假如雌雄二者的数量相等,这些发育迟缓的雌鸟势必要同那些打了败仗的力量较弱的雄鸟进行交配;对于在连续世代的过程中增加雄者的大小、体力和勇敢或是改进其武器,上述那种情况正是所需要的一切。
但在很多场合中,战胜了其竞争对手的雄者如果没有被雌者选中,还是不会占有后者。动物的求偶决不是如人们所想象的那么简单而短促的一桩事。雌者最容易受那些装饰较美的、或鸣唱最动听的、或表演最出色的雄者所挑逗,或者喜欢与之配对;但同时雌者们很可能挑选那些精力比较充沛而活跃的雄者,这一点已通过实际观察在一些例子里得到了证实, (9) 这样,那些最先开始生殖的精力比较充沛的雌者将会在许多雄者中进行选择,虽然雌者们也许不会总是选得最强壮的或武装最好的对象,但它们将会选得那些精力充沛的、武装良好的、并在其他方面最有魅力的对象。因此,这些早期交配的雌、雄双方,有如上面所阐明的,在养育后代方面,就会比其他配偶占有优势;显然这在诸代的漫长过程中足可使雄者不仅增加其体力和战斗力,同样地还可增添其各种各样的装饰物或其他魅力。
与此相反,雄者选择特定雌者的事例就比较罕见得多,在这样场合中,显然只有那些精力最充沛的并且战胜了其他对手的雄者才能最自由地选择雌者;几乎可以肯定,它们将会选择精力充沛的以及具有魅力的雌者。这等配偶在养育后代方面将占有优势,如果雄者在交配季节具有保护雌者的力量,如某些高等动物之所为,或能帮助雌者养育后代,则上述优势尤其明显。如果某一性别爱好和选择相反性别的某些个体,同样的原理也可应用;假定它们所选择的不仅是更有魅力而且也是更精力充沛的个体的话。
两种性别数量的比例
我曾说过,要是雄者的数量大大超过雌者,则性选择就是一桩简单的事情。因此,我对尽可能多的动物的两性比例进行了力所能及的调查,但材料是不充分的。我在这里所提出的只是调查结果的一个简短提要,而将有关细节留在附录中去讨论,以免干扰我的论述的过程。只有对家养动物,才能确定其出生时的性别比例数,但没有留下有关这个目的的任何记录。然而,我通过间接方法搜集了相当可观的统计数字,从这些数字可以看出我们大多数的家养动物在出生时其雌雄二者的数目接近相等。例如,竞赛马21年间的出生记录为25560匹,公马出生数与母马出生数的比例为99.7∶100。细躯猎狗(greyhounds)雌雄出生数不相等的程度比任何其他动物都大,在12年间出生的6878只小狗中,公狗与母狗的比例为110.1∶100。然而能否可靠地推论自然条件下和家养条件下的性别比例是一样的,在某种程度上尚属疑问。因为环境条件的轻微而未知的差异会影响性别的比例。因此,从人类来看,以女性出生率为100,则英国的男性出生率为104.5,在俄国为108.9,而利沃尼亚(Livonia)的犹太人则为120。但我将在本章的附录里再回头讨论这个男性出生数过量的奇妙问题。然而在好望角,若干年内出生的欧洲血统的男孩同女孩的比例数是(90~99)∶100。
对我们现今的目的来说,我们所关心的不仅是出生时期的性别比例数而且还有成熟时期的性别比例数,这就增加了另一个可疑因素;因为有一个十分确定的事实:就人类来说,男性在出生前、出生时以及幼儿时期最初几年内死亡的数目要比女性大得多。公羊羔的情况几乎肯定也是这样,有些其他动物的情况大概亦复如此。有些物种的雄性动物彼此争斗相杀,或者到处互相追逐直至变为衰弱不堪。它们在急切寻求雌者而四处奔走时,也必定常常面临种种危险。许多种类的雄鱼比雌鱼小得多,前者据信常常被后者或是别的鱼类所吞食。有些鸟类的雌鸟看来要比雄鸟早死,它们容易在巢里或照看雏鸟时被消灭掉。就昆虫来说,雌性幼虫常常大于雄性幼虫,从而就更可能遭到吞食。在某些场合中,成熟的雌者比较不爱动而且动作比雄者迟缓,从而不能有效地逃避危险。因此,关于自然状况下的动物,为了断定它们成熟时期的性别比例,我们必定只有依靠估计;除非性别数目的不相等非常悬殊,否则这种方法的可靠性是很小的。尽管如此,就我们所能作出的判断来说,我们可以从附录所列举的事实中作出这样的结论,即,少数哺乳类、多数鸟类、一些鱼类和昆虫类的雄性数量要比雌性数量大得多。
雌雄二者的逐年比例稍有变动,例如竞赛马,某年,每产100匹母马,相应产107.1匹种马,而另一年则为92.6匹,又如细躯猎狗,雄者的比例数从116.3变动到95.3。但是,如果在比英格兰更广阔的区域里搜集更庞大的数字来列表显示,则这等变动可能就会消失。像这样,简直不足以导致性选择在自然条件下发生作用。尽管如此,在某些少数野生动物的场合中,正如附录所表明的,性别比例似乎还在不同季节里,要不,在不同产地出现足够程度的变动以导致性选择发生作用。其所以如此,是因为可以观察到,那些能够战胜其竞争对手的,或对雌者最有魅力的雄者,在某些年代或某些产地所获得的任何优越性大概都会遗传给其后代,而不致在此后消失。在随后的季节里,当出现雌雄数目相等时,每个雄者如果都能得到一个雌者,那些产生较早的而且较为强壮的或更有魅力的雄者至少还会同较弱的或魅力较差的雄者一样有一个留下后代的良好机会。
多 配 性
多配性的实行也会导致由雌雄数目实际不相等所引起的相同结果,因为,如果每个雄者占有两个或更多的雌者,那么就会有许多雄者不能找到配偶;后者无疑将是那些较弱的或魅力较差的雄性个体。许多哺乳类或少数鸟类都是一雄多雌,即多配性的,但我未发现低等动物有这种习性的任何证据。这种动物的智力也许不足以导致它们集拢一群雌者并守住她们。看来多配性和第二性征的发达之间存在着某种关系,几乎是确定无疑了。这一点支持了这样的观点:即雄者的数量优势大概显著地有利于性选择的作用。尽管如此,许多严格单配性的动物,特别是鸟类,还显示了强烈显著的第二性征,而某些少数多配性的动物却没有这等性征。
我们先对哺乳类简略地浏览一下,然后再看看鸟类。大猩猩似乎是多配性的,而且雄者相当不同于雌者。有些狒狒也是如此,它们聚群而居,所含成年雌者为雄者的两倍。南美的卡拉亚吼猴(Mycetes caraya)在毛色、髭须和发音器官方面都呈现十分显著的性差异;而且一个雄者一般有二至三个雌者和它一起生活;白喉卷尾猴(Cebus capucinus) (10) 的雄者和雌者多少有些差别,好像也是多配性的。 (11) 关于大多数其他猴类的这方面情况还了解得很少,但有些物种是严格单配性的。反刍类显然是多配性的,它们所表现的性差异比差不多任何其他哺乳动物类群更加常见;这种情况特别适用于它们的武器,当然也适用于其他性征。大多数的鹿、牛和绵羊都是多配性的,大多数的羚羊也是如此,虽然有些是单配性的。安德鲁·史密斯爵士在谈到南非的羚羊时,指出在12只左右的一个羚羊群里,成熟的公羊很少超过一只。亚洲的高鼻羚羊(Antilope saiga)似乎是世界上最放纵的一雄多雌主义者,因为帕拉斯述说, (12) 这种公羚羊要把全部的竞争对手赶走并把100只左右的母羊和小羚羊集拢为一群;母羊无角而有较柔软的毛,但在其他方面与公羊没有太大差别。马尔维纳斯群岛上的野马和美国西部诸州的野马都是多配性的,但是,公马除了较大的体型、躯体的比例与母马有所不同之外,其他方面的差别很小。公野猪的獠牙和其他一些方面呈现出十分显著的性特征。在欧洲和印度,除了生殖季节之外,公野猪都过着独居生活;但是,正如在印度有很多机会对这种动物进行过观察的埃利奥特(W.Elliot)爵士所认为的那样,雄者在生殖季节同若干雌者相配。这种情况是否适用于欧洲的野猪还难以确定,但有某种证据支持这一点。成年的雄性印度象同野猪一样,在其一生中的大部时间里是独居的,但如坎贝尔(Campbell)博士所指出的,“当它同一些别的象在一起时,从一群雌象中发现的雄象很少多于一只”,较大的雄象把较小的和较弱的雄象赶走或弄死。雄象的粗长獠牙、庞大的体型,体力和耐力都同雌象有所不同;正因为这些方面的差异是如此之大,所以当捕到雄象时其价值要比雌象高出五分之一。 (13) 其他厚皮动物的雌雄二者差别很小或完全没有差别,如迄今所了解的那样,它们都不是多配性动物。我也没有听说过翼手目、贫齿目、食虫目和啮齿目中的任何物种是多配性的,除了啮齿目中的普通家鼠,据一些捕鼠人讲,雄鼠是同若干雌鼠生活在一起的。尽管如此,有些树懒(贫齿目)的雌雄二者在性状以及肩部毛斑的颜色上还有所不同。 (14) 而许多种类的蝙蝠(翼手目)呈现十分显著的性差异,不仅在于雄者具有散发气味的腺体和肚囊,而且在于它们的体色较浅。 (15) 在啮齿类这一大目中,就我所知道的,其雌雄二者很少有差别,即使有差别,也不过是毛的色泽稍有不同而已。
正如我听安德鲁·史密斯爵士说的,南非的雄狮有时同单独一只母狮一块生活,但通常是同较多的母狮在一起,有一回竟发现有五只母狮之多,因而雄狮是多配性的。就目前我所发现的来说,在所有陆栖食肉类中,雄狮是唯一多配性动物,而且只有它呈现了十分显著的性征,然而,如果我们把注意力转到海栖食肉类,正如我们以后将看到的那样,情况就大不相同了,因为海豹科的许多物种表现了异常大的性差异,而且它们显然是多配性的。例如佩隆(Péron)认为,南部海洋(Southern Ocean)的雄海豹经常占有若干只雌海豹,由福斯特(Forster)命名的雄海狮有二三十只雌海狮在其左右。在北部海洋,由斯特勒(Steller)命名的雄海狗,甚至伴随着更多的雌者。正如吉尔(Gill)博士的论述, (16) 有一个有趣事实,即,单配性的物种,“或是那些营小群生活的动物,其雌雄二者之间在体型大小上差别很小;那些社会性的物种或者更确切地说,那些雄者占有许多配偶的物种,其雄者的体型要比雌者大得多”。
在鸟类中,许多物种的雌雄二者之间的差别很大,它们肯定是单配性的。我们在大不列颠见到有些鸟类的性差异十分显著,例如,公野鸭只同单独一只母野鸭交配,乌鶫(blackbird) (17) 和红腹灰雀(bullfinch) (18) 据说都是终身配偶。华莱士先生告诉我说,南美的啁啾燕雀(Chatterers or Cotingidae)以及许多其他鸟类同样也是如此。在若干类群里我未曾发现这些物种究竟是多配性的还是单配性的。莱逊(Lesson)说,性差异非常显著的极乐鸟是多配性的,但华莱士怀疑他是否有充分证据。沙尔文先生告诉我说,他曾倾向于相信蜂鸟是多配性的。非洲产的黑羽长尾鸟(Widow-bird)以其尾羽著称,确实好像是一种多配性动物。 (19) 詹纳·韦尔先生和其他人士都曾向我保证说,一巢之内有三只欧椋鸟来往,似乎是常见之事;但这种情形到底是一雄多雌还是一雌多雄还不能确定。
鹑鸡类(Gallinaceae)所显示的性差异,其强烈显著的程度差不多同极乐鸟或蜂鸟一样,众所周知,其中许多物种都是多配性的;另外一些物种则是严格单配性的。多配性的孔雀或雉同单配性的珠鸡(guinea-fowl)或山鹑(partridge),其雌雄二者之间呈现了多么强烈的对照!关于松鸡族,也有许多相似的例子,如多配性的公松鸡(capercailzie)和公黑松鸡同母鸟差别很大,而单配性的红松鸡和羽脚雷鸟(ptarmigan)的雌雄二者之间的差别就很小。在走禽类(Cursores)中,除鸨类以外,只有少数物种呈现强烈显著的性差异,据说大鸨(Otis tarda)是多配性的。关于涉禽类(Grallatores)只有极少数物种有性差异,但流苏鹬(Machetes pugnax)则是一个明显的例外。蒙塔古(Montagu)相信这个物种是多配性的一种动物。由此看来,鸟类的多配性同强烈显著的性差异的发展之间有一种密切关系。我曾问过动物园的巴特利特(Bartlett)先生,他对鸟类的经验非常丰富,关于公角雉(tragopan,鹑鸡类的一种)是否多配性的问题,他的回答给我留下深刻的印象,他说:“我不知道,但从它鲜艳的羽色来看,可以这样认为。”
值得注意的是,这种只同单独一只母鸟成配偶的本能容易在家养条件下失去。野鸭是严格单配性的,而家鸭则是高度多配性的。福克斯牧师告诉我说,在他邻近的一口大池塘里有一群半驯化的野鸭,猎场看守人射死了其中大量的公野鸭,以致剩下来的公野鸭平均每只摊到七八只母野鸭,但居然也一窝窝地孵出了非常多的雏鸭来。珍珠鸡是严格单配性的,但福克斯先生发现当他将一只公珠鸡同两三只母珠鸡养在一块时,它们繁殖得最为成功。金丝雀在自然状况下是成双成对的,但英国的养鸟人把一只公雀和四五只母雀养在一块,成功地使它们进行了繁殖。我之所以注意到这些事例,是因为要提出野生的单配性物种可能容易地变成暂时的或永久的多配性物种。
关于爬行类和鱼类的习性,我们知道的太少了,以致我们无法说出它们的婚配方式。然而,据说刺鱼(Gasterosteus)是多配性的一种动物, (20) 雄者在生殖季节期间同雌者差别显著。
根据我们所能作出的判断,现对性选择导致第二性征发达所通过的途径作出如下总结。已经阐明,那些在竞争中战胜其雄性对手的最强壮、武装得最好的雄者,同那些在春季最早生殖的精力最充沛而且营养状况最良好的雌者配对以后,将会养育数量最多的精力充沛的后代。如果这等雌者所选中的雄者是魅力较强、同时又是精力充沛的雄者,那么它们将比那些发育迟缓的雌者养育数量较多的后代,因为后者势必要同一些精力较不充沛、魅力较差的雄者配对。如果精力较为充沛的雄者所选中的雌者是魅力较强、同时又是健康较好而且精力较为充沛的雌者,则其后果也将同上述一样;要是雄者保护雌者并且帮助雌者给后代供应食物,则其结果尤其如此。精力较为充沛的配偶在养育数量较多后代方面所获得的这种优势,显然已足够使性选择产生效果了。但是,雄者比雌者在数量上如果占巨大优势,其效果就更加显著,不管这种优势是否只是暂时性的和区域性的或持久性的,不管这种优势是否出现于降生时期或雌者大量夭折以后的时期,也不管这种优势是否间接地由于实行多配所引起的,都是一样。
雄者的变异一般大于雌者
在整个动物界中,除了很少例外,当雌雄二者在外部形态有所差别时,总是雄者的改变较大。因为,雌者一般都保持与同一物种的幼者和同一类群的其他成年成员密切相似的外形。产生这个现象的原因似乎在于几乎所有动物的雄者都比雌者具有较强的激情。因此,正是雄者彼此争斗,孜孜不倦地在雌者面前显示自己的魅力,而那些优胜者将把它们的优越性传给其雄性后代。为什么后代的雌雄二者没有这样都获得父方的性状,将在后面加以探讨。众所周知,所有哺乳动物的雄者都热切地追求雌者,鸟类也是如此。但许多公鸟追求母鸟并不那么积极,而只是在母鸟面前显示其羽衣,作出奇特的表演和纵声鸣唱。少数鱼类的雄者据观察似乎比雌者热切得多,短吻鳄类(alligators)的情况也确是这样,蛙类(Batrachians)的情况尤其明显。正如柯尔比先生所论述的, (21) 整个庞大的昆虫纲的“规律是雄者寻求雌者”。布莱克瓦尔和斯彭斯·巴特(C.Spence Bate)这两位优秀权威人士告诉我,蜘蛛类和甲壳类的雄者在其习性上比雌者更为活跃、更为见异思迁。当昆虫类和甲壳类的某一性别具有感觉器官或运动器官而另一性别却不具有的时候,或是像通常的情况那样,当这等器官在某一性别比在另一性别更加高度发达的时候,就我所能发现的来说,几乎必然是雄者具有这样器官,不然就是雄者的这样器官最为发达,这就阐明了雄者在两性求偶中是较为活跃的一方。 (22)
另一方面,除了极少例外,雌者在求偶中都比雄者缺乏热情。正如有名的亨特先生 (23) 早就观察到的,雌者一般都“需要求爱”,它是腼腆的,而且往往可以见到它在很长一段时间里竭力逃避雄者。每一位观察过动物习性的人都会回忆起一些这类例子。根据下面所列举的种种事实,以及根据完全是由性选择所产生的那些结果,可以阐明雌者虽然相对地比较被动,但一般也实行某种选择,并对一些雄者优先接受其中的一个。或者,它所接受的雄者并不是对它最富有魅力的,而是最少使它厌恶的;雄者的外貌有时使我们相信情况就是如此。雌者方面实行某种选择似乎同雄者热切求偶一样,几乎也是一个普遍的规律。
自然我们会追问,为什么在如此众多而且如此截然不同的各个纲中,雄性动物都变得比雌性动物更加热切求偶,所以是雄者寻求雌者,并在求偶中显示出更为积极的态度。假如雄者和雌者彼此相互寻求,这并不会带来什么好处,而只会招致一些精力的浪费;然而为什么雄者几乎总是寻求者?植物的胚珠在受精后还要接受一段时间的营养;因此花粉必然要被带到雌性器官——依靠昆虫或风力,要不就是依靠柱头的自发的运动,把花粉置于柱头之上;在藻类等植物中则依靠游动精子的运动能力。体制低的水生动物类永久着生于同一地点而且是雌雄异体,其雄性生殖要素(male element)总是始终不变地被运给雌者;我们不难看出这里面的原因,这是由于即使卵在受精前就被排出体外,并且不需要随后的营养和保护,但因为卵大于雄性生殖要素,而且产生的数量远比后者为少,所以卵的运送仍然要比雄性生殖要素的运送困难得多。因此,许多低等动物在这方面和植物是相似的。 (24) 固定于一个地点的水生动物的雄者就是被引导沿着上述那个途径放出它们的精子,自然,任何它们的后裔在等级上上升了并变为能动的以后,还会保持这同样的习性;为了避免精子在经过水中较长一段路程中受到损失的风险,它们就会尽可能地接近雌者。有些少数低等动物,仅是雌者固定不动,这等动物的雄者必定是异性的寻求者。但难以理解的是,为什么一些物种的雄者尽管其原始祖先是自由活动的,也总是获得向雌者接近的习性,而不是反过来雌者向雄者接近。但在所有场合中,为了雄者能有效地进行寻求,赋予它们以强烈的激情就成为必要的了;而更热切的求偶者比不太热切的求偶者留下数量较多的后代这一情况,自然会引致这等激情的获得。
雄者强烈的热切求偶,就这样间接地导致它们发展其次级性征比雌者更加常见得多。但是,雄者如果比雌者更容易发生变异,则其第二性征的发展大概就会得到很大帮助——我经过长期间对家养动物的研究得出了这个结论。阅历很广的冯·纳图西斯(von Nathusius)也强烈地持有同样观点。 (25) 从人类男女两性的比较中也可得出支持这个结论的有力证据。在诺瓦拉地方探险期间,曾对不同种族身体的好多部位进行了大量测量,几乎在每个例子里都发现男人比女人显示了更大的变异范围, (26) 但我将在后面一章再回头来论述这问题。伍德先生 (27) 曾仔细观察过男人肌肉的变异,他强调作出如下结论,“每具尸体上最大数量的肌肉变态都是在男人身上发现的”。在此之前,他曾谈过,“在所有102具尸体上,女人身上所发现的肌肉多余部分的变异只为男人的一半,这一情况同以前所描述的女人较多出现肌肉不足的情况形成了鲜明对照”。麦克利斯特博士也同样谈到 (28) 男人肌肉的变异“大概要比女人更常见”。人类身上所反常出现的某些肌肉,也是在男性身上比在女性身上更为发达,更加常见,虽然关于这个规律据说也有例外。伯特·怀尔德(Burt Wilder)博士 (29) 将152个有多余指的人的例子列成表,其中86个是男人,39个或少于半数是女人,剩下的27个则性别不明。然而,不应忽略女人要比男人更爱掩盖这类生理缺陷。此外,迈耶博士断言男人的耳朵在形态上比女人更易变异。 (30) 最后男人的体温比女人更容易变化。 (31)
雄性具有较大的一般变异性,其原因尚属不明,所知者只是第二性征特别容易变异,而且这种变异通常只限于雄者,我们即将看到,这个事实在某种程度上是可以理解的。很多事例说明,通过性选择和自然选择使雄性动物大不相同于雌者;但是,不依赖选择作用,雌雄二者由于体质上的差异也有按照多少不同的方式发生变异的倾向。雌者在形成卵的时候势必要消耗大量有机物质,而雄者则要把大量精力用于同竞争对手进行剧烈斗争,用于到处寻求雌者,用于呼叫声,用于散发气味的分泌物,等等;但这种消耗一般只集中于一个短时期。雄者在求爱季节的巨大活力似乎常常致使其色彩加强,而这同任何区别于雌者的显著差异并无关联。 (32) 在人类中,甚至在有机界等级上那样低的鳞翅类昆虫,其雄者的体温都高于雌者,此外还伴随着男人的脉搏较慢。 (33) 从总的方面来看,雌雄二者在物质上和精力上的消耗大概是接近相等的,但其消耗的方式和速率却大不相同。
由于刚才详细说明的那些原因,雌雄二者的体质几乎多少都有所不同,至少在生殖季节是这样;而且,虽然它们可能处于完全一样的条件下,却有按照不同方式发生变异的倾向。如果这等变异对任何一性都无用处,就不会被性选择或自然选择所积累和加强。尽管如此,如果激发的原因持久地起作用,这等变异还会成为永久性的;并且按照遗传上一种常见的形式,这等变异首先在哪一性别发生就只会传递给哪一性别。在这样场合中,雌雄二者将呈现永久性的、但不重要的性状差异。例如,艾伦(Allen)先生阐明,就居住在美国北部和南部的大批鸟类来看,得自南部的标本,其羽色比得自北部的标本较深,这大概是由两个地区的气温、光线等差异直接造成的结果。且说,有某些少数事例表明,同一物种的雌雄二者所曾受到的影响好像有所不同;红翼椋鸟(Agelaeus phaeniceus)的雄者在南部其羽色大大加深了;相反,关于北美红雀(Cardinalis virginianus)受到了这样影响的却是雌者;关于欧洲山鹬(Quiscalus major),雌者的色彩极易变异,而雄者的色彩则几乎保持一致。 (34)
许多纲的动物也出现了少数例外:获得十分显著第二性征的——诸如鲜艳的色彩、较大的体型、体力或好斗性,是雌者,而不是雄者。关于鸟类,雌鸟和雄鸟所特有的正常性状有时会完全倒置过来;是雌鸟在求偶时变得更热切,而雄鸟则比较被动,但我们可以从求偶的结果推断,雄鸟仍明显地选择魅力较强的雌鸟。某些雌鸟就是这样获得了更鲜艳的色彩或别的装饰,也获得了比雄鸟更大的力量和好斗性;而这些性状只传给其雌性后代。
可以这样认为:在某些场合中曾经进行了一种双重的选择过程,这就是雄者选择魅力较强的雌者,而雌者也选择魅力较强的雄者。然而,这种过程虽然会导致雌雄二者都发生改变,却不会使某一性别同另一性别产生差异,除非二者的审美力确实不一样;但这是极不可能的一种假设,除人类以外,对任何动物来说都没有考虑的价值。不管怎样,还有许多动物的雌雄二者彼此类似,具有同样的装饰,根据类推,我们可以把这种情形归因于性选择的力量。在这样场合中,可以提出一个似乎比较说得通的假设,即有一种双重的或交互的性选择过程存在;那些精力较充沛和较早熟的雌者选择魅力较强和精力较充沛的雄者,而后者除了那些魅力较强的雌者之外,拒绝接受其他任何对象。但根据我们所了解的动物习性来看,这个观点几乎是不可能的,因为雄者一般都热切于同任何一个雌者交配。对于雌雄二者所共有的装饰,比较可能的解释是,这种装饰是由某一性别、一般是由雄性获得的,然后传递给雌雄二者的后代。如果任何一个物种的雄者在一个相当长的时期内确实远远超过雌者的数量,然后在另一个相当长的时期内,由于条件的改变,却出现了相反的情况,那么一种双重的但不是同时发生的性选择过程就会易于进行,从而使雌雄二者大不相同。
我们以后会知道有许多动物,其雌雄二者都没有鲜艳的色彩,也不具备特别的装饰,而通过性选择双方或只有其中一方的成员很可能获得像白色或黑色那样的简单色彩。上述这些动物没有鲜艳的色彩或其他装饰可能是由于正常的变异从未发生,也可能是因为它们本身就喜欢全白或全黑的颜色。暗淡不鲜艳的色彩常常是为了保护自己,通过自然选择而发展起来的;通过性选择所获得的鲜明色彩,有时似乎会因此招来危险而受到抑制。但在其他场合中,雄者在悠久的年代中可能为了占有雌者而互相斗争,但是,除非那些成功较大的雄者比成功较小的雄者留下数量更多的后代以遗传它们的优越性,否则就不会产生任何效果。如上所述,这要取决于许多复杂的偶然性。
性选择的作用方式不像自然选择那样严峻。自然选择所产生的效果,不论动物的年龄,将会使成功的个体生存,使不成功的个体死亡。在雄者进行竞争的互相冲突中所造成的死亡确不少见。但是,较少成功的雄者一般仅是得不到雌者,或是在生殖季节的后期得到一个发育迟缓而精力不充沛的雌者,再不然,如果它们是多配性的,就只能得到为数较少的雌者;因此它们留下来的后代数量较少而且精力不充沛,甚至绝了后代。关于通过正常选择、即自然选择所得到的那些构造,只要生活条件保持不变,在大多数情况下,其与某些特殊用途有关的有利变异量都有一个限度;但是关于使某一雄者在斗争中或对雌者献媚中胜过另一雄者的那些构造,其有利变异量就没有明确的限度;所以只要这种适宜的变异性一旦发生,性选择的工作就不会停止。这个情况也许可以部分地说明第二性征何以如此频繁地发生变异,而且其变异量又何以如此之大。尽管如此,如果这些性状由于过分消耗动物的生命力,或者由于把它们暴露在任何巨大危险之下而是高度有害的话,那么自然选择还会决定优胜的雄者不致获得这等性状。然而,某些构造——例如某些公鹿的角——还是发达到令人吃惊的极端;在某些场合中,就一般生活条件来说,趋向极端对于雄者一定略有危害。根据这一事实我们认识到,由于在战斗或求偶时战胜了其对手因而留下了大量后代所带给那些雄者的利益,到头来要比由于对生活条件更能完善适应所带来的利益为大。我们还将进一步看到,雄者取媚于雌者的力量有时要比在战斗中战胜其他雄者的力量更为重要,但这决不是以前所能预料到的。
遗 传 规 律
为了理解性选择如何作用于不同纲的许多动物,以及如何在世世代代的过程中产生出一种显著的结果,就必须记住那些已被发现的遗传规律。“遗传”这个术语包含有两个不同的要素——性状的传递和性状的发育;但由于二者一般是相伴进行的,因此它们的区别就往往被忽略了。我们可从那些在生命早期进行传递而只在成年期或老年期才发育完成的性状上见到这二者的区别。从第二性征上可以更清楚地看到这种区别,因为这些性状是通过雌雄双方传递下去的,但只在其中一方发育。当两个具有强烈显著性征的物种进行杂交时,这些性征存在于雌雄双方的情况就显而易见了,这是因为雄性亲本或雌性亲本都会把各自特有的那些性征传递给任何一性的杂种后代。当雌者年老或得病时,偶尔也会发育出雄者所特有的那些性征,例如普通母鸡呈现公鸡的飘垂尾羽、颈部纤毛、鸡冠、脚距、鸣叫,甚至还会呈现公鸡的好斗性,在这里,上述同样的事实也是显而易见的。反过来,关于去势的公鸡,也多少可以清楚看到同样的现象。此外,同年老或得病无关,雄者的有些性征偶尔也会传递给雌者,如在鸡的某些品种中,健康的小母鸡会经常地呈现公鸡的距。但是,事实上这些性征只不过是在母鸡身上得到发育而已;因为在每个品种中,脚距的各个细微构造都是通过雌者传递给其雄性后代的。以后还要举出许多事例来说明雌者多少完整地显示出雄者所特有的性征,这些性征必然是最先在雄者身上发育的,然后再传递给雌者。至于在雌者身上最先发育的那些性征被传递给雄者的相反事例不甚常见;因此举出一个显著的事例,将是有益的。关于蜜蜂,只有雌蜂才用采集花粉的器官为幼虫采集花粉,但在大多数物种中,这种器官在雄蜂身上也部分地得到发育,但这对它却毫无用处,在雄熊蜂(Bombus)身上这种器官则得到了完全的发育。 (35) 虽然我们有某种理由去猜想雄性哺乳动物在原始时期同雌性哺乳动物一样也给幼仔喂奶,但由于其他任何膜翅目昆虫、甚至同蜜蜂有密切亲缘关系的小胡蜂(wasp)都不具有花粉采集器,所以我们没有根据去假定雄蜂在原始时期也曾同雌蜂一样地采集花粉。最后,在返祖的所有场合中,性状的传递是经过两代、三代或更多的世代,然后在某种未知的有利条件下发育起来。靠泛生说(pangenesis)的帮助,我们将会把性状的传递和性状的发育二者之间的这种重要区别牢牢记住。按照这个假说,身体每个单位或每个细胞都会放出芽球(gemmules)、即未发育的微粒,它们被传递给雌雄二者的后代,并且依靠自体分裂而成倍地增加。它们在生命的早期或在连续的世代内保持不发育状态;它们是否会发育成像其所来自的那样的单位或细胞,则取决于在生长的正常次序中同先前发育的其他单位或细胞的亲和力和结合。
在生命相应时期的遗传性
这种倾向已经完全得到证实。一只幼年动物身上出现的一个新性状,不管它是保持终生或转瞬即逝,一般都将在后代的同一年龄中重现并保持同样的时间。另一方面,如果一个新性状出现于成年、甚至老年,它就倾向于在同样老的年龄中重现。一旦发生偏离这个规律的情况时,被传递的性状的出现早于相应年龄要比晚于相应年龄更加常见。由于我已在另一著作中充分讨论了这个问题, (36) 因此,我只准备在这里举出两三个事例以唤起读者对这两个问题的回忆。在鸡的若干品种里,全身披着绒毛的雏鸡,最初长出真羽毛的小鸡以及成年鸡,彼此都有重大差异,就像它们同其共同亲类型原鸡(Gallus bankiva)之间的差异一样。每个品种都把这些性状在其生命相应时期忠实地传递给后代。例如汉堡亮斑鸡(spangled Hamburgs)的雏鸡当初生绒毛时,头部与臀部只有少数黑暗点,但不像许多别的品种那样,呈现纵条纹;它们长出的第一批真羽毛,“具有美丽的线纹”,这就是说,每根羽毛都有无数的横条斑;但它们的第二批羽毛就全部生有亮晶晶的斑点,即每支羽端都具有一个黑色的圆点。 (37) 因此这个品种的变异是在三个不同生命时期中发生和传递的。鸽类提供了一个更为显著的例子,因为作为原始祖先的亲种除了在成年期胸部虹色变得较深之外,并不随着年龄的增长发生羽毛变化;但仍然还有些品种不换两三次甚至四次羽毛就不会获得它们所特有的色彩;羽毛的这些变异都是有规则地被传递下去的。
在一年相应季节出现的遗传性
关于生活在自然状况下的动物,有无数事例说明性状是在不同季节中定期出现的。我们从雄鹿的角可以看到这一点,从北极动物的毛在冬季变厚变白的现象也可以看到这一点。许多鸟类仅在生殖季节获得鲜艳的颜色和其他的装饰。帕拉斯 (38) 说,西伯利亚家养的牛和马到冬季颜色变淡;我本人也曾观察过并听说过关于颜色的类似强烈显著变化,那就是英国有些驮马(ponies)从褐黄色或红褐色变成全白色。虽然我不了解在不同季节皮毛颜色发生变化的这种倾向是否会传递下去,但可能就是这样,因为各种浓淡的毛色都可以被马强烈地遗传下去。这种受季节限制的遗传形式,并不比受年龄和性别限制的遗传形式更加显著。
限于性别的遗传性
性状相等地传递给雌雄二者是遗传的最普通形式,至少那些不呈现强烈显著性差异的动物是这样,许多这类动物确实采取了这种遗传形式。但最先在某一性别身上出现的那些性状,多少都是一般地被传递给那一性别。我在《动物和植物在家养下的变异》一书中已经提出过有关这个问题的充分证据,然而这里不妨再举少数几个例子来说明。有些绵羊和山羊的品种,其公羊的角在形态上同母羊的角大有差异;在家养下所获得的这些差异有规则地传递给相同的性别。猫通常只有母的是玳瑁毛的,而公猫相应的颜色则是暗红色的。在大多数家鸡品种中,每一性别所特有的性状只传递给相同的性别。性状传递的这种形式是如此普遍,以致一旦出现某些品种的变异相等地传递给雌雄双方的情况时,就成了一种反常的现象。还有某些家鸡的亚品种,其公鸡几乎无法区别,而母鸡的颜色则明显不同。原种岩鸽的雌雄二者在外部性状上并无差别;尽管如此,某些家养品种的公鸽羽色还和母鸽羽色有所不同。 (39) 英国信鸽的垂肉和突胸鸽(Pouter)的嗉囊在雄者比在雌者更加高度发育;这些性状虽是通过人类长期选择而被获得的,但雌雄二者之间的轻微差异则完全是由于发生作用的遗传形式,因为这些轻微差异的发生与其说是出于育种者的愿望,莫如说是违背了育种者的愿望。
大多数我们的家养族都是通过轻微变异的积累而形成的,由于有些后续的变异步骤只传递给一性,还有些后续的变异步骤则传递给雌雄两性,因此在同一物种的不同品种中,从雌雄二者极不相似到完全相似之间,可以发现所有的级进。我们已经举出有关家鸡和家鸽诸品种的事例,在自然界里也常常见到类似的情况。关于家养动物,某一性别可能失去其特有的性状而多少变得同另一性别相似,例如,有些家养品种的公鸡失去雄性的尾羽和颈部纤毛,至于自然界的动物是否也有这种情况我不敢乱说。另一方面,在家养下雌雄二者之间的差异可能加大,如母的美利奴绵羊已失去了它们的角。此外,某一性别所特有的性状还会在另一性别突然出现;如有些家鸡亚品种,其年幼的母鸡有距;又如某些波兰鸡亚品种;有理由可信其母鸡原来获得了冠羽,随后又将它传递给了公鸡。根据泛生论假说,所有这些情况都是可以理解的;因为,所有性状都取决于下述情形,即,某些部分的芽球虽存在于雌雄二者,但通过家养的影响,它们在这一性别或那一性别中变为潜伏的或发育的。
为了方便起见,把下面的一个难题安排在以后的一章来讨论较为合适,这就是,最初在雌雄双方都发育的一种性状,是否会通过选择限于只在某一性别中发育。举例来说,如果某个育种者观察到他的一些鸽子(它们的性状通常都是以同等的程度传递给雌雄双方)变为蓝灰色,那么他能否通过长期持续的选择形成一个只是公鸽具有这种颜色而母鸡保持不变的品种呢?我在此只能说,做到这一点虽然并非不可能,但将是极端困难的。因为以蓝灰色公鸽进行繁育的自然结果将使整个类族的雌雄双方都变成这种颜色。然而,如果人们所希望的颜色变异出现了,而且这种变异一开始就限于在雄者一方发育,那么要形成一个雌雄颜色不同的品种,就一点也没有困难,例如一个比利时品种确实就是这样形成的,只是这个品种的公鸽才具有黑色条纹。同样,如果有一只母鸽发生了任何变异,而且这种变异从一开始就限于在母鸽身上发育,那么要培育出一个只有母鸽才有这种特性的品种也很容易,但是如果这种变异从一开始就没有上述这样的性别限制,那么这一过程将极难实现,甚至不可能实现。 (40)
性状的发育时期同该性状向某一性或向雌雄两性传递的关系
为什么某些性状会遗传给雌雄二者,而另外一些性状只遗传给某一性别,即遗传给最初出现这种性状的那一性别,在大多数场合中其原因还是完全未知的。我们甚至无法猜想,为什么在鸽子的某些亚品种中其黑色条纹虽然通过母鸽传递下去,却只在公鸽身上发育,而另一方面其他每个性状又是相等地传递给雌雄双方。另外,为什么猫的龟甲色除了很少例外只在雌者身上发育。就人类而言,完全同样的性状如缺指,多指,色盲,等等,在某一家族中只遗传给男性,而在另一家族中则只遗传给女性,虽然在这两种情况下,通过相反性别或通过相同性别传递都是一样的。 (41) 我们虽然这样无知,但知道有两条规律似乎往往是适用的——即任何一性在其生命晚期最初出现的变异就有只在这相同一性别进行发育的倾向;另一方面,任何一性在其生命早期最初出现的变异就有在雌雄双方都进行发育的倾向。然而,我绝不是假定这就是唯一的决定原因。鉴于我没有在任何地方讨论过这问题,鉴于这个问题对于性选择的重要意义,我必须在此对一些冗长而有些复杂的细节加以论述。
事物的本身存在这种可能性,即在幼年出现的任何性状都有相等地遗传给雌雄双方的倾向,因为雌雄二者在获得生殖力之前,它们的体质并没有多大差异。另一方面,在获得生殖力之后,而且雌雄二者的体质已发生了差异,那么,从某一性别的各个变异着的部分释放出来的芽球(如果我可以再次使用泛生说的术语的话),将和同一性别的组织相结合并由此发育起来,这种和同一性别的固有亲和力比和相反性别的亲和力远远更加可能发生。
最初我根据以下事实来推论有这种关系存在,即成年雄者同成年雌者无论何时并且无论以何种方式有所差异,则雄者也按照同样方式而有别于雌雄二者的幼仔。这个事实的普遍性十分显著:它适用于差不多所有的哺乳类,鸟类、两栖类和鱼类,同样也适用于许多甲壳类,蜘蛛类以及少数昆虫类,如某些直翅目昆虫和蜻蛉科(libellulae)昆虫。在所有这样场合中,凡是雄者通过变异的积累而获得其特有性状者其变异一定是在生命的稍晚时期发生的;否则年幼的雄者也会具有同样的特性;而且同上述规律相符合,这等变异只向成年雄者传递,也只在成年雄者发育。另一方面,要是成年雄者同雌雄双方的幼仔密切类似(除了很少例外,雌雄双方的幼仔都彼此相像),则雄者一般也同成年雌者类似;在大多数这种场合中,凡是老者和幼者通过变异而获得其现有性状者,按照上述规律,这等变异大概是在幼年时期发生的。但这里还有可疑的余地,因为有时性状传递给后代时的年龄要比父母最初出现该性状时的年龄为早,因而父母在成年时发生变异,而在幼年时将它们这等性状传递给后代。还有许多动物,其雌雄二者彼此密切类似,但二者都同各自的幼仔有差别,这时,成年动物的那些性状一定是在生命晚期获得的;尽管如此,这些性状还是传递给雌雄双方,这显然同上述规律相矛盾。我们千万不要忽视出现下述情况的可能性或者甚至盖然性,即在相同的生活条件下发生的相同性质的连续变异会在生命相当晚的时期同时出现于雌雄双方;在这样的场合中,这些变异将在相应的晚年传给雌雄双方;这样,就与上述规律并无真正的矛盾,即,凡是在生命晚期出现的变异都专门传递给最先发生该变异的那一性别。这一规律的适用范围似乎比第二个规律更为普遍,后面这个规律表明任何一性在生命早期出现的诸变异都有传递给雌雄双方的倾向。在整个动物界中究竟有多少事例可适用这两个定理,仅仅对此作个估计也显然是不可能的,因此,我认为只能对一些显著的或有决定意义的事例加以研究,以便得出可以依据的结果。
鹿科提供了进行研究的极好例子。除了一个物种外,在所有物种中,只有公鹿生角,虽然这个性状肯定是通过母鹿传递下去的,而且能在母鹿头上出现反常的发育。另一方面,母驯鹿(reindeer) (42) 也生角;因此按照上述规律,这个物种的角应该远在雌雄二者成熟并呈现体质重大差异之前的生命早期出现的。其他所有物种的角则应该在生命较晚时期出现,从而导致它们的发育只限于一性,这正是整个鹿科祖先最初生角的那一性别。现在属于鹿科不同组(section)的、并且居住在不同地区的七个物种只有公鹿生角,我发现角的最初出现时期不同,公獐(roebuck)出生后九个月生角,其他六个体型较大的物种的公鹿,最初生角时期在出生后十个月、十二个月甚至更长的时间。 (43) 但驯鹿的情况就大不一样,因为奈尔森博士热心地在瑞典的拉普兰为我作了专门调查,他说,出生后四五个星期以内的幼鹿就生角,而且同时出现于雌雄双方。因此我们这里看到鹿科的一个物种的一种构造在生命最早时期的发育,而且只有这一个物种的雌雄二者都生角。
有几个种类的羚羊,只有公羊生角,而更多种类的羚羊则雌雄二者都生角。关于角的发育时期,布赖茨先生告诉我说,动物园里的一头幼南非条纹羚羊(Ant.strepsiceros)有一次生角,但只限于雄者;还有一个亲缘关系密切接近的物种——南非大羚羊(Ant.oreas),它的小羊无论雌雄都生角。这一情况同上述规律完全符合,即,南非条纹羚羊的小公羊虽然已10个月,但从它最终的角的大小看来,那时的角显得很小;另一方面,南非大羚羊的小公羊虽然只有3个月,它的角都比前者的角大得多。在叉角羚羊(pronghorned antelope) (44) 中还有一个值得注意的事实,即只有少数母羊,约1/5有角。她们的角有时虽也有4英寸长,但都处于残迹(退化)状态;因此,要是只就公羊才生角这一点来考虑,则这个物种是处于中间状态的,而且它们的角大约要在出生后5~6个月才长出来。因此,同我们还不太清楚的其他羚羊类的角的发育情况相比,并且根据我们已经清楚的关于鹿、牛等动物的角的发育情况,就可看出叉角羚羊的角是在生命的居中时期出现的——换句话说,既不像牛和绵羊那样早,也不像大型的鹿和羚羊类那样晚。绵羊、山羊和牛的角,在雌雄双方其大小虽不完全一样,但都发育良好,在它们出生时,或刚出生后不久,就可以摸到甚至看到。 (45) 然而,上述规律对于绵羊的某些品种,例如美利奴绵羊似乎就不适用了,在这个品种中只有公羊生角;因为我在调查中, (46) 未能发现这个品种的角的发育时期晚于雌雄二者都生角的普通绵羊。但是,就家养绵羊来说,有角或无角并不是一种十分固定的性状,因为美利奴绵羊有一定比例的母羊也生短角,而有些公羊却不生角,在大多数品种中偶尔也会产出无角母羊。
马歇尔博士最近对鸟类头上常见的突起作了专门研究, (47) 得出以下结论——凡是头上突起只限于公鸟的那些物种,该性状是在生命晚期发育的;凡是头上突起为雌雄二者所共有的那些物种,该性状是在生命很早时期发育的。这一结论肯定同我的上述两项遗传定律显著符合。
在美丽的雉科大多数物种中,公雉和母雉显著不同,它们是在生命相当晚的时期才获得其装饰物的。然而,蓝马鸡(Crossoptilon auritum)提供了一个明显的例外,因其雌雄二者都有尾羽、耳簇毛,而且头部都有深红的天鹅绒般的软毛;我发现这些性状都是在生命的很早时期出现的,与上述遗传规律相符。但其成年公鸡可根据脚距同成年母鸡区别开来,而且同我们的规律相符的是,这些距要到出生六个月后才会开始发育,然而巴特利特(Bartlett)先生向我确言,即使在这个龄期也几乎无法把雌雄二者区别开来。 (48) 公孔雀和母孔雀除了共有的华丽冠毛之外,几乎每一部分的羽毛都明显不同;而冠毛是在生命的很早时期发育的,它的发育远在公孔雀所专有的其他装饰物的发育之前。野鸭也有类似情况,母鸭翅上美丽的绿色灿点虽然比公鸭小些,模糊些,但由于这个性状是二者所共有的,因而它的发育是在生命的早期,另一方面,公鸭的卷曲尾羽和其他装饰物则要在较晚的时期才发育。 (49) 在马鸡那样的雌雄二者非常相像和孔雀那样的雌雄二者极不相像的两类极端事例之间,还可以举出许多中间性的事例,在这些事例中性状的发育顺序是遵循上述那两条规律的。
由于大多数昆虫都是在成熟条件下才从蛹羽化出来,因此发育周期是否能决定性状向性别一方或是双方进行传递,尚无法确定。例如蝴蝶有两个物种,其中一个物种的雌雄颜色不同,另一个物种的雌雄颜色则一样,但我们不知道这两个物种的有色鳞粉是否在同一个相应蛹期发育的。我们也不知道全部有色鳞粉是否同时在同一个蝴蝶种的翅上发育的,在这个蝴蝶种中,某些色斑只为某一性别所具有,还有一些色斑则为两性所共有。发育时期的这种差异最初看来好像很不可能,其实并非如此;因为直翅目昆虫达到成熟状态并不是单单由于一次变态,而是由于连续的蜕皮,有些物种的幼龄雄虫最初同雌虫相像,而只是在稍晚的一次蜕皮中才获得其明显的雄性性状。某些雄性甲壳类在连续蜕皮的过程中也出现了完全相似的情况。
到现在为止,我们只考察了同发育时期有关的性状传递,而且所涉及的只是处于自然状况下的物种的性状传递;现在我们要转来谈谈家养动物,而首先要提到的是畸形和疾病的问题。多余指的出现和某些指骨的缺如,必定在很早的胚胎期就被决定了——大量出血的倾向至少是先天性的,色盲大概也是如此——但这些特性以及其他相似特性的传递往往只限于一性;因此早期发育的性状倾向于传递给雌雄双方的这条规律在这里就完全失效了。但是,如上所述,这一条规律似乎不如相反的那一条规律普遍有效,后者表明,在某一性别的生命晚期出现的性状,专门传递给同一性别。上述不正常的特性远在生殖机能活动之前就已为某一性别所具有,根据这个事实,我们可以推论雌雄二者一定早在极幼小的时期就已经有了某种差异。关于受到性别限制的疾病,我们对其发生的时期了解甚少,以致难以作出可靠的结论。然而痛风病(gout)似乎受我们的规律的支配,因为这种病一般是在成年时由酗酒造成的,并且由父亲传给了子女,而在儿子方面远比在女儿方面表现得显著。
关于绵羊、山羊和牛的各个家养品种,雄者在角、额、鬃毛、颈部垂肉、尾和肩上隆肉的形状及其发育情况方面都和雌者有所不同;按照我们的规律,这些特性不到生命相当晚的时期是不会充分发育的。狗类没有雌雄差异,但某些品种是例外,特别是苏格兰猎鹿狗,其雄者比雌者大得多,重得多;而且我们将在后面一章看到,雄者的体型会持续增长到生命异常晚的时期,按照上述规律,这个情况将说明这种体型的增长只传给雄性后代。另一方面,只限于母猫才有的龟甲色在其出生时就十分明显,这个情况是违背上述规律的。有一个鸽子的品种,只是公鸽具有黑色条纹,这些条纹甚至在雏鸽身上就可以察觉出来;但这些条纹随着每次换毛而日益明显,从而这个情况既是部分地违背了又是部分地证实了上述规律。英国信鸽和突胸鸽的垂肉和嗉囊都是在生命的相当晚期才充分发育的,同上述规律相符合,这等性状充分完善地只传递给公鸽。下面的例子也许属于前面提到的那一类,即雌雄二者在生命的相当晚期以同样的方式发生变异,从而在相应的晚期将其新性状传递给后代的雌雄双方;果真如此,这些情况同我们的规律并不矛盾——根据诺伊迈斯特(Neumeister)的叙述, (50) 有这样一些鸽子的亚品种,其雌雄二者都在两三次换毛期间改变毛色[杏包翻头鸽(Almond Tumbler)也是这样],这等变化虽发生于生命的相当晚期,却为雌雄双方所共有。有一个金丝雀的变种,名为“伦敦获奖者”(London Prize),提供了一个很近似的例子。
关于家鸡的品种,其种种性状是由一性遗传下去,还是由两性都遗传下去,似乎一般是由这些性状的发育时期来决定的。这样,在所有这许多品种中,如果成年公鸡在毛色上同母鸡有重大差异,而且同野生亲种也有重大差异,那么成年公鸡也会同小公鸡有差异,所以新获得的性状一定是在生命的相当晚期出现的。另一方面,在公鸡和母鸡彼此类似的大多数品种中,小鸡的毛色则同其双亲的差不多一样,因此,它们的毛色最初出现于生命的早期。我们在全黑和全白的品种中可以看到这个事实的例证,这些品种的雌雄小鸡和老鸡都彼此相似;我们也不能主张全黑的或全白的羽毛有什么特殊之处可以导致这种性状传递给雌雄双方;因为有许多自然界的物种,只是公鸡的羽色是黑的或白的,而母鸡则是别种颜色的。有一个叫做“杜鹃鸡”的亚品种,它的羽毛具有黑色横条纹,其雌雄双方和小鸡的毛色几乎都一样。塞勃赖特矮脚鸡(Sebright bantam)的雌雄二者都具有花边羽衣,小鸡的翅羽花边虽不完善,但很明显。然而亮斑汉堡鸡提供了一个局部例外的情况;因为其雌雄二者虽不完全相像,但比起原始亲种的雌雄二者,彼此更加相像得多;然而它们的特有羽衣是在生命晚期获得的,因为小鸡也具有明显不同的彩色条纹。至于颜色以外的其他性状,不论野生亲种还是大多数家养品种,只有公鸡才有发达的肉冠;但小西班牙鸡在很早的龄期其肉冠就十分发育,同公鸡肉冠的早期发育相一致,成年母鸡的肉冠也异常之大。在猎鸟的品种中,好斗性的发育早得令人吃惊,在这方面可以举出一些奇妙的证据;这个性状是传递给雌雄双方的,所以由于母鸟的极端好斗,现在一般都分栏展出。关于波兰鸡品种,头部支持鸡冠的骨质突起甚至在小鸡孵化之前就已部分发育了,而鸡冠本身也很快开始生长起来,虽然起初它还是柔弱无力的; (51) 这品种的成年公鸡和成年母鸡都有一个大型骨质突起和一个巨大鸡冠作为特征。
最后,根据我们现在所见到的许多自然界物种和家养族的性状发育时期和性状传递方式之间的关系——例如昭然若揭的下述事实:雌雄二者都生角的驯鹿,其鹿角是在早期生长的,而与此相对的是,只有雄者才有角的其他物种,其鹿角则是在晚得多的时期生长的——我们可以得出如下结论,第一,其性状专门遗传给雌雄任何一方的原因是由于这些性状是在生命晚期发育的,虽然这不是唯一的原因。第二,其性状遗传给雌雄双方的原因是由于这些性状是在雌雄双方的体质还没有多大差别的生命早期发育的,虽然这明显是一个不甚有力的原因。然而,看来雌雄二者之间甚至在很早的胚胎期就一定存在着某种差异,因为早期发育的性状只为某一性别所具有者,并不罕见。
提要和结论
根据上述对遗传法则的讨论,我们认识到双亲的性状常常是,甚至普遍是倾向于在双亲最初发生这等性状的同一龄期、同一季节、同一性别的后代中发育的。但是,这些规律由于不明的原因还远远不是固定不变的。因此,当一个物种发生变异时,那些连续的变化可能随时以不同的方式被传递下去;有的只传递给性别一方,有的则传递给雌雄双方;有的只在一定的龄期传递给后代,有的则不问龄期而传递给后代。不仅遗传法则是极端复杂的,而且诱发和控制变异性的诸原因也是极端复杂的。这样被诱发起来的变异由性选择保存下来并积累起来,而性选择本身又是极端复杂的事情,性选择实际上取决于雄者的求爱热情、勇气和竞争,还取决于雌者的识别力、审美力和意愿。性选择还受到有助于物种普遍福利的自然选择的支配,因此,性选择对任何一性的个体的影响方式或对雌雄双方的个体的影响方式必然是高度复杂的。
当变异发生在某一性别的生命晚期并在同一龄期向同一性别传递时,另一性别及其幼仔都保持不变。当变异发生在生命晚期、但在同一龄期向雌雄双方传递时,则只有幼仔保持不变。然而变异可能在某一性别或在雌雄两性的生命任何时期发生,并在一切龄期向雌雄双方传递,于是这个物种的一切个体就会同样地发生改变。在以下几章将可看到所有这些情况经常在自然界里发生。
性选择在未达生殖年龄之前,决不会对任何动物发生作用。由于雄者求偶的巨大热情,性选择一般是对雄者发生作用,而不对雌者发生作用。这样,雄者就会获得同其竞争对手战斗的武器,获得用以发现雌者并牢牢抓住她的器官,也获得用以刺激雌者或向其献媚的器官。要是雄者在这些方面都同雌者有所差异,那么成年雄者同幼年雄者也会多少有所不同,正如我们已经见到的,这是一个极普遍的法则;根据这个事实我们可以断言,使成年雄者发生改变的那些连续变异,一般不会远在生殖年龄之前出现。每当在生命早期发生一些变异或许多变异时,则幼年雄者就会或多或少地具有成年雄者的一些性状;老年雄者和幼年雄者之间的这类差异可以在动物的许多物种中观察到。
幼年雄性动物大概往往倾向于按照下述方式发生变异;即在幼年时期不仅对它们毫无益处,实际上反而有害处——例如获得鲜艳色彩,这将使它们容易被敌人发现,又如获得像巨大的角那样构造,这将使其在发育过程中消耗掉很多生命力。幼年雄者所发生的这类变异,通过自然选择几乎肯定要被排除掉。另一方面对于成年而有经验的雄者来说,由获得这些性状而带来的利益将会抵消冒受危险和损失生命力这两种危害而有余。
有些变异可以使雄者有一个较好的机会去战胜其他雄者,或者去寻求、占有或魅诱异性;如果这样的变异碰巧发生于雌者,由于它们对雌者毫无益处,它们就不会通过性选择在雌者身上被保存下来。关于家养动物,我们也有良好的证据表明,所有种类的变异如果不加以细心的选择,通过杂交以及意外的死亡,就会很快消失掉。因此,在自然状况下,如果上述这类变异偶尔发生于雌性一方并专门在雌性这一方传递,那么这类变异就极其容易消失。然而,如果雌者发生了变异并把它们新获得的性状传递给其后代的雌雄双方,那么那些对雄者有利的性状将会通过性选择被保存下来,结果雌雄双方都会按照同样的方式发生改变,虽然这样的性状对雌者毫无用处;不过以后我还要回头对这些更为复杂的偶然情况进行探讨。最后,通过性状的传递,雌者可能获得而且显然常常获得来自雄者的一些性状。
在生命晚期发生的并只传递给一种性别的变异,如果关系到物种的繁殖,就会被性选择所利用,而且通过性选择被积累起来;因此,同上述相似的变异,虽然关系到日常的生活习性为什么没有常常通过自然选择而被积累起来,乍一看这好像是一个无法解释的事实。如果这种情况发生了,雌雄二者,譬如说为了捕捉猎物和逃避危险,往往会发生不同的改变。雌雄二者之间的这类差异确会偶尔发生,在低等动物中尤其如此。但是,这意味着雌雄二者在生存斗争中遵循不同的习性,对于高等动物来说,这是少见的事情。然而这个情况和生殖机能大不相同,雌雄二者在生殖功能方面必然有差别。这是因为同生殖机能有关的构造变异,常被证明只对一种性别有价值,而且由于这些变异发生在生命晚期,所以只向同一性别传递,这样保存下来和传递下去的变异,便引起了第二性征的发生。
我将在以下几章讨论属于各个纲的动物的第二性征,并努力把本章所阐明的原理应用于每个事例。我们用于讨论最低等动物纲的时间将很短,而对于高等动物,尤其是对于鸟类,则必须用相当的篇幅详加讨论。请注意,由于已经说明的理由,关于雄者用以寻求雌者并在寻得后牢牢把它抓住的无数构造,我只想举出少数例子用做说明。另一方面,关于雄者用以战胜其雄性对手的以及用以魅惑或刺激雌者的全部构造和本能,将予以充分讨论,因为它们在许多方面都是最有趣的。
附录 关于不同纲的动物的雌雄比例数
就我所知,还没有一个人注意过整个动物界雌雄二者的相关数字,因此,我将在这里列举我所能搜集到的有关这方面的材料,尽管这些材料是极不完整的。这些材料所包含的事例只是少数经过实际计算的,而且其数据也不很多。由于只有对人类的这种比例数了解得比较确切,所以我最先列举这些数据作为一个比较的标准。
人 类
在英国从1857—1866年的10年间,出生婴儿存活的年平均数是707120人,男女的比例为104.5∶100。但在1857年全英国的出生率为男婴与女婴之比为105.2∶100;而在1865年这个比例则为104.0∶100。再分别看看一些地区的情况,如白金汉郡(那里每年大约有5000个婴孩出生)在上述整个10年间,其男女出生的平均比例数为102.8∶100;同时在北威尔士(那里平均年出生数为12873人)则高到106.2∶100。再看看一个更小的地区,叫做拉特兰郡(那里年出生数平均只有739人),以女婴出生率为100,1864年男婴出生率为114.6,而在1862年只有97.0。但是,即使在这样小的地方,整个10年间的平均出生数也有7385人,男婴对女婴的比例为104.5∶100,这就是说,这个比例数和全英国的相同。 (52) 由于一些不清楚的原因,这个比例数有时稍受干扰。因此费伊(Faye)教授说:“挪威有些地区在某一个10年间稳定地缺少男孩,而同时在其他一些地区却出现了相反的情况。”在法国,44年间男女的出生比例为106.2∶100;但这一期间曾在某一县出现过五次女婴出生数超过男婴的情况,在另一县曾出现过六次这种情况。在俄国,男婴的平均出生比例为108.9,而在美国的费城男女出生比例则高达110.5∶100。 (53) 比克斯(Bickes)从大约七千万出生婴孩推算出欧洲男女平均出生比例为106∶100。另一方面,关于在好望角出生的白人婴孩,在连续几年里,如以女婴出生率为100,则男婴出生率竟低至90到99之间。有一个奇特的事实:犹太人的男婴出生比例数决定性地大于基督教人的,例如在普鲁士,其比例为113∶100;在波兰的布雷斯劳(Breslau)为114∶100;在利沃尼亚(Livonia)为120∶100;而基督教人在这些地方的男女出生比例则同普通情况一样,例如在利沃尼亚为104∶100。 (54)
费伊教授述说:“如果在母体中和出生时男女死亡的比例相等,则男性所占的数量优势还要更大。但事实是,在几个区域内我们看到,如以死产女婴为100,则死产男婴为134.6到144.9。4~5岁夭折的婴儿,也是男的比女的多;例如在英国,如以1岁死亡女婴为100,则1岁死亡男婴为126。在法国这个比例数更大。” (55) 斯托克顿·霍夫博士根据男孩的发育不完全比女孩更加常见这一情况对上述这些事实作了部分说明。我们从上述中已经知道男性在构造上比女性容易变异;而重要器官的变异一般是有害的。但男婴的身材、特别是其头部都比女婴为大,这又是另一个原因;因为男婴在分娩时将因此更容易受到伤害。因此死产的男婴就更多了;克赖顿·布朗(Crichton Browne)博士 (56) 是一位有高度权威的鉴定家,他认为男婴在出生后的数年内往往会在健康上受到损害。由于男婴在出生时和出生后一段时期内的死亡率过高,又由于成年男人要面临种种危险以及他们向别处迁徙的倾向,所以在保存有统计记录的一切老殖民地方, (57) 发现女性在数量上都比男性占有相当优势。
处于不同环境和气候条件下的不同国家,如那不勒斯、普鲁士、威斯特伐利亚、荷兰、法国、英国以及美国,其非法出生的男婴数量超过女婴的情况要少于合法出生的, (58) 这个事实乍一看好像是难以理解的。不同的作家曾从不同角度解释这种现象,有的认为是由于婴儿们的母亲一般都很年轻,有的认为是第一次怀孕占了很大比例,等等。然而我们已经知道,男婴由于头部较大,在分娩时要比女婴受到较大的损伤;而非法私生婴儿的母亲们一定更容易比其他妇女进行辛苦的劳动,由于种种原因,如紧紧束腰企图遮盖怀孕,繁重的工作,思想的烦恼等等,她们怀的男婴大概要相应受到损伤。关于出生的活男婴同活女婴的比例,不合法私生者要比合法出生者为小的情况,上述大概是一切原因中的一个最有力的原因。就大多数动物而言,成年雄者的大小之所以超过成年雌者,乃是由于较强的雄者在占有雌者的斗争中征服了较弱的雄者;无疑是由于这个事实,至少某些动物的雌雄二者在出生时的大小就不一样。这样,我们便看到一个奇妙的事实,即我们可以把死亡的男婴多于死亡的女婴(非法私生的婴儿尤其如此)这种现象部分地归因于性选择。
人们往往假设双亲的相对年龄决定后代的性别,留卡特教授曾提出, (59) 他认为,关于人类和某些家养动物,有充分证据足以证明这即使不是决定后代性别的唯一因素,也是一个重要因素。另外,有些人曾认为同妇女状况有关的妊娠期是一个有效的原因;但最近的观察结果否定了这个信念。根据斯托克顿·霍夫 (60) 博士的见解,一年中的季节,父母的贫困或富裕,居住于乡村或城市,同外国移民的杂交,等等,对男女性别的比例全有影响。对人类来说,一夫多妻制也曾被假定是导致女婴出生比例较大的原因;但坎贝尔博士 (61) 曾就暹罗的妾妇细心地研究过这个问题,并且断言,一夫多妻下的男婴同女婴的比例和一夫一妻下的情况相同。几乎没有哪一种动物像英国竞赛马那样的高度多配性,可是我们马上就会看到,它们的雌雄后代在数量上几乎完全相等。现在我将列举一些我所搜集到的有关各种动物雌雄比例数的事实,然后对选择在决定这种后果时究竟起了多大作用加以简要的讨论。
马 类
特格梅尔先生曾经如此热心地从“赛马年历”中将竞赛马自1846—1867年这21年间的出生情况给我制成一个表,其中缺1849年的情况,因该年没有发表过出生统计报告。出生总数为25560, (62) 其中包含12763匹公马和12797匹母马,即公马同母马的比例为99.7∶100。由于这些数字相当大,而且是根据全英国各个地方若干年期间的情况统计出来的,因此我们可以充分有信心地作出如下结论:关于家养马,至少是竞赛马,其所产生的雌雄后代在数量上几乎相等。历年中比例的变动同人类在一个人口稀少的小地区所发生的情况密切相似,例如,以母马出生数为100,则1856年公马的出生比例数为107.1,而1857年仅为92.6。统计表里该比例数的变动是有周期性的,因为在连续六年里,公马数量超过母马;而在每次为四年的两个时期内,母马数量又超过公马。然而,这可能是偶然的;至少我从1866年公布的“户口报告”中十年统计表里查不出人类有任何这种情况。
狗 类
从1857—1868年的12年间,全英国大多数细躯猎狗的出生数字均送给《田野》新闻发表;我再一次感谢特格梅尔先生,蒙他细心地把这些结果列制成表。记录下来的出生数是6878,其中包含3605只公狗和3273只母狗,即,公狗和母狗的比例为110.1∶100。最大一次变动发生在1864年,该年公狗和母狗的出生比例为95.3∶100,而1867年,则为116.3∶100。上述110.1∶100这个平均比例对细躯猎狗来说大概是接近正确的,但它是否也符合其他家养品种的情况,在某种程度上还有疑问。卡波勒斯(Cupples)先生曾向一些大养狗家进行过调查,发现他们毫无例外地全都以为出生的母狗比公狗多;但他指出这种看法的发生可能是由于母狗的价值较低,并且由于因此而来的失望在头脑里产生了比较强烈的印象。
绵 羊
农业家们在绵羊出生几个月后给公羊施行阉割的时期才确定其雌雄比例,因而下面的统计并不表示其出生的比例。另外,我发现每年饲养几千头绵羊的苏格兰大饲养家都坚决相信,在出生后的一两年间,公羊的死亡率比母羊高。因此公羊出生时的比例数要比阉割时期的比例数要大些。这一点同我们所看到的人类情况显著符合,而且这两种情况大概都出于同样的原因。我曾从在英格兰饲养低地绵羊(主要是莱斯特羊,Leicesters)的四位先生收到过最近10~16年间的统计报告;其出生总数为8965头,其中包含4407头公羊和4558头母羊;即公羊和母羊的比例为96.7∶100。关于在苏格兰饲养的切维奥特羊(Cheviot)和黑脸绵羊,我也曾收到过六位饲养家的统计报告,其中有两位养羊的规模很大,主要是1867—1869年间的情况,但有些统计则上溯至1862年。记录总数为50685头,其中包含25071头公羊和25614头母羊,即公羊和母羊的比例为97.9∶100。如果我们把英格兰和苏格兰的统计数字加起来,其总数为59650头,其中包含29478头公羊和30172头母羊,即97.7∶100,因而对阉割年龄的绵羊来说,母羊的数量肯定超过公羊,但这个情况大概不适用于其出生时期。 (63)
牛 类
我曾收到九位先生关于982头刚出生的牛犊的统计报告,这个数字太少,不足信赖;该数字包含477头公牛犊和505头母牛犊;即95.4∶100,福克斯牧师告诉我说,1867年在德比郡(Derbyshire)的一个农庄里出生了34头牛犊,其中只有1头是公的。哈里逊·韦尔(Harrison Weir)先生曾向若干养猪者进行过调查,他们大多数都估计出生的公猪和母猪的比例为7∶6。这些先生们还多年饲养家兔,他们注意到生出来的公兔数量远远大于母兔。但是这些估计的价值不大。
关于在自然状况下生活的哺乳类,我知道的很少,至于普通鼠,我曾收过一些互相矛盾的报告。莱伍德(Laighwood)的埃利奥特(R.Elliot)先生告诉我说,有一位捕鼠者向他确言,雄鼠的数量总是大大超过母鼠,即使还在窝里的幼鼠也是如此。结果,埃利奥特先生接着亲自检查了数百只老龄的鼠,证明上面的说法是正确的。巴克兰得先生饲养过大量白鼠,他也以为雄鼠数量大大超过雌鼠。至于鼹鼠(Moles),据说“雄鼠的数量远远超过雌鼠”, (64) 由于捕捉这种动物是一种专门职业,因而这个说法也许是可信的。史密斯爵士在描述一种南非水羚羊(Kobus ellipsiprymnus) (65) 时说道,在这个种和其他种的羚羊群里,公羚羊的数量比母羚羊少:当地土人以为它们出生时的比例数也是如此;另外有些人以为幼小公羚羊是被赶出了群外的,而史密斯爵士说,虽然他本人从未见过仅由幼小公羚羊所成之群,但别人却断言确有这种情形。看来,这些幼小公羚羊一旦被赶出群外,就会被当地许多野兽吃掉。
鸟 类
关于家鸡,我只收到过一份统计材料,即,斯特雷奇(Stretch)先生饲养过交趾鸡(Cochins)的一个精心育成的品系,在八年期间生出1001只小鸡,判明其中487只为公鸡,514只为母鸡,即94.7比100。关于家鸽,有良好的证据可以证明公鸽不是数量过多就是活得更长;因为这等鸽子永远成双成对,特格梅尔告诉我说,独身公鸽的价钱总是比母鸽便宜。在同一窝里下的两个卵所孵出来的两只小鸽通常都是一公一母;但一位大饲养家哈里逊·韦尔先生说道,他常常从同一窝里育出两只公鸽,而很少从同一窝里育出两只母鸽;此外,育出的两只小鸽中,母鸽较弱,更易夭折。
关于自然状况下的鸟类,古尔德先生及其他人士 (66) 都确信公鸟一般要比母鸟多;但由于许多物种的小公鸟同母鸟相类似,所以母鸟数量自然显得比公鸟多。利登赫尔(Leadenhall)的贝克(Baker)先生用野生的雉卵孵出了大量的雉,他告诉詹纳·韦尔先生说,孵出的公雉和母雉的比例一般是4或5比1。一位有经验的观察家述说, (67) 在斯堪的纳维亚,松鸡和黑琴鸡(black-cock)一窝孵出的小鸡,公多于母;而Dal-ripa(一种雷鸟)到求偶场所来的,公比母多;但有些观察家对后一情况的解释是由于被害兽弄死的母鸟比公鸟多。根据塞尔旁(Selborne)的怀特先生所提供的种种事实, (68) 显然英格兰南部的公鹧鸪数量一定大大超过母鹧鸪;有人向我保证说,在苏格兰情况也是如此。韦尔先生向那些在一定季节大批收购流苏鹬(Machetes pugnax)的商人做过调查,据说公鹬的数量要多得多。这位博物学者还为我向捕鸟人做过调查,他们每年都要捕捉数量惊人的各种活的小型鸟供应伦敦市场,一位可信赖的老人毫不迟疑地回答他说,关于苍头燕雀(chaffinch),公的数量大大地超过母的,他认为公和母的比例高达2∶1,至少是5∶3。 (69) 同样地他还坚决主张,用圈套或在夜间用结网方法捕到的鸟鶫,其公鸟的数量远远超出了母鸟。这些说法显然是可信赖的,因为这个人说,云雀、黄嘴朱顶雀(Linaria montana)和金翅雀(goldfinch)的雌雄二者大致相等。另一方面他肯定普通赤胸朱顶雀的公雀大大超过母雀,但超出的数量在不同年份中也有所不同;有些年头他发现母雀和公雀的比为4∶1。但必须记住,主要捕鸟季节到九月份才开始,因此有些物种可能已部分开始迁徙他方,这时期的鸟群往往只含有母鸟。沙尔文(Salvin)先生特别研究过中美洲的蜂鸟,他确信大多数物种是公的占多数。例如,有一年他捕获了属于10个物种的204个样本,其中包含166只公鸟和仅仅38只母鸟。另外有2个物种,是母鸟占多数,但这个比例数不是随着不同季节就是随着不同产地而明显地变化,因为有个时候蝶鸟(Campylopterus hemileucurus)的公和母之比为5∶2,而在另一个时候, (70) 它们的比例则正相反。关于后面这一点,我还要作点补充,波伊斯(Powys)先生发现在科孚(Corfu)和伊皮鲁斯(Epirus)两地苍头燕雀雌雄二者是分开饲养的,而“母鸟的数量最多”,同时特里斯特拉姆先生发现在巴勒斯坦“公鸟群在数量上似乎大大超过了母鸟群”。 (71) 再者,泰勒先生说,在佛罗里达大丘鹬(Quiscalus major)的“母鸟比公鸟的数量少得多”, (72) 而在洪都拉斯,这个比例又是另一种情况,这个物种在那里具有一雄多雌的特性。
鱼 类
关于鱼类的雌雄比例数只有在捕到其成年或接近成年的鱼以后才能确定,因而对此作出任何公正的结论将有许多困难。 (73) 不育的雌鱼可能容易被误认作雄鱼,如冈瑟博士向我说过的鳟鱼情况就是如此。据信有些物种的雄鱼使卵受精后就很快死去。许多物种的雄鱼比雌鱼小得多,因此有大量雄鱼会从捕获雌鱼的同一张网里逃掉。卡邦尼尔(M.Carbonnier) (74) 特别注意过白斑狗鱼(Esox lucius)的自然史,他说,许多雄鱼由于体型小而被较大的雌鱼所吞食;并且他认为,几乎所有鱼类的雄鱼由于同样的原因比雌鱼面临的危险更大。虽然如此,但对雌雄比例数进行过实际观察的少数事例还表明了雄鱼似乎大大超过了雌鱼。例如,斯托蒙特菲尔德(Stormontfield)养鱼实验的负责人布伊斯特(R.Buist)先生说,1865年,为了取卵,最先捕获上岸的70条鲑鱼中,雄鱼之数竟高达60条。1867年他再一次“对这种雌雄数量极不相称的现象给予了注意。在开头时我们捕获的雄鱼和雌鱼的比例是10∶1”。其后,才获得足够的雌鱼以供取卵之用。他接着说,“由于雄鱼的巨大比例,它们在排卵床上彼此不断地进行战斗和厮杀”。 (75) 这种数量的不相称无疑可部分地归因于雄鱼比雌鱼先由海溯游至河,但这是否为全部原因还难肯定。巴克兰(Buckland)先生记述了有关鳟鱼的情况如下,“雄鱼的数量远远超过雌鱼,是一个奇妙的事实,当捕鱼旺季时必然发生的情况是,所捕获的鱼中雄和雌的比例至少是7或8比1。我还不能完全解释这种情形,这是由于雄鱼数量本来比雌鱼多,还是由于雌鱼靠着隐藏而不是靠逃跑以求得安全”。接着他又说,通过仔细搜查沿岸,可以找到足够数量的雌鱼供作取卵之用。 (76) 李(H.Lee)先生告诉我说,在扑次茅斯勋爵的猎园中为了取卵目的所捕获的212条鳟鱼中,有150条是雄的,62条是雌的。
同样地,鲤科(Cyprinidae)的雄鱼在数量上似乎也超过了雌鱼;但这个科的某些成员,如鲤鱼、丁鱥鱼(tench)、欧鳊(bream)和 雅罗鱼(minnow),都正常地实行动物界少见的一雌多雄制,因为雌鱼在排卵时总是有两条雄鱼陪伴左右,而雌欧鳊则有三到四条雄鱼陪伴着。这个事实如此为人所熟知,以致总是劝告在养鱼池中养丁
雅罗鱼(minnow),都正常地实行动物界少见的一雌多雄制,因为雌鱼在排卵时总是有两条雄鱼陪伴左右,而雌欧鳊则有三到四条雄鱼陪伴着。这个事实如此为人所熟知,以致总是劝告在养鱼池中养丁 鱼时,雄和雌的比例应为2∶1,至少是3∶2。至于鲦鱼,一位杰出的观察家说,雄鱼在排卵床上的数量十倍于雌鱼;当有一条雌鱼来到雄鱼当中时,“她马上就被两条雄鱼紧紧夹在中间;当它们在这种局面下经历一段时间后,又有另外两条雄鱼取而代之”。 (77)
鱼时,雄和雌的比例应为2∶1,至少是3∶2。至于鲦鱼,一位杰出的观察家说,雄鱼在排卵床上的数量十倍于雌鱼;当有一条雌鱼来到雄鱼当中时,“她马上就被两条雄鱼紧紧夹在中间;当它们在这种局面下经历一段时间后,又有另外两条雄鱼取而代之”。 (77)
昆 虫 类
在这个巨大的纲里,几乎只有鳞翅目(Lepidoptera)可以用来判断雌雄二者的比例数。这是因为有许多著名观察家曾特别细心地收集这个“目”的昆虫,并从卵或幼虫状态大量把它们繁殖起来。我曾希望有些养蚕者会保存一个确实的记录,但经过写信到法国和意大利并查阅了各种文献之后,我并没有找到过这方面的材料。一般的意见好像是雌雄二者接近相等,但在意大利,我听卡内斯垂尼(Canestrini)教授说,许多饲养者都认为生出来的雌虫数量超过雄虫。然而这位博物学家还告诉我说,臭椿蚕(Bombyx cynthia)为一年两化,在第一造中雄蚕数量大大超过雌蚕,而在第二造中雌雄二者的数量接近相等,或雌蚕稍多。
关于自然状况下的蝴蝶,其雄者的数量显然占巨大优势,这曾使若干观察家留下深刻的印象。 (78) 例如,贝茨(Bates)先生 (79) 在提到某些产于上亚马孙(Upper Amazons)的100个左右的物种时说,雄虫的数目大大超过雌虫,甚至其比例达到100∶1。在北美,具有丰富经验的爱德华兹(Edwards)估计在凤蝶属(genus Papilio)中,雄虫和雌虫的比例为4∶1;把这一点告诉我的沃尔什(Walsh)先生说,图尔努凤蝶(P.turnus)的情况正是这样。特里门(R.Trimen)先生在南非发现有19个物种都是雄虫占多数; (80) 其中有一个群集于开阔地带的物种,估计其雄虫的数量为雌虫的50倍。还有另一个物种,其雄虫在某些地方为数至多,以致他在七年间只收集到五只雌虫。波旁(Bourbon)岛的马亚尔(M.Maillard)说,凤蝶属的一个物种,其雄虫比雌虫多达20倍。 (81) 垂门先生告诉我说,就他本人所见到的或听到的来说,很少有一种蝴蝶的雌虫数量超过雄虫的;但有三个南非的物种也许是例外。华莱士先生述说, (82) 在马来群岛,鸟翼蝴蝶(Ornithoptera craesus)的雌虫比雄虫常见,也较容易抓到;但这是一种稀有的蝴蝶。我在这里还要作点补充,盖内(Guenée)说,从印度送来的红蛾(Hyperythra,蛾类的一个属)采集品,其雌虫为雄虫的4~5倍。
当把这个昆虫雌雄比例数问题提到昆虫学会进行讨论时, (83) 一般都承认捉到的鳞翅目大多数成年的、即成虫状态的雄虫在数量上超过雌虫,但各种各样的观察家都把这个事实归因于雌虫比较隐匿的习性和雄虫从茧里羽化较早。众所周知,大多数鳞翅目的昆虫以及其他种类的昆虫都有这种情形发生。因此,正如佩尔索纳(M.Personnat)所说的,家养天蚕(Bombyx Yamamai)的雄虫在交配季节开始时并无用处,雌虫在交配季节结尾时因缺少配偶也无用处。 (84) 然而,上述某些蝴蝶在其产地极其普通,这些原因是否可以把其雄虫占大多数的问题解释得足够清楚,很难使我信服。斯坦顿(Stainton)先生多年来对小蛾类给予了密切注意,他告诉我说,当他搜集到的蛾子处于成虫状态时,他以为雄虫的数量为雌虫的十倍,但是,自从他把这些蛾子由幼虫状态大批养育以来,他相信雌虫就占多数了。若干昆虫学家都赞同这个观点。然而,道布尔戴伊(Doubleday)先生以及一些其他人士则持相反的观点,他们确信在他们从卵和幼虫养育的成虫中,雄虫比雌虫所占的比例数为大。
除了鳞翅目雄虫有较大的活动习性,较早从茧里羽化,以及在某些场合中群集于较开阔的地带等这些原因外,关于在成虫状态捕获的鳞翅目昆虫以及从卵或幼虫状态养育起来的鳞翅目昆虫,在雌雄比例方面所存在的明显的或真实的差异,还可以举出其他原因。我听卡内斯垂尼教授说,意大利许多养蚕者都相信蚕蛾的雌性幼虫比雄的更多遭到近代疾病的危害;而斯托丁杰(Staudinger)博士告诉我说,在饲养鳞翅目昆虫时,死在茧里的雌虫比雄虫为多。许多物种的雌性幼虫比雄的大,昆虫采集者自然要挑选最好的标本,这样就会无意识地采集到大量的雌虫。有三位采集者曾告诉我,他们的实践情况就是这样;但华莱士博士确信,大多数采集者如果能够找到比较稀有的种类,他们就会把全部标本都采集下来,因为只有这些稀有种类才值得他们花工夫去饲养。当鸟类遇到幼虫时,大概要把最大的幼虫吞食掉;卡内斯垂尼教授告诉我说,意大利有些养蚕者相信臭椿蚕的第一化幼虫中,被黄蜂消灭的雌虫数量超过雄虫很多。华莱士博士进一步说道,雌性幼虫因比雄的大,所以需要较多的发育时间并消耗较多的食物和水分;因而就要有更长的时间遭到姬蜂(ichneumons)和鸟类等带来的危险,在荒歉之年就会更大量地死亡。因此在自然状况下,鳞翅目雌虫达到发育成熟的数量,很可能要比雄虫少得多;对我们的特殊目的来说,我们所关心的是,当雌雄二者就要繁殖其种类时,即在其成熟时它们的相对数量。
某些蛾类大批雄虫聚集于单独一只雌虫周围的方式,清楚地表明了雄虫的巨大多数,虽然这个事实也许可以由雄虫较早从茧里羽化而得到解释。斯坦顿先生告诉我说,经常可以见到有10~20只筒蛾(Elachista rufocinerea)的雄虫聚集在一只雌虫周围。众所周知,如果把栎枯叶蛾(Lasiocampa quercus)或鹅耳枥天蚕蛾(Saturnia carpini)的一只未交配过的雌虫摆在一个笼子里,大批雄虫就会聚集在它周围,而且如果把雌虫关在一个房间里,雄虫甚至会从烟囱跑下来找它。多勃尔德伊先生相信,仅在一天之内他就见到一只关起来的雌虫吸引来50~100只雄虫。垂门先生把几天前就关着一只枯叶蛾雌虫的盒子放在怀特岛上,马上就有五头雄虫竭力向盒子里钻。在澳大利亚,韦雷奥(M.Verreaux)把放有一只小雌蚕的盒子搁在口袋里,于是招来了一群雄虫跟着他,因而约有200只雄虫随他一起飞进房中。 (85)
道布尔戴先生叫我注意斯托丁杰的鳞翅目价目表, (86) 上面开列了蝴蝶(锤角亚目Rhopalocera)的300个物种或特征十分显著的变种,雄虫和雌虫价格均被标明。很普通的物种的雄虫和雌虫的价格当然都是一样的;但有114个稀有物种,其雄虫和雌虫的价格却不相同;除了一个例外,所有这些物种的雄虫都比雌虫的价格便宜。以这113个物种的价格加以平均,雄虫和雌虫价格之比为100∶149;这一点显然表明雄虫的数量正好成反比地超过雌虫。编入目录的蛾类(缰翅亚目,Heterocera)约有2000个物种或变种,由于雌雄二者习性不同而导致雌虫无翅的种类未编入内。在这2000个物种中,有141个物种的价格因性别而异,其中130个物种的雄虫价格比雌虫便宜,只有11个物种的雄虫比雌虫贵。这130个物种的雄虫平均价格和雌虫平均价格之比为100∶143。道布尔戴先生(在英国没有任何人的经验胜过他)认为,雌雄二者价格的不同和这些物种的生活习性没有任何关系,而只能归因于雄虫数量超过雌虫。但我必须补充一点,即斯托丁杰博士告诉我说,他本人持有不同的意见。他以为由于雌虫的习性比较不活泼,并且由于雄虫从茧里羽化较早,可以说明昆虫采集者所获得的雄虫数量多于雌虫,其结果就使雄虫的价格较低。关于从幼虫状态养育起来的样本,斯托丁杰博士相信,如上所述,雌虫死于茧内者的数量远比雄虫为多。他接着说,关于某些物种,某一性别在某些年间似乎比另一性别占有数量优势,但并非永远如此。
表 2-1

关于对鳞翅目昆虫的直接观察材料,不论是从卵或从幼虫养育起来的,我仅收到如表2-1所示的少数事例。
因此,从这七组茧和卵产生的雄虫数量超过了雌虫,合计雄虫和雌虫的比例为122.7∶100。但整个数据不够大,几乎不足为凭。
总之,证据的来源虽有不同,但均指着同一方向,因此我推论鳞翅目的大多数物种,不论最初从卵孵化时的雄虫比例大小如何,其成熟雄虫的数量一般都比雌虫多。
至于昆虫的其他“目”,我收集到的可靠材料很少。关于鹿角锹甲虫(Lucanus cervus),“其雄虫数量看来比雌虫多得多”;但如科内利乌斯(Cornelius)于1807年所说的,当在德国某个地方异常大量发生这类甲虫时,雌虫数量约超出雄虫六倍。关于叩头虫科(Elateridae)的某一类,其雄虫数量据说多于雌虫,而且“经常可发现两三只雄虫和一只雌虫同在一起; (87) 因此在这里实行的似乎是一雌多雄制”。属于隐翅虫科(Staphylinidae)的扁蟞(Siagonium),其雄虫有角,而“雌虫的数量远远超过雄虫”。詹森(Janson)先生在昆虫学会说道,有一种吃树皮的多毛髓虫(Tomicus villosus),其雌虫多得成灾,而雄虫则少得几乎无人知道。
昆虫的某些物种甚至某些群的雄虫因为无人见过或为数极少,而雌虫又是孤雌生殖的,也就是不需性结合的生殖,因此任何关于其性别比例的议论都几乎没有什么价值;瘿蜂科(Cynipidae)的若干种类提供了这类例子。 (88) 沃尔什先生所知道的形成虫瘿的瘿蜂科昆虫,其雌虫数量为雄虫的4~5倍;他还告诉我说,形成虫瘿的瘿蚊科(双翅目)昆虫的情况也是如此。关于叶蜂科(Tenthredinae)昆虫的一些常见物种,史密斯先生曾从各种大小的幼虫养育成上百个标本,然而从未养出过一只雄虫。另一方面,柯蒂斯(Curtis)说, (89) 他繁育的某一物种——菜叶蜂(Athalia),其雄虫和雌虫之比为6∶1;而同时在田野里捕到的这同一个物种的成熟成虫,其雄虫和雌虫之比正好相反。赫尔曼·米勒 (90) 在蜜蜂科中采集了许多物种的大量标本,并且从茧养育出好多其他标本,然后计算其性别。他发现有些物种的雄蜂数量大大超过雌蜂;而另外一些物种则出现相反情况;还有些物种的雌雄二者接近相等。但同多数情况一样,雄蜂从茧里羽化要早于雌蜂,因而在繁殖季节开始时的数量实际上超过雌蜂。米勒还观察到有些物种在不同产地其雌雄的相对数量大不相同。但是,正如米勒亲自向我陈述的,由于其中一种性别可能比另一种性别更难于被观察到,因此在采纳这些意见时必须小心从事。这样,他的兄弟弗里茨·米勒在巴西曾注意到,同一种蜜蜂的雌雄二者有时群集于花的种类互不相同。关于直翅目(Orthoptera)昆虫,其雌雄二者的相对数量,我几乎一无所知。然而,克尔特说, (91) 他检查过500只蝗虫,其中雄虫和雌虫之比为5∶6。关于脉翅目(Neuroptera),沃尔什先生说,在蜻蜓这个类群的许多物种中,雄虫数量大大超过雌虫,但决非所有物种都是如此。还有一个蜻蜓属(Hetaerina),其雄虫数量一般至少为雌虫的四倍。另外箭蜓属(Gomphus)的某些物种,其雄虫多于雌虫的倍数和上面相同,但另有两个物种,其雌虫数量则为雄虫的2~3倍。关于啮虫属(Psosus)的某些欧洲物种,在采集到的几千只雌虫中可能找不到一只雄虫,但同时,这一属的其他物种,其雌雄二者均系常见。 (92) 麦克拉克伦(Mac Lachlan)先生在英格兰捉到过几百只雌性异幻吸虫(Apatania muliebris),但没有见过一只雄虫;至于雪蠍蛉(Boreus hyemalis),在我们这里见到过的雄虫不过四五只。 (93) 关于大多数这些物种(叶蜂科除外)的雌虫是否属于孤雌生殖的类型,目前还不能证实;由此可见,我们对引起雌雄二者的比例出现如此明显不一致的原因是多么无知。
关于有铰类(Articulata)动物的其他一些纲,我所搜集到的材料还要少一些。布莱克瓦尔先生对蜘蛛纲曾仔细进行过多年观察,他写信告诉我说,雄蜘蛛由于其游动的习性,较为常见,从而显得数量较多。少数蜘蛛种的情况确是这样;但他提到六个属的几个物种,其雌蜘蛛数量似乎比雄的多得多。 (94) 雄蜘蛛的体型比雌的小(这个特性有时会极度发达),并且它们的外貌大不相同,这些情况在某些事例中可能说明它们的采集品为何稀见。 (95)
有些低等甲壳类能进行无性繁殖,这可以说明其雄虫为何极端罕见。冯·赛保德 (96) 曾仔细调查过来自21个产地的不下13000个鲎虫(Apus)标本,他在其中只找到319只雄虫。正如弗里茨·米勒告诉我的,关于另外一些类型如异足虫属(Tanais)和介虫属(Cypris)我们有理由相信其雄虫比雌虫短命得多;这一点大概可以说明雄虫为何稀少,假定雄虫数量一开始就同雌虫相等的话。另一方面,米勒在巴西海岸上采集到的针涟虫科(Diastylidae)和海萤属(Cypridina)的雄虫永远比雌虫多得多。例如在同一天内捉到的后面这一属的一个物种的63个标本中就有57只雄虫;但他认为这个数量优势可能是由于雌雄二者在生活习性上某种尚未弄清的差异所致。关于一种高等巴西蟹,即招潮蟹(Gelasimus),弗里茨·米勒发现其雄者远比雌者的数量为多。根据斯彭斯·巴特先生的丰富经验,有6种常见的英国蟹的情况似乎正好相反,他曾向我说过这6种蟹的名称。
自然选择和雌雄比例的关系
我们有理由设想在某些场合中,人类通过选择曾间接地影响了其自身产生雌雄的能力。有些妇女在其一生所生育的孩子中有一种性别多于另一种性别的倾向,这种情况也适用于许多动物,如牛和马。因此“耶尔德斯雷”公所的赖特先生对我说,他有一匹阿拉伯母马,虽然7次分别与不同的公马交配,仍生下7匹小母马。我在这方面所掌握的证据虽然不多,但根据类推方法可使我相信,专门产生任何一种性别的倾向几乎就像其他每种特性一样,例如,生双胞胎的特性,大概是可以遗传下去的。关于上述倾向,著名权威唐宁(J.Downing)先生写信对我说,似乎可以证明这一点的事实确曾在某些短角牛的族系中发生过。马歇尔上校 (97) 经过仔细调查后最近发现印度有一个山地部落叫托达人(Todas) (98) 的,其一切年龄的人口是由112个男人和84个女人组成的——即男女之比为133.3∶100。托达人的婚姻是一妻多夫制,过去一定实行杀害女婴;但这种风俗目前已停止一个相当时期了。在晚近几年内生育下来的婴儿中,男多于女,其比例为124∶100。马歇尔上校用下面巧妙的方式说明这个事实。“为了说明问题,让我们举出三个家庭作为整个部落的一般代表。比方说一位母亲生育了6个女儿,而没有生儿子。第二位母亲只生了6个儿子。第三位母亲生了3个儿子,3个女儿。按照部落的风俗,第一位母亲杀死4个女儿,保留两个。第二位母亲保留了她6个儿子。第三位母亲杀死2个女儿,保留1个女儿和3个儿子。那么在这三个家庭中总共有9个儿子和3个女儿,由他们来传宗接代。然而当这些男人属于那些生男倾向大的家庭时,则这些家庭中生女的倾向就要小。这种倾向逐代加强,直至像我们所见到的那样,那些家庭就逐渐惯常地生男多于生女了”。
如果我们假定一种产生雌雄的倾向是遗传的,那么杀婴的风俗就几乎肯定要引起上述后果。但是,由于上述数据极为不足,所以我曾搜寻进一步的证据,但不能决定我找到的证据是否可靠。尽管如此,这些事实也许还值得一提。新西兰的毛利人长期以来就实行杀婴,范东先生说 (99) ,“他曾碰到有些妇女弄死了4个、6个甚至7个婴孩,其中多数是女婴。然而,根据最好判断所得到的普遍证据决定性地证明了,这种风俗多年以来几乎已经绝灭。这种风俗的消亡时期大概可以定为1835年。”目前在新西兰人中,正如托达人的情况一样,男性出生数超过女性很多。范东先生述说,“有一个事实是肯定的,虽然无法确定这种男女不成比例的奇特情况是在什么确切时期开始的,但十分明显的是,这种女人减少的过程在1830—1844年间已达到全盛时期,而1844年的未成年人口当时正好出生,并以巨大活力延续到现在”。 (100) 下面的叙述引自范东先生, (101) 但由于数据不够充分,调查不够精确,因此不能期望获得一致的结果。应该记住在这个场合以及下述场合中,每个地方人口的正常状态都是女多于男,至少在所有文明国家里是如此,这主要由于男性在青少年时期的死亡率较高,部分由于在晚年会遇到各种意外事故。1858年新西兰一切年龄的土著人口估计共含男性31667人,女性24303人,即男和女之比为130.3∶100。但在这同一年里,在某些限定地区内,经过非常仔细核实过的数字表明,一切年龄的男性为753人,女性为616人,即男和女之比为122.2∶100。对我们更为重要的是,在1858年这同一年里,同一个地区内的未成年男性为178人,而未成年女性为142人,即125.3∶100。还可以补充,1844年这一年杀害女婴的风俗仅在不久前才停止,某一地区的未成年男性为281人,而未成年女性只有194人,即男和女之比为144.8∶100。
在桑威奇群岛,男人的数量超过女人。该地以前盛行杀婴达到了可怕的程度,但是,正如埃利斯(Ellis)先生所指出的, (102) 还有斯塔雷主教和寇恩(Coan)牧师告诉我的,所杀害者并不限于女婴。尽管如此,另一位显然可以信赖的作者贾夫斯(Jarves)先生 (103) 曾对整个群岛进行过观察,他还说:“可以找到不少妇女,她们承认自己杀死的婴儿有三至六个或八个之多”;他接着说,“女性因被认为比男性的用处较少,更常被弄死”。从我们所知道的世界其他地方发生的情况来看,这个说法是可能的,但要采纳这个说法就得非常谨慎。停止实行杀婴,约在1819年,当时该群岛废除偶像,传教士已经定居下来。1839年曾对考爱岛(Kauai)和奥阿胡岛(Oahu)的一个地区成年的和纳税的男子与女子进行了仔细调查,结果是男性为4723人,女性为3776人,即125.08∶100。同时考爱岛未满14岁和奥阿胡岛未满18岁的男子为1797人,同龄的女子为1429人,在这里男女的比例为125.75∶100。
1850年对所有岛屿的调查表明, (104) 一切年龄的男子总数为36272人,女子33128人,即109.49∶100。未满17岁的男子总数为10773人,同年龄的妇女为9593人,即112.3∶100。根据1872年的调查,一切年龄男女(包括混血儿)之比为125.36∶100。必须记住,所有这些关于桑威奇群岛的统计报告只提供了现存男子和现存女子的比例,并不是出生人口的比例。根据所有文明国家的情况来判断,如果以出生数为据,则男子的比例数还要大得多。 (105)
根据上面的几个事例,我们有理由相信上述杀婴的实行,有助于形成一个产生男性较多的种族。但我决非假定,人类实行杀婴或者其他物种的相似过程乃是男性数量过多的唯一决定性原因。可能有某种未知的法则在人口下降的种族中导致了这种结果,而这个种族的生育力已经多少降低了。除了上面提到的几个原因之外,未开化人的分娩比较顺利,结果其男婴受到的伤害较少,这大概有助于提高产后存活的男婴对女婴的比例。如果我们可以根据最近尚存的为数不多的塔斯马尼亚人后代的特性和居住在诺福克岛上的塔希提人的杂种后代的特性来作判断的话,那么无论怎样说,未开化人的生活和男性数量显著过多之间似乎并不存在任何必要的关联。
由于许多动物的雄者和雌者的习性多少有些不同,而且所面临的危险的程度也不一样,因此,在许多场合中,常常遭到毁灭的一种性别大概要比另一种性别多。但就我所能追查出的各种原因的复杂关系而言,任何一种性别的没有差别的、虽然是重大的毁灭,都无助于改变物种产生性别的能力。至于严格的社会性动物,如蜜蜂或蚂蚁,其不育的和能育的雌虫数量要比雄虫庞大得多,雌虫这种数量优势具有无比的重要性,我们可以看到在任何这等群体中,凡是雌虫具有一种强烈遗传倾向以生产越来越多的雌性后代者,其群体就能最好地繁盛起来;而且在这种场合中,一种产生不相等性别的倾向大概会通过自然选择而被获得。关于群居的动物,有雄者在前面保卫其群体者,如某些狒狒和北美野牛,可以想象到,产生雄者的倾向大概可以通过自然选择而被获得,因为得到更好保卫的那些群的个体将会留下较多的后代。以人类来说,由于男人数量在部落中占有优势而发生的利益,可能就是实行杀害女婴的一个主要原因。
就我们所知道的来说,凡生产雌雄数量相等或生产某一性别超过另一性别的遗传倾向,能使某些个体较其他个体获得直接的利益或害处者,尚无一例。譬如说,有一个个体具有生产雄者多于雌者的倾向,这并不会使它在生存斗争中比其他具有相反倾向的个体得到更大成功,因此这样一种倾向不能通过自然选择而被获得。尽管如此,还有某些动物(如鱼类和蔓足类)在雌者受精的过程中,看来需要有两个以上的雄者参加,因而雄者在数量上占有很大优势,但这种产生雄者的倾向是怎样获得的,其原因还不清楚。我过去曾以为如果生产相等数量的雌雄二者这一倾向对于物种有利,那么它一定会通过自然选择而发生,但我现在认识到了整个问题的复杂性,因此把它留待将来去解决会更妥当些。
第九章 动物界低等纲的第二性征
最低等的动物纲缺少次级性征——灿烂的色彩——软体动物——环形动物——甲壳类,次级性征的强烈发达;二态现象:色彩;成熟以前未曾获得的诸性状——蜘蛛,其雌雄色彩;雄者的摩擦发音——多足类
凡是属于低等诸纲的动物,其雌雄两性结合于同一个体之内者并不罕见,因此第二性征在它们当中不能发育。在雌雄分离的许多场合中,二者都永久地附生于某种支座上,某一方不能寻找另一方也不能为占有另一方而进行斗争。再者,几乎肯定的是,这些动物的感觉器官太不完善,而且心理能力也太低,以致不能彼此欣赏对方的美或其他魅力,也不会感觉到同性之间需要竞争。
因此,我们必须考虑的那种第二性征在原生动物、腔肠动物、棘皮动物以及蠕形动物(Scolecida)等这些纲或亚界中都不会发生;这一事实同关于高等诸纲的第二性征是通过性选择而获得的那一信念相符合,而性选择则依赖于雌雄任何一方的意志、欲望和选择。尽管如此,依然会发生少数明显的例外;例如,我听贝尔德(Baird)博士说,某种体内寄生虫的雄者和雌者的颜色稍有差异;但我们没有理由设想这些差异是由于性选择而被加大的。雄者用来抱握雌者的器官,乃是物种繁殖必不可少的,却同性选择无关,而是通过自然选择获得的。
许多低等动物,无论是雌雄同体还是雌雄异体,都饰以最灿烂的色彩,或具优雅的色调和条纹。例如,许多珊瑚虫和海葵(Actiniae),某些水母(Medusae)、银币水母(Porpita)等等,某些真涡虫(Planariae)、许多海盘车(star-fishes)、海胆类(Echini)、海鞘类(Ascidians)等等。但我们根据已经指明的理由,即,这些动物中有的是雌雄同体,有的是永久附生在其他东西上,以及所有它们的心理能力都低;可以断定这等色彩并不是作为一种性的吸引力,也不是通过性选择而被获得的。应当记住,除非某一性别比另一性别的色彩灿烂得多或鲜明得多,而且除非雌雄之间在习性上没有足够的差异以阐明其色彩的不同,否则我们就没有充分的证据可以证明其色彩是通过性选择而获得的。但是,只有当那些更富于装饰的个体,几乎总是雄者如此,主动在另一性别面前夸示其魅力的时候,其证据才可称为完全;因为我们无法不相信这样夸示是无用的,如果这是有利的,那么性选择几乎不可避免地就会跟着发生作用。然而,当雌雄二者的色彩相同时,如果它们的色彩仅仅和同一类群的某些其他物种的某一性别的色彩明显相似,那么我们就可以把这一结论扩及雌雄双方。
那么,我们怎样来阐明最低等纲的许多动物那种美丽的甚至是灿烂的颜色呢?这等色彩是否常作为一种保护,好像还有疑问;但只要读一读华莱士先生关于这个问题的卓越论著,谁都会承认我们在这个问题上多么容易陷于错误。例如,任何人最初看到水母类的透明性时,大概都不会认为这对保护它们自己有最大益处。但是,赫克尔提醒我们注意,不但是水母,而且许多浮游软体动物、甲壳动物、甚至是小的海洋鱼类都有这种相同的往往带有彩虹色的透明外貌,这样,我们就几乎无法怀疑它们正是这样逃避了海洋鸟类以及其他敌害的注意。贾尔(M.Giard)也认为,某些海绵类和海鞘类的明亮色彩乃是作为一种保护之用。 (106) 明显的色彩对许多动物同样也是有利的。这可以用来警告那些攫食之敌,它们的味道不好或是具有某种特别的防御手段;但为了方便起见,有关这个问题将留在后面去讨论。
由于对大多数最低等动物的知识贫乏,我们只能说,它们的明亮色彩或是由其组织的化学性质所引起的,或是由其组织的细微构造所引起的,而同这种明亮色彩所产生的任何利益无关。几乎没有任何颜色比动脉的血更为漂亮的了;但没有理由来设想这种血的颜色本身具有任何利益;虽然这会给少女的双颊增添美丽,但谁也不会妄说它是为了这个目的而被获得的。又如许多动物,特别是低等动物,其胆汁的颜色富丽。例如,汉考克(Hancock)先生告诉我说,无壳的海参类(Eolidae)是极其美丽的,这主要是因为透过其透明的外膜可以见到胆汁的腺体——这种美丽对这些动物大概不会有什么益处。美洲森林里凋谢的树叶色调,被所有人描写得灿烂耀眼,但没有人认为这等色调对树木有任何一点利益。请记住,化学家们最近合成的同天然有机化合物密切近似的物质何等之多,它们显示出最华丽的颜色,那么在活有机体的复杂实验室中如果没有经常创造出同样颜色的物质,那就是一件奇怪的事情了,虽然它们没有由此得到任何益处。
软体动物亚界
在动物界的这整个大部门中,就我所能发现的来说,绝没有本书所考察的那样第二性征。三个最低等的纲,即海鞘类、苔藓虫类(Polyzoa)和腕足类(Brachiopods)(构成某些作者所谓的拟软体动物门),也不能期望它们有第二性征,因为大多数这等动物都是永久地附生在一个支座上的或是雌雄同体的。在瓣鳃纲(Lamellibranchiata)即双壳类中,雌雄同体并不罕见。紧接的较高一级为腹足纲(Gasteropoda),即单壳类,有雌雄同体的,也有雌雄异体的。但是,在雌雄异体的场合中,雄者从未有过用以寻求、抱持或媚惑雌者的特别器官,也不具有同其他雄者斗争的特别器官。格温·杰弗里斯(Gwyn Jeffreys)先生告诉我说,其雌雄之间的唯一外部差异有时仅仅表现在贝壳形态略有不同;例如,雄滨螺(Littorina littorea)的壳比雌者的狭些,螺旋线细长些。但可以假设,这种性质的差异直接同生殖行为或卵的发育有关。
腹足类动物虽能运动,并且具有不完善的眼睛,但似乎并不赋有足够的心理能力来和同性诸成员在竞争雌者中互相搏斗,这样就不能由此获得次级性征。尽管如此,有肺腹足类动物即蜗牛类(land-snails)在交配之前,还有一个求偶的过程;因为这等动物虽是雌雄同体,但迫于它们的构造还要互相交配。阿加西斯述说,“凡是观察过蜗牛求偶活动的人们都不会怀疑,这种雌雄同体的动物在实行双重交配的过程中存在着对异性进行魅惑的行为”。 (107) 这等动物在某种程度上似乎也容易持久地互相依恋。朗斯代尔(Lonsdale)先生是一位精确的观察家,他告诉我说,他曾把一对罗马蜗牛(Helix pomatia)放在一个食物缺乏的小花园里,其中一只很衰弱。经过短时间后,那个健壮的个体不见了,留下一道有黏液的足迹,原来它翻过了一道墙来到相邻的一个食物丰富的花园里。朗斯代尔先生断定它已将其有病的伴侣抛弃了;但过了24小时后它又回来了,而且显然向其伴侣传达了它的有成效的勘察结果,因为它们于是沿着原路消失在墙外了。
即使软体动物中最高等的纲,像雌雄异体的头足类(Cephalopoda)、即乌贼,就我所能发现的来说,也不具有现在所说的那种次级性征。这是一个令人感到奇怪的情况,因为,凡是见过它们怎样巧妙地逃避一种敌害的人们都会承认这等动物具有高度发达的感觉器官和相当的心理能力。 (108) 然而,某些头足类动物具有一种异常的性征,即,雄性生殖素先集中于一条臂或触手内,这条触手随即断落,依其吸盘附着于雌者,并在一段时间内营独立生活。断落的这条触手同一个独立的动物如此完全相似,以致居维叶把它描述为一种寄生虫,称其为交接腕(Hectocotyle)。但是,把这种奇异的构造归入次级性征,倒不如把它归入初级性征更为合适。
性选择虽然对软体动物似乎不起作用,但是像涡螺、芋螺、扇贝等许多单壳类和双壳类的颜色和形状都很美丽。在大多数场合中,颜色好像没有什么保护作用,正如最低等动物纲的情况那样,颜色大概是组织性质的直接结果,贝壳的样式和刻纹取决于它的生长方式。光的量似乎有一定程度的影响,因为,格温·杰弗里斯先生虽然反复讲过生活在深水里某些物种的贝壳颜色是明亮的,然而我们一般所看到的是其底面以及由套膜所遮盖的部分,其颜色不及上部受光表面的颜色浓。 (109) 在某些场合中,例如生活在珊瑚或色调明亮的海草中的贝类,其明亮的颜色可能是作为保护自己之用的。 (110) 但是,在奥尔德(Alder)和汉考克(Hancock)两位先生的出色著作中可以看到,许多裸鳃软体动物或者海参(sea-slugs)的颜色之美丽同任何贝类无异;根据汉考克先生热心给我的材料看来,这等颜色通常是否作为保护自己之用,似乎极可怀疑。对某些物种来说可能有这种作用,例如有一类是生活在海藻的绿叶上,其本身的颜色也是碧绿的。但许多颜色明亮的、白色的或其他有鲜明颜色的物种并不寻求隐蔽;另外还有某些同等鲜明颜色的物种以及其他暗色的种类生活在石头底下和幽暗的深处。因此对于裸鳃软体动物来说,它们的颜色同其栖息场所的性质显然没有什么密切关系。
这些无壳的海参都是雌雄同体,但它们互相交配,这同蜗牛类的情况一样,而许多蜗牛都有极漂亮的壳。可以这样想象:雌雄同体的两个个体彼此被更富有魅力的美所吸引,因而结合起来并留下后代以继承双亲的更富有魅力的美。但对于体制如此低等的动物来说,这是极不可能的。而且也完全看不出来自一对比较美丽的雌雄同体动物的后代,怎么会比来自一对比较不美丽的雌雄同体动物的后代占有任何优势,以增加其数量,除非精力和美丽的确普遍相符合,就不会有上述情况发生。在这里,我们还没有考虑下述事例,即一定数量的雄者比雌者早熟,以及比较美丽的雄者被精力比较旺盛的雌者所选中。诚然,就有关一般生活习性来说,如果美丽的颜色对于一种雌雄同体动物是有利的,那么,色调比较明亮的个体大概会获得最大成功,并且增加其数量,但这是自然选择而不是性选择的事例。
蠕形动物亚界:环节动物
在这个纲中,当雌雄异体时,雌雄二者有时在如此重要的性状上彼此有所差异,以致会把它们归入不同的属甚至不同的科,虽然如此,这似乎不是可以稳妥地归因于性选择的那种差异。这些动物往往都有美丽的颜色,但由于雌雄二者在这方面并无区别,因此我们很少考虑它们。即使纽形动物(Nemertians),虽然其体制如此低等,“在美丽和颜色的丰富多彩方面也可同无脊椎系列中的任何其他类群相竞争”;然而麦金托什(McIntosh) (111) 博士未能发现这些颜色有什么用途。按照夸垂费什的意见,固定不动的环节动物在生殖时期过后,其颜色就变得暗淡了; (112) 我认为这一点可能是因为它们在那时处于比较不活跃的状况。所有这一切蠕虫状的动物显然都由于太低等,以致雌雄双方的个体都不能尽力去选择一个对象,或者,同一性别的个体也不会在竞争对象中互相搏斗。
节足动物亚界:甲壳动物
在这巨大的纲中,我们首先遇到的是,常常以一种显著方式发育的无可怀疑的第二性征。不幸的是,我们对甲壳动物的习性了解得很不全面,而且还解释不了某一性别所特有的许多构造的用途。关于低等寄生性物种,其雄者体型小,而且只有雄者具有完善的游泳肢、触角和感觉器官;雌者缺少这等器官,其躯体往往只是扭曲的一团。但是,雌雄之间这等异常的差异无疑同它们的广泛不同的生活习性有关,因此不在我们考虑之内。不同科的各种甲壳动物的前触角具有特殊的线状体,这些线状体据信可起嗅觉器官的作用,而雄者的线状体数量远比雌者的为多。即使雄者的嗅觉器官并不特别发达,它们几乎肯定迟早也能找到雌者,因此,嗅觉线状体大概是通过性选择而增加了数量,这是因为具有更多线状体的雄者在寻找对象和产生后代方面都能获得较大成功。弗里茨·米勒描绘过异足虫属(Tanais)的一个显著二态的物种,其雄者有两种不同的类型,决无中间类型存在。其中一种类型具有数量较多的嗅觉线状体,另一种类型则具有较强的而且较长的钳爪或螯,用以抱持雌者。弗里茨·米勒认为这同一物种的两个雄性类型之间的这等差异可能起源于某些个体在嗅觉线状体的数量上发生了变异,同时另一些个体则在钳爪的形状和大小上发生了变异;因此前者能够最有效地寻找雌者,而后者则能最有效地抱持雌者,它们都会留下最大数量的后代以承继各自的优越性。 (113)
在某些低等甲壳动物中,雄者的右前触角和左前触角的构造大不相同,左前触角的简单圆锥状关节同雌者的触角相类似。雄者那条变异了的触角不是中间膨大就是呈一定角度的弯曲,不然就是变成某种优雅的有时是异常复杂的抱握器官(图4)。 (114) 我听卢伯克爵士说,它是用来抱持雌者的,而且为了同一目的,身体同一侧的两条后肢(b)也变成了一种钳状物。在另一科中,只有雄者的下触角,即后触角呈“奇妙的锯齿状”。
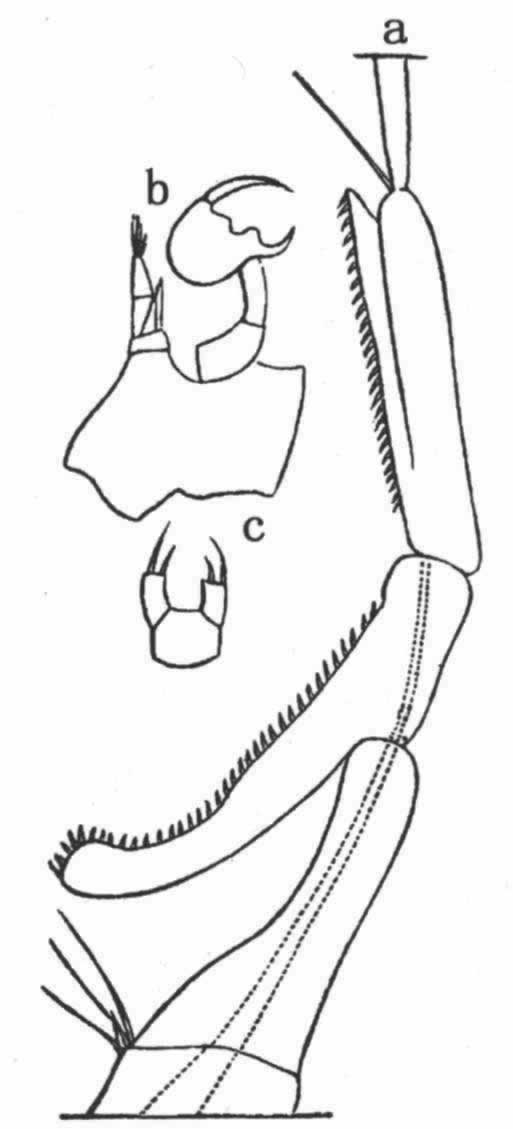
图 4 达氏角水蚤(Labidocera Darwinii)(引自卢伯克原著)
a.雄者右前触角形成抱握器官的部分;b.雄者的一对后胸肢;c.雌者的一对后胸肢。
高等甲壳动物的前肢发育成钳爪或螯,而雄者的这等器官一般比雌者的大,——按照斯彭斯·巴特先生的材料,雄黄道蟹(Cancer pagurus)因为螯很大,其市价要比雌蟹贵五倍。许多物种躯体两侧的螯大小不相等,正如巴特先生告诉我的,右侧的螯一般都是最大的,虽然并非一律如此。这种大小不相等的程度也常常是在雄者比在雌者为大。雄者两只螯的构造往往也有差异(图5,6和7),较小的那只螯同雌者的相类似。它们躯体相对两侧的螯大小不相等以及雄者两侧的螯大小不相等的程度大于雌者会带来什么利益;还有,当两侧的螯大小相等时,雄者的螯为什么又往往大于雌者的,其原因都还弄不清楚。我听巴特先生说,有时它们的螯是如此之大而且如此之长,以致不能用它们取食送至口际。某种淡水雄斑节虾(长臂虾属,Palaemon)的右肢实际上比整个身体还要长。 (115) 这条大型的肢加上它的螯将有助于它同其竞争对手进行战斗;但是,雌者躯体相对两侧的不相等并不是由于这同样的原因。根据米尔恩·爱德华兹所引用的一段叙述, (116) 在招潮蟹(Gelasimus)中,雄者和雌者同穴而居,这阐明它们是成双成对的;雄者用一只非常发达的螯把洞口堵住,因此它在这里是间接用做防御手段的。然而其主要用途大概还是在于抓住和保卫雌者。有些事例,如钩虾(Gammarus),据知就是如此。雄寄居蟹或武士蟹(寄居蟹 (117) ,Pagurus)一连几个星期携带着雌蟹所居住的壳转来转去。 (118) 然而,巴特先生告诉我说,普通滨蟹(Carcinus maenas)在雌蟹刚一脱掉硬壳之后,雌雄就直接交合,雌蟹脱壳后是那样地娇嫩,这时如果被雄蟹强有力的双钳夹住就会受到伤害;但是因为雄蟹在雌蟹脱壳之前就捉住了它并把它带来带去,所以在雌蟹脱壳后再抓住它就不会造成损伤了。
图 5 美人虾属(Callianassa)身体的前部,示明雄者右侧和左侧的螯大小不相等以及构造的不同
(引自米尔恩·爱德华兹原著)注意:绘图人把图画颠倒了,误把左螯画成最大的。

图 6 一种雄跳钩虾(Orchestia Tucuratinga)
(引自弗里茨·米勒原著)
图 7 同图 6,雌者的第二肢
弗里茨·米勒说,Melita的某些物种由于雌者的“倒数第二对足的基节片长成为钩状突起,以便雄者用第一对前肢把它们抓牢”,因而同所有其他端足类(amphipods)都有所区别。这种钩状突起的发育大概由于雌者在生殖行为中可以最牢固地被雄者抓紧,并留下最大数量的后代。另一个巴西的端足类——(达氏跳钩虾,Orchestia Darwinii,图8)呈二态现象;同异足水虱属的情况相似;因为它们有两种雄性类型,其区别在于螯的构造。 (119) 由于无论用哪一支螯肯定都完全可以把雌者抱握住——因为现在这两支螯都用于这个目的——所以这两种雄性类型大概起源于有些个体发生了这样变异,而另一些个体则发生了那样变异;这两种类型由于它们不同形状的器官都曾经产生了某种特殊的而又接近相等的利益。
现在尚未发现雄性甲壳动物为了占有雌者而互相战斗,但这种情况可能是存在的;因为,对大多数动物来说,当雄者大于雌者时,雄者的较大体型似乎是靠了其祖先同其他雄者经历了许多世代的战斗之故。在大多数的“目”中,尤其是在最高等的“目”、即短尾类(Brachyura) (120) 中,雄者都大于雌者;然而雌雄二者遵循不同生活习性的寄生性的属,以及大多数的切甲类(Entomostraca) (121) 都是例外。许多甲壳动物的螯都是十分适于战斗的武器。例如,巴特先生的儿子曾见到一只梭子蟹(Portunus puber)同一只滨蟹进行战斗,后者很快就被打得背朝下,而且每条肢都从躯体上被撕裂下来。有一个具有巨螯的物种叫巴西招潮蟹(Brazilian Gelasimus),当弗里茨·米勒把它们的若干只雄者放入一个玻璃容器时,它们就互相撕裂和残杀。巴特先生曾把一只大型的雄滨蟹放入一盆水中,其中已有一只雌滨蟹同一只较小的雄滨蟹交配,但后者很快就被撵走了。巴特先生接着说,“如果它们战斗过,那么这个胜利是一种不流血的胜利,因为我没见到损伤”。这位博物学家把一只雄海岸钩虾(Gammarus marinus,在英国的海岸上经常可以见到),同它的雌性伴侣分开,它们二者原是与同一个物种的许多个体放在同一个容器里的。当雌者这样被分开之后,很快就同其他个体混合在一起了。经过一段时间后,又把原来那只雄者放回容器里,它在四周游了一会儿后,就向虾群中猛冲进去,没有经过任何战斗,一下子就把其原配带走了。这个事实阐明了,在动物等级上属于低等一个“目”——异足类的雄者同雌者是彼此认识的,而且是互相依恋的。
图 8 达氏跳钩虾(Orchestia Darwinii)(根据弗里茨·米勒原著)
图示两种雄性类型的螯的不同构造。
甲壳动物的心理能力大概比初见时所表现的为高。任何人如果试图去捉一只热带海滨常见的海岸蟹,就会看到它们是多么谨慎和警惕。在珊瑚岛上发现有一种大型的蟹(椰子蟹,Birgus latro),它们能从椰实剔出纤维,在一个深洞底部铺成一个厚床。它以掉下的椰实为食,自外壳逐层撕去其纤维,而且总是从椰子上有三个像眼睛那样凹痕的那一端开始。然后它用其沉重的前螯敲打,打开其中的一个凹眼,再把它翻过来,用其狭窄的后螯取出里面含有丰富胚乳的果心。不过这些动作大概是本能的,因为在进行这些动作时幼蟹同老蟹都完成得一样好。然而下述情况就几乎不能认为也是如此:一位可信赖的加德纳(Gardner)先生 (122) ,当他注视到一只海岸蟹(招潮蟹属)在做巢穴时,曾向洞穴扔了若干贝壳。一只贝壳滚进了洞里,另三只贝壳掉在离洞口几英寸的地方。五分钟左右,这只蟹把掉进洞里的那只贝壳弄了出来,放到离洞口一英尺的外面;然后它又见到掉在附近的那三只贝壳,并且显然想到它们会同样滚进洞里去的,于是又把它们带到第一只贝壳所在的地方。我想这种行动同人类借助于理性的行动之间是很难加以区别的。
巴特先生不知道有任何十分显著的事例表明我们英国的甲壳动物的雌雄颜色有什么差异,而高等动物的雌雄二者在这方面的差异是很常见的。然而在某些场合中,雌雄二者的色调稍有不同,但巴特先生认为这无非是由于它们的不同生活习性所致,譬如雄者游动性较强,这样就受光较多。鲍尔(Power)博士曾试图从颜色来区别产于毛里求斯的一些物种的雌雄性别,但除了虾蛄属(Squilla)的一个物种之外都失败了,这个物种大概就是针形虾蛄(S.styli fera),其雄者被描述为“具有美丽的天蓝色”,而且有一些樱红色的附器,但雌者的外壳则布满模糊的褐色和灰色斑点,“其四周的红色远不如雄者的鲜艳”。 (123) 从这个例子,我们可以猜想到性选择的作用。根据M.伯特(Bert)对水蚤属(Daphnia) (124) 的观察,当把它放进一个通过棱镜的光线所照射的容器里时,我们有理由相信甚至最低等的甲壳动物也能辨别颜色。叶剑水蚤属(Saphirina,切甲类的一个海产属)的雄者具有许多微型盾状体或类细胞体,它们表现有变化不定的美丽颜色;这些颜色是雌者所没有的,有一个物种的雌雄二者都没有这样颜色。 (125) 然而要断定这些奇妙的器官就是用来吸引雌者就未免过于轻率了。弗里茨·米勒对我说,招潮属的一个巴西物种的雌者通体几乎都是一致的灰褐色。雄者头胸部的后部是全白色的,前部是深绿色的,并逐渐变为暗褐色;值得注意的是,这些颜色在几分钟内就有改变的倾向——由白色变成暗灰色甚至是黑色,而绿色也“失去了其大部光泽”。尤其值得注意的是,雄者要到成熟时才获得其鲜明的色彩。它们的数量看来比雌者多得多;它们的螯也大于雌者。在这个属的某些物种中,也可能在它的所有物种中,雌雄二者都是成双成对地居住在同一个洞穴内。正如我们已经看到的,它们也是高度聪明的动物。根据这种种考察,看来这个物种的雄者大概为了吸引或刺激雌者而变得装饰华丽了。
上面刚刚讲过雄招潮蟹要到成熟后,接近准备繁殖的时候才获得其鲜明的色彩。关于雌雄二者之间许多构造上的显著差异,上述一点似乎是这全纲的一个普遍规律。以后我们将会看到同一法则且通用于脊椎动物这一大的亚界。而且在所有场合中,通过性选择所获得的性状都是区别显著分明的。弗里茨·米勒 (126) 提出一些有关这个法则的显著事例,例如雄跳钩虾(Orchestia)要到接近完全成长时才获得巨大的抱握器,其构造和雌者的大不相同,而当雄者幼小时,其抱握器则同雌者的相似。
蛛形纲(Arachnida)(蜘蛛类)
其雌雄二者的颜色一般没有重大区别,但雄者往往比雌者色暗,在布莱克瓦尔(Blackwall)先生的巨著 (127) 中可以看到这一点。然而有些物种的差异却是显著的,例如,雌绿色遁蛛(Sparassus smaragdulus)呈暗绿色,而成年雄蛛的腹部则呈鲜黄色,并具三道浓艳的红色纵条纹。蟹蛛属(Thomisus)某些物种的雌雄二者彼此密切类似,但另外一些物种的雌雄二者则很不相同,许多别的属也有近似的情况。究竟雌雄二者哪一方同该物种所隶属的那个属的正常色彩相差最大,往往很难说。但布莱克瓦尔先生认为,按照一般的规律,还是雄者如此。卡内斯垂尼述说 (128) ,在某些属中,雄者特别容易被识别,而识别雌者就非常困难了。布莱克瓦尔先生告诉我说,雌雄二者在幼小时通常是彼此类似的,在它们成熟之前的连续几次蜕皮期间二者的颜色就往往发生了重大变化。在另外一些场合中,好像只有雄者的颜色发生变化。因此上述具有鲜明色彩的雄遁蛛最初同雌者相类似,只有当它接近成熟时才获得其特有的色调。蜘蛛类具有敏锐的感觉并表现有很大的智力,众所周知,雌蜘蛛对它们的卵常常表现了最强烈的感情,它们用丝网把卵封包起来,随身携带。雄蜘蛛热切地寻求雌者,卡内斯垂尼以及其他人士还见到过雄蜘蛛为了占有雌者而进行战斗。这位作者还说,他曾观察过将近20个物种的雌雄蜘蛛的交配,他肯定地断言,雌蜘蛛拒绝有些雄蜘蛛的求爱,张开了上颚吓唬它们,经过长时间的犹豫之后,最后才接受了它所挑中的一只。根据这几种考察,我们可以多少有些把握地承认,某些物种雌雄二者之间在颜色方面的显著差异乃是性选择的结果。虽然关于这一点我们还没有掌握最好的证据——雄者以其装饰物进行夸示。从有些物种的雄者在颜色上的极度变异性来看,例如条纹球腹蛛(Theridion lineatum),其雄者的这些特征似乎至今还不十分稳定。卡内斯垂尼根据某些物种的雄者在颚的大小和长短上,呈现了互相区别的两种类型这一事实,也作出了同样的结论,这一点使我们想起了上述有关甲壳动物二态性的事例。
雄蜘蛛一般要比雌者小得多,有时竟小到异常的程度, (129) 迫于此,雄者为求偶而向雌者接近时,必须极端小心,因为雌者的羞怯往往会引起危险的攻击。德吉尔(De Geer)见过一只雄蜘蛛“浸沉在准备求爱之中时,被其所注意的对象捉住,包入她的蛛网,然后被吃掉,”他接着说,“这一景象使他充满了恐怖和愤慨”。 (130) 坎勃瑞季牧师 (131) 关于络新妇(Nephila)雄者的极端小型做过如下说明。“万松(M.Vinson)关于小型雄蜘蛛逃避雌者的凶猛攻击,做过生动记载,雄蜘蛛采取同雌者捉迷藏的办法,沿着后者的巨肢,越过后者的躯体,在后者四周滑来滑去。在这样一场追逐中显然最小的雄蜘蛛逃脱的机会最多,而大一点的雄蜘蛛很早就会成为牺牲品;因此一种小的雄性类型就会渐渐受到选择,直至最后缩到最小的可能程度,以适合实行其生殖机能——事实上大概就是我们现在所看到的那样大小,这就是说,它们小得就像雌者身上的一种寄生物,不会引起雌者的注意,或者因为它们太小、太敏捷,以致雌者非常难于捉住它们。”
韦斯特林(Westring)有过一个有趣的发现:即球腹蛛属某些物种的雄者 (132) 具有摩擦发音的能力,而雌者却是哑子。它的发音器的构成是由腹底的锯齿状隆起同坚硬的后胸部相摩擦,但在雌者找不到这种构造的任何痕迹。值得注意的是,有几位作者,其中包括著名的蜘蛛学家瓦尔克纳(Walckenaer),曾宣称蜘蛛类受音乐的吸引。 (133) 根据下一章所描述的直翅目(Orthoptera)和同翅目(Homoptera)昆虫来类推,我们几乎可以肯定这种摩擦发音是用来召唤或刺激雌者,韦斯特林也这样认为。在动物界的等级中向下追溯,关于为了这个目的而发出音响的,这是我所知道的最初的一个事例。 (134)
多足纲
这个纲的两个“目”,无论马陆类(millipedes)或蜈蚣类(centipedes),都没有这等雌雄差异的任何十分显著的事例值得我们更特别关心。然而,有一种球马陆(Glomeris limbata),另外也许还有少数物种,其雄者和雌者的颜色稍有不同,但这种马陆都是一些高度容易变异的物种。关于倍足亚(Diplopoda)纲的雄者,在其躯体某一前节或后节上着生的一对腿变成了可以抱握的钩状物,作为抱持雌者之用。马陆属(Iulus)某些物种的雄者的跗节具有膜质吸盘,其用途也是一样。我们讨论到昆虫类时,将会看到十分异常的情况是,在石蜈蚣(Lithobius)属中,正是雌者在其躯体末端具有抱握的附器以抱持雄者。 (135)
第十章 昆虫类的第二性征
雄虫用以捉住雌虫的各种构造——雌雄之间含义不明的差异——雌雄之间在大小上的差异——缨尾目——双翅目——半翅目——同翅亚目,只有雄虫才具有的音乐能力——直翅目,雄虫的音乐器官,构造的巨大变化;好斗性;色彩——脉翅目,雌雄颜色的差异——膜翅目,好斗性和色彩——鞘翅目,色彩;具有明显作为装饰之用的巨角;战斗;雌雄双方一般都有的摩擦发音器
在庞大的昆虫纲中,雌雄的差异有时表现在运动器官上,但往往是表现在感觉器官上,如许多物种的雄虫所具有的栉状触角和美丽的羽状触角即是。蜉蝣类(Ephemerae)的一种叫做Chlo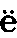 on的,其雄虫具有巨大的柱眼,而雌虫则没有, (136) 某些昆虫的雌者没有单眼,蚁蜂科(Mutillidae)就是如此,同时它们还没有翅。然而我们所关心的主要是使某只雄者或在战斗中或在求偶中能凭其体力,好斗性,装饰,或音乐去战胜其他雄者的那些构造。因此,雄者用以抓住雌者的无数装置,可能简略地一笔带过,但腹端的复杂构造除外,这恐怕是要列为初级器官的, (137) 正如沃尔什先生所说的,“为了使雄者能够牢固地抓住雌者这个表面上毫不重要的目的,大自然创造了何等众多的不同器官,实足令人惊异不止”。 (138) 昆虫的上颚或颚有时就用于这种目的。例如,具角鱼蛉(Corydalis cornutus,一种脉翅目昆虫,同蜻蜓等有某种程度的亲缘关系)的雄者具有大而弯曲的颚,比雌者的颚长达数倍;它们是平滑的,而不是锯齿形的,所以雄者这样抓住雌者时就不致使她受到伤害。 (139) 北美洲有一种大锹甲虫(Lucanus elaphus),其雄者的颚比雌者的大得多,也用于同样的目的,但大概也用于战斗。有一种蠮螉(Ammophila),其雌雄二者的颚密切相似,但用于大不相同的目的:如韦斯特伍德教授所观察的,雄蜂“非常热情,用其镰刀状的颚绕住配偶的颈部把后者抓住” (140) ;雌蜂则用这种器官在沙坝上打洞筑巢。
on的,其雄虫具有巨大的柱眼,而雌虫则没有, (136) 某些昆虫的雌者没有单眼,蚁蜂科(Mutillidae)就是如此,同时它们还没有翅。然而我们所关心的主要是使某只雄者或在战斗中或在求偶中能凭其体力,好斗性,装饰,或音乐去战胜其他雄者的那些构造。因此,雄者用以抓住雌者的无数装置,可能简略地一笔带过,但腹端的复杂构造除外,这恐怕是要列为初级器官的, (137) 正如沃尔什先生所说的,“为了使雄者能够牢固地抓住雌者这个表面上毫不重要的目的,大自然创造了何等众多的不同器官,实足令人惊异不止”。 (138) 昆虫的上颚或颚有时就用于这种目的。例如,具角鱼蛉(Corydalis cornutus,一种脉翅目昆虫,同蜻蜓等有某种程度的亲缘关系)的雄者具有大而弯曲的颚,比雌者的颚长达数倍;它们是平滑的,而不是锯齿形的,所以雄者这样抓住雌者时就不致使她受到伤害。 (139) 北美洲有一种大锹甲虫(Lucanus elaphus),其雄者的颚比雌者的大得多,也用于同样的目的,但大概也用于战斗。有一种蠮螉(Ammophila),其雌雄二者的颚密切相似,但用于大不相同的目的:如韦斯特伍德教授所观察的,雄蜂“非常热情,用其镰刀状的颚绕住配偶的颈部把后者抓住” (140) ;雌蜂则用这种器官在沙坝上打洞筑巢。
许多雄甲虫前肢跗节膨大或具有宽的毛垫,在水生甲虫的许多属中,它们都具有扁圆的吸盘以便雄者能吸附在雌者的滑湿躯体上。有一个更加异常得多的情况,即有些水生甲虫(龙虱属Dyticus)的雌者具有刻着深槽的鞘翅,而条纹龙虱(Acilius sulcatus)雌者的鞘翅上则披着厚厚一层毛,借此以帮助雄者抱持雌者。另外有些水生甲虫(Hydroporus)的雌者为了同一目的,具有刻点的鞘翅。 (141) 细腰蜂雄者(图9)的胫节膨大而成宽阔的角质板,其上布满了微小的膜质点,使它呈现粗筛状的独特外形。 (142) 霉蛰属(Penthe,甲虫的一属)的雄者“显然因同一目的”,其触角中部的几节膨大,而且膨大部分的下表面有毛垫,同步行虫科(Carabidae)跗节的膨大部分完全一样。雄蜻蜓“尾巴尖端的跗器变成几乎数不清的种种奇形怪状,使它们能够用以抱握雌蜻蜓的颈部”。最后,许多雄性昆虫的肢都具有特殊的刺、节或距;或者整个肢弯成弓状或变粗,这是一种性的特征,但决非永远如此;或者一对肢变长了,或者三对肢都变长了,有时长到过分的程度。 (143)
在所有昆虫的目中,许多物种的雌雄二者都表现有含义不明的差异。有一个奇妙的例子是关于一种甲虫的(图10),其雄者的左上颚变得非常之大,因而口器大大歪斜。另一种步行甲虫,阔颚虫(Eurygnathus), (144) 其雌者的头比雄者的宽得多而且大得多,虽然在程度上有所不同,这是沃拉斯顿(Wollaston)先生所知道的独一无二的事例。这种含义不明的事例不胜枚举。鳞翅目中充满了这类事例。其中最特别的一个是,某些雄蝴蝶前肢多少有些萎缩,其胫节和跗节缩小成仅是痕迹的小瘤。雌雄二者常常在翅的脉序上有所不同, (145) 有时其轮廓也相当不同,巴特勒(A.Butler)先生在大英博物馆给我看的Aricoris epitus就是如此。某些南美洲的雄蝴蝶的翅在边缘上有毛簇,后一对翅的中域有角质赘疣。 (146) 若干英国蝴蝶,如翁弗尔(Wonfor)先生所阐明的,只有雄者才部分地披有特殊鳞片。
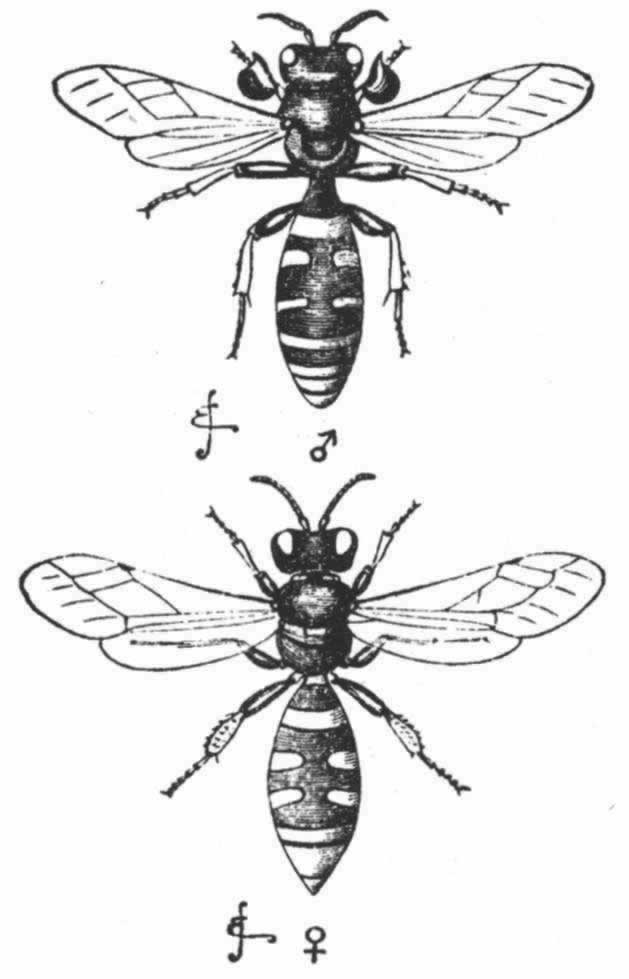
图 9 细腰蜂(Crabro cribrarius)
上图是雄蜂;下图是雌蜂。
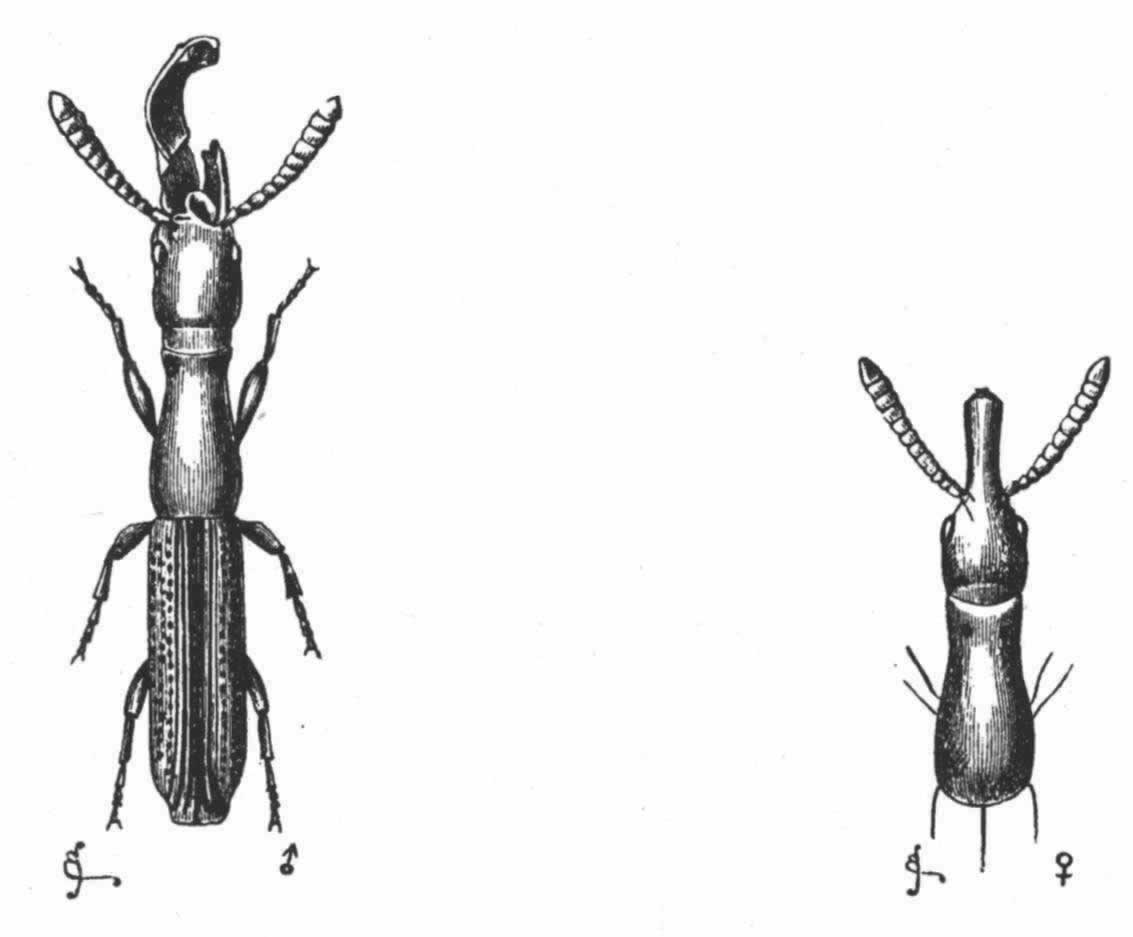
图 10 潜叶吉丁(Taphroderes distortus)(放大)
左图是雄虫;右图是雌虫。
雌性萤火虫发光的用途,引起了许多讨论。雄虫发光很弱,其幼虫以至卵的情况也一样。有些作者设想萤火虫的光是用以吓走敌人,另外有些作者则设想是用以引导雄虫来找雌虫的。贝尔特先生好像终于解决了这个难题: (147) 他发现所有他用做试验的萤科(Lampyridae)昆虫,都是食虫的哺乳类和鸟类所高度厌恶的。因此,这同后面还要加以说明的贝茨先生的观点相符合,他认为许多昆虫密切模拟萤科是为了使食虫动物弄错,而这样逃脱毁灭。他进一步相信发光的物种有个好处,即立刻可被认出它是不可口的。这个解释大概可引申到弹尾目昆虫,其雌雄二者都是高度发光的。至于雌萤火虫的翅为什么不发育,还弄不清楚;但雌虫现在的形态同一种幼虫密切类似,而幼虫是许多动物大量捕食的对象,因此我们就可理解为什么雌虫会比雄虫发出明亮得多的光而且更显眼得多,同样地,为什么幼虫本身也会发光。
雌雄之间在大小上的差异
所有种类的昆虫普遍都是雄者小于雌者,这种差异甚至在幼虫状态中已可察觉。家蚕(Bombyx mori)的雄性茧和雌性茧之间的差异是那么显著,以致在法国是用一种特殊的称重方法将二者分离开来。 (148) 在动物界的低等纲中,雌者体型大于雄者一般似乎是由于前者要育成大量的卵,这在某种程度上也适用于昆虫类。但华莱士博士提出一个可能性大得多的解释。他仔细观察了臭椿蚕和天蚕幼虫的发育,特别是观察了用异常食物饲养的第二造短小幼虫的发育,发现“蚕蛾个体越细小,其变态所需的时间也成比例地越长;正是由于这个原因,雌蛾因为要产生大量的卵,所以大于而且重于雄蛾,而体型较小、易于成熟的雄蛾将先于雌蛾孵化”。 (149) 那么,由于大多数昆虫都是短命的,而且由于它们处于许多危险之中,因此雌蛾如能尽早受精,显然对它是有利的。如果大批雄蛾先行成熟并随时等候雌蛾的出现,就可达到上述目的;正如华莱士先生所指出的 (150) ,这当然也是自然选择带来的结果;因为较小的雄虫先成熟,就会繁殖出大量继承其父本短小体型的后代,而体型较大的雄虫因成熟较晚就要留下数量较少的后代。
然而,雄虫小于雌虫这个规律也有例外,其中有些是容易理解的。在占有雌虫的斗争中,体大和力强对雄虫可能是一种有利条件;在这样事例中,如锹甲虫(Lucanus)雄虫则大于雌虫,另外还有些甲虫,据知彼此并不为了占有雌虫进行战斗,而其雄虫在大小上也超过雌虫;这个事实的意义还不清楚;不过在某些这样场合中,例如,关于巨大的独角仙(Dynastes) (151) 和分枝独角仙(Megasoma) (152) ,我们至少能够知道,雄者没有必要为了先于雌虫成熟而小于雌虫,因为这等甲虫并不是短命的,故有充分时间保证雌雄的交配。另外,雄蜻蜓蜻科(Libellulidae)从不小于雌者,有时则明显地大于雌者; (153) 正如麦克拉克伦先生所相信的,雄蜻蜓要经过一周或二周并呈现出它们特殊的雄性色彩后才会同雌蜻蜓普遍交配。但最奇妙的是关于具有螫刺的膜翅目(Hymenoptera)昆虫的例子,它阐明了像雌雄二者之间的体型差异这样一种微小的性状却受着多么复杂而容易被忽略的关系所支配;因为史密斯先生告诉我说,几乎在整个这一巨大类群中,按照一般规律,雄虫都小于雌虫,而且其羽化先于雌虫一周左右;但在蜜蜂类中,蜜蜂(Apis mellifica)、长袖切叶蜂(Anthidium manicatum)和毛花蜂(Anthophora acervorum)的雄者,以及在掘土蜂类(Fossores)中,艳蚁蜂(Methoca ichneumonides)的雄者都大于雌者。对这种异常现象的解释是,实行一种飞行交配对这些物种是绝对必要的,而雄者为了在空中携带雌者就需要巨大的体力和体型。在这里,虫体的增大是同体型大小和发育期之间的普通关系相违背的,因为雄者虽比雌者大,但比雌者先羽化。
我们现在对几个“目”的昆虫再检查一下,从中选用一些同我们有特别关系的事实。关于鳞翅目(蝴蝶类和蛾类),将另立一章进行讨论。
缨尾目(Thysanura)
这个“目”的体制低等,其成员都是无翅的、颜色暗淡的、体型微小的昆虫,具有丑陋的、几乎是畸形的头和躯体。它们的雌雄二者无区别,但使人感到兴趣的是,它们阐明了即使在动物等级的低下阶段,其雄者也孜孜不倦地向雌者求爱。卢伯克爵士说:“看到这些小动物(黄圆跳虫,Smynthurus luteus)在一起卖弄风骚很是有趣。比雌虫小得多的雄虫绕着雌虫跑,彼此抵撞,迎面而立,退退进进,活像两只相戏的羊羔。然后雌虫假装跑开,雄虫装着一种愤怒的怪模样在后面追,赶到雌虫前面之后,又一次迎面而立;然后雌虫羞怯地转身避开,但雄虫比雌的跑得更快而且更活跃,一溜烟地前后左右追随,而且似乎用其触角鞭打雌的,过了一会它们又迎面对立,用触角相戏,互相之间的一切似乎全都解决了。” (154)
双翅目(Diptera)(蝇类)
雌雄之间的颜色差别很小。据沃克(F.Walker)先生所知,雌雄差异最大者为毛蝇属(Bibio),其雄者略带黑色或为全黑色,雌者是暗褐橙色。华莱士先生 (155) 在新几内亚发现的角蝇属(Elaphomyia)是高度引人注意的,因雄者有角,而雌者全无。角从眼的下方生出,同雄鹿的角奇妙地相似,不是呈叉状就是呈掌状。其中有一个物种的角与躯体的长度相等。大概有人认为这等角是适于战斗的,但有一个物种的角呈美丽的淡红色,黑色镶边,并有一道淡色中央条纹,因为这等昆虫的整个外貌都很优雅,所以更加可能的是,这等角是用做装饰的。有些双翅目的雄蝇相互争斗是肯定的,因为韦斯特伍德教授 (156) 好几次见过大蚊属(Tipulae)就有这种现象。其他双翅目的雄者显然试图以它们奏出的音乐赢得雌者的欢心:米勒 (157) 几次注意到一种蜂蝇(Eristalis)的两只雄者在追求一只雌者;雄者在雌者上面盘旋,在其周围飞来飞去,同时发出很响的嗡嗡声。蚋科(gnat)和蚊科(Culicidae)似乎也靠发出嗡嗡声互相吸引;迈耶教授最近已证实在雌虫发出的声音范围之内,雄虫触角上的毛振动得同音叉的音调相符。长毛的振动同低音调共鸣,短毛的振动同高音调共鸣。兰多依斯(Landois)也宣称他曾用某种特殊音调反复地招来了一整群蚋。还可以补充一点,双翅目的心理官能大概高于其他大多数昆虫,这同它们的高度发达的神经系统是符合的。 (158)
半翅目(Hemiptera)(蝽象类)
道格拉斯(J.W.Douglas)先生对英国物种做过特别研究,他热心向我提供了一份有关这等物种雌雄差异的报告。有些物种的雄者有翅,而雌者无翅;两者在躯体、鞘翅、触角和跗节的形态上都有差异;但由于这些差异的意义不明,故略而不谈。雌者一般都比雄者体大而强壮。英国的物种以及道格拉斯先生所知道的外来物种,其雌雄二者在颜色上通常并无多大差异;但是,约有6个英国物种的雄者比雌者的颜色暗得多,另外还有4个物种,却是雌者的颜色比雄者的暗。有些物种的雌雄二者都有美丽的颜色;由于这些昆虫散发一种非常令人作呕的气味,所以其显著颜色也许就是对食虫动物发出的一种不好吃的信号。在某些少数场合中,它们的颜色似乎直接就是保护性的。例如,霍夫曼(Hoffmann)教授告诉我说,有一种淡红色和绿色的小型物种经常群集在菩提树上,他简直不能把它们同树干上的芽区别开来。
猎蝽科(Reduvidae)的某些物种能摩擦发音,以善鸣黝蠄(Pirates stridulus)的例子而言,据说是由于它们的颈在前胸腔内运动而发音。 (159) 按照韦斯特林的见解,盛装猎蝽(Reduvius personatus)也会摩擦发音。对于非社会性的昆虫来说,除非发音是作为一种性的呼唤,否则发音器官就没有任何用处了,倘不如此,我就没有理由设想这种摩擦发音乃一种性征。
同翅目(Homoptera)
凡是在热带丛林中漫游过的人一定都会对雄蝉发出的喧噪声感到惊奇。雌蝉却默不作声,正如希腊诗人季纳卡斯(Xenarchus)说的,“乐哉蝉之生活,有妻皆默女”。当“贝格尔”号在离巴西海岸四分之一海里的地方抛锚泊船时,在甲板上就可清楚听到这样的噪音;汉考克船长说,远在一英里以外的地方就可听到这种噪音。希腊人过去把它们养在笼里,现在中国人还这样做,为的是欣赏它们的歌唱,所以有些人一定感到这是悦耳的声音。 (160) 蝉科(Cicadidae)通常是在白天歌唱,而樗鸡科(Fulgoridae)好像是夜间的歌手。按照兰多依斯(Landois)的见解, (161) 这声音是由气门唇边的振动而产生的,气门唇边的振动又是由气管发出的一股气流引起的,但对这个观点最近有所争论。鲍威尔博士似乎证明了这种声音是由一块膜的振动而产生的, (162) 而这块膜则是由一块特别肌肉牵动起来的。在唧唧鸣叫的活昆虫身上,可以见到这片膜在振动;在死昆虫身上,若以针尖拨动那块稍微变干和变硬的肌肉,也可听到其固有的声响。雌虫身上也有这整个的复杂音乐器官,但远不如雄虫的发达,且绝不用以发声。
关于这种音乐的目的,哈特曼(Hartman)博士提到美国的周期蝉(即17年蝉Cicada septemdecim)时说道 (163) :“现在(1851年6月6日和7日)四面八方都可听到鼓噪声。我相信这是雄者对雌者发出的召唤。我站在高与头齐的满布嫩芽的栗树丛中,成千上百的雄蝉在我的四周,我看到雌蝉飞来环绕着鼓噪的雄蝉周围。”他接着说:“在我的花园里有一株矮生梨树,这个季节(1868年8月)在它上面产生了50只左右梨蝉(Cic.pruinosa)的幼虫;我好几次注意到雌蝉落在一只正发出响亮声调的雄蝉附近。”弗里茨·米勒从巴西南部写信告我说,他常听到属于一个物种的两三只雄蝉用特别响亮的声调进行音乐比赛:一只刚唱完,另一只马上开始,然后又一只接下去。由于雄蝉之间有那么多的竞争者,因而雌蝉大概不仅是根据音响寻找雄蝉,而且也像鸟类的母鸟一样,会被具有最动听的声音的雄蝉所刺激与诱惑。
关于同翅目昆虫雌雄二者之间的装饰差异,我还没听说过任何十分显著的事例。道格拉斯先生告诉我说,有三个英国的物种,其雄者是黑色的或具有黑色带斑,而雌者则颜色浅淡或黯然无光。
直翅目(Orthoptera)(蟋蟀和蝗虫)
本目中有三个能跳跃的科,其中雄者都以其音乐能力著称,这三个科是:蟋蟀科(Achetidae)、螽斯科(Locustidae)和蝗科(Acridiidae)。有些种螽斯摩擦发音如此响亮,以致夜间在一英里以外都可听到; (164) 某些物种的叫声即使在人听起来也很悦耳,因此亚马孙河一带的印第安人把它们养在柳条笼子里。所有的观察家都一致认为这种叫声不是用来召唤就是刺激不会发音的雌者的。关于俄国的迁移性蝗虫, (165) 克尔特(K rte)举出过一个有关雌者选择雄者的有趣例子。飞蝗(Pachytylus migratorius)的雄者当同雌者交配时,如有另一只雄者走近,它就会因愤怒或嫉妒而唧唧叫起来。家蟋蟀在夜间受到惊扰时就会用它的叫声来警告其伙伴。 (166) 据记载, (167) 北美产的穴居扁叶
rte)举出过一个有关雌者选择雄者的有趣例子。飞蝗(Pachytylus migratorius)的雄者当同雌者交配时,如有另一只雄者走近,它就会因愤怒或嫉妒而唧唧叫起来。家蟋蟀在夜间受到惊扰时就会用它的叫声来警告其伙伴。 (166) 据记载, (167) 北美产的穴居扁叶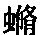 (Platyphyllum concavum,螽斯科的一种)登上树木的顶枝,一到傍晚就开始“发出嘈杂的喧叫,竞争者的叫声也同时在邻近的树上呼应,整个小树林回响着“凯提—底得—施—底得”的叫声,彻夜不休”。贝茨先生谈到欧洲田蟋蟀(蟋蟀科的一种)时说道,“一到傍晚就可看到雄蟋蟀呆在洞口唧唧地叫,一直叫到一只雌蟋蟀到来时,于是叫声就由高音转为低音,同时这个成功的演奏家用其触角爱抚着它所赢得的配偶”。 (168) 斯卡德(Scudder)博士用一支羽茎在纸夹上摩擦发音就能刺激一只这种昆虫发出叫声来呼应。 (169) 冯·西博尔德(Von Siebold)已经发现雄虫和雌虫的显著的听觉器官位于前肢。 (170)
(Platyphyllum concavum,螽斯科的一种)登上树木的顶枝,一到傍晚就开始“发出嘈杂的喧叫,竞争者的叫声也同时在邻近的树上呼应,整个小树林回响着“凯提—底得—施—底得”的叫声,彻夜不休”。贝茨先生谈到欧洲田蟋蟀(蟋蟀科的一种)时说道,“一到傍晚就可看到雄蟋蟀呆在洞口唧唧地叫,一直叫到一只雌蟋蟀到来时,于是叫声就由高音转为低音,同时这个成功的演奏家用其触角爱抚着它所赢得的配偶”。 (168) 斯卡德(Scudder)博士用一支羽茎在纸夹上摩擦发音就能刺激一只这种昆虫发出叫声来呼应。 (169) 冯·西博尔德(Von Siebold)已经发现雄虫和雌虫的显著的听觉器官位于前肢。 (170)
这三个科的发音方法各不相同。蟋蟀科雄者的两个鞘翅具有相同的器官,田蟋蟀(Gryllus campestris,图11)的这类器官,像兰多依斯所描述的, (171) 是由131至138个锐利的、横向的脊或齿(st)构成的,这等脊或齿位于鞘翅脉之一的下表面。这种具齿的翅脉同位于相对一翅上表面的一道突出而平滑的硬翅脉(r)迅速摩擦。先是一翅向另一翅擦过去,然后又是逆向地擦过来。两翅同时稍微抬高,以便提高共鸣的效果。在某些物种中,雄者的鞘翅基部具有一片云母状的板。 (172) 图12表明蟋蟀属另一个物种叫家蟋蟀(G.domesticus)的翅脉下表面的齿。格鲁勃博士曾阐明这等齿是在选择作用的帮助下由覆盖于翅和躯体之上的小鳞片和毛形成的,关于鞘翅目(Coleoptera)的齿,我得出了相同结论。但格鲁勃进一步阐明这等齿的发展, (173) 部分地是直接由于一翅在另一翅上摩擦所产生的刺激。
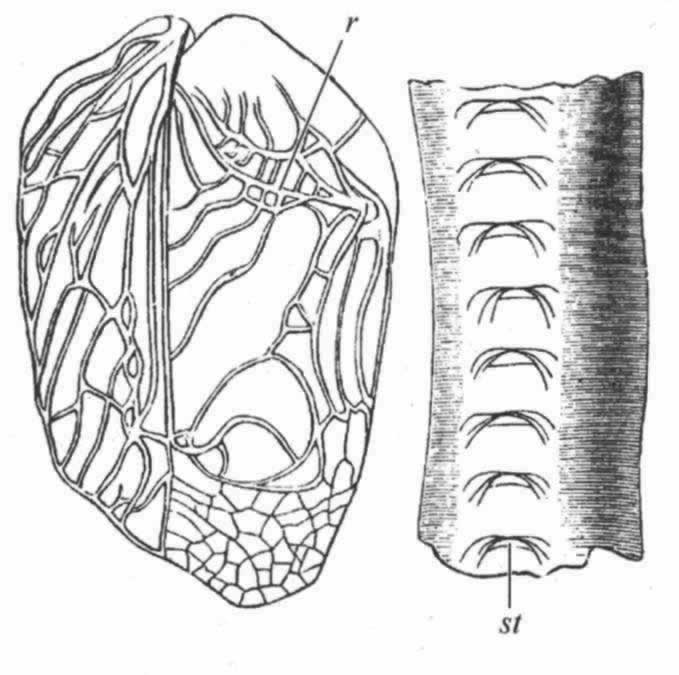
图 11 田蟋蟀(Gryllus campestris)(引自兰多依斯)
右图是放大很多的翅脉底面的一部分,st表示上面的齿。
左图是翅鞘背面图,上面有翅脉的光滑突起r,用它与齿(st)交互摩擦。
图 12 家蟋蟀(Gryllus domesticus)
翅脉上的齿(引自兰多依斯)
在螽斯科中,相对的两个鞘翅彼此在构造上有所不同(图13),其摩擦动作不同于蟋蟀科,不能逆向进行。左翅的作用有如提琴的弓,位于作为提琴的右翅之上。左翅底面的翅脉之一具有细齿,在相对的右翅上面的具有突起的翅脉上擦过。在我看来,我们英国的普通绿螽斯(Phasgonura viridissima)的锯齿状翅脉似乎是与相对另一翅的圆形后角相摩擦后面这张翅的边缘较厚,褐色,很锐利。在右翅上,而不是在左翅上,有一块云母般的透明小板,被翅脉包围着,称为响板。该科另一成员,葡萄隐螽(Ephippiger vitium),有一种奇妙的次要变化;因其鞘翅大为缩小,但“前胸后部隆起成圆屋顶状,而超出鞘翅之上,这大概是为了增强声音的效果”。 (174)
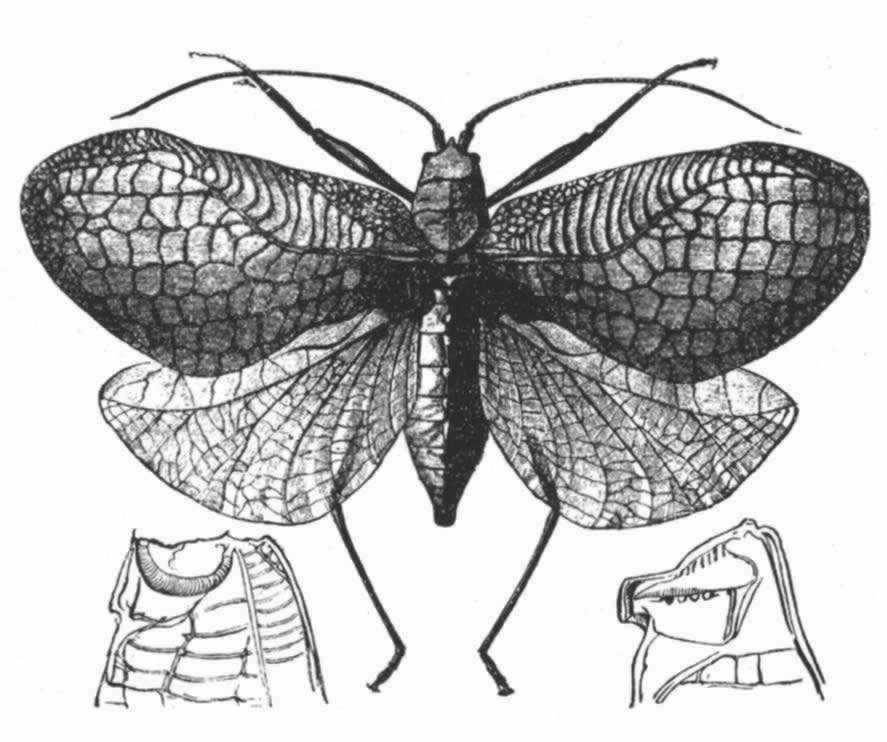
图 13 Chlorocaelus Tanana(引自贝茨)
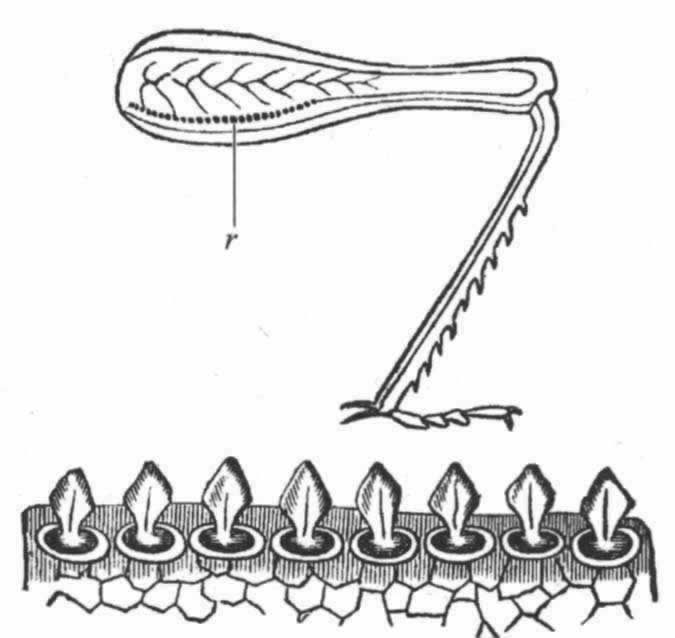
图 14 草蝗(Stenobothrus pratorum)的后肢r摩擦发音的脊;下图是放大很多的组成这条脊的齿(引自兰多依斯)
因此,我们看到螽斯科的音乐器官(我相信在这一“目”中包括有最强有力的演奏者)比蟋蟀科的更加分化或更加特化了,蟋蟀科的两个鞘翅在构造上都是一样,功能也一样。 (175) 然而,兰多依斯在螽斯科的一种、即黑螽斯属(Decticus)中,发现右翅鞘底面有一行既短又窄、仅是残迹状态的小齿,其右翅位于左翅之下,从不做琴弓之用。我在普通绿螽斯的右鞘翅底边观察到同样的残迹构造。因此我们可以有信心地推定,螽斯科是从现存的蟋蟀科那样的一种类型传下来的,这个类型的两张鞘翅底面都有锯齿状翅脉,而且同样都可作为琴弓之用;但在螽斯科中,这两张鞘翅就逐渐分化而且完善了,按分工原理,一张专门作琴弓用,另一张则作提琴用。格鲁勃博士持有相同的观点,他曾阐明残迹齿状物一般见于右翅的下面。蟋蟀科这种比较简单的器官是经过怎样步骤发生的,我们还弄不清楚,但大致的情况可能是这样:两张鞘翅的基部就像它们现在那样地彼此重叠;其翅脉摩擦所产生的音响是嘎嘎的,同现在雌虫鞘翅摩擦发出的声音一样。 (176) 雄者偶尔或意外发出的这样嘎嘎声,如果对雌者曾起过哪怕是一点爱情召唤的作用,大概就会容易地在性选择的作用下通过翅脉粗糙化的变异而得到加强。
在最后一科也是第三个科即蝗科中,其摩擦发音则是按照很不相同的一种方式进行的,按照斯卡德博士的说法,其叫声远不及前两个科那样尖锐。在其腿节内表面(图14,r)有一列纵向的、小巧玲珑的口针状弹性齿,齿数85到93个; (177) 这些齿状物在鞘翅的锐利而突出的翅脉上擦过,就这样引起鞘翅振动而发出声响。哈里斯说, (178) 当一只这种雄虫开始鸣奏时,它先“把后腿的胫节弯到股节之下,那里预先设计有一道小沟以容纳之,然后把腿轻快地上移动。两边的提琴并不一起演奏,而是先奏一个再奏另一个,交替进行”。在这一科的许多物种中,其腹基凹陷,成一大空腔,据信这是作为共鸣板之用的。属于本科一个南非的属,叫做牛蝗(Pneumora)(图15),在这里我们遇到了值得注意的新变异;其雄者从腹部两边斜着各伸出一道具有小缺刻的脊,与后股节互相摩擦。 (179) 值得注意的是,尽管这种雄虫有翅(雌虫无翅),但股节不像通常那样同翅鞘相摩擦;不过这一点也许可用后肢异常短小的情况来解释。我未能检查其股节的内面,但根据类推,可以判断那里大概有细齿。牛蝗属(Pneumora)的诸物种在摩擦发音方面所发生的变异比其他任何直翅类昆虫都更为深刻;因为,其雄者全身已经变成一个乐器,体内充满空气而膨胀,像一个透明大气胞,以便增强共鸣的效应。垂门先生告诉我说,在好望角这些昆虫每到夜间就发出令人吃惊的喧嘈声。
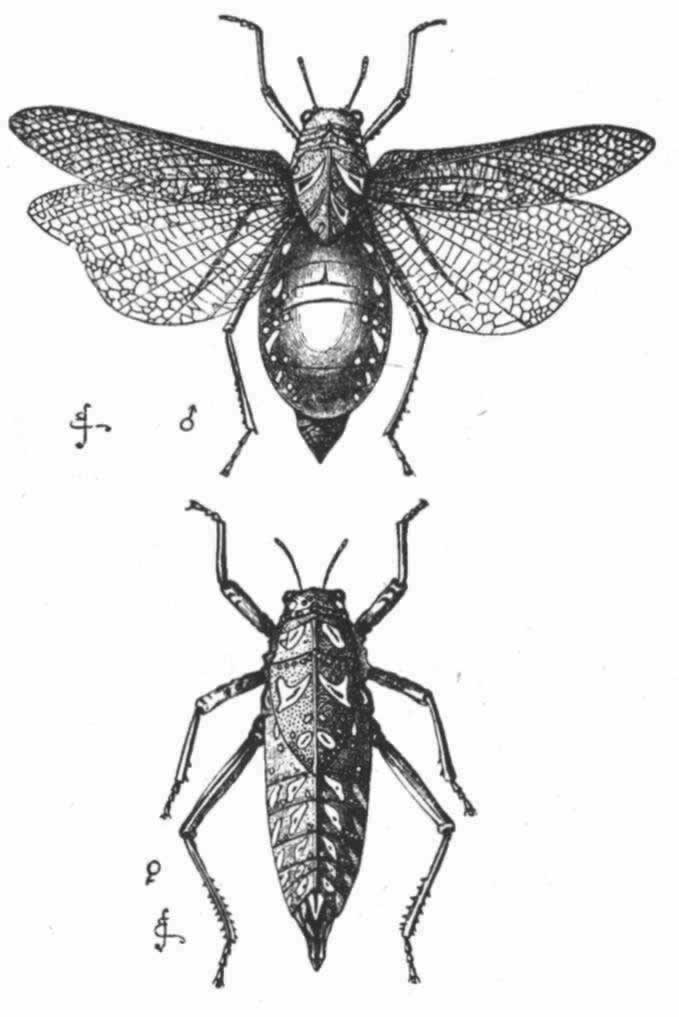
图 15 牛蝗(Pneumora)(根据大不列颠博物馆的标本绘制)
上图为雄虫;下图为雌虫。
在以上三个科中,雌虫几乎总是缺少某种有效的音乐器官,但这个规律也有少数例外,因为格鲁勃博士曾阐明葡萄隐螽(Ephippiger vitium)的雄虫和雌虫都有这类器官,尽管它们的这类器官仍存在某种程度的差异。因此,我们不能假定这类器官是由雄者传递给雌者的,而许多其他动物的次级性征似乎就是这样。它们必定是分别在雌雄双方独立发展起来的,这些雄虫和雌虫一到求偶季节无疑就互相召唤。螽斯科大多数其他昆虫的雌者[据兰多依斯说黑螽斯属(Decticus)除外]具有雄者所特有的摩擦发音器官的残迹,这大概是由雄者传递来的。兰多依斯还在蟋蟀科雌虫的鞘翅底面和蝗科雌虫的腿节上找到这类残迹物。在同翅目中,雌虫也具有丧失作用的这种特有的音乐器官;此后我们还会在动物界其他部门中遇到许多这类的事例,即雄者所特有的构造在雌者身上表现为残迹状态。
兰多依斯观察了另一个重要事实,即,蝗科雌虫腿节上摩擦发音的齿其状态终生保持不变和其最初出现于雌雄幼虫期的状态是同样的。另一方面,雄者的这类器官则继续进一步发育,当它们最后一次蜕皮、即已经成熟并准备繁殖的时候,这类器官就获得了完善的构造。
从现在已经举出的事实来看,我们知道直翅目雄虫的发音手段是极其多种多样的,而且和同翅亚目所使用的发音手段完全两样。 (180) 然而在整个动物界里我们常常发现可用极其多样的手段来达到同一个目的,这似乎是由于世世代代以来整个体制经历了各种各样的变化,在一部分跟着一部分发生变异时,不同的变异都给同一个总目的带来了好处。直翅目这3个科的以及同翅目的各种各样的发音手段给我们留下的深刻印象是,这类构造为了雄虫召唤和诱惑雌虫是高度重要的。根据斯卡德博士的卓越发现, (181) 我们现在知道直翅目有用之不尽的时间在这方面发生变异,所以我们就无须对它们的变异量之大感到惊奇了。这位博物学家最近在新不伦斯威克泥盆纪形成的地层中发现了一只化石昆虫,它具有“螽斯科雄虫的著名的鼓膜或摩擦发音器官”。这种昆虫虽然在大多数方面同脉翅目(Neuroptera)有关系;但似乎把两个有关系的目即脉翅目和直翅目连接起来了,有许多很古老的类型都是如此。
关于直翅目,我还要略谈一二。有些物种非常好斗:当两只雄的田蟋蟀(Gryllus campestris)关在一块时,它们就会斗到其中一只被杀死为止;螳螂属(Mantis)的一些物种被描写得像骑兵挥舞马刀那样地运用其剑状前肢。中国人把蟋蟀养在小竹笼里,使它们相斗 (182) ,就像斗鸡一样。在颜色方面,有些外来的蝗虫装饰得很漂亮;后翅有红、蓝、黑的斑点;但就整个这个“目”来说,雌雄二者在颜色上很少有大的差异,它们的鲜艳色彩大概不是由于性选择所致。鲜明颜色对这些昆虫的用处可能是要引起其他动物注意它们是不好吃的。例如,有人观察过 (183) 把一只色彩鲜艳的印度蝗虫给鸟类或蜥蜴去吃时,它们总是拒绝食用。然而已经知道有几个例子表明在这个“目”中雌雄体色有差异。有一种雄的美国蟋蟀 (184) 被描写白得像象牙一般,而雌者的颜色则变化不定,从接近白色到青黄色或微黑色的都有。沃尔什先生告诉我说,Spectrum femoratum[竹节虫科(Phasmidae)的一种]的雄性成虫“具有发亮的褐黄色;雌性成虫则呈暗淡无光的灰褐色;而雌雄两性的幼虫都是绿色的。最后,我还要提一下,有一种奇异种类的蟋蟀 (185) ,其雄者具有“一个长的膜质附器,就像一幅面纱似的将其脸部盖住”,但什么是它可能的用途,还不清楚。
脉翅目(Neuroptera)
除颜色外,在这里没有什么值得一谈。在蜉蝣科(Ephemeridae)中,雌雄二者在其暗淡颜色上往往稍有差异, (186) 但这大概不至于使雄者因此就能吸引雌者。蜻科(Libellulidae)以鲜艳的绿色、蓝色、黄色和朱红的金属色彩来装饰自己,雌雄二者在体色上常有差异。例如,像韦斯特伍德教授所述说的 (187) ,有些色 科(Agrionidae)昆虫的雄者“具有浓艳的蓝色和黑色翅膀,雌者则呈优雅的绿色而翅膀无色”。但在红色
科(Agrionidae)昆虫的雄者“具有浓艳的蓝色和黑色翅膀,雌者则呈优雅的绿色而翅膀无色”。但在红色 (Agrion Ramburii)中,其雌雄二者的颜色正好同上述相反。 (188) 北美有一个大属,叫做宽角阎虫属(Hetaerina),只有雄者在每个翅基部具有洋红色的美丽斑点。有一种蜻蜓(Anax junius),其雄者的腹基部呈鲜艳的绀青蓝色,而雌者的则呈草绿色。另一方面,在一个亲缘关系密切的箭蜓属(Gomphus)中以及另外一些蜻蜓属中,雌雄二者在体色上的差别很小。在整个动物界的亲缘关系密切的诸类型中,与此相似的情况经常出现,即,有些雌者和雄者的体色差别很大,有些差别很小,有些则完全没有差别。虽然许多种蜻蜓的雌雄二者在体色上差别很大,但往往很难说何者更漂亮;而且如我们刚刚见到的,在色
(Agrion Ramburii)中,其雌雄二者的颜色正好同上述相反。 (188) 北美有一个大属,叫做宽角阎虫属(Hetaerina),只有雄者在每个翅基部具有洋红色的美丽斑点。有一种蜻蜓(Anax junius),其雄者的腹基部呈鲜艳的绀青蓝色,而雌者的则呈草绿色。另一方面,在一个亲缘关系密切的箭蜓属(Gomphus)中以及另外一些蜻蜓属中,雌雄二者在体色上的差别很小。在整个动物界的亲缘关系密切的诸类型中,与此相似的情况经常出现,即,有些雌者和雄者的体色差别很大,有些差别很小,有些则完全没有差别。虽然许多种蜻蜓的雌雄二者在体色上差别很大,但往往很难说何者更漂亮;而且如我们刚刚见到的,在色 属的一个物种中,雄者和雌者的正常体色却互相颠倒了。它们在任何情况下所获得的颜色大概都不是作为保护之用的。对这一科昆虫曾经密切研究过的麦克拉克伦先生写信告诉我说,蜻蜓——昆虫世界的暴君——是任何昆虫当中最不容易受到鸟类和其他敌害的攻击的,他相信它们的鲜艳色彩是用来吸引异性的。某些蜻蜓显然受特殊的颜色所吸引:帕特逊先生曾观察到 (189) 其雄者为蓝色的色
属的一个物种中,雄者和雌者的正常体色却互相颠倒了。它们在任何情况下所获得的颜色大概都不是作为保护之用的。对这一科昆虫曾经密切研究过的麦克拉克伦先生写信告诉我说,蜻蜓——昆虫世界的暴君——是任何昆虫当中最不容易受到鸟类和其他敌害的攻击的,他相信它们的鲜艳色彩是用来吸引异性的。某些蜻蜓显然受特殊的颜色所吸引:帕特逊先生曾观察到 (189) 其雄者为蓝色的色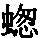 科成群地落在一根钓丝的蓝色浮子上,同时另外两个物种却受耀眼的白色所吸引。
科成群地落在一根钓丝的蓝色浮子上,同时另外两个物种却受耀眼的白色所吸引。
有一个有趣的事实,首先是谢尔沃(Schelver)注意到的,即,在隶于两个亚科的几个属中,其雄虫最初从蛹的状态羽化时在体色上同雌虫的一模一样;但不久它们的身体就呈现出显著的乳蓝色,这是由于有一种可溶于乙醚和酒精的油类分泌出来的缘故。麦克拉克伦先生相信窄腹蜻蛉(Libellula depressa)的雄虫要在变态后经过近两周的期间,即当雌雄准备交配时,才发生这种颜色的变化。
按照布劳尔(Brauer)的说法, (190) 脉翅科的某些物种表现了一种奇妙的二态现象,有些雌虫具有正常的翅,同时另外一些雌虫则“像同种雄虫的翅一样具有很丰富的网脉”。布劳尔“用达尔文的原理解释这现象,假定翅脉紧密相接乃是雄虫的一种第二性征,这种性征并不像一般情况那样传递给所有雌虫,而是突然地传递给一部分雌虫”。麦克拉克伦先生给我讲过另一个二态现象的例子,是关于色 属(Agrion)的几个物种的,在这些物种中有些个体是橙色的,它们必定是雌虫。这大概是返祖的一例;因为在纯系的蜻蜓科中,当雌雄二者在体色上有所差异时,则雌虫都是橙色或黄色的,所以假定色
属(Agrion)的几个物种的,在这些物种中有些个体是橙色的,它们必定是雌虫。这大概是返祖的一例;因为在纯系的蜻蜓科中,当雌雄二者在体色上有所差异时,则雌虫都是橙色或黄色的,所以假定色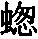 属起源于某个原始类型,这个原始类型在其次级性征上同典型的蜻蜓类相类似,那么只在雌虫方面出现按照这种方式发生变异的一种倾向也就无足为奇了。
属起源于某个原始类型,这个原始类型在其次级性征上同典型的蜻蜓类相类似,那么只在雌虫方面出现按照这种方式发生变异的一种倾向也就无足为奇了。
许多蜻蜓虽然都是大型的、强有力的而且凶猛的昆虫,但麦克拉克伦先生相信,除了色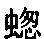 的一些体型较小的物种外,他还没有见过雄蜻蜓互相搏斗的情形。在这个“目”的另一类群中,即白蚁类(Termites),当它们大群出动时,可以看到雌雄二者相互追逐,“雄蚁追在雌蚁后面,有时两只雄的共追一只雌的,以巨大的激情互相竞争,看谁能占有雌者”。 (191) 据说有一种啮虫,叫做白书生(Atropos pulsatorius)会用颚发出喧嘈声,此呼彼应。 (192)
的一些体型较小的物种外,他还没有见过雄蜻蜓互相搏斗的情形。在这个“目”的另一类群中,即白蚁类(Termites),当它们大群出动时,可以看到雌雄二者相互追逐,“雄蚁追在雌蚁后面,有时两只雄的共追一只雌的,以巨大的激情互相竞争,看谁能占有雌者”。 (191) 据说有一种啮虫,叫做白书生(Atropos pulsatorius)会用颚发出喧嘈声,此呼彼应。 (192)
膜翅目(Hymenoptera)
无与伦比的观察家法布尔(M.Fabre) (193) 在描述一种类似黄蜂的昆虫——砂蜂属(Cerceris)的习性时说道:“为了占有某只特殊雌虫,雄虫之间屡屡发生争斗,雌虫则坐以观战,一旦胜负分晓,它就安然地同胜利者一块飞去。”韦斯特伍德说,有一种叶蜂科(Tenthredinae)的雄虫“在争斗中互相用上颚紧紧揪住不放”。 (194) 由于法布尔提到节腹泥蜂属的雄虫努力去获得一只特殊雌虫的情况,因此应好好记住隶于这个“目”的昆虫经过一段长时间后仍有互相识别的能力,而彼此深深依恋。例如,胡伯尔的精确观察是无可怀疑的,他曾把某些蚂蚁分开,四个月后,它们又碰到原来属于同一群体的其他蚂蚁,彼此都能相识并用触角相互爱抚。如果碰到的是陌生的蚂蚁,就不免于争斗。再者,当两群蚂蚁发生战争时,有时在一片混战中同一边的蚂蚁也会互相攻击,但它们很快就会发现错误,其中一只连忙安慰另一只。 (195)
在这个“目”中,体色依性别不同而有轻微差异是常见的,但除蜜蜂科外,很少有显著差异;然而某些类群的雌雄体色是那么鲜艳——例如青蜂属(Chrysis)常见的体色是朱红和金绿色——以致诱使我们把这种结果归因于性选择。根据沃尔什先生的见解 (196) ,姬蜂科(Ichneumonidae)雄虫的体色几乎普遍都比雌虫的浅。另一方面,叶蜂科雄虫的体色一般都比雌虫的深。在树蜂科(Siricidae)中,雄雌的体色常常不同,因此钢青小树蜂(Sirex juvencus)的雄虫具有橙色的带斑,而雌虫则呈暗紫色;但很难说二者之中何者装饰得更好。在鸽形树蜂(Tremex columbae)中,雌虫的体色比雄虫的鲜明。史密斯先生告诉我说,有几个物种的雄蚁呈黑色,而雌蚁则呈褐黄色。
我听同一位昆虫学家史密斯先生说,在蜜蜂科中,特别是在独居的物种中,雌雄的体色常常不同。雄者一般都较鲜明,在熊蜂属(Bombus)以及在Apathus这一蜂属中,雄虫体色的变异比雌虫的大。青条花蜂(Anthophora retusa)的雄虫具有一种浓艳的暗黄褐色,而雌虫则呈全黑色。木蜂属(Xylocopa)几个物种的雌虫也是如此,而雄虫则是鲜黄色的。另一方面,有些物种的雌虫,如金黄地花蜂(Andraena fulva),其体色比雄虫的鲜明。体色上的这类差异几乎不能以下面的说法来解释,即雄虫缺乏自卫能力因而需要这样的保护色,而雌虫则可凭借其螫针来很好地进行自卫。对蜜蜂习性做过特别研究的米勒 (197) 把这种体色差异主要归因于性选择。蜜蜂对颜色有一种敏锐感觉是肯定的。他说,雄蜂热切地寻求雌蜂并为占有它而斗争,他把这种竞争看做是导致某些物种的雄蜂的颚大于雌蜂的原因。在某些场合中,雄蜂的数量不论在季节的早期或在所有时间和所有地点或地区性都比雌蜂多得多;反之,在另外一些场合中,雌蜂的数量又超过雄蜂。有些物种的较美丽的雄蜂似乎是雌蜂选择的对象;另外一些物种的较美丽的雌蜂似乎又是雄蜂选择的对象。结果在某个属中有几个物种的雄蜂在外貌上彼此差异很大,而雌蜂几乎没有差别;在另一个属中情况则相反。米勒相信,某一性别通过性选择所获得的颜色往往以不同的程度传递给另一性别,就像雌蜂的花粉采集器官往往会传递给雄蜂一样,尽管对后者来说这种器官是根本无用的。 (198)
欧洲蚁蜂(Mutilla Europaea)会摩擦发出喧嘈的声音,按照古罗 (199) (Goureau)的说法,其雌雄二者都有这种能力。他认为声音是由第三腹节同前一个腹节摩擦发出的,我发现在这等表面有很细的同心的隆起线;但在头与前胸分节处突出的骨片上也有这样的隆起线,如果用针尖在该骨片上一划,就会发出其特有声响。由于雄虫有翅而雌虫无翅,因此两者都有发音能力是相当奇怪的。众所周知,蜜蜂类以嗡嗡的叫声表达某些像愤怒那样的感情;按照米勒的说法,有些物种的雄蜂当追求雌蜂时会发出一种特别的歌声。
鞘翅目(Coleoptera)(甲虫)
许多甲虫的颜色都同它们常来常往的地面相似,从而避免被其敌害发觉。其他物种,如南美亮壳甲虫(diamond-beetles) (200) ,乃饰以美丽的颜色,组成条纹,斑点,十字花纹以及其他优雅的式样。除非在某些食花物种的场合中,这等颜色几乎不能直接作为保护之用;但根据萤火虫散发萤光的同样原理,这等颜色可能作为一种警号或识别手段。由于甲虫雌雄二者的颜色一般相像,所以我们无法证明这等颜色是通过性选择获得的;但至少这是可能的,因为这等颜色可能先在性别的一方发育然后再传递给另一方;这个观点对那些具有其他十分显著第二性征的类群在某种程度上甚至也是可能适用的。盲甲虫当然不能见到彼此的美丽,听小沃特豪斯(Waterhouse)先生说,它们虽然往往有光滑的外鞘,但决不会呈现鲜艳的色彩;但对于其颜色的晦暗可能做如下解释,即,由于它们一般居住在洞穴中和其他阴暗地方的缘故。
有些天牛(Longicorns),特别是某些锯天牛科(Prionidae)甲虫,却在甲虫雌雄颜色无差别这个规律之外。大多数这等昆虫都是大型的,颜色华丽。我在贝茨先生的采集品中看到,锯天牛属(Pyrodes) (201) 的雄虫颜色一般与其说比雌虫红些莫如说暗些,雌虫或多或少地具有美丽的金绿色。另一方面,有一个物种的雄虫是金绿色的,而雌虫则具有鲜艳的紫红二色。在斑蛾属(Esmeralda)中,雌雄二者的颜色差别如此之大,以致被列为不同的物种;有一个物种,其雌雄二者都具有美丽的鲜绿色,但雄者的胸部则呈红色。总之,按照我所能判断的来说,雌雄颜色不同的锯天牛类,其雌虫颜色要比雄虫的更艳丽,这一点同经过性选择获得颜色的普遍规律是不相符的。
图 16 咖啡独角仙(Chalcosoma atlas)
上图是雄虫(缩小);下图是雌虫(原大)。

图 17 小犀头(Copris isidis)
左图是雄虫
许多甲虫雌雄之间最明显的一个区别就是雄虫由头部、胸部和唇基等处长出的巨角,在少数场合中是从躯体底面长出来的。在庞大的鳃角组中,它们的角同各种四足兽,如公鹿、犀牛等的角相似,不论其大小还是其各式各样的形状都令人吃惊。为了免去描述,我举出了一些比较显著的类型的雄虫和雌虫的绘图(图16到图20)。雌虫一般以小瘤或隆起的形式来表示角的残迹;但有些雌虫甚至连最细小的残迹物也没有。另一方面针角亮蜣螂(Phanaeus lancifer)雌虫的角几乎同雄虫的一样发达;该属以及小犀头属(Copris)的另外一些物种,其雌虫的角虽也发达,但比雄虫的稍差。贝茨先生告诉我说,这等角的差别同本科某些亚部之间更为重要的性状差异并不一致。例如,在黑团蛂属(Onthophagus)的同一部中,有的物种只生单独一只角,另外的物种则生两只角。
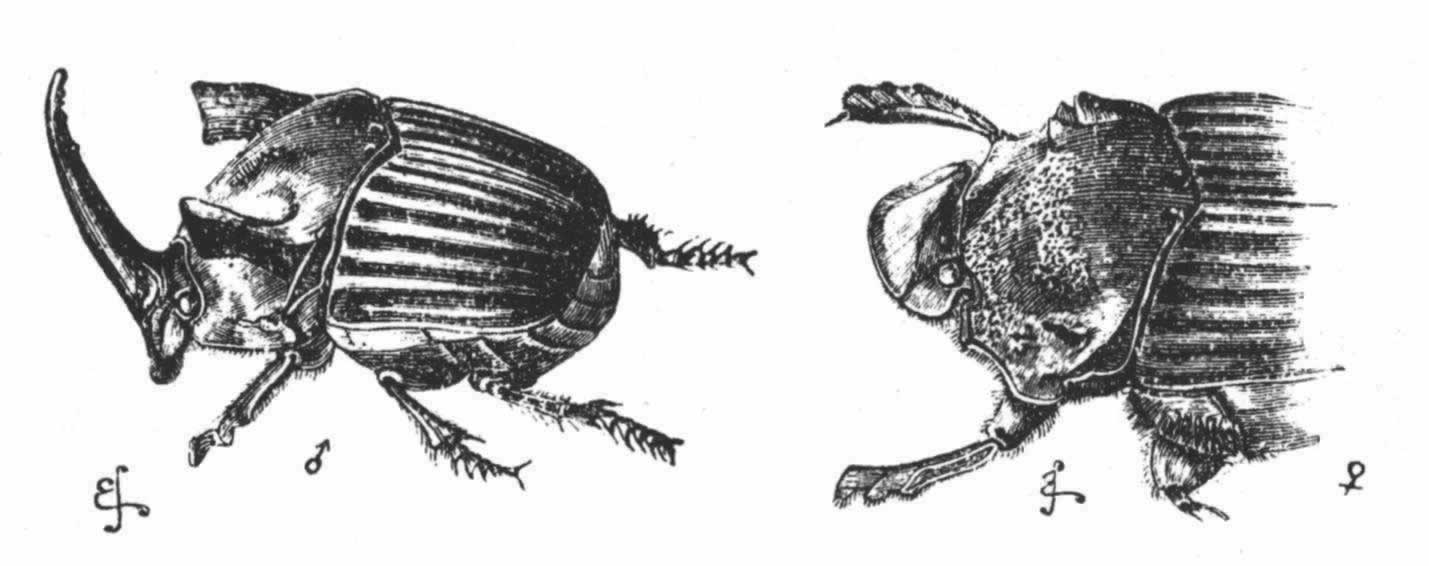
图 18 地区亮蜣螂(Phanaeus faunus)

图 19 广东金龟子(Dipelicus cantori)

图 20 黑团蛂(Onthophagus rangifer)(放大图)
几乎在所有情况下,这等角都以它们的极端变异性而著称,故可形成一个级进的系列,从具有最高度发达的角的雄虫到角已退化到仅仅能同雌虫加以区别的其他雄虫。沃尔什先生 (202) 发现在闪亮蜣螂(Phanaeus carnifex)中,有些雄虫的角长为其他雄虫的三倍。贝茨先生对一百只以上黑团蛂(Onthophagus rangifer)(图20)的雄虫进行了调查之后,认为他终于发现了一个物种,它的角没有发生过变异;但进一步的研究证明事实正好相反。
角的异常之大以及在近亲类型中角的构造的巨大差异都表示这些角是为了某种目的而形成的,但同一物种的雄虫的角表现了极端的变异性,这便引导我们推论这种目的并不具有确定的性质。这些角没有露出曾用于任何正常工作的摩擦痕迹。有的作者设想 (203) 雄虫到处漫游远比雌虫为甚,所以它们需要角以抵御敌害,可是由于这些角往往都是钝的,因此它们似乎并不适于防御之用。最明显的猜测乃是雄虫用这等角彼此相斗;可是从未见过雄虫相斗;贝茨先生详细检查了为数众多的物种以后,也没有能够从它们残断或破碎的状态中找到任何充分的证据来证明这等角曾用于相斗。如果雄虫是惯常的斗士,那么,它们的躯体大概就会通过性选择而增大,以至超过雌虫的躯体;但贝茨先生对金龟子科(Copridae)的一百个物种以上的雌雄二者作了比较之后,也没有在发育良好的个体中找到任何这方面的显著差异。此外,Lethrus是属于鳃角组这同一大部的一种甲虫,据知其雄虫是相斗的,但它们没有角,虽然它们的上颚要比雌虫的大得多。
有一种结论说这等角是作为一种装饰而被获得的,这同下述事实最相符合:即这等角已发展到如此巨大的地步,却还没有固定下来——在同一物种中角的极端变异性以及在亲缘密切的物种中角的多样性都阐明了这一点。最初看来,这种观点好像极不可能;但我们以后将在许多远为高等的动物中,如鱼类、两栖类、爬行类和鸟类,发现各种各样的脊突、瘤状物、角和肉冠显然都是为了这唯一目的而发展起来的。
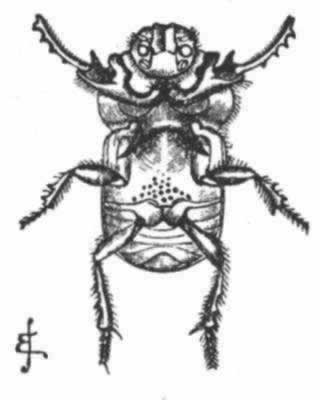
图 21 叉角蜣螂(Onitis furcifer)
雄虫底面图
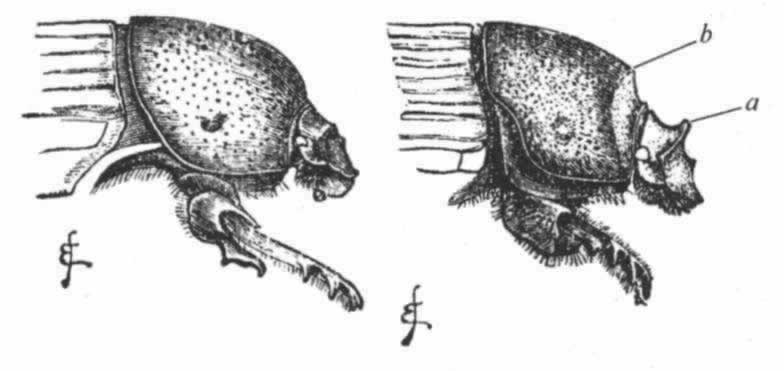
图 22 左方为叉角蜣螂雄虫的侧面图,右方为雌虫图
a.头角的残迹;b.胸角或胸突的残迹。
有叉角蜣螂(Onitis furcifer)(图21),其雄虫以及本属一些其他物种的雄虫在其前肢节上都具有奇特的突起,并在其胸部底面生有一只大型叉角或一对角。根据其他昆虫来判断,这等构造可能有助于雄虫紧紧抱住雌虫。雄虫虽然在躯体的上部表面连一点角的残迹也没有,但雌虫头上却明显地呈现着一个单角的痕迹(图22,a),并在胸部有一个胸突(图22,b)。雌虫这种微小的胸突显然是雄虫所特有的一种突起的残迹,虽然这个特殊物种的雄虫完全没有这种突起,因为野牛布蜣螂(Bubas bison)(次于Onitis的一个属)的雌虫在其胸部具有一个同样的小突起,而雄虫却在同一部位长出一个大型突起。因此几乎毫无疑义的是,叉角蜣螂的雌虫头上的那个小点(甲)以及两三个亲缘相近的物种的雌虫头上的小点,都是代表头角的一种残迹,这种头角实为许多鳃角组甲虫的雄者所共有,如亮蜣螂(Phanaeus)(图18)就是如此。
旧的信念认为这等残迹物是为了完成自然界的计划而被创造出来的,这同实际情况非常不符,以致我们在这一科中所看到的正常状态正好完全相反。我们可以合理地猜测最初是雄虫生角,后来以残迹的状态把它们传递给了雌虫,正如其他许多鳃角组甲虫的情况那样。为什么雄虫后来失去了角,我们还不清楚;但由于其躯体底面发育了巨大的角和突起,这可能是由补偿原理所引起的;而且由于这只限于雄虫才有,所以雌虫上部的残迹角就不会这样消失掉。
迄今为止我们所举的例子都是关于鳃角组甲虫的,还有少数其他雄甲虫属于两个大不相同的类群,即象虫科(Curculionidae)和隐翅虫科(Staphylinidae),也都有角——前者的角在躯体的底面,后者的角则生于头部和胸部的上面。 (204) 在隐翅虫科中,同一物种的雄虫的角变异多端,正如我们在鳃角组甲虫中所看到的那样。在扁鳖(Siagonium)中,我们看到一个二态现象的例子,因其雄虫可分成两组,在躯体大小及角的发达等方面都有巨大差异,但无居间的级进。关于隐翅虫(Bledius)的一个物种(图23),也是属于隐翅虫科的,韦斯特伍德教授说道,“在同一地方能够找到的雄虫标本,有的胸部中央角很大,但头角完全处于残迹状态;有的胸角则非常之短,但头部突起却是长的”。 (205) 这里我们显然看到了一个补偿的例子,刚才提到的雄叉角蜣螂失去上部角的设想,也可借此得到解释。
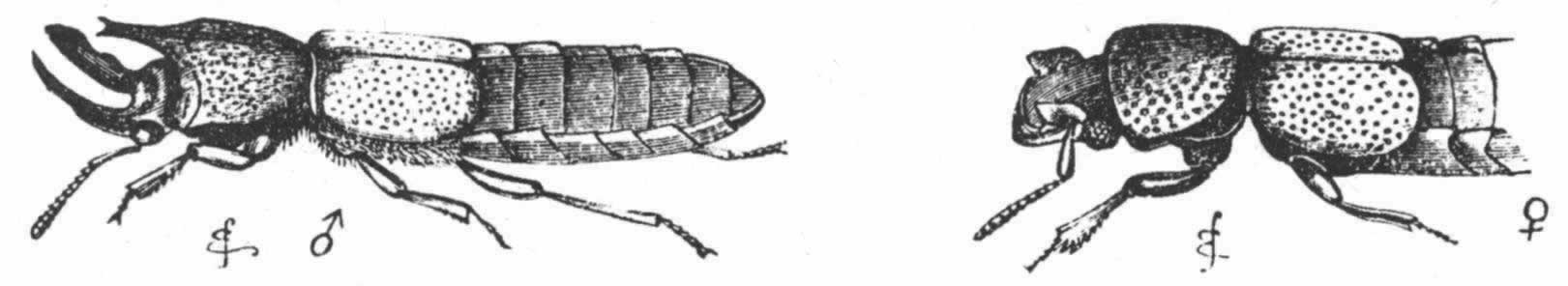
图 23 一种隐翅虫(Bledius taurus)(放大图)
左图雄虫;右图雌虫。
战斗的法则
有些雄甲虫似乎不适于战斗,然而为了占有雌虫也照样卷入冲突。有一种喙很长的线状甲虫,叫Leptorhynchus angustatus,华莱士先生 (206) 见过两只这种雄虫“为一只雌虫而战斗,后者则在一旁忙于钻孔。这两只雄虫用喙相互冲撞,用爪抓来抓去,砰砰地打来打去,显然处于激怒中”。然而较小的雄虫“很快跑开了,承认自己打败了”。在少数场合中,雄甲虫由于具有比雌甲虫上颚大得多的刻齿的巨大上颚,很适于战斗。普通的鹿角锹甲虫(Lucanus cervus)就是如此,其雄虫比雌虫约早一周从蛹羽化,因而往往可以见到若干雄虫追逐同一只雌虫。在这个季节内,它们的冲突进行得很激烈。当戴维斯先生 (207) 把两只雄虫和一只雌虫关在一个盒内时,大的雄虫猛钳小的雄虫,直到后者放弃了它的要求而后已。一位朋友告诉我说,有个小孩常把雄虫放到一块看它们相斗,他注意到它们就像高等动物那样都比雌虫勇敢而凶猛。要是捉拿雄虫的前部,它们就会抓住他的指头不放,而雌虫虽有更强大的上颚却不会这样。锹甲科(Lucanidae)许多种类的雄虫以及上述Leptorhynchus的雄虫,都大于雌虫而且更有力量。大头粪金龟(Lethrus cephalotes,鳃角组甲虫的一种)的雌雄二者同住一穴;雄虫的上颚比雌的大。如果有一只陌生雄虫在繁殖季节企图撞入洞穴里来,就会受到袭击;雌虫不是处于被动地位,而是堵住洞口,并不断地从后面推其伴侣向前以资激励;战斗将一直持续到入侵者被杀死或逃走才告结束。 (208) 另一种疤痕金龟子(Ateuchus cicatricosus)的雌雄二者成双成对地生活在一起,而且似乎彼此非常依恋;雄虫鼓励雌虫去滚动粪球,产卵其中;如果雌虫被移走,雄虫就变得非常焦躁不宁。若雄虫被移走,雌虫就会停止一切工作,而且如布律勒里(M.Brulerie) (209) 所相信的,它将留在同一地点不去,直到死去。
锹甲科雄虫的巨大上颚在大小和构造两方面都是极其易变的,在这方面,同许多鳃角组和隐翅虫科(Staphylinidae)雄虫的头角和胸角相类似。因而介于装备最好的类型和装备最差或退化的类型之间的一个完整系列得以形成。普通锹形虫的、可能还有其他许多物种的上颚虽是用做战斗的有效武器,但其上颚之大是否也能如此解释尚属疑问。我们已知道北美洲的大锹甲(Lucanus elaphus)是用上颚去抓握雌虫的。由于它们如此显眼并具有如此优美的分枝,再加上长度大,因而并不十分适于抱握雌虫;我脑子里交织着这样猜测,即它们可能附带有装饰的作用,正如上述各个不同物种的头角和胸角那样。智利南部的巨颚甲虫(Chiasognathus grantii)雄虫——属于同一科的一种美丽甲虫——具有异常发达的上颚(图24);它勇猛而好斗;当遇到威胁时,它就转过身来,张开巨颚,同时摩擦发出高叫。但其上颚不够有劲,挟住我手指还感不到真正苦痛。
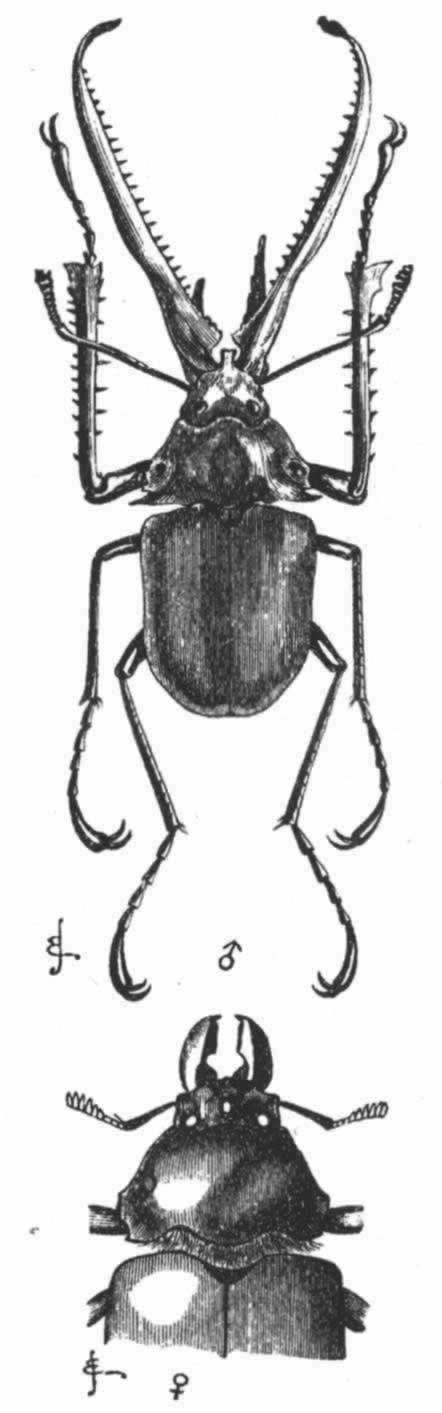
图 24 巨颚甲虫(Chiasognathus grantii)缩小图
上图为雄虫;下图为雌虫。
意味着具有相当的知觉能力和强烈情欲的性选择对鳃角组甲虫比对任何其他科的甲虫似乎更加有效。有些物种的雄虫具有战斗的武器;有些物种成对生活,显示有相互的爱情;许多物种受到刺激时都有摩擦发音的能力;许多物种具有异常大的角,显然是作为装饰之用;有些物种具昼间活动的习性,它们的颜色都很华丽。最后,世界上最大的几种甲虫都属这一科,林纳和法布尔都把这一科分类在鞘翅目之首。 (210)
摩擦发音器
许多差别很大的科的甲虫都具有这类器官。这样发出的声音有时在几英尺、甚至几码外仍可听到, (211) 但是这种声音是无法同直翅目发出的声音相比的。这种音锉一般是由一个稍微升起的窄表面构成的,其上横亘着很细的平行肋状突起,有时如此之细,致成虹色,而且在显微镜下显出一种很漂亮的模样。在某些场合中,如歹粪金龟属(Typhoeus),其音锉整个周围表面布满了硬毛状或鳞片状微小突起,差不多成为平行线,由此逐渐过渡到音锉的肋状突起。这一过渡的完成是靠着那些微小突起汇集成一条直线,而且变得更突出和平滑。躯体的邻接部位上有一条硬脊作为音锉的刮具之用,但在某些场合中,为了这种用途,该刮具已经发生过特殊改变。它迅速地刮过音锉,或者反过来,音锉擦过刮具。

图 25 埋葬虫(兰多依斯提供)
r.两个音锉。左图是高度放大的音锉一部分。
这类器官所处的位置很不相同。埋葬虫(Necrophorus) (212) 有两片平行的音锉(图25,r),位于第五腹节背面,每片音锉 (213) 由126到140条细肋状突起构成。这些肋状突起同鞘翅的后缘互相刮拨,后者的一小部分伸出其一般轮廓之外。许多负泥虫科(Crioceridae)甲虫、四星锯角叶甲虫(Clythra 4-punctata,叶甲虫科Chrysomelidae的一种)以及拟步行虫科(Tenebrionidae)某些甲虫等 (214) 的音锉都位于腹部的背端,即臀板或前臀板之上,也是用鞘翅按上述同样方式刮拨。属于另一科的异角类(Heterocerus),其音锉位于第一腹节的两侧,而用腿节上的隆起线刮拨。 (215) 某些象虫科(Curculionidae)和步行虫科(Carabidae) (216) ,其发音部分的位置则完全颠倒,因其音锉位于鞘翅的下表面,接近翅尖或沿着翅的外缘那一部分,而腹节的边缘则用做刮具。赫氏龙虱(Pelobius Hermanni,龙虱科Dytiscidae或水甲虫的一种)有一条坚固的隆起线靠近鞘翅接合缝的边缘并与之平行,且诸肋状突起横过其上,这些肋状突起中央粗而两端逐渐变细,上端特别细;当在水中或空中把这种昆虫抓住时,它就用腹部的极度角质化边缘刮拨音锉,发出一种唧唧叫声。大量的长角甲虫(Longicornia)的这类器官位置则完全不同,其音锉位于中胸,而同前胸互相摩擦;兰多依斯在英雄天牛(Cerambyx heros)的音锉上数出238条很细的肋状突起。
许多鳃角组昆虫都有摩擦发音能力,但发音器官的位置大不相同。有些物种摩擦发出的声调很高,以致当史密斯先生捉到一只砂蛂(Trox sabulosus)时,站在旁边的一位猎物看守人竟以为他逮住了一只老鼠;但我没有发现这种甲虫特有的发音器官。在推丸蜣螂(Geotrupes)和歹粪金龟(Typhoeus)中,有一条窄隆起线斜穿(图26,r)每只后足的基节(在G.stercorarius中,有84条肋状突起),由一个腹节特别突出的部分向它刮拨。近亲的镰刀形角金龟子(Copris lunaris)沿着其鞘翅边缘的接合缝有一条非常窄而细的音锉,靠近基部的外缘还有另一片短音锉;但据勒孔特(Leconte)说 (217) ,有些其他金龟子的音锉则位于腹部的背面。独角仙(Oryctes)的音锉位于前臀板;据这同一位昆虫学家说,有些其他独角仙类(Dynastini)的音锉则位于鞘翅的底面。最后,韦斯特林说,褐绢金龟(Omaloplia brunnea)的音锉位于前胸腹,而刮则位于后胸腹板,这样,发音部分所在的位置是躯体的下表面,而不是像长角甲虫那样在上表面。
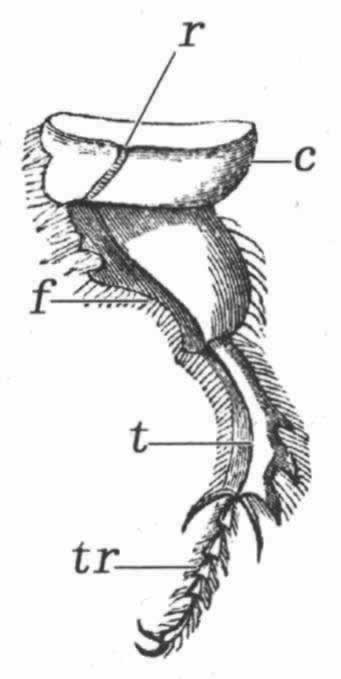
图 26 粪蜣螂(Geotrupes stercorarius)后肢图(引用Landois)
r.音锉;c.基节;f.股节;t.胫节;tr.跗节。
我们由此看到鞘翅类不同科的摩擦发音器官位置的多种多样,实是惊人,但在其构造上却没有多大差别。在同一科中,有些物种具有这等器官,而另外一些物种就没有。这种差异是可以理解的,我们如果设想各种甲虫躯体的任何坚硬而粗糙的部分原先偶然相触,互相摩擦而发出模糊或嘶嘶的声音;而且由于这样发出的声音有点用处,那么其躯体的粗糙表面就会逐渐发展成为正规的摩擦发音器官。现在有些甲虫当行动时,不管有意或无意地还会发出一种模糊的声音,然而它们并没有任何适于这种用途的特殊器官。华莱士先生告诉我说,长臂金龟子(Euchirus longimanus,一种鳃角组甲虫,其雄虫的前肢奇长)“在移动时靠腹部伸缩发出一种低沉的嘶嘶声;如果把它捉住,它就会用后腿同鞘翅边缘互相摩擦而发出一种刺耳的声音”。这种嘶嘶声显然是由于一个窄音锉沿着每张鞘翅边缘的接合缝擦过而发出的;我用它的腿节的粗糙表面同其对应的鞘翅凸凹不平的边缘互相摩擦,同样也能发出那种刺耳的声响;然而我无法在这里找出任何特殊的音锉,这种昆虫如此之大,我很不可能把这种音锉忽略掉的。在考察了高脊步行虫(Cychrus)并读过韦斯特林关于这种甲虫的著述后,我感到尽管它有发音能力,但是否有任何真正的音锉,似乎很有疑问。
根据直翅目和同翅亚目来类推,我曾预期在鞘翅目昆虫中会发现不同性别有不同的摩擦发音器官;但详细检查过若干物种的兰多依斯并没见到过这种差异;韦斯特林和克罗契(G.R.Crotch)先生制作了许多标本送给我,他们也没有见过这种差异。由于这类器官的巨大变异性,即便有任何差异,如果是轻微的话,也是难以被察觉的。例如我检查的第一对埋葬虫(Necrophorus humator)和Pelobius,其雄虫的音锉要比雌虫的大得多;但后来检查的标本就不这样了。有三只粪蜣螂雄虫的音锉在我看来要比三只雌虫的音锉更厚,色更暗,也更隆起,因此,为了弄清楚不同性别的摩擦发音能力是否不同,我儿子F.达尔文先生搜集了57只活标本,用同样方法拿着,按照它们叫声的大小分成两堆。然后他检查了所有这些标本,发现这两堆雄虫和雌虫的比例很接近。史密斯先生保存了许多Monoynchus pseudacori(象虫科)的活标本,他认为其雌雄二者都会摩擦发音,而其发音程度显然是相等的。
尽管如此,在某些少数鞘翅目昆虫中,摩擦发音能力肯定还是一种性征。克罗契先生发现Heliopathes(拟步行虫科)的两个物种只有雄虫具有摩擦发音器官。我检查了驼背拟步行虫(H.gibbus)的五只雄虫,在最末腹节背面全有一个相当发达的音锉,其一部分分而为二;而在同样数目的雌虫中,甚至连一个音锉的痕迹也没有,最末这个腹节的膜是透明的,而且比雄虫的这种膜薄得多。一种拟步行虫(H.cribratostriatus)的雄者有一个同样的音锉,只是其一部分并不分而为二,而雌虫则完全缺少这种器官;此外雄虫在鞘翅尖端的边缘上,在鞘翅接合缝的每一边,有三四条短的纵向隆起线,其上横亘着极细的肋状突起,这等隆起线同腹部的音锉平行也相类似;这等隆起线究竟是作为独立的音锉之用还是作为腹部音锉的一个刮具,我还无法断定:雌虫一点也没有后述这种构造的痕迹。
还有,在鳃角类独角仙属(Oryctes)的三个物种中,我们看到一个近似的例子。钩角独角仙(O.gryphus)与尖鼻独角仙(O.nasicornis)雌虫的前臀板音锉上的肋状突起在连续性和清晰性上均不及雄虫;但主要的差异还在于这个体节的全部上表面,当把它放在适当光线中时,即可见到它上面覆盖着毛,而雄虫并没有这种毛,或仅以非常微细的绒毛为其象征。应该注意,在所有鞘翅目昆虫中,音锉的有效部分都是无毛的。塞内加尔独角仙(O.senegalensis)雌雄之间的差异更加强烈显著,当把这个特殊腹节弄干净作为透明物体观察时,就可以最清楚地看到这种差异。雌虫的这整个表面覆盖着分散的带刺小脊突;而雄虫的这些脊突在向腹端延伸的过程中逐渐会合,变得愈益规则,愈益没有毛刺;因而这个腹节的四分之三被极细的平行肋状突起所覆盖,是为雌虫所根本没有的。然而当把一个软化了的标本的腹部前后推动时,独角仙属所有这三个物种的雌虫都会发出一种轻微的嘎嘎声或唧唧声。
在拟步行虫属和独角仙属的场合中,雄虫的摩擦发音乃是为了召唤或刺激雌虫,几乎是无可疑问的了;但对多数甲虫来说,摩擦发出的叫声显然是用于雌雄的相互召唤。甲虫类在各种情绪下的摩擦发音,也同鸟类一样,除向配偶鸣唱外,还为了许多目的来使用它们的叫声。巨大的巨颚甲虫当愤怒和挑战时就要摩擦发出鸣叫,许多物种如果被捉住因而无法逃脱时,由于绝望或恐惧也会发出鸣叫;沃拉斯顿(Wollaston)和克罗契二位先生在加那利群岛用敲打空心树干的方法可以引起仙人掌象虫属(Acalles)的甲虫摩擦发出鸣叫,因而探知它们的所在。最后,金龟子(Ateuchus)雄虫摩擦发出鸣叫以鼓励雌虫工作,当把雌虫移走后,也因悲痛而摩擦发出鸣叫。 (218) 有些博物学家相信甲虫发出这种叫声是为了把它们的敌害吓走;但我不能想象一只四足兽或鸟既然能吞食一只大甲虫,怎么会被这么轻微的一种声音所吓倒。摩擦发音是用于性的召唤,这个信念得到了下述事实的支持,即方格斑纹窃蠢虫(Anobium tessellatum)以滴答声互相呼应而闻名,而且据我亲自的观察,它们也向一种人为的轻拍声呼应。道布尔戴先生也告诉过我,他不时见到一只雌虫发出滴答声, (219) 过一两小时后发现它同一只雄虫在交配,还有一次被凡只雄虫包围起来了。最后,许多种类的雌雄甲虫起初很可能是靠它们躯体上邻接的坚硬部分彼此摩擦而发出轻微的声音来相互寻找;当那些声音最响亮的雄虫或雌虫最能成功地寻得配偶时,它们躯体不同部位上的皱纹通过性选择就会逐渐发展成为真正的摩擦发音器官。
第十一章 昆虫类的第二性征(续) ——鳞翅目(蝶类和蛾类)
蝴蝶的求偶——斗争——滴答响声——雌雄共有的颜色,或者雄虫的颜色更鲜艳——实例——非由于生活条件的直接作用——适于保护的颜色——蛾类的颜色——美的夸耀——鳞翅目的知觉能力——变异性——雄虫和雌虫颜色差异的原因——拟态,雌蝴蝶比雄蝴蝶的颜色更鲜艳——幼虫的鲜明颜色——关于昆虫第二性征的提要和总结——鸟类同昆虫的比较
在这个大“目”中,使我们最感兴趣的是,同一物种雌雄二者在颜色上的差异以及同一属不同物种之间在颜色上的差异。本章的绝大部分都要用来讨论这个问题,但在这之前我要先对其他一两个问题略作陈述。常常可以见到若干雄虫群集在同一只雌虫周围,向它求爱。它们的求偶看来是一件延续很久的事,因为我屡屡注视一只或一只以上的雄虫环绕一只雌虫旋转,直到我看得累了的时候还没有结果。巴特勒(A.G.Butler)先生也告诉我说,他曾几次注视过一只雄虫花了整整一刻钟的时间去向一只雌虫求爱,但后者顽固地拒绝它,最后停息在地面上并合拢双翅,以逃避它的求爱。
蝴蝶虽是脆弱的动物,却都好斗,有一只被捉住的“大彩虹蝶” (220) (Emperor butterfly)就是因为同另一只雄蝶冲突而把两片翅尖搞裂了。科林伍德(Collingwood)先生提到婆罗洲蝴蝶经常发生斗争时说道,“它们以最大速度互相围着旋转,似乎激起了极大愤怒而凶猛异常”。
有一种蝶(Ageronia feronia)发出的一种声音就像齿轮在弹簧轮挡下通过时的响声一样,在几码外都能听到:我只是在里约热内卢见到两只这种蝴蝶在一条不规则的路线上互相追逐时才注意到这种声音的,因此这种声音可能是雌雄在求偶时发出的。 (221)
某些蛾类也发音,黾蛾(Thecophora fovea)的雄蛾就是一例。布坎南·怀特(F.Buchanan White) (222) 先生有两次听到 榉青实蛾(Hylophila prasinana)的雄蛾发出一种急促的刺耳声音,他相信,就像蝉属的发音那样,这声音是由具有肌肉的一片弹性膜产生的。他还引用了盖内的说法,即,毛蛾(Setina)显然是靠“位于胸部的两只鼓状大囊”之助,发出一种钟表那样的滴嗒声,而这类器官“在雄蛾身上远比在雌蛾身上发达得多”。因此,鳞翅目的发音器官同性机能似乎有某种关系。我所指的不是骷髅天蛾(Death's Head Sphinx)发出的那种人所熟知的声音,因为在这种蛾刚从茧羽化不久就可听到这种声音。
榉青实蛾(Hylophila prasinana)的雄蛾发出一种急促的刺耳声音,他相信,就像蝉属的发音那样,这声音是由具有肌肉的一片弹性膜产生的。他还引用了盖内的说法,即,毛蛾(Setina)显然是靠“位于胸部的两只鼓状大囊”之助,发出一种钟表那样的滴嗒声,而这类器官“在雄蛾身上远比在雌蛾身上发达得多”。因此,鳞翅目的发音器官同性机能似乎有某种关系。我所指的不是骷髅天蛾(Death's Head Sphinx)发出的那种人所熟知的声音,因为在这种蛾刚从茧羽化不久就可听到这种声音。
霍尔一向观察到天蛾有两个物种散发出麝香气味,这是雄蛾所特有的; (223) 在较高等的动物纲中,我们将会碰到许多只有雄者才散发香气的事例。
许多蝶类和某些蛾类都极其美丽,无论何人,必加赞赏;或可这样提问:它们的颜色及其变化多端的式样是怎样形成的?是由于这些昆虫所暴露于其中的物理条件直接作用的结果吗?同时并不因此取得任何利益吗?还是由于作为一种保护手段,或者为了某种未知的目的,或者为了某一性别可以吸引另一性别,世世代代的变异被积累起来并由它们所决定的吗?再者,某些物种雌雄二者的颜色差异很大,而同一属其他物种雌雄二者的颜色却彼此相像,其意义是什么呢?在试图回答这些问题之前,一定要先举出大量的事实来。
关于我们美丽的英国蝴蝶,像红纹蝶(admiral)、孔雀蛱蝶(peacock)、画美人蛱蝶(Vanessae)以及其他许多蝴蝶,其雌雄二者都是彼此相像的。热带产的艳丽的长翅蝶(Heliconidae)和斑蝶(Danaidae)科的大多数也是如此。但某些其他热带类群以及我们英国的某些蝴蝶,像闪紫蝶(purple emperor),橙色翅尖蝶(orange-tip),等等(紫蛱蝶Apatura Iris和黄斑襟粉蝶Anthocharis cardamines),其雌雄二者在颜色上的差异不是很大就是很小。有些热带物种的颜色之壮丽实非语言所可形容。甚至在同一个属中,我们也常常会发现有的物种雌雄之间表现的差异非常之大,而另外的物种雌雄之间又彼此密切近似。下述大多数事实均蒙贝茨先生见告,并审阅过这里的全部讨论,在南美的Epicalia这一属中他告诉我说,他知道有12个物种的雌雄二者常常出没于同一处所(蝶类并非永远如此),所以它们不会受到不同外界条件的影响。 (224) 这12个物种中有9个,其雄蝶乃所有蝴蝶中最鲜艳者,它们同颜色比较平淡的雌蝶有如此巨大差异,以致后者先前曾被放入不同的属中。这九个物种的雌蝶在其一般色彩上都彼此相似;而且同世界各地所发现的若干亲缘关系密切近似属的物种的雌雄双方都相类似。因此我们可以推论这9个物种,大概还有该属的所有别的物种都是起源于一个颜色几乎相同的祖先类型。第10个物种的雌蝶仍保持相同的一般色彩,但雄蝶与之相似,因此它的颜色远不及前面九个物种绚丽,而且差别悬殊。第11和第12个物种的雌蝶失去了普通的样子,因其颜色几乎与雄蝶一样华丽,只是比后者稍差一点而已。因此,后面这两个物种的雄蝶的鲜明色彩似乎已传给了雌蝶;而第10个物种的雄蝶则保持或复现了雌蝶的以及该属原始类型的平淡颜色。这三个例子的雌雄二者尽管表现的方式相反,却很相似。在亲缘关系相近的Eubagis属中,某些物种雌雄二者的颜色都是平淡的而且近似;而大多数物种的雄蝶都装饰着多种多样美丽的金属色泽,同雌蝶差异很大。这整个属的雌蝶都保持着同样的一般色彩,因此它们互相之间的类似要大于它们和同种雄蝶之间的类似。
在凤蝶(Papilio)属中,安尼阿斯蝶(AEneas)类群的所有物种均以它们显明和差别悬殊的色彩而著称,它们也证明了雌雄二者之间在差异量上常见级进倾向。在少数物种中,例如斑点凤蝶(P.ascanius),雌雄二者彼此相似;在另外一些物种中,雄蝶的色彩或比雌蝶鲜明,或比雌蝶华丽得很多。同英国画美人蝶有亲缘关系的胥蝶(Junonia)属提供了一个几乎同样的情况,因为尽管该属大多数物种的雌雄二者都缺少华丽色彩而且彼此相像,但有些物种如青铜色六月蝶(J.anone)的雄者颜色则比雌者鲜明些,还有胥蝶的少数物种(例如J.andremiaja)的雄者同雌者如此不同,以致可能把雄者误认为是一个完全不同的物种。
A.巴特勒先生在大不列颠博物馆向我指出过另一个显著事例,即热带美洲蚬蝶(Theclae)属的一个物种,其雌雄二者几乎一样,极其美丽;另一个物种的雄者具有同样华丽的颜色,但雌者整个上部表面则都是一致的暗褐色。我们常见的英国灰蝶(Lycaena)的小型蓝蝴蝶表明其雌雄二者之间在色彩上有种种差异,几乎同上述外来的属一样,虽然不如后者那样显著。小丘灰蝶(Lycaena agestis)雌雄二者的翅都是褐色的,边上镶有像瞳眼的橙色小点,彼此都如此相似。雄爱琴岛灰蝶(L.agon)的翅是鲜蓝色的,镶着黑边,而雌蝶的翅都是褐色的,其镶边同小丘灰蝶(L.agestis)的翅很近似。最后,竖琴灰蝶(L.arion)雌雄二者的翅都是蓝色而且很相像,但雌蝶翅缘稍黑,上面的黑点稍淡;有一个鲜蓝色的印度物种雌雄二者彼此还更相像。
我列举上面这些细节是为了阐明,第一,当蝴蝶的雌雄二者出现差异时,按照一般规律总是雄者较为美丽,而且同该种所隶属的那一类群的普通色彩距离较远。因此,在多数类群中若干物种的雌蝶之间的相似远比雄蝶之间的相似更为密切。然而在某些场合中,雌蝶的颜色却比雄蝶更为艳丽,以后我还要谈到这一点。第二,上述这些细节清楚地使我们认识到同一属的雄蝶和雌蝶从其颜色毫无差异开始,经常出现各种级进,直至彼此的颜色如此不同,以致在昆虫学家们把它们归入同一属之前,长期以来一直把它们看做是两个属。第三,我们看到,当雄蝶和雌蝶彼此近似时,这似乎是由于雄蝶将其色彩传给了雌蝶,要不就是由于雄蝶保持或恢复了这一类群的原始色彩。还应注意的是,在雌雄二者有所差异的那些类群中,通常是雌蝶多少有些类似雄蝶,所以当雄蝶美丽到异常程度时,雌蝶也几乎总要呈现某种程度的美丽。根据雌雄二者在差异量上的许多级进事例,根据同一类群普遍具有的一般色彩,我们可以断定导致某些物种只有雄蝶才有鲜艳色彩以及另外一些物种雌雄二者都具有鲜艳色彩的原因一般是相同的。
由于这么多华丽的蝴蝶都产于热带,因此人们往往假定它们的颜色乃是由于该地区的巨大热量和湿度所致;但是,贝茨先生 (225) 在比较了许多产于温带和热带亲缘相近的昆虫类群之后,证明这个观点不能成立;当同一物种的色彩鲜艳的雄蝶和色彩平淡的雌蝶栖息在同样的地方,吃同样的食物并遵循着完全同样的生活习性时,这个证明就成为无可争辩的了。即使雌雄二者彼此相像,我们也几乎不能相信它们灿烂鲜艳的色彩乃是它们组织性质和周围环境条件的毫无目的之结果。
所有种类的动物,一旦为了某种特殊目的而发生了颜色变异,根据我们所能判断的来说,这如果不是为了直接或间接的保护,就是为了性别之间的一种吸引。有许多蝴蝶的种,其翅的上表面都是颜色暗淡的,这多半可以使它们得以避免被发现和逃避危险。但是,蝴蝶在停息时特别容易受到其敌害的攻击;而且大多数种类在停息时都把翅垂直地竖立于背上,于是只有翅的下表面暴露于外界的视线之中。因此,正是这一面的颜色往往模拟它们通常停息于其上的物体色彩。我相信是勒斯勒尔(R ssler)博士首先注意到某些画美人蝶以及其他蝴蝶合拢的双翅同树皮的颜色相似。还可以举出许多类似的显著事实。最有趣的一个例子是华莱士先生 (226) 记载的一种印度和苏门答腊的普通蝴蝶(木叶蝶,Kallima),当它停息在矮树丛上时就像变魔术一样地消失了;因为它把头和触角都藏在合拢的双翅中间,这样从形状,颜色和翅脉来看就和一片带叶柄的枯叶无异。另外还有些例子:翅的下表面具有灿烂的颜色,仍然是作为保护之用。例如红纹蚬蝶(Thecla rubi)的双翅合拢时其颜色是翡翠绿,同黑莓树的嫩叶相类似,这种蝴蝶在春天常常停息于其上。还应注意的是,有许多物种的雌雄二者上表面的颜色差异很大,而下表面的色彩却非常近似或完全一样,这也是作为保护之用的。 (227)
ssler)博士首先注意到某些画美人蝶以及其他蝴蝶合拢的双翅同树皮的颜色相似。还可以举出许多类似的显著事实。最有趣的一个例子是华莱士先生 (226) 记载的一种印度和苏门答腊的普通蝴蝶(木叶蝶,Kallima),当它停息在矮树丛上时就像变魔术一样地消失了;因为它把头和触角都藏在合拢的双翅中间,这样从形状,颜色和翅脉来看就和一片带叶柄的枯叶无异。另外还有些例子:翅的下表面具有灿烂的颜色,仍然是作为保护之用。例如红纹蚬蝶(Thecla rubi)的双翅合拢时其颜色是翡翠绿,同黑莓树的嫩叶相类似,这种蝴蝶在春天常常停息于其上。还应注意的是,有许多物种的雌雄二者上表面的颜色差异很大,而下表面的色彩却非常近似或完全一样,这也是作为保护之用的。 (227)
虽然许多蝶类上下两面的暗淡色彩无疑是便于隐蔽,可是我们不能把这一观点引申到上表面具有鲜艳夺目色彩的那样一些物种,如英国的红纹蝶、孔雀兰蝶、画美人蝶、粉蝶(Pieris),或是常出没于开阔沼地的大燕尾凤蝶——因为这些蝴蝶的颜色使它们可以被每个活的动物都能看到。这些物种的雌雄二者都彼此相似;但山黄粉蝶(Gonepteryx rhamni)的雄蝶是深黄色的,而雌蝶的颜色要淡得多;而黄斑襟粉蝶(Anthocharis cardamines)只有雄蝶的翅尖具有鲜明的橘黄色。在这些例子中,无论雄蝶与雌蝶都会惹起注目的,因而认为它们的颜色差异同正常的保护有任何关系的说法都是不可信的。魏斯曼(Weismann)教授说,有一种灰蝶的雌者当停息在地上时就把她褐色的翅展开,从而几乎无法把它识别出来 (228) ;另一方面,其雄者好像明知其翅膀上表面的鲜蓝色会招来危险,当停息时就把它们合闭在一起,这说明蓝色决不能用于保护。尽管如此,惹起注目的色彩作为表示它们是不好吃的一个警号,对许多物种可能还是间接有利的。因为某些其他例子表明美丽色彩是通过模拟其他美丽物种而获得的,后者也居于同一地方并由于它们对其敌害有某种防卫作用而得以避免受到攻击;但另一方面我们还必须对模拟的物种之美丽色彩进行解释。
正如沃尔什先生向我说过的,上述黄斑襟粉蝶的和一个美国物种(美国襟粉蝶,Anth.genutia)的雌蝶大概向我们表明了该属亲种的原始色彩;因为该属有四五个散布很广的物种,其雌雄二者的色彩几乎是一模一样的。正如上述的几个例子,我们可在这里推断黄斑襟粉蝶和美国襟粉蝶(Anth.genutia)的雄蝶离开了该属的通常形式。产于加利福尼亚州的蛛襟粉蝶(Anth.sara),橘色翅尖的性状在雌蝶方面也得到了局部发育;但颜色比雄蝶要淡些,在其他一些方面也稍有差异。有一个亲缘相近的印度蝴蝶类型叫做齿小蠹(Iphias glaucippe),其橘色翅尖的性状在雌雄双方都得到了充分发育。正如巴特勒先生向我指出的,这个齿小蠹(Iphias)双翅的下表面同一片淡色的叶子奇异地相似;而我们英国的黄斑襟粉蝶的下表面则同野生欧芹的头状花相似,它们常在晚间停息于其上。 (229) 这些都迫使我们相信下表面的色彩乃是为了保护的同样理由,又使我们不得不否认翅尖具有鲜明的黄色是为了同样的目的,特别是当这个性状只限于雄蝶时,尤其如此。
大多数蛾类在整个或大部分白天都不活动,而且其翅垂放;为了逃避外界的发现,它们整个上表面的颜色浓淡和着色方式,正如华莱士先生所说的,常常令人赞叹不已。蚕蛾(Bombycidae)科和夜蛾(Noctuidae)科 (230) 当停息时,其前翅一般重叠而把后翅掩盖起来;因此后翅大概具有灿烂的色彩而不致遭到大的危险;它们的后翅事实上往往都是这样着色的。蛾类在飞翔时常常能够逃避其敌害,尽管如此,由于其后翅这时完全暴露于外界的视线之中,一般说来其灿烂色彩的获得一定还是要冒一点危险的。但下述事实阐明,我们要对这个问题作出结论应该如何地谨慎。普通黄色后翅蛾的毛夜蛾(Triphaena)往往在白天或傍晚飞来飞去,由于它们后翅的颜色,那时是易于被察见的。人们自然要认为这可能是危险的一个根源;但詹纳·韦尔先生相信这实际上是逃避危险的一种手段,因为鸟类所注意到的是这等色彩灿烂而易碎的表面,并非它们的躯体。例如,韦尔先生把一只健壮的一种黄毛夜蛾(Triphaena pronuba)标本放进他的鸟舍里,马上就受到一只驹鸟的追逐;但是,把这只鸟的注意力吸引住的是标本的彩色翅膀,经过50次左右的尝试,而且蛾翅反复被撕裂成碎片后,才把它捉住。他用一只燕子和缘饰毛夜蛾(T.fimbria)在露天做过相同的实验;但这种蛾子的巨大体形妨碍了燕子把它捉获。 (231) 由此我们想起了华莱士先生的一段叙述,他说在巴西森林和马来群岛有许多非常漂亮的普通蝴蝶,它们虽有宽阔的翅膀,但都不善于飞翔;它们“被捉获时,其翅往往因被刺穿而破裂,好像它们曾被鸟类捉住后又逃脱了。倘若翅膀同虫体的比例小得多,那么这种昆虫的致命部位看来就可能更加频繁地受到打击和刺穿,因此翅膀的增大可能有间接的利益”。 (232)
夸 耀
许多蝶类和有些蛾类的灿烂色彩都是为了夸示而特别安排的,所以它们容易被看见。在夜间,颜色是不会被看见的,毫无疑问,夜蛾作为一个整体来说,其装饰远不及具有夜息昼出习性的蝶类华丽。但某些科的蛾类,如斑蛾(Zygaenidae)科、若干天蛾(Sphingidae)科、燕蛾(Uraniidae)科、某些灯蛾(Arctiidae)科和天蚕蛾(Saturniidae)科,在白天或傍晚四处飞翔,它们之中有许多都是极其美丽的,同严格夜出昼息的类型相比,其颜色要灿烂得多。然而也有少数夜出物种具有鲜艳色彩的例外情况曾被记录下来。 (233)
关于夸示,还有另一类证据。如上所述,蝶类当停息时便竖起它们的翅,但在晒太阳的时候,往往把双翅交替地竖起或垂放,这样,翅的两面就充分可见;虽然下表面的色彩往往暗淡,以为保护,但有许多物种,其翅的下表面同上表面装饰得一样,非常华丽,而且其式样有时迥然不同。有些热带物种,其翅的下表面色彩甚至比上表面还要鲜艳。 (234) 英国蛱蝶(Argynnis)只有下表面才装饰着闪闪的银光。尽管如此,作为一般规律,上表面大概暴露的更加充分,其颜色要比下表面更灿烂,更多样化。因此,在鉴定不同物种之间的亲缘关系时,下表面对昆虫学家们一般可以提供更为有用处的性状。弗里茨·米勒告诉我说,在巴西南部他的住宅附近发现了蝶蛾(Castnia)属的三个物种:其中两个物种的后翅颜色都是暗淡的,这两种蝴蝶停息时,其后翅总是被前翅所掩盖;但第三个物种的后翅是黑色的,其上有美丽的红色和白色斑点,这种蝴蝶不论在什么时候停息,它们的后翅都充分展开以显示其色彩。尚有其他这等事例可以举出。
现在我们转来谈谈蛾类这庞大的类群,我听斯坦登先生说,它们习惯上不把翅的下表面暴露得很清楚,我们很少发现这一面的色彩比上表面更灿烂或与之相当。关于这一规律的某些例外,不论是真实的还是表面的,如合欢螆(Hypopyra)的例子 (235) ,都必须加以注意。特里门(Trimen)先生告诉我说,在盖内的伟大著作中有三只蛾的绘图,它们的下表面要鲜艳得多。例如,澳洲枯叶蛾(Gastrophora)的前翅上表面是淡灰赭石色的,而下表面则饰以华丽的钴蓝色眼点,位于一块黑斑的中央,黑斑之外围绕着一层橙黄色,再外一层是浅蓝白色。但关于这三种蛾子的习性还不清楚;因而对它们色彩的不寻常式样无法加以说明。垂门先生也告诉我说,某些别的尺蠖蛾(Geometrae)类 (236) 和四裂的夜蛾(Noctuae)类,其翅膀的下表面或比上表面更色彩斑驳,或更鲜艳灿烂;但其中一些物种具有这样的习性:“它们的翅完全竖立于背上,并在相当长的时间内保持着这种姿势”,这样就把下表面暴露于外界的视线之中。另外有些物种,在停息于地上或草本植物上时,不时突然而轻微地抬起它们的翅。因此,某些蛾类翅的下表面比上表面的颜色鲜明就不会像最初看来那么令人感到异常了。天蚕蛾科中有些蛾是所有蛾类中最美丽的,它们的翅像英国的天蚕蛾那样,饰有漂亮的眼点;伍德先生 (237) 观察到它们的一些活动同蝶类相似:“例如,它们的翅好像为了夸示其美而轻轻起伏,昼出的鳞翅目昆虫比夜出的鳞翅目昆虫更具有这种夸示的特性。”
虽然许多颜色灿烂的蝶类的雌雄二者差异颇大,但根据我所能发现的,没有一种颜色灿烂的英国蛾类,其雌雄二者在颜色上有很大差异,几乎任何外国物种也是如此。然而,有一种美国蛾,叫做河神天蚕蛾(Saturnia Io)的,有人描述其雄者具有深黄色的前翅,其上奇妙地点缀着紫红色斑点;而雌蛾的双翅却是紫褐色的,点缀着灰色的线条。 (238) 雌雄颜色不同的英国蛾为全褐色,或为各式各样的暗黄色,或接近白色。有几个物种,其雄蛾的颜色比雌蛾暗得多, (239) 这些物种都属于一般在下午四处飞翔的类群。另一方面,正如斯坦登先生告诉我的,在许多属中,雄蛾的后翅要比雌蛾的白些——关于这个事实,鸣夜蛾(Agrotis exclamationis)提供了一个生动的实例。忽布蝙蝠蛾(Hepialus humuli)的这种差异更为显著;其雄蛾是白色的,雌蛾呈黄色并带有较暗的斑纹。 (240) 这些事例或可说明雄蛾像这样表现得较为显眼,大概是为了在黄昏四处飞翔时比较容易被雌蛾看到。
根据上述若干事实来看,不能认为蝶类和某些少数蛾类通常是为了保护自己而获得其灿烂的色彩。我们已经看到,它们所安排和展示的色彩和优雅样式好像都是为了夸示其美。因此,我被引导着去相信,雌者爱好颜色比较灿烂的雄者,要不后者最能使雌者激动;因为,根据我们所能知道的来说,若依其他设想,都无法说明雄者装饰的目的何在。我们知道,蚁类和某些鳃角类甲虫都能感到彼此依恋之情,而且蚁类在间隔数月之后还能认出它们的伙伴。因此,在系统上同这等昆虫大概居于差不多相等或完全相等位置上的鳞翅目,具有充分的精神能力来赞赏灿烂的色彩,在理论上并非是不可能的。鳞翅类肯定是凭借颜色来发现花的。常常可以见到蜂鸟天蛾(Humming-bird Sphinx)在若干距离以外向绿叶丛中一束花猛扑过去;有两位海外人士向我保证说,这等蛾曾反复光临一间屋子墙上画的花,而且徒劳地试图把它们的喙插进去。弗里茨·米勒告诉我说,巴西南部有几种蝴蝶准确无误地爱好某些颜色胜于爱好其他颜色:他观察到它们时常光临五六个植物属的灿烂红花,但从不光临同一花园里的同一属或其他属植物开白花或黄花的物种;我也收到过同样意义的其他报道。多勃尔德伊先生告诉我说,普通白蝶常飞向地面上的一块小片纸,无疑是把它错误地当做自己的同种。科林伍德(Collingwood)先生 (241) 当谈到在马来群岛收集某些蝶类的困难时说道,“把一只死标本钉在一条易见的小树枝上,往往会把一只正在匆忙飞翔中的同种昆虫引到捕虫网容易达到的范围之内,如果它同死标本的性别不同,就尤其容易用此法捕获”。
如上所述,蝶类的求偶是一个冗长的过程。有时雄蝶因竞争而互斗,也可看到有许多雄蝶在追逐同一只雌蝶或聚在其周围。那么,除非雌蛾喜爱某只雄蛾胜过其他雄蛾,否则雌雄配合必定完全委于机会,看来这似乎是不可能的。另一方面,如果雌蛾时常或者甚至偶尔选择更美丽的雄蛾,则后者的颜色将逐渐日益增其鲜明,按照普遍的遗传规律,这种颜色将传递给雌雄双方或只传给性别的一方。如果在第九章附录中根据种种证据所作出的结论是可信的话,即许多鳞翅目的雄虫数量至少在成虫状态时远远超过雌虫,则性选择的过程将会大大被推进。
然而有些事实同雌蝶喜爱比较美丽雄蝶的信念不相符合;例如,有几位昆虫采集者向我保证说,常常可以看到生气勃勃的雌蝶同伤损的、憔悴的或光彩暗淡的雄蝶交配;但这几乎总是由于雄虫早于雌虫出茧羽化的一种情况。关于蚕蛾科的蛾,其雄蛾和雌蛾一进入成虫状态即行交配;因为它们的口器处于线迹状态而无法取食。正如几位昆虫学家向我说的,雌蛾几乎处于麻木状态,对其配偶似乎毫不显示选择之意。欧洲大陆和英国的一些饲养家告诉我说,普通家蚕蛾(B.mori)就是这样情况。华莱士博士对饲养臭椿蚕(B.cynthia)有丰富的经验,他相信这种雌蛾毫无选择或偏爱的表示。他曾把三百只以上的这种蛾子饲养在一起,并且经常发现最强壮的雌蛾同发育不全的雄蛾相配。雄蛾对雌蛾好像并不如此;因为,正如他所信的,较强壮的雄蛾置衰弱的雌蛾而不顾,却被最富生命力的那些雌蛾所吸引。尽管如此,蚕蛾科虽颜色暗淡,但由于它们那优雅而复杂的色调,在我们看来往往还是美丽的。
迄今为止,我所谈到的只是雄虫颜色比雌虫更为鲜明的那些物种,并且我把雄虫的美丽归因于雌虫许多世代以来总是选择更有吸引力的雄虫,并与之交配。相反的事例虽然很少,但也是有的,即雌虫的颜色比雄虫更为灿烂;在这种情况下,正如我相信的雄虫所选择的乃是比较美丽的雌虫,因而使雌虫慢慢地增添了它们的美丽。在各个不同的动物纲中都有少数物种的雄者选择比较美丽的雌者,而不是心甘情愿地接受任何雌者,这似乎是动物界的普遍规律,不过为什么如此,我们还弄不清楚。但是,如果同鳞翅目所发生的一般情况相反,雌者数量远比雄者多得多,则雄者大概就可能挑选更美丽的雌者。巴特勒先生让我看过大英博物馆入藏的几个Callidryas的物种.其雌虫之美有和雄虫相等者,另外还有大大超过雄虫者;因为只有雌虫的翅缘才满布着艳红色和橙色,并具有黑色斑点。这些物种的雄虫色彩比较平淡,彼此密切相似,这说明在这里发生变异的是雌虫;而在雄虫更富有装饰的那些事例中,发生变异的乃是雄虫,雌虫则保持密切相似。
在英国我们看到一些类似的情况,尽管不是那么显著。蚬蝶(Thecla)属的两个物种只有雌虫在其前翅上具有一种鲜紫色或橙色块斑。草地褐蝶(Hipparchia)的雌雄颜色差别不大;但有一种复面草地褐蝶(H.janira),其雌虫在翅上有一种显著的鲜褐色块斑;另外有些物种的雌虫也比雄虫颜色更为鲜艳。还有,可食粉蝶(Colias edusa)与黄纹豆粉蝶(C.hyale)的雌虫“在其黑色翅缘上有橙色或黄色斑点,在雄虫方面只表现为细条纹”;在粉蝶(Pieris)属中,雌虫“在其前翅上装饰着黑色斑点,而这在雄虫的翅上只有部分表现”。那么,已知许多蝶类的雄者在其飞行结婚期间支撑雌者;但刚刚提到的那些物种却是雌者支撑雄者;因此雌雄双方所起的作用正好相反,这正如它们相对的美色也颠倒了一样。在整个动物界中,雄者在求偶过程中一般比较积极主动,由于雌者所接受的是吸引力较强的雄性个体,因而雄者的美似乎因此而增加了;但对这等蝶类来说,在最后的交尾仪式中乃是雌者居于比较积极主动的地位,所以我们可以设想它们在求偶过程中也起了同样的作用;在这样场合中,我们就能理解它们更加美丽的原因。上述系引自梅尔多拉(Meldola)先生的论述,他在总结时说道:“虽然我不相信昆虫的颜色是由性选择的作用而产生的,但不能否认这些事实明确地证实了达尔文先生的观点。” (242)
由于性选择主要依靠变异性,因此必须对这个问题稍作补充,关于颜色的变异性,并没有什么困难问题,因为可以举出任何数目的鳞翅目昆虫都是高度易变的。举一个明显的例子就够了。贝茨先生给我看过两种凤蝶(Papilio sesostris和P.childrenae)的一整套标本,后者的雄蝶在前翅的翡翠绿色美丽块斑的宽窄方面,在白斑的大小方面,以及后翅上艳红色的华丽条纹方面,都有很大变异;因此在最绚丽的雄蝶和最不绚丽的雄蝶之间形成了强烈对照。某种凤蝶(Papilio sesostris)的雄者远不及另一种凤蝶(P.childrenae)的雄者美丽;在其前翅上绿色斑块的大小以及后翅上偶尔出现的艳红色小条纹等方面也同样稍有变异,后一性状看来好像是来自本种的雌蝶;因为这个物种的雌蝶以及安尼阿斯蝶(Aeneas)类群其他许多物种的雌蝶都具有这种艳红色条纹。因此,在P.sesostris最鲜明的标本和P.childrenae最暗淡的标本之间只有一小段间隔,显然仅就变异性来说,凭借选择作用不断地给任何一个物种增添其美丽,并非困难之事。这里的变异性差不多只限于雄蝶;但华莱士先生和贝茨先生都曾指出 (243) ,有些物种的雌蝶是极易变异的,而雄蝶则几乎保持不变。我在后面一章将有机会说明许多鳞翅目昆虫的翅上美丽眼斑是显著易变的。这里我愿补充说明这些眼斑给性选择学说提供了一难点;因为在我们看来这些眼斑虽然非常富有装饰性,但从来没有只见于一种性别而不见于另一性别的,而且两种性别的眼斑也从来没有很大差异。 (244) 对这个事实目前还无法解释,但如果以后能发现眼斑的形成是由于翅膀组织有某种改变,譬如说,这种变化发生于很早的发育时期,则根据我们所知道的遗传规律,我们就可期待这种性状将会传递给雌雄双方,尽管它只在一种性别发生并完成。
总之,虽有许多严重的反对主张,但大多数鳞翅目的色彩鲜艳物种的颜色大概是由于性选择的结果,当然在即将谈到的某些事例中,其鲜明色彩的获得乃是由于作为保护的拟态。在整个动物界中,雄者因其热情一般都乐于接受任何雌者;而雌者则通常要尽力选择雄者。因此,如果性选择曾对鳞翅目发生过有效作用,则在雌雄相异的场合中,雄者的色彩应是更灿烂的,而事实无疑也正是如此。在雌雄双方都有着灿烂的色彩并彼此类似时,雄者所获得的这等性状大概传递给了雌雄双方。我们作出这个结论的根据是,即使在同一属中,雌雄二者之间在颜色上存在着从颜色差异非常之大到完全一样的等级。
但或可提问,关于雌雄二者之间的颜色差异,除性选择外,难道不能用其他方法来解释吗?例如,在若干事例中,据知同种的雄蝶和雌蝶栖息于不同的场所, (245) 前者一般曝于日光之下,后者则出没于幽暗的森林中。因此不同的生活条件可能会直接作用于雌雄二者;但看来这似乎不可能如此, (246) 因为处于成虫状态的雄蝶和雌蝶只是在很短时期内暴露于不同的生活条件下,而它们在幼虫期则都暴露于同样的生活条件下。华莱士先生相信,性别之间的差异,其原因在于雄者的变异并非那样多,而在所有或几乎所有的场合中大都是由于雌者为了保护自己而获得了阴暗的色彩。在我看来,情况正好相反,远为可能的是,通过性选择主要发生变化的正是雄者。雌者的变化是比较小的。这样,我们就能理解亲缘关系密切的雌者之间的类似为什么一般甚于雄者之间的类似。于是这些雌者向我们大致显示了它们所隶属的这个类群的亲种的原始色彩。然而由于某些连续变异传递给了雌者,它们几乎总得要或多或少地发生改变,而雄者正是通过这些连续变异的积累而增添了美丽。但是,我无意否定在某些物种中仅是雌者为了保护自己而发生了特别变异。在大多数情况下,不同物种的雄者和雌者在其漫长的幼虫状态中将暴露于不同的生活条件下并因此受到不同的影响;然而雄者由此发生的任何细微的颜色变化一般都要被性选择所引起的灿烂色彩所掩盖。在我们论到鸟类时,关于雌雄二者之间的颜色差异有多大程度是由于雄者为了装饰目的而通过性选择发生的变异,或者有多大程度是由于雌者为了保护自己而通过自然选择发生的变异,对这整个问题我还要加以讨论,因此在这里稍微谈一下就可以了。
如果雌雄二者同等遗传的方式更为普遍,在所有这种情况下,色彩鲜明的雄者的受到选择也倾向于引起雌者色彩变得鲜明;而色彩阴暗的雌者的受到选择也倾向于使雄者色彩变得阴暗。如果这两个过程同时进行,它们则倾向于相互发生作用,其最终结果将决定于如下情况:究竟是雌者由于暗淡的色彩而得到保护并占有多数,以成功地留下了更多的后代,还是雄者由于色彩鲜明并因此觅得配偶而占有多数,以成功地留下了更多的后代。
为了说明性状常常只传递给一种性别的现象,华莱士先生表示相信雌雄二者比较普通的同等遗传方式可以通过自然选择转变为只向一种性别遗传的方式,但我还没有支持这种观点的任何证据。根据在家养状况下所发生的情况,我们知道新性状常会出现,它们一开始只传递给一种性别;经过对这等变异的选择,只使雄者具有鲜明的色彩,同时或者随后只使雌者具有暗淡的色彩,这并没有一点困难。按照这种方式,某些蝶类和蛾类的雌者可能是为了保护自己而变得颜色暗淡并与其同种的雄者大有差异。
然而,如果没有明显的证据,我无论如何也不愿承认在大量的物种中有两个复杂的选择过程在进行,它们各自要求把新性状只传递给一种性别——即雄者靠着击败其竞争者而变得颜色更灿烂,而雌者由于逃避其敌害而变得颜色更暗淡。例如普通黄粉蝶(Gonepteryx)的雄者,其黄色比雌者强烈得多,虽然其雌者的黄色也是同等显著的;其雄者获得鲜明色彩可能是作为一种性的吸引,但是要说雌者是为了保护自己而特别获得了暗淡的色彩似乎是不大可能的。黄斑襟粉蝶(Anthocharis cardamines)的雌者不像雄者那样具有美丽的橙色翅尖;结果它同我们花园中常见的粉蝶密切类似;但我们还没有证据可以证明这种类似对它是有益的。另一方面,由于它同居住在世界不同地方的该属若干其他物种的雌雄二者都相类似,因而它可能只是在很大程度上保持了其原始的色彩。
最后,正如我们所见到的,不同的考察都引出了下面的结论,即对大多数色彩灿烂的鳞翅目昆虫来说,主要通过性选择发生变异的乃是雄者,雌雄二者之间的差异量大都决定于起作用的遗传方式。遗传是受到如此众多未知的法则和条件所支配的,其作用方式在我们看来是捉摸不定的; (247) 因此,在某种程度上我们可以理解为什么近亲物种的雌雄颜色或是惊人地不同或是完全一样。由于变异过程的所有后续步骤都必须通过雌者来传递,因而这类步骤就容易在雌者方面多少发展起来;于是我们就可以理解,在所有亲缘相近的物种中,雌雄二者从极端不同到毫无差别之间常常会出现一系列的级进。可以补充说明的是,这等级进的例子是如此普遍,以致不能支持我们在下述设想,即我们在这里所见到的雌者实际上经历了转变的过程,并为了保护自己而失去其鲜明色彩。因为我们有各种理由可以断定随便在任何时候大多数物种都是处于固定状态的。
拟 态
这一原理是由贝茨先生在一篇令人钦佩的论文中首次阐明的, (248) 因而对许多暖昧不明的问题提供了大量解释。以前曾有人观察过属于完全不同科的某些南美洲蝶类同长翅蛱蝶(Heliconidae)科在每一条纹和每一色调上如此密切类似,以致如果不是一位有经验的昆虫学家,就无法将它们区别开来。由于长翅蝶科所具者乃是本来的色彩,而别的蝶类则偏离了它们所属的类群的正常色彩,因此后者显然是模拟者,而长翅蛱蝶科则是被模拟者。贝茨先生进一步观察到,模拟的物种较少,而被模拟的物种则很多,这两组昆虫混在一起生活。长翅蝶科为颜色鲜明而美丽的昆虫,但它们的种和个体数量非常之多,他根据这个事实断定它们必定靠某种分泌物或气味来保护自己免受敌者的攻击;这一结论现已得到广泛的证实, (249) 尤其是得到了贝尔特先生的证实。因此,贝茨先生推论模拟有所保护的物种的蝶类通过变异和自然选择而获得了它们目前那种不可思议的伪装,以图被误认为是那些有所保护的种类而逃避被吞食的危险。这里只想说明一下模拟的蝶类的鲜明色彩,而对被模拟的蝶类的鲜明色彩则不做任何解释。我们对后者的色彩必须按照本章以前讨论的诸例所用的同样方式进行说明。自从贝茨先生的论文发表之后,华莱士先生在马来亚地区,特里门先生在南非,并且赖利先生在美国都观察到了同等显著的相似事实。 (250)
鉴于有些作者感到非常难于理解拟态过程的最初步骤如何能够通过自然选择而被完成,因此最好注意这一过程很久以前在颜色上并无显著差别的类型之间大概就开始了。在这种情况下,即使是一种轻微的变异大概也是有益的,如果这种变异能使其中一个物种更像另一个物种;以后被模拟的物种也许通过性选择或其他途径而被改变到极端的程度,如果这等变化是渐进的,则模拟者大概会容易地沿着同一轨道而随着变化,直至同它们原始状态的差别达到同等的极端程度;于是它们最终便获得了同它们所隶属的那个类群的其他成员完全不同的一种外貌或颜色。还应记住,鳞翅目的许多物种在颜色上都容易发生大量而突然的变异。本章已举出过少数例子,在贝茨先生和华莱士先生的论文中还可找到更多的例子。
有几个物种,其雄虫和雌虫是相似的,并且模拟另外一些物种的雌雄二者。但垂门先生在已经提到的那篇论文中举出三个例子,说明被模拟的类型的雌雄颜色彼此不同,而模拟的类型的雌雄颜色也以同样的方式而有所不同。还记载过若干事实,说明只有雌者才模拟色彩鲜艳的有所保护的物种,而雄者则保持“其直系同性的正常外貌”。在这里使雌者发生改变的连续变异显然只传递给了雌者一方,然而,在这许多连续变异中有些传递给了雄者并发展起来是可能的,如果不是获得这类变异的雄者因此失去对雌者的吸引力而被淘汰了的话;所以只有从一开始就严格限于传递给雌者的那些变异才会保持下来。贝尔特(Belt)先生在一项叙述中对这些意见曾作了部分证明, (251) 他说有的Leptalides的雄者在模拟有所保护的物种时仍以隐蔽的方式保留了它们一些原始状态。例如,雄虫“的下翅上半部是纯白色,其余部分则布满了黑的、红的以及黄的条斑和点斑,同它们所模拟的物种相似。雌虫则无此白色斑纹,雄虫通常用上翅将下翅掩盖起来而使它得以隐蔽,因此当它们把它展示于雌虫之前因而满足了后者对Leptalides所隶属的那一‘目,之正常颜色的根深蒂固的爱好时,我不能想象它有任何别的用处可以比得上作为求偶时的一种吸引”。
蠋的鲜明颜色
回顾许多蝶类的美丽,使我想起有些幼虫的颜色也是灿烂的;由于性选择在这里不可能起作用,因此,除非对其幼虫的鲜明色彩能以某种方式进行解释,否则把成年昆虫的美丽归因于性选择就未免轻率了。第一,可以看出蠋(幼虫)颜色同成年昆虫颜色并无任何密切的相关。第二,它们鲜明的颜色在任何正常意义上都不是作为保护之用的。贝茨先生向我说过这方面的一个例子:有一种天蛾幼虫生活于南美开阔的大草原(Ilanos)上一株树的大绿叶子上,这是他看到过的颜色最鲜明的一种幼虫,长约4英寸,横向有黑和黄的带斑,头、足和尾均呈鲜红色。凡是路过的人,即使在许多码以外都会看到它。至于每只过路的鸟无疑也会看到它。
于是我请教了华莱士先生,他是一位解决难题的天才。他考虑了一下回答说:“大多数幼虫都需要保护,这一点可从下述情况推论出来,即,有些种类的幼虫具有棘状突起或刺激性的毛,许多幼虫都是绿色的,同它们所取食的叶子颜色相似,或者同它们生活于其上的小树枝相似。”还有一个关于保护的事例,是曼塞尔·威勒(J.Mansel Weale)先生提供给我的,可以补充谈谈,即,有一种蛾的幼虫,生活于南非的含羞草上,把自己伪装得同其周围的棘刺完全区别不出来。根据这等考察,华莱士先生想象颜色显著的幼虫可能由于有一种恶味而得到保护的;但是,由于它们的皮极嫩,一受到创伤肠子就容易脱出,一只鸟只要用它的嘴轻轻把它们一啄,对它们来说就如同被吞食一样地可以致死。因此,正如华莱士先生所说的,“仅仅味道讨厌也许还不足于保护一只幼虫,除非有外在的标志向其可能的破坏者表示其牺牲品是一种味道恶劣的食物”。在这等情况下,一只幼虫能被所有鸟类和其他动物马上确定地认出它们是不好吃的,这也许对它非常有利。这样,最绚丽的颜色可能就有用了,并且通过变异和最易被识别的个体的生存而获得了这种颜色。
这一假说最初看来好像很大胆,但一把它提到昆虫学会, (252) 却受到了种种发言的支持。詹纳·韦尔先生在一个鸟舍里养过大量的鸟,他告诉我说,他做过许多试验,发现所有夜出日伏习性的而且表皮光滑的幼虫,所有绿色的幼虫,以及所有模拟树枝的幼虫,一概都被他养的鸟贪婪地吃掉,对此毫无例外。那些有毛的和有刺的种类一律都被拒而不食,四个颜色显著的物种亦复如此。当这些鸟拒绝不吃一种幼虫时,它们用摇头和擦净鸟喙来明确表示讨厌这种味道。 (253) 巴特勒先生也把三种颜色显著的幼虫和蛾子给一些蜥蜴和蛙吃,它们虽爱吃其他种类的幼虫和蛾子,但对这三种却拒而不食。因此,华莱士先生观点的可能性得到了证实,即,某些幼虫变得颜色显著乃是为了其自身的利益,以便使其敌害容易地把它们认出,这同药商把毒药装在有色瓶里出售乃是为了人类安全几乎是同样道理。然而我们现在还不能这样来解释许多幼虫多种多样的雅致颜色;但是,任何一个物种在某一既往时期如果由于模拟周围的物体或者由于气候的直接作用等等,而获得了暗淡的、具有点斑的或条纹的外貌,那么,当这个物种的色调变得强烈而鲜明时,它的颜色几乎肯定不会一致;因为,仅仅为了使一种幼虫易于辨认,大概不会按照任何一定方向进行选择的。
有关昆虫类的摘要和结论
就上述几个“目”来看,我们知道雌雄常常在种种性状上有所差异,但其意义一点也弄不明白。雌雄二者的感觉器官和运动手段也常有差异,因而雄者可以迅速发现雌者并抵达那里。雌雄二者更常见的区别还有:雄者具有种种构造以抱持所找到的雌者。然而,我们这里所涉及的这些种类的雌雄差异仅仅是次要的方面而已。
在几乎所有的“目”中,有些物种的雄者,即使是娇弱的种类,据知也是高度好斗的;有少数物种具有特殊的武器,同其竞争者战斗。但战斗之规律在昆虫中几乎不像在高等动物中那样普遍。因此,可能只有在少数场合中,才会发生雄虫变得比雌虫更大更强壮。相反,它们通常都小于雌虫,以便在较短时间内可以完成发育,而为雌虫的出现准备下大量雄虫。
在同翅目的两个科、直翅目的三个科中,只有雄虫才具备有效状态的发音器官。这些器官在繁育季节不停地使用,这不仅是为了召唤雌虫,显然也是为了同其他雄虫竞赛以诱惑和刺激雌虫。凡是承认有任何种类的选择作用的人在读了上述讨论后,都不会再争论这些音乐器官是通过性选择而获得的。在其他四个目中,一种性别的成员,更普通的是雌雄双方的成员都具有可以发出各种声音的器官,这些声音显然只是作为呼唤之用。当雌雄双方都具有这类器官时,声音最高的或叫得最持久的个体也许比那些声音较低的个体先找到配偶,所以它们的器官大概是通过性选择而获得的。仔细想一想或者仅仅雄虫或者雌雄双方具有多样发音手段者不少于六个目,是会有所启发的。我们由此可以认识到性选择如何有效地引起了变异,这等变异如在同翅目中那样,同体制的重要部分是有关联的。
根据上一章所举出的原因,许多鳃角组昆虫以及某些其他甲虫,其雄者所具有的巨角大概是作为装饰物而获得的。因昆虫的体积小,我们就容易低估了其外貌。如果我们能想象披着一身亮光闪闪的青铜铠甲,而且长着复杂的大角的雄性独角仙类(Chalcosoma)(p200,图16)放大到一匹马、哪怕是一只狗那样大小,那么它也许是世界上最壮观的动物之一。
昆虫的颜色是一个复杂而暧昧不明的问题。当雄虫同雌虫的差别轻微而且双方颜色都不鲜艳时,这可能是由于雌雄二者在差别轻微的途径上进行变异的,而且这等变异各向其自身性别那一方传递下去,并不因此带来任何好处也不带来任何害处。当雄虫的颜色鲜艳并同雌虫差别显著时,如某些蜻蜓类和许多蝶类那样,那么雄虫的颜色可能是由于性选择所致;如果雌虫保持一种原始的或很古老形式的颜色,则由于上述作用所发生的变异就是轻微的了。但在某些场合中,雌虫显然由于单独传递给它的变异而变得颜色暗淡,以作为直接保护自己的一种手段;几乎肯定的是,有时雌虫也会变得颜色鲜艳,以模拟栖息在同一地区的其他具有保护性的物种。当雌雄二者彼此相似而颜色又都暗淡时,在很多场合中它们的颜色之所以变得这样,无疑是为了保护自己。当雌雄二者都具有鲜明颜色时,有些情况也同上述一样,因为它们这样来模拟具有保护性的物种,或同周围的物体、如花朵等相类似;或使其敌害注意到它们是不好吃的。在另外一些场合中,雌雄二者彼此相似又都颜色鲜艳,尤其是它们的颜色如果是为了夸耀,我们就可断定雄虫获得的这种颜色乃是作为一种性吸引,并把这种颜色传递给了雌虫。任何时候当在整个类群中普遍存在着同样的颜色形式时,而且我们如果发现有些物种的雄虫和雌虫的颜色差别很大,而另外一些物种的雌雄颜色差别很小或完全没有差别,并且有中间的级进把这两个极端的状态连接起来,那么,这等情况尤其足以引导我们作出上述结论。
正如鲜明的颜色常常由雄者部分地传给雌者一样,许多鳃角组昆虫和某些其他甲虫异常巨大的角也是如此。还有,同翅目和直翅目雄虫所特有的发音器一般以残迹(退化)状态甚至以几乎完善的状态传递给雌虫,但没有完善到有任何用途。还有一个同性选择有关的有趣事实,即,某种同翅目雄虫的摩擦发音器要到蜕最后一次皮时才完全发育,某些雄蜻蜓的颜色也要在从蛹羽化后不久并准备繁育时才完全发育。
性选择意味着,为异性所喜爱的是更富吸引力的个体;而对昆虫来说,当雌雄二者有所差异时,除少数例外,更富装饰的以及偏离这个物种所隶属的那种模式更远的,乃是雄虫——因为正是雄虫热切地寻求雌虫,所以我们必须假定雌虫常常地或者偶尔地偏爱更美丽的雄虫,因而后者获得了它们的美。雄虫具有许多抱持雌虫的独特装置,如巨颚,黏着垫、刺、延伸的腿等,用以抓住雌虫,从这等情况看来,在大多数或所有的“目”中,雌性昆虫可能具有拒绝任何特殊雄虫的能力;因这些装置表明在求偶活动中还有某种困难,所以雌者的同意似乎是必要的。根据我们所了解的各种昆虫的知觉能力和爱情来判断,认为性选择曾起了很大作用并无不可信之理。但关于这个问题我们还没有直接的证据,而且还有些事实同这个信念正相抵触。尽管如此,当我们见到许多雄虫追逐着同一只雌虫时,我们还是几乎无法相信这种配合是完全由于盲目的机会——即雌虫既不尽力实行选择,也不受雄虫的华丽颜色或它们所具有的其他装饰物所影响。
如果我们承认同翅目和直翅目的雌虫欣赏其雄性配偶的和谐音调,而且它们的各种发音器官是通过性选择而完成的,那么其他雌性昆虫欣赏形状上颜色上的美丽,并因而导致雄虫获得这类性状,看来也很少有不可信之理。但由于颜色非常易变,并且由于为了保护自己颜色如此经常地发生改变,所以难于断定性选择所起的作用究竟占有多大比率。这一点在直翅目、膜翅目以及鞘翅目的那些“目”中尤其难于得到断定,它们的雌雄二者在颜色上很少有重大差异。因为在这里除类推法外并无其他方法可循。如上所述,在鞘翅目中有一鳃角大类群,某些作者把它置于该目之首,我们时常看到其中雌雄之间的相互依恋,我们还发现某些物种的雄虫为了占有雌虫的斗争而具有武器,另外一些雄虫则具有异常大的角,许多雄虫具有摩擦发音器官,其他则装饰着辉煌的金属色泽。因此,所有这等性状大概都是通过同样的途径、即性选择而获得的。至于蝶类,我们则掌握有最好的证据,因为雄蝶有时尽力夸示其美丽的颜色;这种夸示除非是用于求偶,否则我们就无法相信它们为什么要这样做。
当我们讨论鸟类时,我们将看到它们在其次级性征方面同昆虫类是最相似的。于是,许多雄鸟都非常好斗,有些还具有特别的武器,同其竞争者战斗。它们还具有在繁育季节中用来发出声乐和器乐的器官。它们往往装饰着各式各样的肉冠,角状物,垂肉和羽毛,并且还具有美丽的颜色,这些显然都为了夸耀其美。我们将会发现,正如昆虫类那样,有某些群的雌雄二者都同样美丽,都同样地具有一般限于雄虫才有的那种装饰物。在其他类群中,雌雄二者则都颜色平淡,且无装饰。最后,还有某些少数异常的例子,表明雌者反比雄者更为美丽。我们将会常常发现,在同一个鸟的类群中,从雌雄之间毫无差异到雌雄之间具有极端差异,其间有各种等级。我们还会看到,母鸟像雌性昆虫那样,往往或多或少具有一些雄者所固有的并只对它们有用的那种性状,但这等性状是残迹的、即不发育的。确实,在鸟类和昆虫类之间所有这些方面的相似性是非常密切的。能应用于一个纲的无论什么解释大概也能应用于另一个纲,而这种解释,正如以后我们将进一步详细加以阐明的,就是性选择。
第十二章 鱼类、两栖类和爬行类的第二性征
鱼 类
雄者的求偶和争夺——雌者的较大体型——雄者,鲜明色彩和作为装饰的附器;其他奇特的性状——雄者只在繁殖季节才获得色彩和附器——雌雄二者均具鲜艳色彩的鱼类——保护色——不能根据保护原理来说明雌者色彩的较不显著——雄鱼筑巢并照顾卵和幼鱼。两栖类:雌雄二者在构造和色彩上的差异——发音器官。爬行类:龟类——鳄类——蛇类,颜色有时是保护性的——蜥蜴类及其斗争——作为装饰的附器——雌雄二者在构造上的奇特差异——色彩——雌雄差异之大几乎与鸟类相同
我们现在讨论的是脊椎动物门的大亚界,先从最低等的纲、即鱼类开始。横口鱼类(Plagiostomous fishes,鲨类,鳐鱼类)和银鲛类(Chimaeroid fishes)的雄者都有用以守住雌者的鳍脚,就像许多低等动物所具有的各种这样构造一样。除鳍脚外,许多红鱼类的雄者在其头部都生有坚固锐利的刺丛,沿着“它们胸鳍的上部外表面”也有数行。有些物种的雄者生有这种刺丛,而其体躯的其余部分则是光滑的。刺丛只在繁殖季节才临时发育起来,京特(Günther)博士怀疑它们靠着把躯体两侧向内和向下弯曲而起抱握器官那样的作用。有一个值得注意的事实,即,有些物种,如刺背鳐鱼(Raia clavata),其背上生有钩状大刺者为雌鱼而非雄鱼。 (254)
毛鳞鱼(Mallotus villosus,鲑科的一种)只有雄鱼才具有一条密集的毛刷状鳞隆起,当雌鱼在海滨沙滩飞泳和产卵时,有两个雄鱼凭借毛刷状鳞隆起之助各在一边以挟持之。 (255) 和毛鳞鱼大不相同的毡毛单角魨(Monacanthus scopas)也有一种多少相似的构造。正如京特博士向我说的,其雄者在尾部两侧生有一团肉冠似的坚硬直刺;在一个6英寸长的标本身上这种刺约为1.5英寸;雌鱼在同一部位则生有一簇硬毛,可同牙刷的硬毛相比拟。还有一种魨鱼(M.peronii),其雄鱼生有的那种毛刷同后一物种的雌鱼所生的那种毛刷相似,而雌鱼尾部两侧则是光滑的。在同一属的某些其他物种中,可以看出其雄鱼尾部稍现粗糙,而雌鱼的尾部则完全是光滑的;最后,在其他物种中,则雌雄二者的尾部两侧都是光滑的。
许多鱼类的雄者都要为占有雌者而斗争。例如,有人描写光尾刺鱼(Gasterosteus leiurus)的雄者当雌者跑出其隐藏处来到前者为雌鱼做好的巢进行观察时,表现得“欣喜欲狂”。“它在雌鱼的周围钻来钻去,又钻到储备在巢里的物资那里,马上又折回雌鱼这里,当雌鱼不前进时,它就试图用吻去推雌鱼,然后又用尾巴和边刺试着把雌鱼推进巢里”。 (256) 据说雄鱼是多配性的, (257) 它们特别勇敢而好斗,而“雌鱼则都十分温和”。雄鱼之间的斗争常常是不顾死活的,“因为这些短小的斗士紧紧地互相缠住达数秒钟之久,翻过来滚过去,一直到它们显得体力已完全耗尽为止”。至于尾部粗糙的刺鱼(G.trachurus),其雄鱼在互斗中绕来绕去,相互撕咬并以竖起之侧刺试图把对方刺穿。这同一位作者接着说,“这些狂暴的小东西撕咬起来很凶猛。它们还运用其侧刺造成如此致命的效果,以致我曾见到在一次战斗中有一条雄鱼确实把其对手完全撕开使之沉下水底而死去”。当有一条鱼被征服时,“即行停止再向雌鱼献殷勤;它的华丽颜色随之减退;忍辱于其安静的伙伴之中,但在若干时间内它还是征服者所经常迫害的对象。” (258)
雄鲑鱼同小刺鱼一样好斗,我听京特博士说,雄鳟鱼(trout)也是如此。肖(Shaw)先生见过两条雄鲑鱼的一次激烈斗争持续了整整一天,渔场监督布伊斯特(Buist)先生也告诉我说,他从珀思(Perth)的桥上常常看到在雌鱼产卵时,雄鱼把它的竞争者赶走。这些雄鱼“总是在产卵床上互相厮打不已,许多受伤严重而造成相当的死亡,还有许多在体力竭尽的状态下在岸边游动,显然已处于垂死之中”。 (259) 彪斯特先生告诉我说,1868年6月斯托蒙特菲尔德养鱼场的管理员访问了泰恩河北段,发现有300条死鲑鱼,其中只有一条雌鱼,其余都是雄鱼,他相信它们是在厮斗中丧生的。
雄鲑鱼最奇异之处是,它们在生殖季节,除色彩出现轻微变化外,“下颚延长并在颚端生出一个朝上翻卷的软骨突起,当上下颚闭合时,该突起就占满了上颚颚间骨之间的那个深腔”。 (260) (图27和图28)英国鲑鱼这等构造的变化只发生在繁殖季节;但洛德(J.K.Lord) (261) 先生认为,在美洲西北部所产的一种狼鲑(Salmo lycaodon)这种变化却是永久性的,并且以前溯游到河里来的那些较老的雄鱼表现得最为显著。这些老雄鱼的下颚已经发展为一个巨大的钩状突起,上面的尖齿生长规则,长度往往超过半时。按照劳埃德 (262) 先生的见解,当一条雄欧洲鲑鱼猛攻另一条雄鱼时,这种临时性的钩状构造既加强了鱼嘴的力量,也为它提供了防护;但美洲雄鲑鱼极其发达的牙齿则可以同许多雄性哺乳动物的獠牙相比拟,这意味着这等尖牙与其说是为了防护的目的,倒不如说是为了进攻的目的更为恰当。
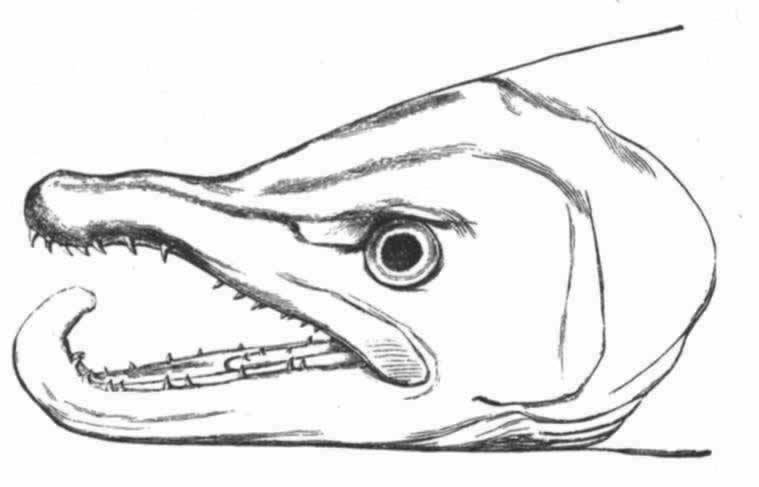
图 27 在生殖季节普通雄鲑鱼(Salmo salar)头部
(这幅图以及本章其他插图都是由著名画家G.福特先生在京特博士诚恳指导下根据大英博物馆的标本画出的)
图 28 雌鲑鱼头部
鲑鱼不是雌雄牙齿相异的唯一鱼类,许多魟鱼也是如此。成年雄刺背鳐鱼(Raia clavata)具有朝后长的尖锐牙齿,而雌鱼的牙齿则阔而平,有如铺石路面;所以同一物种雌雄二者牙齿的这等差异要比同科不同属之间的通常差异更甚。雄鱼的牙齿要到成年时才变得尖锐,它们的牙齿在幼小时就像雌鱼那样是阔而平的。正如第二性征所屡屡发生的情况那样,鳐鱼类的某些物种(如R.batis)在成年时,其雌雄二者都具有尖利的牙齿;在这里,为雄者所固有并首先为它所获得的一种性状似乎传递给了雌雄双方的后代。斑鳐(R.maculata)雌雄二者的牙齿同样也是尖形的,但只在它们完全成熟时才如此;而雄鱼获得这种性状的时期则早于雌鱼。关于某些鸟类,俟后我们还会遇到相似的情况,即雄鸟获得雌雄二者在成熟时所共有的羽衣的时期,多少要比雌鸟为早。至于魟鱼类的其他物种,即使雄鱼在年老的时候也从不生长尖锐的牙齿,因而其成年的雌雄二者的牙齿都同它们幼小时一样,也同上述物种的成熟雌鱼的牙齿一样,长得阔而平。 (263) 由于鳐鱼是一种勇敢、强壮而贪婪的鱼类,我们可以想象其雄鱼之所以需要尖锐的牙齿乃是为了同其竞争对手进行争斗;但由于它们具有已发生变异而适于抱握雌鱼的许多部分,所以它们的牙齿也可能用于这一目的。
关于鱼的大小,卡邦尼尔 (264) 主张几乎所有鱼类的雌者都大于雄者;京特博士也不知道有任何一个事例可以说明雄者确比雌者为大。至于某些鳉类(Cyprinodonts),其雄者的大小甚至不及雌者的一半。正因为许多鱼类的雄者经常互斗,它们没有通过性选择的作用变得比雌鱼大而壮,确令人感到诧异。根据卡邦尼尔的意见,雄鱼因体形小而蒙受损害,这个物种若是肉食性的,那么它们就容易被本种的雌鱼所吞食,也无疑会被别种鱼所吞食。雄鱼需要力强体大以与其他雄鱼进行争斗,而雌鱼体形的增大,在某种意义上其重要性必然大于此者。这也许是为了可以大量产卵之故。
在许多物种中,只有雄鱼才饰有鲜明的色彩;或者说,雄鱼的色彩比雌鱼鲜明得多。有时,雄鱼也具有附器,但它们对雄鱼的正常生活用途并不比尾羽对孔雀更大。我感谢京特博士的善意,他向我提供了大部分的下述事实。有理由猜想许多热带鱼类的雌雄二者在颜色和构造上都有差异,英国鱼类在这方面就有一些显著的事例。雄 (Callionymus lyra)“因其宝石般的鲜艳色彩”,而有宝石
(Callionymus lyra)“因其宝石般的鲜艳色彩”,而有宝石 (gemmeous dragonet)之称。从海里捕获的这种活鱼,其躯体带有各种浓淡不同的黄色,其头部具有亮蓝色的条纹和斑点;其背鳍呈淡褐色并有暗色纵带斑;其腹鳍、尾鳍和臀鳍均呈蓝黑色。其雌鱼或称泥色
(gemmeous dragonet)之称。从海里捕获的这种活鱼,其躯体带有各种浓淡不同的黄色,其头部具有亮蓝色的条纹和斑点;其背鳍呈淡褐色并有暗色纵带斑;其腹鳍、尾鳍和臀鳍均呈蓝黑色。其雌鱼或称泥色 (sordid dragonet),曾被林奈以及其后的许多博物学者当做一个不同的物种;雌鱼们的颜色是红褐的,没有光泽,背鳍呈褐色,其他诸鳍则为白色。雌雄二者在头部和嘴部的大小比例上以及眼睛着生的位置上也有差异; (265) 但最显著的差异还在于雄者背鳍的特别延长(图29)。萨维尔·肯特(W.Saville Kent)先生说道,“我就圈养鱼种所作的观察得知,这等独特的附器同鹑鸡类雄者的垂肉、羽冠以及其他异常附器所从属的目的一样,都是为了雄者向其配偶献媚之用”。 (266) 幼小雄鱼在构造和颜色上都同成年雌鱼相似。在整个
(sordid dragonet),曾被林奈以及其后的许多博物学者当做一个不同的物种;雌鱼们的颜色是红褐的,没有光泽,背鳍呈褐色,其他诸鳍则为白色。雌雄二者在头部和嘴部的大小比例上以及眼睛着生的位置上也有差异; (265) 但最显著的差异还在于雄者背鳍的特别延长(图29)。萨维尔·肯特(W.Saville Kent)先生说道,“我就圈养鱼种所作的观察得知,这等独特的附器同鹑鸡类雄者的垂肉、羽冠以及其他异常附器所从属的目的一样,都是为了雄者向其配偶献媚之用”。 (266) 幼小雄鱼在构造和颜色上都同成年雌鱼相似。在整个 属 (267) 中,雄鱼的斑点一般远比雌鱼的鲜明得多,还有几个物种,其雄鱼不仅背鳍长得多,而且臀鳍也长得多。
属 (267) 中,雄鱼的斑点一般远比雌鱼的鲜明得多,还有几个物种,其雄鱼不仅背鳍长得多,而且臀鳍也长得多。
图 29  (Callion ymus lyra)
(Callion ymus lyra)
上图为雄鱼;下图为雌鱼。
注意:下图较上图更为缩小。
蠍杜父鱼(Cottus scorpius)亦称海蝎鱼(sea-scorpion),其雄者比雌者细而小。它们之间在色彩上也有巨大差异。正如劳埃得先生 (268) 所说的,“当产卵时这种鱼的色彩极为鲜艳,任何人要是没有见过这种情况,他将难于想象这种混合的鲜艳色彩正是在那时装饰起来的,而在其他方面这种鱼并没有任何美丽之处。”杂种隆头鱼(Labrus mixtus)的雌雄二者虽在色彩上很不相同,但都是美丽的;其雄者呈橙色并带有亮蓝色的条纹,雌者呈鲜红色,背上有一些黑斑点。
在大不相同的鳉科中——国外的淡水鱼类——雌雄二者在种种性状上有时差异很大。黑帆鳉(Mollienesia petenensis) (269) 雄者的背鳍极其发达,其上有一行颜色鲜明的大而圆的眼状斑;而雌者的背鳍却较小,形状也不同,其上只有不规则的曲线形的褐色斑点。雄者臀鳍的底边稍有延长,且色暗。一个亲缘关系相近的类型叫做剑尾鱼(Xiphophorus Hellerii)(图30),其雄者尾鳍的下缘发展成一条长的丝状物,正如京特先生对我述说的,其上有色彩鲜明的线条。这等丝状物不含任何肌肉,显然它对这种鱼不会有任何直接的用处。同 属情况一样,其雄鱼当幼小时在色彩和构造上都同成年雌鱼相似。这类性差异可以严格地同鹑鸡类如此常见的性差异相比拟。 (270)
属情况一样,其雄鱼当幼小时在色彩和构造上都同成年雌鱼相似。这类性差异可以严格地同鹑鸡类如此常见的性差异相比拟。 (270)

图 30 剑尾鱼(Xiphophorus Hellerii)
上图为雄鱼;下图为雌鱼。
产于南美洲淡水中的一种鲇鱼叫做有须鲇鱼(Plecostomus barbatus (271) (图31),其雄者的嘴和内鳃盖骨边缘布满硬毛胡须,而这在雌者方面连一点痕迹也没有。这等硬毛具有鳞片的性质。同属的另一个鱼种,从其雄者的头前部伸出一些柔软易弯的触须,而雌者则缺如。这等触须乃是真皮的延长物,因此和上述鱼种的硬毛不是同源;然而,几乎无法怀疑它们都是用于同一目的。至于这个目的究竟是什么,就难于猜测了;在这里它们似乎不可能是用做装饰,但我们几乎无法设想这等硬毛和易弯的丝状物只对雄者有任何正常的用途。那个叫做怪银鲛(Chimaera monstrosa)的陌生怪物,其雄者头顶长出一个钩形骨指向前方,其顶端变圆且为锐刺所覆盖;而雌者则完全没有这等冠饰,但这等冠饰对雄者可能有什么用途,则全属未明。 (272)
到目前为止所谈到的那些构造都是在雄鱼到达成熟后才成为永久性的;但在鳚属(Blennies)以及其他亲缘相近的属中, (273) 有些种类只在繁殖季节,其雄鱼头上的冠饰才发育起来,而且其躯体的色彩也同时变得更加鲜艳。此等冠饰只是作为一种临时性的装饰,看来很少有疑问,因为雌鱼丝毫没有呈现这种痕迹。同属的其他物种,其雌雄二者均有冠饰,至少有一个物种其雌雄二者都不具有这样构造。阿加西斯教授对我说 (274) ,在雀鲷科(Chromidae)的许多鱼类中,如Geophagus,尤其是丽鱼(Cichla),其雄者在前额上有一显著的突起部,而在雌者和幼小雄者的前额上则完全缺如。阿加西斯教授接着说,“我常常见到这些鱼类前额突起部在产卵季节为最大,而在其他季节则完全消失,以致雌雄二者这时从头部侧面轮廓看来一点也显不出什么差别。我丝毫不能确定它对任何特殊机能有何帮助,亚马孙河上的印第安人对它的用途也一无所知”。此等突起部在定期出现方面同某些鸟类头上的肉瘤相似,但它们是否用做装饰,目前仍有疑问。

图 31 有须鲇鱼(Plecostomus barbatus)
上图为雄鱼的头;下图为雌鱼的头。
阿加西斯教授和京特博士对我说,有些鱼类的雄者在色彩上同雌者永不相同,这些鱼类往往在繁殖季节就会变得更加鲜艳。还有大量的鱼类也是如此,其雌雄二者的色彩在繁殖以外的所有季节里都完全相同。可举出丁鱥(tench)、拟鲤(roach)和鲈鱼(perch)为例。雄鲑鱼当此繁殖季节,“其双颊呈现橙色条纹,这使它具有隆头鱼(Labrus)的外观,其周身也现金橙色。而雌者的颜色则是暗黑的,故通常称为黑鱼”。 (275) 大南乳鱼(Salmo criox)雄者所发生的变化也与此类似,甚至更大;湖红点鳟(S.umbla)雄者的色彩当此繁殖季节同样也比雌者更为鲜明。 (276) 美国网纹狗鱼(Esox reticulatus)的色彩,尤其是雄者的色彩,在繁殖季节变得极其浓烈、鲜艳和富于虹彩。 (277) 雄光尾刺鱼(Gasterosleus leiurus)是这许多显著事例中的又一个,沃林顿(Warington)先生 (278) 描述它在繁殖季节所表现的“美丽非笔墨所可形容”。而雌鱼的背和眼的颜色单调,均为褐色,腹部白色。反之,雄刺鱼的眼乃“最艳丽的绿色,且具金属光泽,如同某些蜂鸟的绿羽。其喉部和腹部均呈鲜明的艳红色,背部为灰绿色,整条鱼看来多少有点透明,犹如体内有个白热的光源在闪闪发亮”。繁殖季节一过,这等色彩就全变了,其喉部和腹部的红色变淡了,背部的绿色变深了,发亮的色调消失了。
关于鱼类求偶的问题,自本书第一版问世以来,除已举出的刺鱼事例外,又观察到了一些其他例子。正如我们所知道的,杂种隆头鱼的雌雄二者在色彩上互不相同,肯特(W.S.Kent)先生说,这种雄鱼“在池塘沙滩做好一深穴,然后不厌其烦地极力诱使同种的一条雌鱼与之同居,它在雌鱼和已竣工的新居之间游来游去,对于雌鱼的依从显然表示了最大的热望”。海管鱼(Cantharus lineatus)的雄者当繁殖季节,则呈深铅黑色,然后离开鱼群并挖穴为巢。“现在每条雄鱼均走上警卫各自巢穴的岗位,并向任何其他雄鱼猛烈进攻,把它们赶走。然而对其异性的配偶,雄鱼的行为就大不相同。许多雌鱼眼下因怀卵而躯体膨大,雄鱼便以力所能及的各种办法,逐一地把雌鱼极力引诱至它所准备好的巢穴,以便把它们满怀的无数鱼卵产在那里,然后,雄鱼就以最大的小心保护和守卫着这些鱼卵”。 (279)
卡邦尼尔举出一种中国红鲤(Chinese macropus)雄者求偶以及夸示其美的显著事例。他对这等圈养中的鱼类进行了仔细观察。 (280) 雄鱼的色彩最为漂亮,比雌鱼要美得多。它们在繁殖季节为占有雌鱼而争斗;雄鱼在求偶行动中把鳍展开,鳍上具有斑点并饰以色彩鲜明的鳍刺,据卡邦尼尔说,其方式有如孔雀开屏一样。然后它们非常活泼地在雌者周围窜来窜去,炫耀“其鲜艳动人之色彩以便吸引雌鱼的注意,而雌鱼对雄鱼的这等动作也未尝不感兴趣,于是它们慢慢随着雄鱼一起游去,似乎很乐于同雄鱼作伴而待在一起”。雄者赢得了其配偶之后,就从嘴里吹出空气和黏液形成一个小泡沫盘。接着它把雌鱼产的受精卵集拢在嘴里,这一现象曾使卡邦尼尔大为吃惊,以为这些卵要被雄鱼所吞食。但实际不然,雄鱼迅速地把卵附着在泡沫盘中,然后就守卫鱼卵、修补泡沫并对孵化的幼鱼进行照顾。我之所以要陈述这些细节,是由于我们马上就要见到有些鱼类的雄者是把卵搁在嘴里孵化的;那些不相信逐渐进化原理(the principle of gradual evolution)的人们大概要问这样一种习性何由而始;但是,我们如果知道某些鱼类是像上述那样收集和携带鱼卵的,那么这个难题就很容易理解了;因为,要是把卵存放于泡沫中的这件事因任何一种原因而受到耽搁,那么,也许就会获得把卵放在嘴里孵化的这种习性。
让我们回到更为直接的主题上来。情况是这样摆着:就我所知,如无雄鱼在场,雌鱼决不愿意产卵;而如无雌鱼在场,雄鱼也决不给卵授精。雄鱼为占有雌鱼而互相争斗。有许多物种,其雄鱼当幼小时在颜色上同雌鱼相似;但一到成年,就变得比雌鱼鲜艳得多,而且始终保持着这种颜色。另外有些物种,其雄鱼的颜色只在求偶季节才变得比雌鱼鲜艳,或者更富于装饰。雄鱼孜孜不倦地向雌鱼求偶,有一个例子,像我们所见过的,雄鱼卖力地在雌鱼面前夸示其美色。难道能认为它们在求偶过程中的这等行为是毫无目的的吗?若有目的,则雌鱼一定要尽力进行某种选择,而且选取它们最中意的或使它们最受刺激的那些雄鱼。如果雌鱼尽力进行这等选择,那么所有上述有关雄鱼装饰的事实就可以借性选择之助立刻得到说明。
其次我们势必要追问,某些雄鱼通过性选择获得其鲜明色彩的这个观点,能否依据性状向雌雄双方同等传递的法则,引申到雌雄二者色彩鲜艳的程度和式样都相同或几乎相同的那些类群上去。像隆头鱼这样的一个属,它含有世界上一些最绚丽的鱼类——以孔雀隆头鱼(L.pavo)为例,有人用可以谅解的夸张手法描绘说, (281) 它的鳞片是用闪闪发亮的黄金制成的,周身还镶饰着天青石、红宝石、蓝宝石、翡翠和紫水晶——关于这个属,我们多半可以接受上述说法;因为我们已经知道,该属至少有一个物种的雌雄二者在色彩上是大不相同的。至于有些鱼类,正如许多低等动物那样,其华丽的色彩可能是其组织性质以及环境条件的直接结果,而并未借助于任何种类的选择。金鱼(Cyprinus auratus),根据普通鲤鱼的金色变种来类推,也许是一个恰当的例子,因为它的华丽色彩大概是由一种单纯的突然变异所形成的,而这种突然变异乃是它所处的圈养条件引起的。然而,更为可能的是这等颜色是通过人工选择而被加强的,因为从遥远的古代这个物种在中国就被精心培育出来了。 (282) 在自然条件下,像鱼类这样高级体制的动物,生活于如此复杂的关系中,要说从如此一种巨大的变化中既没有受到某种祸害也没有得到某种益处,因而没有受到自然选择的干预,它们就会变得色彩鲜艳,看来是不大可能的。
那么,对于雌雄色彩都华丽的许多鱼类,我们将作出怎样的结论呢?华莱士先生认为 (283) ,有些物种常常出没于礁石之间,那里富有珊瑚虫和其他色彩鲜明的有机体,因而这等物种的色彩也变得鲜明,以免被其敌害发现;但根据我的回忆,它们却因此表现得极其显眼。在热带的淡水中并无色彩鲜艳的珊瑚虫或其他有机体可供鱼类模仿;但亚马孙河的许多鱼种却有着美丽的色彩,印度的肉食性鲤科有许多鱼类也饰有“各种色彩鲜明的纵线条”。 (284) 麦克莱兰(M'Clelland)先生在描写这些鱼类时竟离奇地设想“其色彩之特别鲜艳”是给“那些注定要来抑制其数量增殖的翠鸟、燕鸥以及其他鸟类提供一种较易识别的标志”;然而今日博物学家们很少承认任何动物之所以变得显眼乃是为了加速其自身的毁灭。某些鱼类变得显眼可能是为了警告那些猛禽和猛兽,表示它们是不好吃的,就像我们讨论鳞翅目幼虫时所阐明了的那样;但我相信,还不知道有任何一种鱼,至少是任何一种淡水鱼,因味道不好而被食鱼的动物所拒绝。总之,关于雌雄色彩都鲜艳的鱼类,最近理的观点是,这等色彩作为一种性的装饰先由雄者所获得,而后把它同等地或几乎同等地传递给了雌者。
我们现在必须考虑的是,当雄者在色彩上或其他装饰上以显著的方式同雌者有所差异时,是否只是雄者发生了变异,而且这等变异只遗传给其雄性后代;或者,雌者发生了特殊变异,为了保护自己而变得颜色暗淡,这等变异是否只遗传给雌者。无可怀疑,许多鱼类所获得的色彩乃是作为一种保护;凡是察看过比目鱼(flounder)斑点累累上部表面,谁都无法忽视这等表面同它所栖息于其上的海底砂床的相似性。再者,某些鱼类能通过其神经系统的作用在短时间内改变其颜色以同周围物体相适应。 (285) 关于利用其颜色以及利用其形状来保护自己的动物,见诸记载的最显著事例之一(根据保藏的标本所能判断的来说),是由京特博士举出的, (286) 即,具有红光四射的丝状体的海龙(pipe-fish)同海草之间几乎无法加以区别,而它是用抱握性的尾部缠附于海草之上的。但现在所要考虑的问题在于是否只有雌鱼才为了保护的目的而发生变异。我们可以知道,假定雌雄双方为了保护的目的通过自然选择都发生了变异,那么任何一方所发生的变异绝不会超过另一方。除非是其中一方暴露于危险之中的期间比另一方较长,或者逃避这样危险的能力比另一方较差;然而在这些方面鱼类的雌雄二者看来并没有什么差别。如果说有任何差别的话,充其量也无非是雄鱼因其体形一般较小,而且较常游动,所以比雌鱼所面临的危险较大而已;尽管如此,一旦雌雄之间出现差异时,几乎总是雄鱼的色彩更为显著。鱼卵一经产下马上就会受精;如果这个过程像鲑鱼的情况那样,要持续数日的话,则雌鱼在这整个产卵期间里一直都会由雄鱼伴随左右。 (287) 在大多数场合中,鱼卵受精后就被其双亲弃置不顾,而失去了保护,因此就产卵而论,雄鱼和雌鱼所面临的危险是相等的,而且对于受精卵的产生,双方的重要性也是同等的;因此,任何一性的色彩鲜艳的个体,无论其鲜艳程度或大或小,其遭受毁灭或得到保存的倾向大概都是同等的,而且双方对其后代色彩的影响大概也是同等的。
属于若干个科的某些鱼类会筑巢,其中有些还会照顾刚孵化出来的幼鱼。颜色鲜明的锯隆头鱼(Crenilabrus massa和C.melops)的雌雄二者共同合作用海草和贝壳等材料修筑其巢穴。 (288) 但某些鱼类的雄者则单独承担了所有这项工作,以后还专门负责照顾幼鱼。颜色暗淡的刺鰕虎鱼类(gobies)的情况正是如此, (289) 其雌雄二者的颜色据知并无差异,刺鱼(Gasterosteus)的情况也是如此,其雄者的色彩在产卵季节则变得鲜艳。光尾刺鱼(G.leiurus)的雄者在一段长时间内以堪称模范的谨慎和警惕履行其作为一个“保姆”的义务,当幼鱼离巢太远之际,它还不断地徐徐把它们引还巢去。它还勇敢地把所有敌害赶走,包括其本种的雌鱼在内。倘若雌鱼产卵完毕后马上就被某种敌害所吞食,这对于雄鱼的确可能是一个不小的安慰,因为要不如此,它就得不停地把雌鱼从巢里赶走 (290) 。
栖息于南美和锡兰的某些其他鱼类,属于两个不同的“目”者,其雄鱼有一种异常的习性,把雌鱼产下来的卵集拢在嘴里或鳃腔里孵化。 (291) 亚加西兹教授告诉我说,亚马孙河的鱼种的雄者就有这种习性,“它不仅在一般时期比雌鱼颜色鲜明,而且这种差异在产卵季节比在其他任何时期还要大。Geophagus的鱼种也同样如此;在这个属中,雄鱼的前额在生殖季节有一个显著的突起物发育起来。关于鲷鱼类的各个物种,阿加西斯教授同样告诉我说,在以下各种场合中都可观察到其颜色上的性差异,即,“无论它们把卵产于水生植物之间的水里或把卵产于穴中,任其孵化,不再进一步给予照顾;还是在河泥里筑造浅巢,就像英国的刺盖太阳鱼(Pomotis)那样坐在其上。还应注意到这等坐巢鱼在它们各自所属的科中乃是颜色最鲜明的物种。例如,Hygrogonus是亮绿色的,具有黑色大眼斑,眼斑环以最显著的艳红色”。在鲷鱼类的所有物种中,是否只有雄鱼才坐守卵上,尚属不明。然而,鱼卵受到双亲保护或不受到保护这一事实显然对于雌雄之间的颜色差异并无多大影响或根本没有影响。更加显然的是,在雄鱼专门负责守护鱼巢和幼鱼的所有场合中,颜色较鲜明的雄鱼之毁灭,其对于本族性状的影响比颜色较鲜明的雌鱼之毁灭所造成的影响要深远得多,这是因为在卵的孵化或养育幼鱼期间,雄鱼的死亡将会招致幼鱼的死亡,所以就不能将其特性遗传给幼鱼;尽管如此,在许多这等场合中,雄鱼的色彩还是比雌鱼的更为显著。
在大多数总鳃类(Lophobranchii,海龙,海马等)中,雄鱼的腹部不是具有袋囊就是具有半圆形的凹陷,用以承受雌鱼产下来的卵并孵化之。雄鱼还显示了对幼鱼的强烈依恋。 (292) 雌雄二者的色彩通常差异不大;但京特博士相信雄海马的色彩比雌海马的鲜明。然而,剃刀鱼(Solenostoma)属提供了一个奇妙的例外, (293) 因其雌者色彩之强烈以及斑点之多均远比雄者为甚,而且只有雌者具袋状腹囊以孵化鱼卵;所以剃刀鱼的雌者在后述这一点和所有其他总鳃类都不相同,而在其色彩比雄者更为鲜艳方面也几乎和所有其他鱼类不同。雌者性状这种双重的颠倒,看来不可能是一种偶然的巧合。由于专门照顾鱼卵和幼鱼的若干鱼类的雄者在色彩上比雌者更为鲜艳,还由于这里提到的剃刀鱼的雌者负有同样的责任,而且其色彩比雄者更为鲜明,因而可以这样辩说:雌雄两性中对后代繁荣较为重要的那一性,其显著色彩必定在某种方式上是保护性的。但从雄者色彩比雌者更为鲜艳的大量鱼类来看,无论这等色彩是永久性的还是临时性的,雄鱼的生命对于本种的繁荣绝不比雌鱼的更为重要,因此上述这种见解几乎无法成立。当我们讨论鸟类时,还要遇到相似的情况,即,雌雄二者的正常属性完全颠倒了,到那时我们将会提出一个也许是合理的解释,这就是说,雄者选择更富吸引力的雌者,而不是按照整个动物界的正常规律,由雌者选择更富吸引力的雄者。
总之,我们可以断定,关于雌雄色彩或其他装饰性状有所差异的大多数鱼类,雄者最先发生了变异,然后将其变异传递给同一性别,并且通过以吸引或刺激雌者来实现的性选择把这等变异积累起来。然而,在许多场合中,这等性状或是部分地或是全部地转移给了雌者。再者,在其他一些场合中,雌雄二者为了保护自己而着有相似的色彩;但似乎没有任何事例表明,只有雌者为了保护的目的而在颜色或其他性状上特殊地发生了变异。
需要加以注意的最后一点是,据知鱼类会发出各种各样的声响,有的声响据描写犹如音乐一般。对这个问题特别注意的迪福塞(Dufossé)博士说,这等声音是由不同鱼类以若干方法故意发出的:或靠咽头骨摩擦而发声——或靠附于鳔上的某些肌肉振动而发声,而鳔则用做一种回声板——有的还靠鳔的内肌振动而发声。鲂鮄属(Trigla)用后面这种方法发出一种纯正而拖长的声音,约在八音度范围之内。但使我们最感兴趣的一个事例是鼬鳚属(Ophidium)有两个物种,只有它们的雄者才具发音器官,这种器官是由具有专门肌肉的、同鳔相连接的、能动的诸小骨构成的。 (294) 欧洲海洋中的荫鱼类(Umbrinas)据说在20 (295) 深的海底发出来的鼕鼕响声还可以被听到;罗歇尔(Rochelle)地区的渔民断言,“只有其雄鱼在产卵时期才会发出这样响声;其声音可加以模仿,因此不用诱饵即可将它们捕获”。 (296) 根据这个说法,尤其是根据鼬鳚属的事例,几乎可以肯定,脊椎动物中这个最低等鱼纲的发音器官,就像如此众多的昆虫类以及蜘蛛类那样,至少在某些场合中是作为把雌雄聚合在一起的一种手段,而且这等发音器官是通过性选择而发展起来的。
(295) 深的海底发出来的鼕鼕响声还可以被听到;罗歇尔(Rochelle)地区的渔民断言,“只有其雄鱼在产卵时期才会发出这样响声;其声音可加以模仿,因此不用诱饵即可将它们捕获”。 (296) 根据这个说法,尤其是根据鼬鳚属的事例,几乎可以肯定,脊椎动物中这个最低等鱼纲的发音器官,就像如此众多的昆虫类以及蜘蛛类那样,至少在某些场合中是作为把雌雄聚合在一起的一种手段,而且这等发音器官是通过性选择而发展起来的。
两 栖 类
有尾目(Urodela)
我从有尾两栖类开始。蝾螈的雌雄二者往往在色彩和构造上都大不相同。在某些物种中,雄者的前肢于生殖季节有抱握爪发育起来;雄蹼足北螈(Triton (297) palmipes)的后足在此季节有游泳蹼,而在冬季又几乎完全被吸收了;因而这时雄者的足遂与雌者的相类似。 (298) 毫无疑问这等构造有助于雄者对雌者的热切追求。它向雌者求爱时即迅速摆动其尾端。关于英国的普通小蝾螈(斑北螈,Triton punctatus和冠北螈,T.cristatus),雄者在生殖季节有一条深缺刻的多锯齿冠饰沿其脊背和尾巴发育起来,而到冬季就消失了。圣乔治·米伐特(St.George Mivart)先生告诉我说,这种冠饰无肌肉,因此不能用于运动。由于这种冠饰的边缘在求偶季节变得色彩鲜明,所以这是一种雄性的装饰,几乎没有疑问。还有许多物种的躯体呈现了差别非常悬殊的色调,平时是灰黄色,但到生殖季节则变得比较鲜艳。例如英国普通小蝾螈(斑北螈)的雄者,其“上面为灰褐色,下面为黄色,到春季又变为一种鲜艳的橙色,并有圆形黑斑分布于身体各处”。这时,其冠饰的边缘也呈鲜红色或紫色。其雌者平时为黄褐色,杂以褐色斑点,其底面的颜色则往往十分单调。 (299) 幼者的色彩是暗淡的。卵在排出过程中即受精,随即被其双亲弃之不顾。由此我们可以断定,雄者是通过性选择获得其十分显著的色彩和装饰性附器的;这等性状或专向雄性后代传递,或向雌雄双方传递。
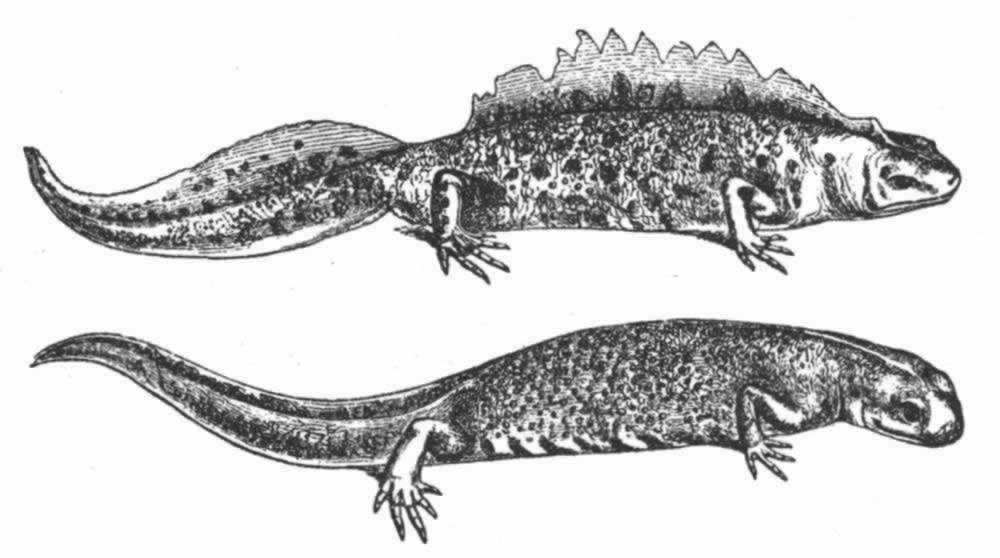
图 32 冠北螈(Triton cristatus)
(原体大小之半,采自贝尔的《英国爬行类志》)上图为生殖季节的雄者;下图为雌者。
无尾目(Anura)或蛙类(Batrachia)
许多蛙类和蟾蜍类的色彩显然是作为一种保护之用的,如雨蛙的鲜绿色彩以及许多陆栖物种的斑驳而暗淡的色调就是如此。我曾见过的色彩最显著的蟾蜍是黑蟾蜍(Phryniscus nigricans) (300) ,其躯体的整个上表面黑如墨水,脚蹠以及腹的局部则有最鲜明的朱红斑点。在拉普拉塔(La Plata)的灼热太阳下,它在不毛的沙地或开阔的草原上到处爬行,不会不被每一只路过的动物所看见。这等色彩对它大概是有利的,因为这可以使所有猛禽类知道这种动物是一种味道恶劣的食物。
尼加拉瓜有一种小蛙,“披着一身红色和蓝色的鲜艳装束”,它不像大多数其他物种那样把自己隐蔽起来,而在大白天到处蹦跳,贝尔特先生说 (301) ,他一见到它那泰然自若的样子,就深信它是不可食的了。经过若干次试验后,他才成功地诱使一只幼小雌鸭衔住一只这种幼蛙,但马上就把它丢掉了;这只鸭子“甩动脑袋走来走去,犹如试图甩掉某种讨厌的味道一样”。
关于雌雄颜色的差异,无论是蛙类还是蟾蜍类,京特博士都不知有任何显著的事例;但他常常能凭雄者比雌者的稍为浓烈的色彩就可以把雄者同雌者区别出来。关于外部构造,他也不知道雌雄二者有任何显著差异,除了雄者的前肢上有些突起在生殖季节变得发达起来,借此以抱持雌者。 (302) 这等动物没有获得更为强烈显著的性征是令人奇怪的;因为,它们虽是冷血动物,但其激情却是强烈的。京特博士告诉我说,他有几次发现一只不幸的雌蟾蜍因被三四只雄者如此紧紧地抱住而窒息闷死。霍夫曼(Hoffman)教授在吉森(Giessen)见过蛙类当生殖季节终日争斗不止,而且进行得如此激烈,以致其中一只的躯体竟被撕裂。
蛙类和蟾蜍类有一种有趣的性差异,即雄者具有发出音乐声响的能力;但说到音乐,如果把这个名词应用于雄菜蛙和某些其他物种所发出的那种压倒一切的不谐和声响,按照我们的欣赏力来说,似乎是非常不适当的。然而,有些蛙类的鸣唱无疑还是悦耳的。在里约热内卢附近,我惯于在傍晚坐下来,倾听那些栖息在水草上的小雨蛙(Hylae)发出来的谐和而美妙的声调。这各种声音主要是雄者在生殖季节发出的,犹如英国普通蛙在这种场合中也哇哇地乱叫一样 (303) 。与此事实相一致的是,雄者的发音器官比雌者的更为高度发达。在一些属中,只有雄者才具有与喉相通的囊 (304) 。例如,食用蛙(Rana esculenta)“的囊是雄者所特有的,在哇哇鸣叫时囊中充满了空气,成为球状的大气胞,位于头部两侧的咀角附近”。这样,雄者的哇哇叫声表现得十分有力;而雌者的叫声只不过是一种微弱的呻吟而已 (305) 。在这一科的某些属中,发音器官的构造大有差异,在所有场合中,它们的发达大概可以归因于性选择。
爬 行 类
龟类(Chelonia)
龟类和海龟类都没有呈现十分显著的性差异。在某些物种中,雄者的尾比雌者的较长。有些物种的雄者,其腹甲或其甲壳的下表面轻微凹陷而同雌者的背甲隆起相吻合。美国雄锦龟(Chrysemys picta)前足的爪为雌者的两倍长,在交配时使用。 (306) 加拉帕戈斯群岛有一种巨龟,名为黑陆龟(Testudo nigra),其雄者成熟后的躯体据说大于雌者;雄者只在交配季节期间,而不在其他期间,发出一种嘶哑的吼声,可闻于一百多码以外;反之,雌者决不发出叫声。 (307)
关于印度的丽陆龟(Testudo elegans),据说“其雄者在争斗中互相冲撞时所发出的声音可闻于一定的距离”。 (308)
鳄类(Crocodilia)
其雌雄二者的颜色显然无差异,我也不知道有雄者互斗之事,尽管这是可能的,因为有些种类的雄者在雌者面前极尽夸示自己之能事。巴特兰姆(Bartram) (309) 描写过在咸水湖中有一只雄性短吻鳄,为了尽力赢得雌者,而在湖中兴浪作波和大声吼叫,“鼓气到快要爆裂的程度,头尾高举,在水面上跳跃转圈,犹如一个印第安人的酋长在练习他的武艺一样”。在求爱季节,鳄的颌下腺散发出一种麝香气味弥漫在它经常出没的地方 (310) 。
蛇类(Ophidia)
京特博士告诉我说,其雄者总是小于雌者,而且一般具有比较细而长的尾巴;但在外部构造上,他不知道有任何其他差异。关于颜色,他依据雄者的更为强烈显著的色彩,几乎总能把它同雌者区别开来;例如,英国雄蝮蛇背上的那种之字形黑色带斑比雌者的更为清晰显著。北美响尾蛇雌雄之间的差异就愈益明显得多了,如动物园管理员指给我看的,其雄者周身均具有更为灰白的黄色,所以立刻可以把它同雌者区别开来。南非洲有一种蛇叫做牛头蛇(Bucephalus capensis),它表现有某种相似的差异,因其雌者“从不像雄者那样在其身体两侧如此充分地具有黄的斑驳”。 (311) 反之,印度有一种毒蛇,名为双突齿食螺蛇(Dipsas cynodon),其雄者呈黑褐色,腹部的一部分为黑色,而雌者则呈微红色或淡绿黄色,腹部或一律为黄色或有黑色大理石斑纹。该地还有一种蛇,叫做Tragops dispar,其雄者为鲜绿色,而雌者则呈青铜色。 (312) 某些蛇类的色彩无疑是保护性的,如树蛇类(tree-snakes)所呈现的绿色以及生活在沙质地方的那些物种所呈现的各种浓淡颜色的斑驳都是如此;但有许多种类,如英国的蛇和蝮蛇,其色彩是否用来隐蔽自己仍是一个疑问;对许多具有极为漂亮色彩的外国蛇的物种来说,这一疑问就更大了。有些蛇的物种,其色彩在成年时和幼小时很不相同。 (313)
蛇类肛门香腺的功能在生殖季节变得活跃了, (314) 蜥蜴类的肛门腺以及鳄类的颌下腺都是如此。由于大多数动物都是雄者寻求雌者,所以这等香腺与其说是为了把雌者引至雄者所在的地点,莫如说是为了刺激雌者或向它献媚。雄蛇虽然显得那样懒惰,却也多情;因为,曾见过许多条雄蛇集拢在同一条雌蛇的周围,甚至后者是一条死蛇,也会如此。尚未发现雄蛇因竞争雌蛇而互斗。它们的智力要比可能预料到的为高。它们在动物园里很快就懂得不能去碰撞用来打扫其笼子的铁栅门;费城的基恩(Keen)博士也告诉我说,他养的某些蛇类经历四五次后就懂得避开一种活套,用这种活套,起初很容易就把这些蛇逮住。锡兰有一位杰出的观察者莱亚德(E.Layard)先生见过 (315) 一条眼镜蛇把头伸入一个窄洞去吞食一只蟾蜍。“由于增加了蟾蜍这个障碍,它无法从窄洞缩回来;发现了这一点后,就勉强吐出了这口开始挣脱的宝贵食物;但这同蛇的哲学太不相容了,于是蟾蜍再次被它弄住,这条蛇经过剧烈的努力以图逃脱后,不得不再把口中的食物舍掉。然而这一次它得到了教训,于是咬住蟾蜍的一只腿,把它拖出,然后吞之大吉”。
动物园的管理员证实,某些蛇类,如响尾蛇属(Crotalus)和蟒蛇属(Python),都能把他同别的人区别开。关在同一个笼里的眼镜蛇(Cobras)相互之间显然有某种依恋之情。 (316)
然而,不能因为蛇类具有某种推理力、强烈的激情以及相互的爱情,就认为它们也同样赋有充分的鉴赏力以欣赏其配偶的鲜艳色彩,因而通过性选择使鲜艳色彩成为物种的装饰。但某些物种的极端美丽是难于以任何其他方法加以说明的,举例说,南美的珊瑚蛇类(coral-snakes)呈艳红色,并具黑色和黄色的横带斑。我记得很清楚,当我在巴西第一次见到一条珊瑚蛇滑过一条小路时,它的美色使我感到了何等惊奇。正如华莱士先生根据京特博士的材料所说的 (317) ,全世界除南美洲外,没有任何一处地方的蛇类有这种特殊颜色,而且在南美洲发现的这种蛇至少有四个属。其中一个属叫做Elaps,是有毒的;第二个大不相同的属,是否有毒尚属疑问,其他两个属则完全是无害的。属于这几个不同属的物种均栖息于同一地区,而且彼此如此相像,以致“除非博物学者大概谁也不能把无害的种类和有毒的种类加以区分”。因此,正如华莱士先生所相信的,无毒的种类大概是为了保护自己,根据拟态的原理,而获得其色彩的;这是因为其敌害自然会认为它们是危险的。然而,有毒的Elaps鲜明色彩的形成原因尚有待于阐明,这个原因也许就是性选择。
蛇类除嘶嘶鸣叫外还能发出其他声音。剧毒的龙首蝮蛇(Echis carinata)其躯体两侧各有数行构造特殊的斜鳞片,鳞片边缘为锯齿形;当引起这种蛇激动时,这等鳞片就互相摩擦,产生“一种奇妙而拖长得近似嘶嘶的声音”。 (318) 关于响尾蛇所做的咔塔咔塔响声,我们终于得到了一些确实的材料:因为奥盖(Aughey)教授说 (319) ,他曾两次把自己荫蔽起来,在一段不远处注视着一条响尾蛇盘蜷昂首,持续发出间隔短暂的嘎啦嘎啦响声达半小时之久:最后他见到另一条蛇来到了,它们相遇后即行交配。因此,他满意地认为这种嘎啦嘎啦声的用途之一就是把雌雄二者引到一块。遗憾的是,他不能确定在原处保持不动而招引异性的到底是雄者还是雌者。但根据上述事实,决不能说这等嘎啦嘎啦声对这些蛇类就不会有其他用途,例如作为对某些动物的警告大概就是一种用途,否则这等动物也许要对它们进行攻击。关于这等声响会使它们的捕获物吓得瘫痪的若干记载,我也不能完全置之不信。还有某些其他蛇类,把它们的尾部对着周围的树干迅速摆动,而发出一种清晰的声响,我在南美亲自听过一条蝮蛇(Trigonocephalus)就发出这种声音。
蜥蜴类(Lacertilia)
蜥蜴类的一些种类的雄者,也许是许多种类的雄者都因竞争雌者而互斗。例如南美树栖的冠饰安乐蜥(Anolis cristatellus)就极其好斗:“在春季和初夏期间,两只成熟的雄者相遇时很少不发生争斗的。它们最初相遇之际,频频点头三四次,同时喉下的襞状部或喉囊便膨胀起来;双眼闪耀着愤怒的光芒,左右摆尾数秒钟,好像是聚集气力,然后彼此猛扑,上下翻滚,互相用牙齿紧紧地咬住不放。这场冲突一般以战斗的一方失去尾巴而告结束,常常是胜利者把对方的尾巴吃掉。”这个物种的雄者远远大于雌者, (320) 根据京特博士所能确定的来说,这一点乃是一切蜥蜴类的一般规律。安达曼群岛的红裸趾虎(Cyrotodactylus rubidus)只有雄者具肛前孔(preanal pores),根据类推,这种孔大概是用以散发香气的。 (321)
雌雄二者的各种外部性状往往大不相同。上述安乐蜥的雄者沿着其脊背和尾巴生有一条冠饰,可以随意竖起;但雌者没有呈现这等冠饰的丝毫痕迹。印度聋蜥(Cophotis ceylanica)的雌者也有一脊冠,但远不及雄者的发达。正如京特博士告诉我的,许多鬣鳞蜥类(Iguanas)、避役类(Chameleons)以及其他蜥蜴类的情况也是如此。然而,在某些物种中,雌雄二者的冠饰是同等发达的,如瘤疣鬣鳞蜥(Iguana tuberculata)即是。在赛塔蜥蜴(Sitana)这一属中,只是雄者才具有一个大喉袋(图33),它可以像一把扇子那样折叠起来,呈蓝、黑、红三种色彩;但这等色彩只在求偶季节才有所表现。雌者没有这种附器,甚至连一点痕迹也没有。按照奥斯汀先生的材料,冠饰安乐蜥的雌者也有喉袋,呈艳红色并具黄色大理石花纹,尽管它是处于残迹状态的。再者,另外还有某些蜥蜴类,其雌雄二者都有同等发达的喉袋。在这里,我们看到了同上述许多事例一样的情况,即,在属于同一类群的诸物种中,同一种性状,有的只限于雄者才有,有的在雄者方面远比在雌者方面发达,有的在雌雄两方面都同等发达。飞蜥属(Draco)的小蜥蜴类借附于肋骨的膜伞在空中滑翔,其色彩之美丽实非语言所能形容,它的喉部具有皮质附器,“犹如鹑鸡类的垂肉”。当这种动物激动时,这等附器就会竖起。雌雄二者都有这等附器,然而它在雄者发育成熟时最为发达,这时,中央附器有时竟有头的两倍长。同样地,大多数物种沿着颈部也生有一条矮冠,完全成熟的雄者的这种矮冠比雌者的或幼小雄者的都发达得多。 (322)

图 33 小赛塔蜥蜴(Sitana minor)具有膨胀喉袋的雄者
(采自京特的《印度的爬行类》)
图 34 斯氏角蜥(Ceratophora stoddartii)
上图为雌者;下图为雄者。
据说有一个中国的物种在春季成双成对地生活在一块,“要是有一只被捕获,另一只就会自树上坠至地面,泰然就擒”——我推测这是出于绝望。 (323)
某些蜥蜴类雌雄二者之间尚有其他更显著的差异。有一种角蜥(Ceratophora aspera),其雄者在吻端生有一种附器,长达头部的一半,圆柱状,复以鳞片,易弯,显然能竖立,但在雌者方面这等构造完全是残迹的。同属的第二个物种在其易弯的附器顶部有一个小角,是由一个末端鳞片形成的。而第三个物种斯氏角蜥(C.Stoddartii,图34)的整个附器已变成一支角,通常为白色,但当这种动物激动时,就会呈现带紫的色彩。后面这个物种的成熟雄者的这支角长达半英寸,但雌者和幼小雄者的这支角则十分小。这等附器,如京特博士对我说的,可同鹑鸡类的肉冠相比拟,而且显然是用做装饰的。
图 35 双角避役(Chamaeleo bifurcus)
上图为雄者;下图为雌者。
图 36 欧氏避役
上图为雄者;下图为雌者。
在避役属中,我们见到雌雄之间的差异已达到顶点。栖息于马达加斯加的雄性双角避役(C.bifurcus,图35)在其头骨的上部生出两个坚硬的巨大骨质突起,就像头部其余部分那样地复以鳞片,这等构造上的奇异改变在雌者方面仅显示一点痕迹而已。再者,非洲西海岸的欧氏避役(Chamaeleon owenii图36),其雄者的吻部和前额生有三只奇异的角,而在雌者方面则连一点痕迹也没有。这种角是由一种骨的赘生物构成的,复以平滑的外鞘,外鞘乃躯体普通外皮的一部分,因而它们同公牛、山羊或其他具有鞘角的反刍动物的角在构造上是相同的。尽管这三只角和双角避役头骨上那两个巨大的延长物在外观上如此大不相同,但我们几乎无法怀疑它们在这两种动物的组织中都是为着同一个总目的服务的。每个人所产生的第一种猜测将认为雄者利用这种角互斗,由于这些动物很善于争吵, (324) 因此这个观点也许是正确的。伍德先生也告诉我说,他有一次见到两只小避役(C.pumilus)在树枝上激烈厮斗;用头猛冲,彼此试图咬住对方,然后休息一会儿,接着又继续厮斗。
有许多蜥蜴类,其雌雄二者的颜色有轻微差异,雄者的色彩和条纹比雌者的较为鲜明和轮廓较为清楚。例如,上述聋蜥属和南非洲的棘趾蜥(Acanthodactylus capensis)的情况就是如此。南非绳蜥属(Cordylus)的雄者不是比雌者红得多就是绿得多。印度黑唇树蜥(Calotes nigrilabris)的雌雄差异还要大,雄者嘴唇为黑色而雌者嘴唇乃绿色。英国的普通胎生小蜥蜴(Zootoca vivipara),“其雄者的躯体底面和尾基呈鲜橙色并具黑色斑点;雌者的这些部分则呈浅灰绿色而且无斑点”。 (325) 我们已经知道塞塔蜥蜴只是雄者生有一个喉袋,并呈华丽的蓝、黑、红三种色彩。智利的瘦蜥蜴(Proctotretus tenuis)只有雄者才呈现蓝色、绿色和红铜色的斑点。 (326) 在许多场合中,雄者全年都保持一样的色彩,但在另外一些场合中,雄者的色彩在生殖季节则变得鲜明得多;我再补充一个例子,即玛丽亚树蜥(Calotes maria)的雄者在这生殖季节,其头部呈鲜红色,而躯体其余部分则为绿色。 (327)
许多物种的雌雄二者,其美丽的色彩完全相似,把这等色彩设想为保护色是毫无理由的。关于那些生活于草木之中的鲜绿色种类,这样的颜色无疑是为着荫蔽它们自己而服务的。在巴塔戈尼亚(Patagonia) (328) 北部我见过斑点蜥蜴(Proctotretus multimacula—tus)一受惊就伸平其躯体,闭上眼,于是凭其斑驳的色调几乎无法把它同周围的沙地分别开。不过,如此众多蜥蜴类所装饰的鲜艳色彩,以及它们所具有的各种奇异的附器,大概还是作为一种魅力先由雄者获得的,然后此等性状或只被传递给其雄性后代或被传递给其雌雄双方的后代。性选择对爬行类所起的作用确实就像对鸟类所起的作用那样,差不多是同等重要的:雌者的色彩不及雄者的显著,正如华莱士先生所相信的鸟类情况那样,是不能用雌鸟在孵卵期间面临着较大的危险来解释的。
第十三章 鸟类的第二性征
性差异——斗争的法则——特殊武器——发声器官——器乐——爱情的滑稽表演和舞蹈——永久性和季节性的装饰物——一年两次和一年一次的换羽——雄者夸耀其装饰物
鸟类的第二性征,同任何其他动物纲相比,也许不会引起其构造发生更重要的变化,但鸟类的第二性征却更加多种多样而且更加显著。因此,我将用相当长的篇幅来讨论这个问题。雄性鸟类有时也具有用于相斗的特殊武器,虽然这并不多见。它们用各种各样的声乐和器乐来魅惑雌鸟。从它们躯体的各个部分生出各种各样优美的肉冠、垂肉、隆起物、角、鼓气的囊、顶结、裸羽轴、羽衣以及修长的羽毛,用以装饰自己。它们的喙、头部周围的裸皮以及羽毛常常具有华丽的色彩。雄鸟有时靠舞蹈要不在地上或天空作出古怪的滑稽表演来表达它们的求爱。至少有一个事例表明雄鸟散发出一种麝香气味,我们可以设想这是用来魅惑或刺激雌鸟的;因为杰出的观察家拉姆齐(Ramsay)先生 (329) 说道,澳洲麝鸭(Biziura lobata)“在夏季月份里散发这等气味的,只限于雄鸭,同时有些个体可以把这种气味保持全年;甚至在繁殖季节我也从未打下过一只具有任何麝香气味的雌鸭”。这种气味在交配季节是如此强烈,以致远在见到这种鸟类之前就可发觉这种气味了。 (330) 总之,鸟类大概是所有动物中的最善于审美者,当然不及人类,它们几乎具有同我们一样的审美力。这一点可从下述情况得到说明,即,我们喜爱倾听鸟类的鸣唱,我们的妇女,不论是文明的还是未开化的,都爱用鸟类羽毛来装饰头部,而且还爱戴宝石,但其色彩几乎并不比某些鸟类的裸皮和垂肉的色彩更为鲜艳。然而,人类既开化之后,他们的美感显然是一种愈益复杂得多的感觉,而且这是同种种理智的观念(intellectual ideas)联系在一起的。
当讨论我们这里所特别关切的性征以前,我想先略微谈一谈雌雄之间显然取决于不同生活习性的某些差异;因为这等情况在低等动物纲中虽是普通的,但在高等动物纲中却是罕见的。栖息于胡安·费尔南德斯(Juan Fernandez)群岛上的Eustephanus属的两种蜂鸟长期被人认为是不同的物种,但现已弄清楚,正如古尔德(Gould)先生告诉我的,它们原来是同一物种的雄者和雌者,二者在喙的形状上有轻微差异。在蜂鸟的另一个属(Grypus)中,雄者的喙缘为锯齿状,喙端为钩状,因而同雌者的喙大不相同。新西兰的新态鸟(Neomorpha),正如我们已经见过的,其雌雄二者因取食方式的关系,喙的形状出现了更加广阔的差异。已经观察到金翅雀(Carduelis elegans)有某种类似的差异,因为詹纳·韦尔先生曾向我保证说,捕鸟人能根据其雄鸟的稍长的喙,而把它识别出来。常常发现成群的这种雄鸟吃起川续断(Dipsacus)的种子,它们的长喙可以啄到这些种子,而雌鸟更常吃的却是水苏(betony)或玄参属(Scrophularia)植物的种子。以这样一种轻微的差异为基础,我们可看到雌雄二者的喙如何通过性选择而形成巨大差异。然而,在上述某些场合中,雄鸟的喙最初发生变异可能同其他雄鸟进行斗争有关,而以后这又导致了生活习性的轻微变化。
斗争的法则
几乎所有的雄性鸟类都极其好斗,它们用喙、翅和腿互相争斗。每年春季我们可以看到英国的歌鸲鸟(robins)和麻雀(sparrows)就是如此。所有鸟类中体型最小的为蜂鸟,而它却是最好争吵的鸟类之一。戈斯(Gosse)先生 (331) 描述过一对蜂鸟的一次争斗,它们互相咬住对方的喙不放,在空中来回旋转,直到几乎落地;孟斯·德奥卡(M.Montes de Oca)当谈到蜂鸟的另一属时说道,两只雄鸟如果在空中相遇很少不发生激烈冲突的:当把它们一块关进笼里时,“它们之间的战斗结果多半是其中之一的舌头被撕裂开,因而以后肯定因不能进食而死去。” (332) 至于涉禽类(Waders),普通黑水鸡(Gallinula chloropus)的雄鸟“在求偶时,为争夺雌鸟而激烈争斗:它们几乎直立水中,用脚互相踢打”。有人见过两只这种雄鸟如此争斗了半小时之久,直到其中一只抓住了另一只的头,要不是旁观者加以干涉的话,被抓住的那只雄鸟大概要被弄死;而雌鸟在这整个期间犹如一个安静的观众,站在一旁看热闹。 (333) 布赖茨先生告诉我说,有一种同鷭亲缘相近的鸟(凤头董鸡Gallicrex cristatus),其雄鸟大于雌鸟三分之一,它们在繁殖季节非常好斗,因而东孟加拉当地人把它们养起来,让它们相斗。在印度也饲养各种其他的鸟用于相同的目的,例如,红鹎(Pycnonotus haemorrhous)就是“斗志昂扬地进行争斗”。 (334)
多配性的流苏鹬(Machetes pugnax,图37)以其极端的好斗性而闻名;其雄鸟的体形大大超过雌鸟,它们在春季日复一日地聚集于某一特定的地点,那里就是雌鸟打算产卵的地点。捕鸟人根据草皮被践踏得有点光秃的情况就可发现这种地点。雄鸟在这里厮打,很像斗鸡那样,互相用喙啄住不放,彼此以翅相击。这时,颈部周围的长羽毛直竖,据蒙塔古上校说,“它像一面盾牌似的扫过地面,以保护躯体比较脆弱的部分”;关于鸟类的任何构造当做盾牌用的情况,这是我所知道的唯一例子。然而,从其颈羽的种种富丽色彩看来,大概这主要是作为一种装饰之用的。就像大多数好斗的鸟类那样,它们似乎做好了随时战斗的准备,一旦把它们关在一起,就往往互相残杀;但蒙塔古观察到它们的好斗性在春季变得较强,这时其颈部长羽就充分发育了;而且在这期间,任何一只鸟的最小动作都会引起一场普遍的战争。 (335) 关于蹼足鸟类的好斗性,举出两个例子来说明就足够了:在圭亚那,“野麝鸭(Cairina moschata)的雄者在繁殖季节就会发生血腥的战斗;凡发生过这等战斗的地方,河面上便有一段距离布满了羽毛。” (336) 似乎不适于战斗的鸟类便进行激烈的冲突。例如,鹈鹕(pelican)的较强雄者会把较弱者赶走,用其巨大的喙猛啄,以其翅膀进行沉重的打击。雄沙锥(snipe)在互斗时,“用嘴以能想象的最奇妙方式又曳又推”。某些少数鸟类据信从不相斗,按照奥杜邦(Audubon)的材料,美国的金黄色啄木鸟(Picus auratus)就是如此,虽然“跟随这种雌鸟的放荡追求者竟有半打之多”。 (337)

图 37 流苏鹬(Machetes pugnax)
(采自布雷姆(Brehm)的《Thierleben》)
许多鸟类的雄者都大于雌者,这无疑是许多世纪以来较大、较强壮的雄者在胜过其竞争对手方面占有优势的结果。有几个澳大利亚的物种,其雌雄二者在体形上的差异已达到了极端。例如,雄麝鸭(Biziura)和雄Cincloramphus cruralis(同英国鹨的亲缘接近)经过实测各为其雌者的两倍大。 (338) 另外有许多鸟类,则是雌鸟大于雄鸟。如上所述,对这种现象提出的解释,常常认为是由于雌鸟在养育幼鸟方面承担了大部分工作,但这种解释是不够充分的。以后我们将会见到,在某些少数场合中,雌鸟显然是为了战胜其他雌鸟并占有雄鸟而获得其较大的体形和较强的体力。
许多鹑鸡类的雄者,尤其是一雄多雌的种类,都具有同其竞争对手进行战斗的特殊武器,叫做距,使用这种武器能产生可怕的效果。一位可信赖的作者曾做过这样记载 (339) :在德比郡(Derbyshire)有一只鸢(kite)袭击一只带着小鸡的雌斗鸡,这时,雄斗鸡飞奔来救,跃起一踢,用距准确地刺穿了侵略者的眼和头骨。把距从鸢的头骨中拔出是不容易的,而且鸢虽死了,仍牢牢地抓住对手不放,因而这两只鸟紧紧地联在一起了;但把雄斗鸡解脱出来之后,才知道它只不过受了点轻伤而已。雄斗鸡大无畏的勇气是有名的:有位先生很久以前曾目睹过下面的残酷景象,他告诉我说,有一只斗鸡因斗鸡场的某种事故而双腿折断了,它的主人打赌说,如果能把这只斗鸡的腿接好使它直立,它就会继续斗下去。结果骨折处被接好了,这只斗鸡又勇敢地投入战斗,直到它受到了致命的一击。在锡兰有一个亲缘相近的野生物种,名为斯氏原鸡(Gallus stanleyi),据知它们在“保卫其配偶”时进行殊死的战斗,因而常常发现斗者之一死于战斗之中。 (340) 一种印度石鸡(Ortygornis gularis)的雄者具有坚固而锐利的距,它们如此喜欢争吵,以致“你所捕杀的几乎每只鸟的胸部都有以往战斗的累累伤痕”。 (341)
几乎所有鹑鸡类的雄者,每当繁殖季节都进行猛烈的冲突,即使无距者亦复如此。松鸡(capercailzie)和雄黑松鸡(Tetrao urogallus和T.tetrix)都是一雄多雌者,它们有约好的固定地点,群集于此,进行战斗并向雌鸟献媚达数周之久。柯瓦列夫斯基博士告诉我说,他在俄国曾经看到松鸡相斗过的场所,那里的雪全被血染红,当几只雄黑松鸡“进行一场大战之后”,也弄得羽毛四处飞扬。在德国,把雄黑松鸡求偶的歌舞称为巴尔兹(Balz),老布雷姆对此做过奇妙的记载。这种鸟几乎连续地发出最奇怪的叫声:“高举其尾,展开为扇,昂起头和颈,所有羽毛全都竖起,并展其双翅。然后它朝不同的方向跳跃几步,有时是绕圈跳跃,并用其喙的下部抵住地面,而且抵得如此用劲,以致颏部羽毛纷纷被磨掉。当做这些动作时,它拍击双翅,转了一圈又一圈。它的热情越高,就变得越活泼,直到最后这种鸟看来就像一个疯狂的动物。”在这样的时候,雄黑松鸡是如此精神贯注,以致几乎变得什么也看不见,什么也听不到了,但同雷鸡相比,还有逊色,因此可在同一地点一只接一只地把它们射杀,甚至可徒手把它们挨个捉住。雄鸟在做完这些滑稽表演之后就开始相斗:同一只雄黑松鸡为了证明其体力胜过若干敌手,要在一个清晨里走访几处巴尔兹舞场(Balz-places),而这等场所在连续数年内都是保持不变的。 (342)
具有长尾羽的孔雀与其说它像个战士,不如说它更像个纨绔子弟,但它们有时也发生猛烈的冲突:福克斯(W.Darwin Fox)牧师告诉我说,离切斯特不远的地方有两只相斗的孔雀变得如此激怒,以致它们飞越了整个城市仍在厮打,直到它们降落在圣约翰塔顶上才算结束。
鹑鸡类所具有的距一般只是单独一个;但多距的鸟类每只腿上则具有两个或两个以上的距;已经见过一种血雉(Ithaginis cruentus)的腿上有五个距。距一般只限于雄鸟才有,而在雌鸟腿上仅表现为小瘤、即残迹物;但爪哇绿孔雀(Pavo muticus)的雌者以及布莱思(Blyth)先生告诉我的小型火背雉(Euplocamus erythropthalmus)的雌者都有距。在鹑鸡属(Galloperdix)中,通常是雄鸡每只腿上有两个距而雌鸡只有一个距。 (343) 因此可以把距视为一种雄性构造,偶尔或多或少地传给了雌鸡。就像大多数其他次级性征那样,同一物种的距无论在数量上或发育程度上都是高度容易变异的。
各种各样的鸟类在其双翅上有距。但埃及鹅(Chenalopex aegyptiacus)只有“光秃而不锐利的小瘤而已”,而这等小瘤大概向我们展示了在其他物种中发展起来的真距的最初步骤。具有翅距的鹅,即距翅鹅(Plectropterus gambensis),其雄者的距比雌者的大得多;正如巴特利特先生告诉我的,它们用这等翅距进行争斗,因此在这种场合中,翅距是作为性的武器来用的;但按利文斯通(Livingstone)的见解,它们主要是用来保卫幼者的。叫鸟(Palamedea)(图38)在每张翅上都装备了一对距,这是一种非常可怕的武器,据了解,只要用它一击就会把狗打得哀号而逃。但在这种场合或在某些具有翅距的秧鸡类(rails)的场合中,雄鸟的距并不见得比雌鸟的大。 (344) 然而,在某些鸻类(plovers)中,必须把其翅距视为一种性征。例如,英国普通凤头麦鸡(Vanellus cristatus)的雄者在繁殖季节其翅肩上的小结节变得更为突出,而且互相争斗。跳凫属(Lobivanellus)的某些物种有一种相似的小结节在生殖季节就会发展成“一支短的角质距”。澳大利亚的裂跳凫(L.lobatus)的雌雄二者都有距,但雄者的距比雌者的大得多。与此亲缘相近的一种鸟,叫做Hoplopterus armatus,它的距在生殖季节并不增大;但在埃及曾见过这种鸟互相争斗,它们争斗的方式同英国田凫一样,在空中突然转向对方,从侧面互相攻击,时常会造成致命的后果。它们还这样把其他敌对者赶走。 (345)

图 38 角叫鸟(Palamedea cornuta)
(采自布雷姆)图示其双翅距,以及头顶的丝状物。
求偶的季节同时也是相斗的季节,但有些鸟类的雄者,如斗鸡和流苏鹬的雄者,甚至野火鸡和松鸡类的年青雄者 (346) ,不论何时,只要一相遇就会随时发生争斗。雌鸟的在场是这等可怕的战争之根源(teterrima belli causa)。孟加拉的印度绅士们挑起梅花雀(Estrelda amandava)小而美的雄者进行斗争的方法是,把三个小鸟笼排成一行,中间那只鸟笼关的是一只雌鸟,两头各关着一只雄鸟,过一会儿把这两只雄鸟放开,它们就立即开始了一场殊死的战斗。 (347) 当许多雄鸟聚集在同一个约定的地点进行争斗时,就像松鸡类和其他种种鸟类的情况那样,一般都有雌鸟呆在旁边 (348) ,争斗结束后,它们就和胜利的斗士相配。但在某些场合中,交配是在争斗之前而不是在争斗之后进行的:例如,按照奥杜邦的材料 (349) ,弗吉尼亚夜鹰(Caprimulgus virginianus)的若干雄鸟“以一种非常殷勤的方式向雌鸟求偶,当雌鸟刚一作出选择后就同意追击所有其他入侵者,并把它们赶出其领地之外”。雄鸟一般在交配前就试图把其竞争对手赶走或杀死。然而,看来雌鸟并不见得总是喜爱胜利的雄鸟。柯瓦列夫斯基博士的确曾向我保证说,雌松鸡有时会携一只年青的雄鸟私奔而去,后者是不敢同年长的雄鸟一块进入争斗场所的,苏格兰的雌赤鹿偶尔也会如此。当两只雄赤鹿在单独一只雌赤鹿面前进行争夺时,胜利者通常无疑会遂其意愿;但有些这类斗争是由于到处乱跑的雄者试图破坏已经成为配偶的安宁所引起的。 (350)
即使是最好斗的物种,其雌雄的交配也可能不完全取决于雄者单纯的体力和勇气;因这等雄者一般都有种种装饰物来打扮自己,这些装饰物在生殖季节往往变得更鲜艳,并殷勤地在雌者面前进行夸示。雄者还用爱情的呼叫、鸣唱和滑稽表演来尽力献媚或刺激雌者,这种求爱在许多场合中是一种费时甚久的事。因此,雌者对异性的魅惑大概既不是无动于衷,也不总是被迫顺从胜利的雄者。更加可能的是,在雄者冲突之前或以后,雌者受到了一定雄者的刺激而不知不觉地爱上了它们。关于伞松鸡(Tetrao umbellus),一位杰出的观察家 (351) 甚至相信其雄者的相斗“完全是假装的,它们的表演是为了向集合于雄者周围而表示赞美的雌者显示其最大的优越性;因为我从来没有找到过一只受了伤的英雄,而所看到的折断的羽毛,很少超过一支”。以后我还要重新来讨论这个问题,但我愿在这里补充一点:关于美国的一种狂热松鸡(Tetrao cupido),约有20只雄者集合于一特定地点,大摇大摆地走来走去,空中到处回响着其喧嚣声。当从一只雌者得到第一个响应后,雄者便投入了猛烈的相斗,结果是弱者败走了;但这时,据奥杜邦说,胜败双方都寻求雌者,因此,雌者必须立即作出选择,否则争斗必定还要重新发生。再者,美国有一种草地鹨(Sturnella ludoviciana),其雄者激烈相斗,“但一见到雌者,就疯狂般地全都随着它飞去”。 (352)
声乐和器乐
鸟类的鸣声是用以表达其种种感情的,诸如痛苦、恐惧、愤怒,胜利或单纯的欢乐。有时它显然是用以激起恐怖,如某些雏鸟所发出的嘶嘶叫声。奥杜邦说,他所驯养的一只夜鹭(Ardea nycticorax Linn.)常常在一只猫来近时躲藏起来,然后“突然跳起,发出一种最可怕的叫声,显然以猫之惊慌逃走为乐”。 (353) 普通家养公鸡找到一口好吃的食物时就会咯咯地召唤母鸡,母鸡又会咯咯地召唤其小鸡。母鸡下蛋时,“频频地重复同一种叫声,以高于六度音程而截止,最后这声调持续较久”, (354) 以此来表达其欢乐。有些群居性的鸟类显然是为了寻求帮助而互相呼唤,当它们从这棵树飞往那棵树时,就靠着嘁嘁喳喳的叫声互相呼应而保持鸟群的完整。在鹅类(geese)和其他水禽类夜徙的期间,我们可以听到夜空中带头鸟所发出的响亮鸣叫和后继者相呼应的鸣叫声。某种鸣叫声是作为危险信号发出的,猎人懂得这种信号所给他带来的损失,而且这种信号能为同一种鸟和其他种鸟所理解。战胜其竞争对手之后,家公鸡便引颈长鸣,公蜂鸟也嘁嘁喳喳地鸣叫起来。然而,大多数鸟类主要是在繁殖季节才发出真正的鸣唱以及种种奇特的叫声,这是用以献媚异性,或仅仅是用以召唤异性的。
关于鸟类鸣唱的目的,博物学者们的看法有很大分歧。蒙塔古堪称最精细的观察者,但他主张“鸣禽类和许多其他鸟类的雄者一般并不寻求雌者,而正好相反,它们在春季的事务都是停息于某一显眼的地点,纵声唱出其多情的音调;对此,雌者依其本能自能领会并前往该地择其配偶”。 (355) 詹纳·韦尔先生告诉我说,夜莺的情况肯定如此。毕生从事养鸟的贝奇斯坦(Bechstein)断言,“雌金丝雀(canary)总是选择最善于鸣唱的公鸟,雌燕雀在自然状态下也是百里挑一地去选择最使它感到高兴的那些善于鸣唱的公鸟”。 (356) 毫无疑问,鸟类非常注意彼此的鸣唱。韦尔先生曾向我说过一个例子:有一只红腹灰雀(bullfinch)被教会了鸣唱一支德国圆舞曲,它演奏得那么好,以致它的身价竟达10个吉尼(guineas) (357) ;当这只鸟首次被放进一间养着其他鸟类的屋内并开始鸣唱时,所有其他鸟类,约为20只红雀(1innets)和金丝雀,都排列在各自鸟笼里最靠近它的一边,以最大的兴趣倾听这个新来客的演奏。许多博物学者都相信鸟类的鸣唱几乎完全是敌对和竞赛所引起的结果,并非为了向其配偶献媚。这就是戴恩斯·巴林顿和塞尔伯温的怀特(White)的见解,他们对这个问题都有特别的研究。 (358) 然而,巴林顿承认,“鸟类的善于鸣唱者比其他鸟类占有无比的优势,这是捕鸟人所熟知的”。
雄鸟之间在鸣唱方面肯定有激烈的竞争。玩鸟的人比赛他们所养的鸟,看哪只鸟鸣唱的时间最长,雅列尔先生告诉我说,第一流的鸟有时会一直鸣叫到坠落在地而几乎死去,或按贝奇斯坦的说法 (359) ,因肺部一条血管破裂而完全死去。不管其原因可能是什么,正如韦尔先生对我说的,雄性鸟类在鸣唱季节往往会突然死去。鸣唱的习性有时显然同爱情完全无关,因为有人曾描写过一只不育的杂种金丝雀 (360) 在镜子里见到自己形象时就鸣唱起来了,随后就向自己形象猛扑过去;把它和一只雌金丝雀关在同一个笼子时,它同样向雌鸟愤怒地进行攻击。由鸣唱行为所激起的妒忌性经常被捕鸟人所利用,把一只唱得好的雄鸟隐藏起来并加以保护,同时把一个剥制的鸟暴露在视线之内,并在其周围放置涂上粘鸟胶的小枝。正如韦尔先生告诉我的,有一个人用这种方法在一天之内所捉到的雄性欧洲苍头燕雀(Chaffinch)就有50只之多,另一次则高达70只。鸟类在鸣唱能力和鸣唱爱好方面的差异非常之大,一只普通雄性欧洲苍头燕雀的价钱虽只有6便士,但韦尔先生见到捕鸟人有一只这种鸟竟索价3镑之多。对一只真正善于鸣叫的鸟的检验方法是,把鸟笼在其主人头上转动时,它还会继续鸣唱。
雄性鸟类因竞争而鸣唱,也因献媚雌者而鸣唱,二者完全不矛盾;也许可以期待这两种习性会同时发生作用,就像夸示本身之美和好斗那两种习性同时发生作用一样。然而,某些作者争辩说,雄鸟的鸣唱不能用以迷惑雌鸟,因为某些少数物种,诸如金丝雀、知更鸟、百灵鸟(lark)和欧洲苍头燕雀的雌者,尤其当它们处于寡居的状态时,正如贝奇斯坦所说的,都会纵声唱出委婉动听的曲调。在某些这等事例中,鸣唱的习性可以部分地归因于雌鸟所受到的高度喂养和被圈禁, (361) 因为这就扰乱了同物种繁殖有关的一切正常功能。关于雄鸟的次级性征部分地传递给雌鸟,已举出过许多事例,因而某些物种的雌鸟具有鸣唱的能力就完全不足为奇了。还有人争辩说,雄鸟的鸣唱不是作为一种魅惑,因为某些物种的雄鸟,例如知更鸟,在秋季也鸣唱。 (362) 动物为了某种真实的利益在某一时期所遵循的无论什么本能,在另一时期还会以实践这种本能为乐,这是最常见的事。我们不是多么经常地见到飞行自如的鸟类显然由于取乐而在空中滑行和翱翔吗?猫戏弄捕得的鼠,鸬鹚戏弄捕得的鱼。织布鸟(Ploceus)被关进笼里时,仍在鸟笼的铁丝柱之间灵巧地编织草叶而自娱。习惯于在生殖季节相斗的鸟类一般在所有时期都准备进行战斗;雄雷鸟有时在秋季也会在它们通常集会的场所举行其巴尔森(Balzen)或勒克斯(1eks)舞会。 (363) 因此,雄性鸟类在求偶季节过后还继续鸣唱以自娱,就完全不足为奇了。
正如前章所阐明的,鸣唱在某种程度上是一种艺术,而且通过实践会大大提高。鸟类可被教会鸣唱各种不同的曲调,即使叫得难听的麻雀也曾学会像一只红雀那样地鸣唱。它们可学得养父养母的歌声 (364) ,有时也可学得邻居的歌声。 (365) 所有普通的鸣禽都属于燕雀类(Insessores)这一目,它们的发音器官要比大多数其他鸟类的发音器官复杂得多;但也存在一个奇特的事实,即,某些燕雀,诸如渡鸦、乌鸦和喜鹊,虽然从来不鸣唱,自然也不会发出抑扬的音调,但它们都具有这种正规的发音器官。 (366) 亨特断言 (367) ,关于真正的鸣禽,其雄鸟的喉肌都比雌鸟的更为强有力;但除这点轻微的差异外,雌雄二者的发声器官并无任何差别,尽管大多数物种的雄鸟鸣唱起来要比雌鸟好听得多,而且更加连绵不断。
值得注意的是,彻底善于鸣唱者皆为小型鸟。然而澳洲的琴鸟属(Menura)必须除外,因为,像半成熟火鸡那样大小的阿氏琴鸟(Menura Alberti)不仅模仿其他鸟类鸣叫,而且“它自己的啭鸣声也极其美妙而富有变化”。其雄鸟集合起来组成“科罗伯瑞舞场(corroborying places) (368) ”,它们在那里鸣唱,像孔雀那样地高举并展开其尾羽,同时双翅下垂。 (369) 同样值得注意的是,善于鸣唱的鸟类很少具有鲜艳的色彩或其他装饰物。以我们英国的鸟类来说,欧洲苍头燕雀和金翅雀除外,最善于鸣唱的都是色彩平淡的。鱼狗(King-fisher)、蜂虎(bee-eater)、德国佛法僧(roller)、戴胜(hoopoe)啄木鸟等都发出刺耳的叫声;并且色彩鲜艳的热带鸟类几乎都不是善于鸣唱者。 (370) 因此,鲜明的色彩和鸣唱的能力似乎是可以互相取代的。我们可以看到如果羽衣色彩不变得鲜明,或者鲜明的色彩危及物种的生存,那么就可能采用其他手段来魅惑雌鸟,而悦耳的音调就提供了这样的手段。

图 39 狂热松鸡(Tetrao cupido)雄者
(采自伍德)
某些鸟类雌雄二者的发音器官差异很大。狂热松鸡(图39)的雄者在其颈部两侧各有一个无毛的橙色囊,这种囊在繁殖季节即行膨大,此时雄鸟便发出奇妙的空洞叫声,在相当的距离以外都可听到。奥杜邦证实说,这种叫声同这等器官密切关联(这使我们回想起某些雄蛙在嘴的两侧各有一个气囊),因为他发现如果一只驯养的这种鸟的一个囊被刺破,其叫声就大大减弱,如果两个囊都被刺破,叫声则完全丧失。雌鸟的“颈部也有一块多少相似的、虽然稍微小一些的裸皮,但它不能膨胀”。 (371) 另一种细嘴松鸡(Tetrao urophasianus)的雄者向雌者求爱时,其“黄色的无毛食管膨胀得非常之大,足有其躯体的一半”;于是它发出各种嘎嘎的、深沉而空洞的声调。是时颈羽竖起,双翼低垂,跑来跑去,其长而尖的尾羽展开有如一把扇子,表现了种种奇形怪状。而这种雌鸟的食管则无任何值得注意的地方。 (372)
现在似乎已弄清楚,欧洲雄性大鸨(Otis tarda),至少还有其他四个物种的雄鸟,它们的大喉袋并非以前所设想的那样用以存水,而是同繁殖季节中所发出的那种类似“喔克(oak)”的特殊叫声有关系。 (373) 一种栖息于南美的形似乌鸦的伞鸟(Cephalopterus ornatus,图40)以其覆盖了整个头部的巨大顶结而被称为伞鸟(umbrella-bird),这种顶结是由羽毛形成的,羽根呈白色,裸露无毛,其顶端则生有暗蓝色的羽毛,它们竖起来便形成直径不下于五英寸的一个大圆顶。这种鸟的颈部有一条长而细的圆筒状肉质附器,其上厚厚地被覆着一层鳞片状的羽毛。它大概一部分用为装饰物,同时也用为一种回声器,因为贝茨先生发现它“同气管和发音器官的异常发育”有关系。当这种雄鸟发出那种独特的深沉、高昂而持久的嘹亮声调时,肉质附器就膨胀起来了。而雌鸟的羽冠和颈部附器则处于残迹状态。 (374)

图 40 伞鸟(Cephalopterus ornatus)的雄者
(采自布雷姆)
各种蹼足鸟类和涉禽类的发音器官极为复杂,且在雌雄二者有一定程度的差异。在某些场合中,气管是盘旋的,像支弯管乐号,且深深嵌入胸骨内。关于野天鹅(Cygnus ferus),其成熟雄者气管所嵌入的程度比成熟雌者和年青雄者更深。雄秋沙鸭(Merganser)的气管扩大部分具有一对附加的肌肉。 (375) 然而,有一种叫做斑点鸭(Anas punctata)的,其雄者的骨质扩大部分仅比雌者的稍微发达一点。 (376) 但鸭科(Anatidae)雌雄二者在气管方面的这等差异,其意义何在,尚属不明;因为雄鸭并不总是喧叫得更凶;例如,普通家鸭,其雄者不过是嘶嘶地叫,而雌者却大声嘎嘎地叫。 (377) 有一种小鹤(Grus virgo),其雌雄二者的气管均深陷胸骨中,但表现了“某种性别变化”。雄黑鹳(black stork)在支气管的长度和弯曲度两方面都表现了十分显著的性差异。 (378) 因此,在这等场合中,如此高度重要的构造也因性别而发生了变异。
雄性鸟类在繁殖季节所发出的许多奇妙的叫声和音调,究竟是用以魅惑雌鸟,或仅仅作为一种召唤,是难以猜测的。或可假定,鸠和许多鸽类的柔和咕咕叫声是取悦雌者。雌野火鸡在早晨鸣叫时,雄鸡则答以一种不同于咯咯叫声的音调,同时竖起羽毛,沙沙地抖动翅膀鼓起垂肉,并在雌鸡面前高视阔步并噗噗喷气。 (379) 雄黑松鸡的鸟语(spel)肯定是用以召唤雌者的,因为,据知一只关在笼中的雄者曾用这种鸟语把四五只雌者从远处召来。但是,由于雄黑松鸡连续做此鸟语达数小时并持续数日不断,并且在雷鸟的场合中还“伴以炽烈的激情”,因此这就使我们设想那些光临的雌者是这样被迷住了。 (380) 据知,普通秃鼻乌鸦(rook)的叫声到繁殖季节就会改变,所以从某一方面来看这是性的变化。 (381) 然而关于那种刺耳的尖叫声,如某些种类的金刚 鷱(macaws)的叫声,我们又该怎样说呢;这些鸟类的不善于欣赏音乐声调是否就像它们不善于欣赏色彩那样呢?根据它们的羽毛呈鲜黄色和蓝色这种不协调的颜色对比,可以判断它们欣赏色彩的能力显然是低劣的。从这样叫声不会获得任何利益,的确是可能的,许多雄性鸟类的高声鸣叫可能是由于它们当受到强烈的爱情、嫉妒和愤怒等感情的刺激时连续使用其发音器官的遗传效果所造成的;但关于这一点,在我们讨论四足兽时还要谈到。
鷱(macaws)的叫声,我们又该怎样说呢;这些鸟类的不善于欣赏音乐声调是否就像它们不善于欣赏色彩那样呢?根据它们的羽毛呈鲜黄色和蓝色这种不协调的颜色对比,可以判断它们欣赏色彩的能力显然是低劣的。从这样叫声不会获得任何利益,的确是可能的,许多雄性鸟类的高声鸣叫可能是由于它们当受到强烈的爱情、嫉妒和愤怒等感情的刺激时连续使用其发音器官的遗传效果所造成的;但关于这一点,在我们讨论四足兽时还要谈到。
到目前为止,我们所谈论的仅仅是鸟类的发声,但各种鸟类的雄者在其求偶期间所发出的鸣叫都可称之为器乐。孔雀和极乐鸟收拢其羽根,格格作响。雄火鸡以翼擦地,发出沙沙响声,松鸡的某些种类也这样发声。另一种北美松鸡叫伞松鸡,当它将尾羽竖起时,其颈羽即行张开,“向藏在附近的雌鸟夸耀其美”,按照海蒙德(R.Haymond)先生的材料,这时它用双翼急击其背而发出鼓声,并非像奥杜邦所想的是以双翼击其躯体两侧。这样发出的声音,有些人把它比作远方的雷声,另外有些人则把它比作快速擂鼓之声。雌鸟从不发出这样鼓声,“但它径直飞往雄鸟发出这种声音的场所”。喜马拉雅山的黑鹇(kalij-pheasant)的雄者,“常常以其双翅发出一种独特的鼓声,就像摇动一张僵硬的布块所产生的那种声音”。非洲西海岸的小型黑色织布鸟常常小群地在围绕一小块空地的灌木丛中聚会,又是鸣唱,又是以其抖动着的双翅在空中滑翔,“这样弄出来的一种急速转动的呼呼声,犹如一个小孩喋喋不休的语声”。一只鸟跟着一只鸟这样地进行表演,达数小时之久,但这种情况仅仅在求偶季节才发生。某些欧夜鹰属(Caprimulgus)的雄者只在这个季节,而不是在另外的时候,才用其双翅奏出一种奇特的隆隆声。啄木鸟的各个物种用喙敲击树枝而发出一种响亮的声音,敲击时头部的往复动作是如此迅速,以致“在一瞬间它的头好像是在两处”。这样发出的声音可在相当远的地方听到,但无法被描写出来;而且我可以肯定,任何人首次听到这种声音时,都不会猜出它的声源在何处。由于这种刺耳的声响主要是在生殖季节发出的,因而它被认为是一种爱情之歌;但更严格地说,这也许是一种爱情的呼唤。当把雌鸟从巢里赶出来时,曾经观察到雌鸟就这样呼唤其配偶,后者以同样方式应答并很快出现在雌鸟的面前。最后,雄戴胜(Upupa epops)会把声乐和器乐结合起来;因为,正如斯温赫(Swinhoe)先生所观察的,在生殖季节,这种鸟先吸进空气,然后用喙端垂直地在一块石头或一棵树干上轻轻敲击,“这时使劲地把气从其管状喙呼出,于是就产生了正确的声音”。如果不用喙这样敲击某些物体,所发出的声音就完全不一样。同时吸进空气,食管也因而大为膨胀:这大概起了一个回声器的作用,不仅戴胜如此,鸽类和其他鸟类也都如此。 (382)

图 41 普通丘鹬(Scolopax gallinago)的外侧尾羽
(采自《动物学会学报》,1858年)
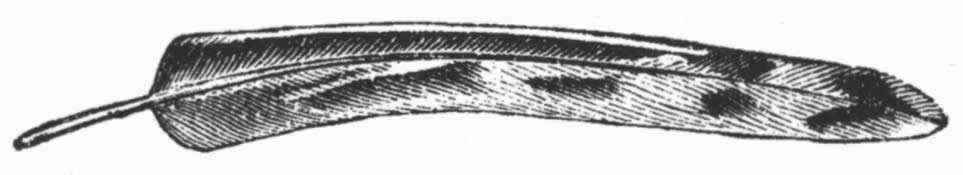
图 42 马缰丘鹬(Scolopax frenata)的外尾羽
上述事例所表明的是,声音的产生系借助于早已存在的而且在其他方面所必需的构造;但下述事例所表明的却是,某些羽毛乃是为了发声的特殊目的而发生变异的。普通丘鹬(Scolopax gallinago)所发出的鼓声、羊叫声、马嘶声或雷鸣声(不同的观察家对此有不同的表达),无论何人听到,都一定要感到惊奇。这种鸟在交配季节飞到“也许高达一千英尺的天空”,弯弯曲曲地飞了一会儿之后,就展开尾羽,抖动着双翼,以惊人的速度沿着一条曲线降落地面。只有在这样快速降落时它才能发出这种响声。这样发声的原因,在梅费斯(M.Meves)以前没有一个人能加以说明,直到梅费斯才观察到:其尾部两边的外侧羽毛具有特殊的构造(图41),羽轴坚硬,呈马刀形,羽轴上的斜羽枝极长,羽枝外侧的短毛紧紧结合在一起。他发现如果吹动这些羽毛,或把它们绑牢在一条长而细小棍上,迅速在空中挥动,就可重现这种活鸟所发出的那样鼓声。其雌雄二者都具有这种羽毛,但雄者的一般要比雌者的大些并发出一种更为深沉的声调。某些物种,如马缰丘鹬(S.frenata),其尾部两侧各有四支羽毛大大变异了(图42),而爪哇丘鹬(S.javensis)尾部两侧大大变异了的这种羽毛(图43)则不少于8支。把不同物种的这等羽毛在空中挥动时,就发出不同的音调;美国的韦氏丘鹬(Scolopax wilsonii)当快速向地面降落时所发的声音有如挥鞭。 (383)

图 43 爪哇丘鹬(Scolopax javensis)的外尾羽

图 44 亮羽蜂鸟(Selasphorus platycercus)的初级翼羽(采自沙尔文先生的绘图)
上图为雄者的;下图为雌者的。
美洲所产的鹑鸡类一种大型鸟,叫做单色镰翅冠雉(Chamaepetes unicolor)的,其雄者的第一初级翼羽顶端弯曲,而且远比雌者的为细。有一种亲缘相近的鸟,即Penelope nigra,沙尔文(Salvin)先生观察到一只这种雄鸟在往下飞时“展开双翼,发出一种冲击折裂之声”,犹如一棵树倾倒的声音一般。 (384) 印度的耳鸨(Sypheotides auritus)只有雄者的初级翼羽才大大变尖;据知有一个亲缘相近的物种,其雄者当追求雌者时会发出一种嗡嗡之声。 (385) 在鸟类的一个大不相同的类群中,即蜂鸟,某些种类只有雄者的初级翼羽的羽轴膨大甚阔,或其羽枝在近尖端处陡然削细。例如,亮羽蜂鸟(Selasphorus platycercus)这种蜂鸟的雄者到成年时,其初级翼羽的顶端就这样削细(图44)。它在花丛中飞来飞去时发出“一种尖锐的几乎同口哨一样的声音”; (386) 但沙尔文先生并不认为这种声音是有意发出的。
最后,美洲产燕雀类小鸟(Pipra或Manakin)的一个亚属的几个物种,正如斯克莱特(Sclater)先生所描述的,其雄者的次级翼羽以更加显著的方式发生了变异。色彩鲜艳的P.deliciosa,有3支第一次级翼羽的羽茎变粗,且向体部弯曲;第四和第五支次级翼羽(图45,a)的变化还要更大些;而第六和第七支次级翼羽(图45,b,c)的羽轴则“加粗到异常的程度并形成一个坚固的角质块”。羽枝的形状同雌鸟的相应羽枝(图45,d,e,f)相比,也大大改变了。甚至支撑着这些独特羽毛的翼骨,据弗雷泽说,在雄鸟方面也要粗得多。这种小型鸟类发出一种异常的声响,其头一个“尖锐的声调就像抽鞭子那样的噼啪声”。 (387)

图 45 Pipra deliciosa的次级翅羽(采自斯克莱特先生,见《动物学会会报》,1860年)
上面三根羽毛a,b,c系采自雄者;下面三根相应的羽毛d,e,f系采自雌者。a和d,为雄者和雌者第五次级翅羽的上表面。b和e,为第六次级翅羽的上表面。c和f,为第七次级翅羽的下表面。
许多鸟类的雄者在生殖季节所发出的声乐和器乐声音的多样性,以及发出这些声音的方法的多样性,都是高度值得注意的。这样,我们对于它们在性的用途上的重要性便提高了认识,并由此可以回忆到从昆虫类所得出的结论。不难想象鸟类发声所经历的步骤有如下述:它们的声调最初仅仅是作为一种召唤或用于某种其他目的,继而可能改进成为一种有旋律的爱情歌唱。在鸟类用变形的羽毛发出鼓声、口哨声或轰鸣声的场合中,我们知道有些鸟类在求偶期间会拍击、抖动其未变形的羽毛或使它们嘎啦嘎啦地作响;如果雌鸟被引导去选择那些最佳的表演者,那么在躯体任何部分具有最坚固或最厚密或最尖细的羽毛的那些雄鸟大概是最能成功的;这样,其羽毛大概就会以缓慢的程度发生几乎任何程度的改变。当然,雌鸟不会注意到其形状的每个细微的连续改变,而只是注意到由此所产生的声音。奇怪的是,在这同一个动物纲中,其声音是如此不同,诸如鹬的尾巴所发出的鼓声,啄木鸟的喙所发出的笃笃轻敲声,某些水禽粗里粗气的类似喇叭的叫声,斑鸠的咕咕声,以及夜莺的歌唱,却全会取悦于若干物种的雌鸟。但我们决不能用一个一致的标准去衡量不同物种的欣赏能力;而且也不能用人类的欣赏标准去做这种衡量。我们应该记住,即使对人类来说,有些不协调的声音,如铜锣的锣声和芦笛的刺耳声,都会使未开化人感到悦耳动听。贝克(Baker)爵士说, (388) “如同阿拉伯人的胃喜爱刚从动物身上取下的热乎乎的生肉和冒着热气的肝脏一样,他的耳朵也喜爱同样粗俗而不谐和的音乐,却不喜欢听所有其他音乐”。
求爱的滑稽表演和舞蹈
有些鸟类的奇特求爱姿态已经顺便提到过了,因此在这里不需多加补充。在北美有一种松鸡叫做尖尾松鸡(Tetrao phasianellus),在生殖季节的每天早晨,它们成群地在某个选中的平坦地点相会,沿着直径约15或20英尺的圆圈,一圈又一圈地奔跑,因而地面被踩得光秃秃的,犹如一个仙环(蘑菇圈) (389) 。猎人称此为石鸡舞(partridge-dances),在这等鸟舞中,鸟类表现了最奇特的姿态,它们跑圆圈,有的向左,有的向右。奥杜邦描述过一种苍鹭(Ardea herodias)的雄者,它们在雌者面前迈着长腿非常威严地走来走去,显示着对其他竞争对手的蔑视。关于令人讨厌的一种吃腐肉的兀鹫(Cathartes jota),同一位博物学家说道,“其雄者在求爱季节开始时所作出的姿态和炫耀都是极其滑稽可笑的”。某些鸟类求爱的滑稽表演是在飞翔中而不是在地面上进行的,如我们所见过的非洲黑织布鸟就是如此。英国小白喉雀(Sylvia cinerea)春天常在某些灌木上空几英尺或几码高的地方飞翔,“它以一种间歇的古怪动作拍着翅膀,并鸣唱不已,然后落到它的栖木上”。英国大鸨当追求雌者时作出一种无法形容的奇特姿态,沃尔夫曾绘过它的图。在这样时期,亲缘相近的孟加拉鸨(Otis bengalensis)则“急拍双翼,垂直地高飞入空,竖起羽冠,鼓起颈羽和胸羽,然后落至地面”;这等表演要重复几次,同时哼出一种特殊的声调。那些碰巧在近旁的雌鸟“听从了这种舞蹈式的召唤”,等它们一到,雄鸟就像雄火鸡那样地拖着双翼,并展开尾羽。 (390)
但最为奇特的例子乃得自澳大利亚鸟类三个亲缘相近的属,即著名的造亭鸟(Bowerbirds)——它们无疑是某些古代物种的共同后裔,这些物种最先获得了造亭的奇异本能以进行其求爱的滑稽表演。这等用羽毛、贝壳、骨头和叶子装饰起来的亭子(图46),正如我们以后就要见到的,系建于地面之上,其唯一用途是为了求偶,因为它们的巢是筑在树上的。雌雄二者在造亭上互相帮助,但雄鸟是主要的劳动者。这等本能是如此强烈,甚至在圈养的条件下也照样造亭,斯特兰奇(Strange)先生曾描述过他在新南威尔士一间鸟舍里所养的一些萨丁造亭鸟(Satin bower-birds)的习性。 (391) 他说:“雄鸟不时在鸟舍里到处追逐雌鸟,然后向亭子走去,啄起一根华丽的羽毛或一张大的叶片,发出一种奇异的叫声,把全身羽毛竖起,绕亭奔跑并变得如此激动,以致它的双眼好像就要从头部迸出;它不断地举起一翼,然后再举起另一翼,发出一种低沉的哨声,并像家养的公鸡那样,好像从地上啄到了什么东西,这样的表演一直要继续到最后雌鸟向它温柔地走去为止。”斯托克斯(Stokes)船长描写过另一个物种即大型造亭鸟的习性及其“游戏室”,他见过这种鸟“飞前飞后,轮流从贝壳的每一边把它衔起,并用嘴把它带过拱道,借此以自娱”。这等奇异建筑乃专门作为聚会场所之用,雌雄二者皆集此取乐,并进行求偶,造此建筑一定要花费这种鸟的巨大劳动。例如,一个胸部淡黄褐色的物种所造的亭子,长度近四英尺,高度近18英寸,而且是在一层厚厚的树枝所铺成的平台上建造起来的。

图 46 造亭鸟(Chlamydera maculata)及其所造之亭(采自布雷姆)
装 饰
我首先要讨论的事例是,雄鸟的装饰物为它所专有,或者其装饰程度远远高于雌鸟,在下一章所讨论的事例是,雌雄具有同等的装饰,最后所讨论的事例是比较罕见的,即雌鸟的色彩多少比雄鸟的更为鲜艳。鸟类的天然装饰物也如同未开化人和文明人所使用的人工装饰物一样,其主要装饰部位是在头部。 (392) 这等装饰物,如本章开头所说的,极其多种多样。头的前部或后部的羽饰有各种不同的形状,有时能竖起或展开,借此来把漂亮的色彩充分显示出来。耳部偶尔生有漂亮的簇毛(见前面图39)。头部有时像雉那样地被以天鹅绒般的柔毛;或裸露无毛而具有生动的色彩。喉部有时也装饰着一把胡子、垂肉或肉瘤。这等附器一般都是色彩鲜明的,虽然它们在我们眼里并不总是那么有装饰性,但无疑是作为装饰之用的;因为当雄鸟向雌鸟进行求偶时,这等附器往往胀大并呈现生动的色彩,就像雄火鸡的情况那样。在这种时候,雄角雉(Ceriornis temminckii)头部周围的肉质附器就膨胀起来而成为喉部一个大垂肉和两只角,这两只角分别位于其漂亮顶结的两边;这时这等附器便呈现我有生以来所见过的最浓烈的蓝色。 (393) 非洲犀鸟(Bucoraxabyssinicus)鼓起颈部的深红色囊状垂肉,低垂双翼并展开尾羽,“形成了十分雄壮的外观”。 (394) 甚至雄鸟的眼球虹膜,其颜色有时也比雌鸟的更为鲜艳;鸟喙的情况也常常是这样,例如,我们英国的普乌鸫即然。还有一种犀鸟(Buceros corrugatus)的雄者,其整个喙部和巨大头盔的色彩都比雌者的更为显著;而“下颚两侧的斜沟,乃雄鸟所特有的”。 (395)
此外,头部还往往支持着肉质附器、丝状物以及坚固的突起物。这等附器若非雌雄二者所共有,则总是只限于雄者所有。马歇尔博士对这等坚固的突起物做过详细描述,他指出它们或是由包在皮里面的松质骨所形成或是由上皮组织和其他组织所形成。 (396) 哺乳动物的真角永远生于额骨之上,但在鸟类,各种骨都为了这一目的而发生了变异;而在属于同一类群的诸物种中,这种突起物可能具有骨髓,也可能完全没有,在这两个极端之间有一系列的中间级进把它们连接起来。因此,正如马歇尔博士所正确指出的,种类最不相同的变异通过性选择为这等装饰附器的发展作出了贡献。延长了的羽毛、即羽饰几乎发生于躯体的各个部分。喉部和胸部的羽毛有时发展成美丽的轮状绉领和项圈。尾羽常常增加了长度,如同我们看到的孔雀尾部覆羽(tail-coverts)以及锦雉(Argus pheasant)尾部本身的羽毛就是如此。至于孔雀,甚至其尾骨也发生了改变以支持沉重的尾部覆羽。 (397) 锦雉的躯体并不大于家鸡,但从其喙端到尾端的长度却不下于5英尺3英寸, (398) 而其饰以美丽眼斑的次级翅羽,其长度也将近三英尺。一种非洲小型夜鹰(Cosmetornis vexillarius)在生殖季节有一支初级翼羽长达26英寸,而该鸟本身的长度才仅仅10英寸。在另一个亲缘相近的夜鹰属中,其延长了的翅羽的羽干除末端着有圆盘羽毛外全都裸露无毛。 (399) 此外,另一个夜鹰属中,其尾羽的发达甚至还要惊人。一般说来,尾羽的延长往往比翼羽为甚,因为翼羽的任何过分延长都会有碍飞翔。这样,我们就可看见在亲缘密切相近的鸟类中雄鸟通过大不相同的羽毛的发育而获得了同类的装饰物。
有一个奇妙的事实:属于很不相同的类群的物种,其羽毛却按照几乎完全一样的特殊方式进行改变。例如上述一种夜鹰,其翼羽的羽干都是裸露无毛的,至其末端才着生圆盘羽毛,有时人们称它为勺状羽毛或球拍状羽毛。这种羽毛在摩特鸟(Eumomota superciliaris)、鱼狗、燕雀、蜂鸟、鹦鹉、几种印度庄哥鸟(卷尾贝鸟属,Dicrurus和毛虫鵙属,Edolius,其中之一的圆盘羽毛与羽干成直角)以及某些极乐鸟类的尾部也有发生。极乐鸟的头部也装饰着相似的羽毛,其上有美丽的眼斑,某些鹑鸡类的鸟也有这种情况。有一种印度耳鸨(Sypheotides auritus),组成其耳簇的羽毛约4英寸长,其末端也着有圆盘羽毛。 (400) 正如沙尔文先生所明确指出的 (401) ,最独特的一个事实是,摩特鸟啄去羽枝使其尾羽呈球拍状,而且进一步指出,这种不断的自残行为产生了某种程度的遗传效果。
此外,在各种大不相同的鸟类中,羽枝呈丝状或羽毛状,诸如某些苍鹭类、彩鹳类(ibises)、极乐鸟类以及鹑鸡类都是如此。在其他场合中,羽枝消失了,整个羽干全部裸露无毛;而阿波达极乐鸟(Paradisea apoda)尾部的这等裸羽干竟达34英寸长; (402) 在巴布亚极乐鸟(P.papuana图47),这等裸羽干就短得多而且细得多。这样没有羽枝的小羽毛看来就像火鸡胸部的鬃毛一般。正如人类赞赏时装的飞速变换那样,雄鸟羽毛在构造或色彩上的任何一种变化似乎也会受到雌鸟的赞赏。在大不相同的类群中,羽毛按照相似的方式发生改变这一事实,无疑主要决定于所有羽毛都具有几乎相同的构造和发育方式,因而有按照同样方式发生改变的倾向。在我们那些属于不同物种的家养品种中,我们常常看到它们的羽毛有一种发生相似变异的倾向。例如,若干物种都生有顶结。有一个已绝灭的火鸡变种,其顶结是由裸露无毛的羽翮形成的,其顶端着生柔软的绒羽,因而多少同上述球拍状羽毛相类似。在鸽和鸡的某些品种中,羽毛呈丝状,羽干有某种裸化的倾向。塞瓦斯托波尔(Sebastopol)鹅的肩羽大大延长了,蜷曲,甚至呈螺旋状,其边缘为丝状。 (403)

图 47 巴布亚极乐鸟(Paradisea papuana)(引自T.W.伍德)
关于色彩,几乎不需在此多谈了,因为,每个人都知道许多鸟类的色彩是多么华丽,而且,这等色彩的配合又多么协调。鸟类的颜色往往具有金属的和彩虹的光泽。在圆点的周围有时环以一层或多层浓淡不同的色带,因而变成了眼斑。关于许多鸟类雌雄二者在颜色方面的巨大差异,也无须多赘。普通孔雀提供了一个显著的事例。雌极乐鸟的色彩暗淡而且缺少任何装饰物,反之,雄极乐鸟大概是所有鸟类中最精于装饰者,其装饰如此多种多样,以致见者无不赞叹。在阿波达极乐鸟双翼下生出来的金橙色长羽,当垂直竖起并使之颤动时,有人把这种情景描写成犹如形成了一种太阳晕轮,位于中央的头“看去就像一个由绿玉做成的小太阳,其光线乃是由两支羽毛形成的”。 (404) 另有一个最美丽的物种,其头部却是秃的,“具有一种鲜艳的钴蓝色,其上有几道横穿而过的天鹅绒般的黑色羽毛”。 (405)
雄蜂鸟(图48和图49)之美几乎可与极乐鸟相匹敌,凡是见过古尔德先生的佳作或其丰富采集品的人都会承认这一点。很值得注意的是,这些鸟类的不同装饰方法是何等之多。它们羽毛的几乎每一部分都被利用了,而且发生了变异;在属于几乎每个亚群的一些物种中,正如古尔德先生向我指出的,这种变异已达到了令人吃惊的极端。这等情况同我们看到的人类为了装饰所培育出来的那些观赏品种的情况非常相似;某些个体最初在某一性状上发生了变异,而同一物种的其他个体则在其他性状上发生了变异;人类抓住了这等变异,并把它们大大地加以扩充——如扇尾鸽的尾羽、毛领鸽(jacobin)的羽冠、信鸽的喙和垂肉等等表明了上述一点。这两类事例之间的唯一不同之处在于:一方面是由于人类选择的结果,而另一方面,如蜂鸟类、极乐鸟类等,乃是由于雌者选择了比较美丽的雄者的结果。

图 48 花冠蜂鸟(Lophornis ornatus),雄鸟和雌鸟(采自布雷姆)

图 49 长尾蜂鸟(Spathura underwoodi),雄鸟和雌鸟(引自布雷姆)
我只再谈谈另一种鸟,它是以雌雄二者色彩的极其强烈对照而闻名的,这就是著名的南美铃鸟(Chasmorhynchus niveus),远在3英里左右尚可辨别其鸣声,每一个人最初听见它的鸣声时,无不感到惊奇。其雄鸟呈纯白色,其雌鸟呈暗绿色;而白色在中等大小和没有侵害习性的陆栖物种中,是很罕见的色彩。正如沃特顿(Waterton)所描述的,这种雄鸟还有一个3英寸左右的螺旋形管从喙的基部伸出来。它的颜色漆黑,点缀着微细的绒毛。此管和腭相通,可充气膨胀,不膨胀时则挂在一边。这个属包含四个物种,其雄鸟很不相同,而雌鸟则如斯克莱特先生在一篇很有趣的论文中所描述的那样,彼此密切相似,于是这向我们提供了有关共同规律的一个最好例证,即,在同一类群中雄者相互的区别远比雌者相互的区别为大。在第二个物种裸颈铃鸟(C.nudicollis)中,雄鸟同样是雪白色的,只有喉部和眼睛周围的一大块裸皮除外,这种裸皮在生殖季节呈艳绿色。在第三个物种三丝铃鸟(C.tricarunculatus)中,雄鸟的头部和颈部都是白色的,而躯体的其余部分则呈栗褐色,这个物种的雄者有三条丝状突起物,其长度为其躯体的一半——一条从喙的基部伸出来,另两条从嘴的两角伸出来。 (406)
成年雄鸟的彩色羽衣和某些其他装饰物或保持终生,或在夏季和繁殖季节定期地更新。在繁殖季节里,它们的喙及其头部周围的裸皮也常常改变颜色,诸如某些苍鹭类、鹳类、鸥类以及刚刚提到的一种铃鸟等都是如此。白彩鹳的双颊、喉部能鼓起的皮肤以及喙的基部在那时都变为艳红色。 (407) 有一种秧鸡,叫做冠董鸡(Gallicrex cristatus),在这期间其雄者的头顶有一块红色大肉冠发育起来了。红嘴鹈鹕(Pelecanus erythrorhynchus)喙上的一个薄角质突起也是如此;因为,在生殖季节过后,这种薄角质突起就像雄鹿头上的角那样地脱落了,有人发现在内华达州一个湖中小岛的岸边上,布满了这种脱落下来的奇异残骸。 (408)
羽衣色彩的季节性变化决定于:第一,每年两次的换羽,第二,羽毛自身色彩的实际变化,第三,暗色羽毛边缘的周期性脱落,或者,决定于这三个过程或多或少的结合。暂时性羽毛边缘的脱落可同其幼鸟绒毛的脱落相比拟。因为在大多数场合中,绒毛是由第一真羽的顶端长出来的。 (409)
关于每年两次换羽的鸟类,可分下列五种:第一,雌雄二者彼此相类似,而且无论在什么季节它们的毛色都不改变,如鹬类、燕鸻类(swallow plovers,Glareolae)和杓鹬类(curlews)皆是。我不知道它们的各羽是否要比夏羽厚些和暖和些,不过毛色既不改变,保暖似乎是换羽两次的最可能的目的。第二,雌雄二者相类似,但冬羽同夏羽稍有差异,如红脚鹬(Totanus)和其他涉禽类的某些物种皆是。然而冬羽和夏羽的差异如此轻微,以致这种差异几乎不会给它们带来任何益处;也许,这可能是这些鸟类在两个季节中所处的不同条件直接作用的结果。第三,还有许多其他鸟类,其雌雄二者彼此相类似,但夏羽和冬羽则大不相同。第四,有些鸟类,其雌雄二者在色彩上彼此不同,但雌者虽两次换羽却整年都保持着同样的色彩,而雄者则经历色彩的变化,有时变化颇大,如某些鸨类就是如此。第五,也是最后一种,有些鸟类,其雌雄二者无论夏羽和冬羽均彼此不同;但雄者在周而复始的每个季节里所经历的色彩变化,其程度要比雌者为大——在这方面流苏鹬(Machetes pugnax)提供了一个良好的事例。
关于夏羽和冬羽在色彩上相异的原因或目的,在某些事例里,例如雷鸟, (410) 可能在两个季节中都是作为一种保护之用的。倘若夏羽和冬羽的差异轻微,这也许像已经说过的,可能是生活条件直接作用的结果。但对许多鸟类来说,夏羽作为装饰品几乎是无可怀疑的,即使雌雄二者彼此相类似,也是如此。我们可以断定,许多苍鹭类、白鹭类等的情况都是如此,因为它们只在繁殖季节才获得美丽的羽饰。还有,这等羽饰、顶结等,虽为雌雄二者所具有,但雄者的比雌者的偶尔稍为发达,而且雄者的羽饰和装饰物只同其他鸟类雄者所具有的相类似。我们还知道圈养可以影响雄性鸟类的生殖系统,因为圈养常常会抑制其次级性征的发育,但对其他任何性征却没有直接的影响;巴特利特先生告诉我说,在动物园中圈养的漂鹬(Tringa canutus)有八九个样本整年保持其不加装饰的冬羽,根据这一事实,我们可以推论许多其他鸟类的夏羽虽为雌雄二者所共有,却带有完全的雄性性质。 (411)
根据上述事实,尤其是根据某些鸟类无论雌雄在每年任何一次换羽时都不改变色彩,或改变得如此轻微以致这种改变对它们几乎没有任何用处,以及根据其他物种的雌者虽两次换羽却还整年保持着同样的色彩,我们可以断定,鸟类所获得的每年换羽两次的习性并非为了雄鸟在繁殖季节呈现一种装饰的性状,而是原先为了某种不同目的所获得的两次换羽习性,后来在某些场合中为了取得婚羽而被利用了。
某些亲缘相近的物种有规律地经历每年两次换羽,而其他物种只是一年换羽一次,最初一看,这似乎是一个令人惊奇的情况。例如,雷鸡每年换羽两次甚至三次,而黑琴鸡每年仅换羽一次。印度某些色彩华丽的花蜜鸟类(honey-suckers,太阳鸟类Nectariniae)以及色彩暗淡的鹨(Anthus)的某些亚属一年换羽两次,而其他仅每年换羽一次。 (412) 但是,已知各种鸟类换羽方式的种种级进向我们表明:鸟类的物种或整个类群最初怎样获得了其每年两次换羽的习性,或者怎样一度获得了这种习性而后又失掉了。某些鸨类和鹬类的春季换羽远远是不完全的,有些羽毛更新了,有些羽毛颜色改变了。还有理由可以相信,关于正常换羽两次的某些鸨类和形似秧鸡的鸟类,其较老的雄鸟有些整年保持其婚羽不变。在春季可能只有少数高度变异的羽毛增添于羽饰之中,如某些印度卷尾鸟(Bhringa)的圆盘形尾羽,以及某些苍鹭背部、颈部、胸部上的延长羽毛,就是如此。按照上述这些步骤,就可使婚羽的脱换越来越完全,直到最终便获得完全的换羽两次的习性。有些极乐鸟类整年保持其婚羽,这样就仅换羽一次;另外一些极乐鸟类在繁殖季节过后其婚羽就立即脱落,这样,便进行换羽两次;还有其他极乐鸟的婚羽只在头一年的繁殖季节脱落,此后则不,所以后面这些物种的换羽方式乃处于中间地位。许多鸟类每年保持这两种羽衣的期限长短也大不相同,所以其中一种羽衣也许保持全年,而另一种羽衣就完全消失了。例如,流苏鹬在春季只能把它的颈羽保持两个月。在纳塔尔(Natal),雄黑尾长羽鸟(Chera progne)在十二月和正月获得其美丽的羽衣和长尾羽,一到三月它们就脱落了,因此它们大约只保持3个月。换羽两次的大多数物种可把其装饰性的羽毛保持6个月左右。然而,野生原鸡(Gallus bankiva)的雄者可把其颈部长羽保持9个月或10个月;当这等羽毛脱落后,下面的黑色颈羽就充分显现出来了。但是,就这个物种的家养后裔来说,雄鸟的颈部长羽马上就会被新羽所置换,因此在这里我们看到,羽衣的一部分由两次脱换变为在家养状况下的一次脱换了。 (413)
众所周知,普通公鸭(Anas boschas)在繁殖季节过后就失去其雄性羽衣达3个月之久,在这期间它的羽衣同雌鸭的一样。雄针尾鸭(Anas acuta)失去其雄性羽衣的时间要短些,为6周或2个月。蒙塔古说:“在这么短的时间内换羽两次是一种最异常的情况,这似乎是向人类的一切推理进行挑战。”但是,相信物种渐变的人在发现所有级进类型时决不会感到惊奇。如果雄尖尾鸭在更短的期间内获得其新羽衣,那么,新的雄性羽毛几乎必然要和旧的羽毛混在一起,而且新旧羽毛又会和雌者所固有的羽毛混合在一起;一种亲缘不远的鸟,即红胸秋沙鸭(Merganser serrator)的情况就是如此,因为,据说其雄者“经历了羽衣的一种变化,从而使它的羽衣变得同雌者的有几分相像”。这个过程稍微再加快一点,则换羽两次就会完全消失了。 (414)
如上所述,有些雄鸟的色彩在春季变得更为鲜明,并非由于春季换羽,而是由于其羽色发生了实际变化,要不,就是由于其色彩暗淡的暂时性羽毛边缘脱落了。如此而引起的色彩变化所持续的时间可能有长有短。在春季,白鹈鹕(Pelecanus onocrotalus)的全部羽衣都具有美丽的玫瑰色彩,胸部有柠檬颜色的斑点;但这等色彩,正如斯克莱特先生说的,“保持不久,一般约在六周或两个月后就消失了”。某些鷽类的羽毛边缘于春季脱落,这时其色彩变得更鲜明,而燕雀类其他的种则不经历这样的变化。例如,美国的暗色燕雀(Fringilla tristis)同许多其他美国的物种一样,只在冬季过后才呈现其鲜明的色彩,而确切表现了这种鸟的习性的英国金翅雀,以及在构造上同这种鸟还要接近的英国黄雀(siskin),则不经历这样的年度变化。但是,亲缘接近的物种在羽衣上有这类差异并不为奇,因为属于同一科的普通红雀,其艳红色的前额和胸部只在英国的夏季才呈现出来,而在马德拉,这等色彩则可保持全年。 (415)
雄鸟夸耀其羽衣
一切种类的装饰物,不论是永久获得的或暂时获得的,均为雄鸟孜孜不倦地加以夸示,显然这是为了刺激、吸引或魅惑雌鸟。但是,当没有雌鸟在场时,雄鸟有时也夸示其装饰物,如松鸡类在其巴尔兹舞场偶尔发生的情况,又如孔雀也有这种情况;然而,孔雀显然渴望得到某种观众,正如我常常看到的,它在家禽甚至在猪的面前也显示其华丽的羽衣。 (416) 所有曾经密切注意过鸟类习性的博物学家们,不论所注意的是自然状况下的或是圈养状况下的,都一致承认雄鸟乐于夸耀其美。奥杜邦屡次谈到雄鸟用各种办法尽力献媚雌鸟。古尔德先生在描述了一只雄蜂鸟的某些特性之后说道,他不怀疑它在雌鸟面前能够最有效地表现其特性。杰尔登博士 (417) 坚持认为雄鸟的美丽羽衣乃是用于“魅惑和吸引雌鸟的”。伦敦动物园的巴特利特先生用最有力的字眼向我表达了他自己对这种效果的同样看法。

图 50 美洲巨冠黄鸟(Rupicola crocea)的雄者
(引自T.W.伍德)
出现于印度森林里的下述情景必定是很壮观的:“忽然出现了20或30只孔雀,在感到喜悦的雌者之前,雄者夸示其华丽的尾羽,意气扬扬,昂首阔步。”野生雄火鸡竖起其灿烂的羽毛,展开其具有精美轮纹的尾羽和具有条纹的翼羽,再加上其艳红色和蓝色的垂肉,一起形成了一副美丽的模样,虽然这种模样在我们的眼里是奇形怪状。有关各种松鸡的相似事实已经列举过了。现在让我们转来谈谈鸟类的另一个“目”。雄性美洲巨冠黄鸟(Rupicola crocea,图50)是世上最美丽的鸟类之一,它具有华丽的橙色,有些羽毛奇妙地缩短而成羽状。其雌鸟为褐绿色并蒙上红晕,而且其羽冠比雄鸟的小得多。朔姆布尔克(R.Schomburgk)爵士描述过它们的求爱,他发现它们的一个聚会场所,那里有10只雄鸟和两只雌鸟。场所的直径为4~5英尺,其中没有一片叶子,而且平滑得就像用人手整理过的一样。一只雄鸟“在跳跃着,显然使若干其他鸟感到高兴。它随即展翅昂首,或展开羽尾如扇;接着以一种跳跃的步法大摇大摆地走来走去,直到累了为止,这时它急促地发出某种声调,跟着让位给其他雄鸟。于是,有三只雄鸟相继占领了这块空地,然后自我欣赏地退下去休息”。印第安人为了猎取其鸟皮,守候在它们的一个聚会场所,等这些鸟都热衷于跳舞之际,就能用毒箭一只又一只地射杀四五只。 (418) 至于极乐鸟类,有一打或一打以上羽毛丰满的雄鸟集合于一株树上举行当地土人所谓的舞会:它们在这里飞来飞去,高举双翼,竖起其优美的羽毛并使之颤动,就像华莱士先生所说的,这整株树好像充满了飘动的羽毛。当它们这样进行时,竟变得如此凝神专注,以致一个好射手几乎可把整个舞会上的鸟全部射下来。当把这些鸟圈养在马来群岛时,据说它们很注意保持其羽毛的整洁;常常伸开其羽毛加以检查,并把每一个尘粒都清除掉。有一位养了几对这种鸟的观察家并不怀疑雄鸟的夸示其美是为了取悦于雌鸟。 (419)

图 51 孔雀雉(Polyplectron chinquis),雄鸟
(引自T.W.伍德)
金雉(gold pheasant)和云实树雉(amherst pheasant)在求偶期间不仅展开和抬高其华丽的颈羽,而且如我亲自看到的,还把它扭曲并使之斜对着雌鸟,不论它站在哪一边都是如此,这显然是为了把一个大的表面显示在雌鸟之前。 (420) 同样地,它们还把其美丽的尾羽和尾覆羽稍微转向雌鸟那一边。巴特利特先生见过一只在求偶活动中的雄孔雀雉(Polyplectron,图51),并给我看了按当时姿态制作的一个标本。这种鸟的尾羽和翼羽都饰以美丽的眼斑,就像孔雀尾羽上的眼斑那样。当雄孔雀夸示自己时,便展开并竖起其尾羽,使其同躯体相横切,这是因为它站在雌鸟之前,需要同时显示其鲜蓝色的喉部和胸部。但团花雉胸部的色彩是暗淡的,而且眼斑并不限于尾羽才有。因而团花雉不是站在雌鸟之前;但它略为斜斜地抬高和展开其尾羽,把对着雌鸟那边的一翅展开并低垂下来,而把另一边的一翅高举起来。依此姿势,遍布全身的眼斑就可以同时暴露在对此赞赏的雌鸟眼前,从而构成一幅宽阔灿烂的景象。雌鸟不论转到哪个方向,雄鸟张开的双翼羽和斜举的尾羽也会转向雌鸟那一边。雄红胸角雉的行为也几乎一样,虽然它不展开翼羽,却把向着雌鸟那一方的躯体上的羽毛竖起,它们在其他时候隐蔽不显,一竖起来则几乎一切具有美丽斑点的羽毛都同时显示出来了。
锦雉提供了一个更为显著得多的事例。其极为发达的次级翼羽只限于雄鸟才有,每张翼羽饰以一行20~30个直径1英寸以上的眼斑。这些羽毛还具有雅致的斜条纹和成行的暗色斑点,犹如把虎皮和豹皮上的纹彩结合起来一样。这些美丽的装饰物平时隐而不露,等到雄鸟在雌鸟面前夸示自己时才显露出来。这时它竖起尾羽并把翼羽展开,成为一把几乎笔直的大圆扇或一张大盾牌,置于躯体的前方。它的颈和头均保持在一边,因而被大圆扇所遮住(图52);但这种雄鸟为了望见正向其夸示自己的雌鸟,有时把头从两支翼羽之间伸出去(像巴特利特先生所见过的),于是表现了一副奇形怪状。这必定是这种鸟在自然状况下的一种常见的习性,因为巴特利特先生和他的儿子在检查从东方送来的一些完整鸟皮时,发现在那两支翼羽间有一处磨损得很厉害,好像鸟头曾经常常在该处伸进伸出。伍德先生认为这种雄鸟也能越过圆扇的边缘从一边窥视雌鸟。
翼羽上的眼斑是一种不可思议的装饰物,正如阿盖尔公爵所说的, (421) 其浓淡如此合宜,以致它们形状凸出得就像松松置于球穴中的一只球。我在大英博物馆看到过这种标本,它两翼展开,向下垂放,但它使我大失所望,因为这等眼斑显得扁平,甚至凹陷。不过古尔德先生很快就把情况给我讲清楚了,因为,雄鸟当在自然状况下夸示其美的位置上竖起羽毛时,光线是从上方照射下来的。因而各个眼斑立刻就会显得类似那种所谓球与穴的装饰物了。这等羽毛曾给几位艺术家看过,他们对其色彩的浓淡适宜无不表示赞赏。似乎应该这样提问:这种色彩浓淡适宜的艺术性装饰物会不会依据性选择的手段而形成的呢?不过把这个问题推延到我们下一章讨论级进的原则时再予以回答将会方便些。
以上所说的是关于次级翼羽的情况,至于初级翼羽,在大多数鹑鸡类中其色彩都是一致的,但在锦雉中则同样是不可思议的。它们具有柔和的褐色以及大量的暗黑斑点,每个斑点都是由2~3个小黑点组成的,并围以暗黑环带。同暗蓝色羽干相平行的有一处空白,它的轮廓是由一支位于真羽之内的次级羽毛形成的,这正是其主要的装饰所在。其里层部分着有较淡的栗色并有微小的白点密布其上。我曾把这等羽毛给若干人士看过,其中有许多人对它的赞赏甚至超过了对球与穴的那些装饰物的赞赏,他们还声称与其说它是自然生成的,莫如说它更像是一种艺术作品。在通常所有情况下这些羽毛完全隐而不现,只有当它们和长长的次级羽毛一起全部展开而形成一把大扇或一面盾牌的时候才充分显示出来。

图 52 雄锦雉侧面图,正在雌鸟之前夸耀自己。
伍德先生根据在自然界的观察绘制而成
雄锦雉的情况是显著有趣的,因为它提供了很好的证据来说明最优雅的美可能是作为一种性的魅诱而无其他目的。我们必须断定情况确系如此,因为在雄鸟进行求偶之前,其次级翼羽和初级翼羽完全不显露,而且球与穴那种装饰也不完全充分显露。锦雉的色彩并不鲜艳,因此它求爱的成功似乎决定于其巨大的羽型以及精心制作的最优雅样式。许多人将会宣称,一只雌鸟能够欣赏浓淡合宜的色彩和雅致的样式乃是极不可信的。这诚然是一个不可思议的事实:雌鸟大概具有近乎人类水平的鉴赏力。凡是认为能够可靠地估计低等动物的鉴别力和欣赏力的人,可能都会否定锦雉能够欣赏这种优雅的美;但是,这时他将被迫承认,雄鸟在求偶活动中所表现的异常姿势,借此以充分显示其非常美丽的羽衣,乃是无目的的。这是我永远不会承认的一个结论。
虽然那么多的雉类以及亲缘接近的鹑鸡类都不厌其烦地在雌鸟面前夸示其羽衣,但是,正如巴特利特先生告诉我的,值得注意的却是,颜色暗淡的蓝马鸡(Crossoptilon auritum)和欢乐雉(Phasianus wallichii)的情况却非如此,所以说这等鸟类似乎意识到了它们没有多少可以夸示的美。巴特利特先生从未见过这两个物种的雄鸟相互争斗,虽然他观察欢乐雉的机会不如观察角雉的机会那样好。詹纳·韦尔先生也发现一切雄鸟如果具有色彩浓艳或特征强烈显著的羽衣,就比同一类群中那些色彩暗淡的物种更好争吵。例如,金翅雀就远比红雀好斗,乌鸫也比画眉好斗。同样地,羽衣发生季节性变化的那些鸟类也在它们装饰最华丽的期间变得更加好斗得多。某些颜色暗淡的鸟类无疑也会互相进行殊死的战斗,但是,当性选择发挥其高度影响并使任何物种的雄鸟具有鲜明色彩时,似乎也往往使这些雄鸟具有一种好斗的强烈倾向。我们在讨论哺乳动物时将会遇到差不多相似的事例。另一方面,同一物种的雄鸟既获得鸣唱的能力又获得灿烂的色彩却是罕有的;但这两方面所获得的利益也许是一样的,这就是魅诱雌鸟的成功。尽管如此,还必须承认,若干色彩灿烂的鸟类,其雄者的羽毛为了发出器乐鸣叫的缘故也曾经发生过特别的变异,虽然这种美,至少按我们的鉴赏标准来说,是无法同许多鸣禽类所发出的声乐鸣叫之美相比拟的。
我们现在转来谈谈没有高度装饰的雄鸟,它们在求偶时仍将其可能有的无论什么吸引力都显示一番。这等事例在某些方面比上述那些事例更为奇妙,但很少为人所注意。感谢韦尔先生为我提供了下述事实,他长期圈养过许多种类的鸟,包括所有英国的燕雀科(Fringillidae)和鹀科(Emberizidae)的鸟。这些事实就是从他好心寄给我的大量有价值的记录中选出来的。红腹灰雀为了求爱而走近雌鸟之前时,噗地一下鼓起其胸部,因此其艳红色的羽毛立刻得见,这比在任何位置上都显示得更清楚。与此同时,它把黑尾低垂,从这一边扭转到那一边,作出一副可笑的样子。欧洲苍头燕雀也站在雌鸟之前,这样来显示其红色胸部和“蓝钟”——养鸟行家以此名其头;同时双翼微张,使其肩部的纯白带斑显露无遗。普通红雀鼓起其玫瑰色胸部,微张其褐色的双翅和尾部,以使这等羽毛的白色边缘最充分地显露出来。然而,要断言双翅的展开仅仅是为了显示之故,必须要谨慎,因为某些鸟类的翅膀并不漂亮,但也会这样做。家养雄鸡的情况就是这样,但它所展开的那个翅膀总是对着雌鸡的,同时以翅擦地而过。雄金翅雀的行为不同于所有其他鷽类:它的双翅是美丽的,肩部黑色,翼羽上散布着白色斑点,其尖端呈黑色,边缘为金黄色。当它向雌鸟求偶时,其躯体摆来摆去,并迅速将其略微张开的双翅先转到一边,然后再转到另一边,于是产生了金光闪闪的效果。韦尔先生告诉我说,没有其他英国鷽类在求偶期间这样转来转去的,即使亲缘相近的雄金雀也是一样,这大概因为其美丽并不因此而有所增添。
大多数英国的鹀类(buntings)都是颜色平淡的鸟;但雄苇鹀(Emberiza schaeniculus)的头部羽毛到春天就脱去其污色的毛尖,而获得一种优美的黑色;这等羽毛在求偶活动中就会竖起来。韦尔先生曾经养过澳大利亚产的环喉雀(Amadina)的两个物种:A.castanotis是一种体型很小而色彩朴素的燕雀类,具有一条黑尾,白臀,以及漆黑的尾上覆羽(upper tail-coverts),后者每根羽毛上都有三个显著的椭圆形白色大斑点。 (422) 这个物种当向雌鸟求偶时,便把这等杂色的尾覆羽微微张开并以很奇特的方式进行摇晃。雄拉塔环喉雀(Amadina lathami)的行为则很不相同,它们在雌鸟之前展示其具有鲜艳斑点的胸部、猩红色的臀部以及猩红色的尾上覆羽。根据杰尔登博士的材料,我在这里还可以补充一点:印度红鹎(Pycnonotus haemorrhous)具有鲜红色的尾下覆羽,可以想象,这等尾覆羽永远不会充分展示的;但这种鸟“一旦激动时,也往往会把这等尾覆羽横向地张开,因而即使从上面也能看到它们”。 (423) 某些其他鸟类的鲜红色尾下覆羽,即使不进行夸示,也能看见,大型啄木鸟(Picus major)的情况就是如此。普通鸽子的胸部具有彩虹色的羽毛,大家一定都看到过这种雄鸽当向雌鸽求偶时,便把胸部鼓起,这样就会使胸部羽毛显示到充分的程度。澳大利亚有一种具有漂亮的青铜色翅膀的鸽子,叫做冠毛野鸽(Ocyphaps lophotes),其行为,如韦尔先生向我描述的,则迥然不同:当雄鸽站在雌鸽之前时,低垂其头几乎达到地面,张开并高举其尾,并半张其双翅。然后它交替地使其躯体缓慢起落,因而那些具有彩虹色金属光泽的羽毛立刻尽收眼底,并在阳光下闪闪发亮。
现在已经举出了足够的事实来阐明雄鸟多么细心地显示其种种魅力,而且它们是极其熟练地进行这种显示。当它们用嘴来啄理其羽毛时,它们经常有机会进行自我欣赏并学习如何最好地展示其美。但是,由于同一物种的所有雄鸟都以完全一样的方式来显示自己,因此,这种行为最初也许是有意的,以后就变成为本能的了。果真如此,我们就不应责备鸟类有意识地进行虚夸,然而当我们见到一只把尾羽展开并使其抖动着的孔雀大摇大摆地走来走去时,它似乎就是骄傲与虚夸的唯一典型。
雄者的各种装饰物对它们肯定具有最高的重要性,因为在某些场合中,它们获得这等装饰物是以面临飞行或奔跑的巨大阻力为代价的。非洲夜鹰(Cosmetornis)在交配季节有一支初级翼羽发展成很长的飘带,因而大大减慢了其飞行速度,虽然它在其他时候是以飞得快而著称的。雄锦雉的次级翼羽“非常笨重”,据说这“几乎完全剥夺了它的飞翔能力”。雄极乐鸟的美丽羽毛使它们在大风之际处于困境。南非的雄黑羽长尾鸟(Vidua)的极长的尾羽使“它们飞翔吃力”,一旦这等尾羽脱落后,它们就飞得同雌鸟一样好了。由于鸟类总是在食物丰富时进行繁殖,因此雄鸟在寻找食物时大概不会由于它们行动的阻力而遇到很多不便;但几乎无可怀疑的是,它们一定会更容易地被猛禽类所击落。我们也无法怀疑孔雀的长尾以及锦雉的长尾和翼羽一定会使它们更容易被任何四处觅食的山猫所捕获,否则就不会如此。甚至许多雄鸟的鲜明色彩也必定会使它们易于被各种敌害所发现。因此,正如古尔德先生说过的,这种鸟类大概一般都具有一种胆怯的性情,好像意识到了它们的美就是危险的根源,它们比颜色暗淡、性情较为温顺的雌鸟或者比尚未装饰的幼小雄鸟更难被发现或者更难接近。 (424)
一个更为奇异的事实是,某些鸟类的雄者具有进行战斗的特殊武器,它们在自然状况下如此好斗以致常常互相残杀致死,这等鸟类由于具有某些装饰而身受其苦。斗鸡者修剪斗鸡的颈部纤毛,割去其肉冠和垂肉,据说这时它们才取得了斗鸡的称号。一只尚未取得斗鸡称号的公鸡,正如特格梅尔先生所主张的,“是处于一种可怕的劣势,它的鸡冠和垂肉容易被其对手啄住,雄斗鸡总是向它所啄住的地方进行打击,一旦它啄住其对手时,就把对手完全控制在自己的力量之下了。即使假定这只雄斗鸡没有被杀死,未经修剪者所流的血也远比修剪者多得多”。 (425) 幼小雄火鸡在相斗时总是啄住对方的垂肉,我相信成年火鸡也是按照同样的方式彼此争斗。也许有人会反对说,肉冠和垂肉并非装饰性的,因而在这方面不会对它们有什么用处;但是,即使以我们的眼光来看,光泽闪闪的黑色雄西班牙鸡之美也会被其白脸和鲜红色肉冠大大加强;雄红胸角雉在求偶时便鼓起华丽的蓝色垂肉,凡是见过这种情景的人将会毫不迟疑地承认它要达到的目的正是在于美观。根据上述事实,我们清楚地看到了雄鸟的羽饰以及其他装饰物对它们一定具有最高的重要性;我们进一步看到这种美甚至有时比相斗的胜利还更重要。
第十四章 鸟类的第二性征(续)
雌鸟实行选择——求偶历时甚久——丧偶的鸟类——心理属性和审美力——雌鸟对特殊雄鸟所表现的爱好和憎恶——鸟类的变异性——变异有时是突发的——变异的法则——眼斑的形成——性状的级进——孔雀、锦雉和蝶鸟诸例
当雌鸟和雄鸟在美丽方面或在鸣唱能力方面或在演奏我所谓的器乐能力方面有所差异时,几乎总是雄鸟胜过雌鸟。这些属性,如我们刚刚见过的那样,对雄鸟显然具有高度的重要性。如果只在一年的一部分时间表现有这等属性,这总是在生殖季节以前。只有雄鸟才尽力显示其种种的魅力,并常常在地面或空中于雌鸟之前进行奇怪的滑稽表演。每只雄鸟都要把其竞争对手赶走,要是办得到的话,就要把它们杀死。因此,我们可以断言,雄鸟的目的就在于诱使雌鸟与之交配,为了达到这个目的,它试图用各种方法去刺激她,媚惑她;这就是所有仔细研究过活鸟习性的人们所持的见解。但是,还留下一个同性选择有非常重要关系的问题尚待解决,即,同一物种的每只雄鸟是否都同等地刺激和吸引雌鸟呢?或者,雌鸟是否实行选择并且偏爱某些雄鸟呢?后面这个问题,可用许多直接的和间接的证据予以肯定的回答。究竟是那些属性来决定雌鸟的选择,殊难断言;但我们在这里也有某些直接的和间接的证据可以证明雄鸟的外在魅力在很大程度上是决定雌鸟选择的因素;虽然雄鸟的精力、勇敢以及其他心理属性无疑也起了作用。我们将从间接的证据开始。
求偶历时甚久
某些鸟类的雌雄二者日复一日地在一个约定的场所相会,历时颇久,这大概部分地决定于鸟类求偶是一件费时的事情,而且部分地决定于交配行为是反复进行的。例如,在德国和斯堪的纳维亚,黑松鸡所举行的巴尔兹(balz)或勒克斯(leks)舞会从三月中开始,经过四月份整整一个月,一直到五月才结束。在勒克斯舞会上聚会的鸟竟达40或50只之多,甚至还要多;而且以后连续数年往往都在这同一场所聚会。雷鸟的勒克斯舞会从三月底开始,到五月中、甚至到五月底才结束。在北美,尖尾松鸡(Tetrao phasianellus)的“鹧鸪舞”“要持续一个月或者还要长些”。无论北美或西伯利亚东部的其他种类的松鸡差不多都遵循相同的习性。 (426) 捕鸟人根据草被踏光的情况可以发现流苏鹬相聚的小丘,这说明此处是它们长期出没的场所。圭亚那的印第安人十分熟悉岩鸽的清洁的活动场所,他们可以期望在那里找到漂亮的雄岩鸽;新几内亚土人知道极乐鸟聚会于其上的那些树,10~20只羽饰丰满的这种雄鸟常集合于此。在后面这个例子里,没有明确提到在这些树上是否有雌鸟来会,但是,捕鸟人如果没有被特别询及,大概不会谈到有无雌鸟在场,因为她们的鸟皮是毫无价值的。一种非洲织巢鸟(Ploceus)在繁殖季节集合起来举行小型舞会并表演其优美的舞蹈动作达数小时之久。大量的独居丘鹬(Scolopax major)每于黄昏时节在沼泽中相聚;以后连续几年它们为了同样目的仍然常常出没于同一场所;在那里可以看到它们“像许多大老鼠似的”跑来跑去,高耸其羽毛,拍打其双翼,并发出最奇异的叫声。 (427)
在上述鸟类中有些据认为是一雄多雌者,如黑松鸡、雷鸟、雉松鸡(pheasantgrouse)、流苏鹬、独居鹬即是,大概还有其他鸟类也是这样。对这等鸟类来说,可以认为其雄鸟之强者大概只要把弱者赶走后,马上就可以占有尽可能多的鸟;但如果雄鸟必须去刺激或取悦于雌鸟的话,那么我们就能理解为什么需要那样长的时间进行求偶,而且需要在同一个地点集合那么多雌雄二者。某些严格单配的物种也同样举行结婚集会;斯堪的纳维亚有一种松鸡似乎就是如此,其勒克斯舞会从三月中旬开始一直到五月中旬才结束。澳大利亚琴鸟(Menura superba)做成的“小圆丘”,以及阿氏琴鸟(M.alberti)给自己扒成的浅穴,都被当地土人称为“克罗伯瑞舞场”,据认为那里就是雌雄二者相聚的场所。澳大利亚琴鸟的集会有时规模很大。最近一位旅游者发表的一文章说, (428) 他曾到过一处地方,其下为一茂密丛林所覆盖的山谷,他听到从那里发出了“一阵使他十分震惊的喧哗”;他慢慢地走近该处,惊奇地看到了约有150只华丽的琴鸟“列阵相争,并以无法形容的狂怒进行战斗”。造亭鸟的亭子乃其雌雄二者在生殖季节常去之处;“雄鸟在此相遇并为了取悦于雌鸟而互相争斗,雌鸟则集合于该处向雄鸟卖弄风情”。该属有两个物种,它们许多年都常常在同一个亭子相聚。 (429)
普通喜鹊(Corvus pica,Linn.),如达尔文·福克斯牧师告诉我的,常从德勒密尔(Delamere)森林各处集合起来,以庆祝其“盛大的喜鹊婚礼”。数年前这种鸟的数量特别多,因而一个猎场看守人在一个早晨就打死了19只雄鸟,另一人一枪就打死了栖息在一起的7只鸟。再者,它们有早春集合于特殊地点的习性,在那里可以看到成群的这种鸟叽叽啁啁乱叫,有时互相争斗,并在树的周围喧闹着飞来飞去。这种鸟显然把这全部情况看做是极其高度重要的事情之一。在集会后不久它们就各自分散了,于是福克斯先生和其他人士曾见到它们就在这一季节交配。凡是一个物种没有大量成员存在的任何地区,自然无法在那里举行盛大集会,因而同一物种在不同地方可能有不同的习性。例如,韦德伯恩(Wedderburn)先生向我说过一个例子:黑松鸡在苏格兰只举行一次例会,而在德国和斯堪的纳维亚这等集会是如此闻名,以致获得了专用名称。
丧偶的鸟类
根据现在提出的事实,我们可以得出结论说,属于大不相同的类群的鸟,其求偶往往是一件费时、微妙而麻烦的事情。甚至还有理由推测,最初看起来这好像是不可能的,即,栖息于同一地区、属于同一物种的某些雄鸟和雌鸟并非总是相互喜欢,因而不互相交配,已经发表过的许多记载表明,一对配偶中的雄鸟或雌鸟如果被射杀,很快就会有另一个来代替。在喜鹊比在任何其他鸟类更会经常看到这种情况,恐怕这是由于其外貌和鸟巢惹人注目之故。著名的詹纳说,在维尔特郡(Wiltshire),一天之内就射杀了一对喜鹊中的一只不下7次之多,“但全无用处,因为剩下的那只喜鹊很快又找到了另一只配偶”;而最后这一对照样养育幼鸟。新配偶一般要在隔天才会找到;但汤普孙先生举出过一个例子表明在同一天傍晚就换了一个配偶。即使在鸟卵孵化之后,若有老鸟之一被杀,也会找到一只配偶;卢伯克爵士的猎场看守人最近观察到的一个例子 (430) 表明,这种情形发生在两天之后。首先的和最明显推测将是,雄喜鹊的数量一定比雌喜鹊多得多;而且在上述场合中,以及在能够举出的许多其他场合中,被杀死的只是雄鸟。这种推测对某些事例显然是适用的,因为德勒密尔森林的猎场看守人向福克斯先生保证说,以前有大量的喜鹊和食腐肉的乌鸦在其鸟巢附近被相继打死,而且被打死的全是雄鸟;他们对这一事实的解释是,雄鸟在把食物带给孵卵的雌鸟时比较容易被打死。然而,麦克吉利夫雷根据一位优秀观察家的材料,举出一个事例表明,在同一个窝里相继被打死的3只喜鹊都是雌鸟;另外还有一个事例表明,有6只喜鹊连续被打死,当时它们都相继在抱同一窝的卵,从抱窝这一点来看,它们多数可能是雌鸟;但是,我听福克斯先生说过,雌鸟一旦被打死,雄鸟就要代之孵卵。
卢伯克爵士的猎场看守人曾反复地用枪射死了一对松鸦(Garrulus glandarius)中的一只,但不能详说其射击次数,射杀一只以后,总能发现另一只未亡者又再婚配了。福克斯先生,勃恩德(Bond)先生以及其他人士都曾用枪打死过一对食腐肉的小嘴乌鸦(Corvus corone)中的一只,但其巢很快又有一对鸦住上了。这些鸟类都是相当普通的,但游隼(Falco peregrinus)则是罕见的,然而汤普孙先生说道,在爱尔兰“如果其成熟的雄者或雌者在生殖季节中任何一个被打死了(这并非是不常有的事),另一只配偶在很短几天内就会被找到,因此,隼鹰尽管有了这类伤亡,肯定还会产出足够的幼鸟来补充。”詹纳·韦尔先生所知道的滩头堡(Beachy Head)的隼也是如此。同一位观察家告诉我说,三只红隼(Falco tinnunculus),全系雄者,在先后光顾同一鸟巢时都相继被打死了;其中两只都具成熟的羽衣,第三只则具前一年的羽衣。一位苏格兰的可信赖的猎场看守人向伯贝克(Birkbeck)先生保证说,即使是罕见的金雕(Aquila chrysa tos),倘若一对中有一只被打死了,很快就会找到顶替它的另一只。短耳鸮(Strix flammea)也是如此,“未亡者很容易找到一只配偶,虽然射杀不断进行”。
tos),倘若一对中有一只被打死了,很快就会找到顶替它的另一只。短耳鸮(Strix flammea)也是如此,“未亡者很容易找到一只配偶,虽然射杀不断进行”。
塞尔伯恩(Selborne)的怀特,即举出鸮的实例者,进一步说,他知道有一个人相信山鹑的交配会由于雄者的相斗而受到干扰,所以常常去射死它们;他虽然几次使同一只雌鸟丧偶,但她总是很快地又找到了一个新配偶。同一位博物学家又因麻雀夺去了家燕的巢,命令把前者射杀;但一对中所留下的“不论是雄者或雌者,立刻就会得到一只配偶,这样连续进行几次都是如此”。有关苍头燕雀、夜莺以及红尾鸲的情况,我还可以补充几个近似的例子。就红尾鸲(Phaenicura ruticilla)来说,一位作家感到非常惊奇的是,抱窝的雌鸟如何能够那么快地作出有效的表示,使雄鸟知其为寡者而来就之;之所以叫之感到惊奇还因为附近并不常见这个种鸟。詹纳·韦尔先生向我说过一个非常相似的事例;他在布莱克希思(Blackheath)从未见过野红腹灰雀,也没有听过它的鸣叫,然而当他养在笼子里的一只雄鸟死后,一般在几天之内就会有一只野生雄鸟飞来栖于丧偶的雌鸟附近,而雌鸟的叫声并不高。根据同一位观察家的材料,我还要再举另一个事实;有一对紫翅椋鸟(Sturnus vulgaris),其中之一在早晨被打死,到了中午一个新配偶就被找到了;这一只又被打死,但到晚上以前这一对又配齐了。因此那个忧伤的寡鸟或鳏鸟在同一天之内就三次得到了安慰。恩格尔哈特(Engleheart)先生也告诉我说,在布莱克希思有一处房屋,欧椋鸟在这所房屋一个空穴内筑巢,几年以来他常常把配偶的一只打死,但失去的那一只的位置总是立刻就会补上。在某一个季节里,他作了记录,发现从同一个巢打死了35只,其中有雌鸟、也有雄鸟,但二者的比例如何,他说不清楚:尽管如此,在经历了所有这样灾祸以后,一窝鸟还是生育出来了。 (431)
这些事实十分值得注意。怎么会有那么多的鸟随时可以立即顶替雄者或雌者任何一方所失去的一个配偶呢?我们在春季所看到的喜鹊、松鸦、小嘴乌鸦、山鹑和其他一些鸟类总是成双成对的,从未见过它们是独身的;乍一看,这种情况是极其复杂的。但是,同一性别的鸟,当然不会真正地相配,虽然如此,有时也成对或成小群地生活在一起,据知鸽子和鹧鸪的情况即是。有时鸟类还会三只一组地生活在一起,有人观察到椋鸟、小嘴乌鸦、鹦鹉和山鹑就是如此。关于山鹑,已知有两只雌者和一只雄者生活在一起以及两只雄者和一只雌者生活在一起。在所有这类场合中,这种结合大概容易破裂;因为三者之一随时都会同一只寡鸟或一只鳏鸟相配。偶尔会听到某些鸟类的雄者在过了特定的季节很久以后还纵声高唱其求爱的歌曲,这表明它们已经失去或从未得到过一只配偶。一对配偶中的一只如死于事故或疾病,就会使另一只成为自由而孤单的;有理由相信雌鸟在生殖季节特别容易夭折。此外,巢窝被毁的鸟,或不育的配偶,或发育迟缓的个体,大概都容易诱使其一离去,而且还会出于乐趣和义务去抚养虽非自己所生的后代。 (432) 这种偶然发生的事情大概可以说明大多数上述事例。 (433) 尽管如此,在同一地区内,正值生殖季节的高峰期间,成对之鸟损失其一,竟有如此众多的雄鸟和雌鸟随时准备补上,也还是一个奇怪的事实。为什么这等孤独诸鸟没有彼此立刻相配呢?难道我们没有某种理由来推测,由于鸟类的求偶看来在许多场合中都是一件费时而麻烦的事,所以会偶尔出现某些雄鸟和雌鸟在特定季节内没有能够成功地激起彼此的爱情,因而没有结为配偶吗?詹纳·韦尔先生就曾做过这样的推测。当我们看到了雌鸟会偶尔对特殊的雄鸟表示何等强烈的憎恶和偏爱之后,可知这种推测似乎就不那么不可能了。
鸟类的心理属性及其对美的鉴赏力
在我们进一步探讨雌鸟究竟选择魅力较强的雄鸟还是接受它们所可能碰到的头一只雄鸟这个问题之前,大致地考察一下鸟类的精神能力将是合宜的。它们的理智一般被认为是低等的,这种意见也许是正确的;但还可以举出导致相反结论的一些事实。 (434) 然而,低级的理解力同强烈的感情、敏锐的知觉以及对美的鉴赏力是并存的,我们从人类可以看到这种情形;而我们在这里所要讨论的正是后面这些属性。往往听说,鹦鹉如此深深地相互依恋,以致有一只死了,另一只会长期悲伤憔悴;但詹纳·韦尔先生认为大多数鸟类感情的强度是被夸大了。尽管如此,当配偶的一只在自然状况下被打死之后,还会听到未亡者在此后几天要发出一种痛苦的鸣叫;圣约翰先生(Mr.St.John)举出了各种事实来证明已成配偶的鸟类有相互依恋之情。 (435) 贝内特先生述说 (436) ,中国所产的美丽鸳鸯,如果其雄者被偷走之后,剩下的雌鸳鸯尽管有另一只雄鸳鸯在雌者面前显示其全部的魅力,殷勤地向其求爱,仍然郁郁不乐。三周以后那只被偷走的雄鸳鸯又出现了,于是这一对鸳鸯立即以极大的喜悦彼此认出来了。另一方面,如我们已经见到的,欧椋鸟同一天内三次失偶,三次换配新偶,而感到欣慰。鸽子对地点具有非常卓越的记忆力,据知它们离开原地九个月后还能飞回,可是,如我听哈里逊·韦尔先生所说的,如果有一对自然终生匹配的鸽子在冬天被分开少数几个星期,又分别同其他鸽子相配,那么,此后把原来那一对鸽子再放到一起时,彼此还能相认者,即使有的话,也是罕见的。
鸟类有时表现有仁慈的感情;它们喂养甚至属于不同物种的幼鸟,不过也许应认为这是一种错误的本能。它们还喂养双目失明的同种的成年鸟,本书前一部分对此已有所论及。布克斯顿先生做过一项奇妙的记载,表明一只鹦鹉照管一只异种的冻伤了而残废的鸟,将其羽毛弄干净,保护它免受其他在花园周围自由飞翔的鹦鹉的攻击。更为奇妙的是,这些鸟对于同伙的欢乐明显表示了某种同情。当一对白鹦(cockatoos)在一株合欢树上做巢时,“同种的其他鹦鹉对这件事表示了高度的兴趣,其态至为滑稽可笑”。这等鹦鹉还表现有无限的好奇心,而且明显地具有“财产和所有权的观念”。 (437) 它们有良好的记忆力,因为在动物园里经过几个月后它们还能明确地认出以前的主人。
鸟类具有敏锐的观察能力。已配的每一只鸟当然都认识其伴侣。奥杜邦说,“模拟画眉”(Mimus polyglottus)有一定数量一年到头都留在路易斯安那(Louisiana),而其余的那些则向东部各州迁徙;这些鸟一回来马上就会被其南方同胞认出,而且总要遭到它们的攻击。笼养的鸟能辨认不同的人,它们对某些个人并无明显原因的强烈而持久的憎恶或喜爱证明了这一点。我听说过不少关于松鸦、山鹑、金丝雀,尤其是灰雀在这方面的事例。赫西(Hussey)先生描述过一只驯养的鹧鸪如何奇异地认出了每个人;它的爱和憎都很强烈。——这只鸟似乎“喜欢华丽的颜色,谁穿上新上衣或戴上新帽子没有不引起它的注意的”。 (438) 休伊特先生描述过某些鸭的习性(乃野鸭的最近后代),它们一见陌生的狗或猫来到,就急速纵身入水,竭力逃避;但它们同休伊特先生的狗和猫如此熟识,甚至卧于其旁晒太阳。它们见到陌生就避去,喂养它们的妇女如果在衣服方面有任何重大改变,它们也会避开。奥杜邦说,他驯养过一只野火鸡,它一见到任何陌生的狗总是跑掉;它曾逃入森林,几天后奥杜邦以为他见到了一只野火鸡,就叫他的狗去追它,当狗追到时却不攻击这只火鸡,原来它们彼此早就是老相识了。 (439)
詹纳·韦尔先生相信鸟类特别注意其他鸟类的色彩,这有时是出于嫉妒,有时表示彼此是亲属。例如,他把一只具有黑色头饰的苇鹀放进他的鸟舍,除一只红腹灰雀外,没有引起任何鸟对这只新客的注意,而这只红腹灰雀的头同样也是黑色的。这只红腹灰雀是很安静的,以前从未和任何同伴争吵过,包括另一只头部尚未变黑的苇鹀在内:但是,这只黑头苇鹀受到的虐待如此之凶,以致不得不把它移走。蓝顶雀(Spiza cyanea)在生殖季节呈鲜蓝色;虽素性温和,但也攻击仅头部呈蓝色的蓝顶雀(S.ciris),甚至把后面这不幸者的头皮完全剥掉。韦尔先生也不得不把一只歌鸲从他的鸟舍移走,因为它在其中大肆攻击所有在羽衣上带有红色的鸟类,对其他鸟则不攻击,事实上它弄死了一只红交嘴雀(crossbill),而且几乎把一只金丝雀弄死。另一方面,他也观察到,当把某些鸟类首次放进鸟舍时,它们就飞向那些色彩最像它们的物种,并落于其旁。
由于雄鸟在雌鸟之前很细心地显示其漂亮的羽衣和其他装饰物,所以这等雌鸟欣赏那些求婚者的美,显然是可能的。然而要获得有关雌鸟审美力的直接证据却是困难的。当鸟类注视其镜中之影时(关于这方面的例子已有许多记载),我们无法肯定这不是出自它对一个假想竞争对手的嫉妒,虽然有些观察家的结论与此相反。在其他场合中,要把单纯的好奇和鉴赏区别开来也是困难的。正如利尔福(Lilford)爵士所说的, (440) 吸引流苏鹬向任何明亮目标飞去的恐怕就是好奇心,因此在爱奥尼亚群岛(Ionian Islands),“它不顾反复射击,急向一块颜色明亮的手绢飞下”。用一面小镜子在太阳底下晃动使其闪闪发光,这样就可以把普通云雀从天空引至地面而大批捕获它们。喜鹊、乌鸦和其他某些鸟类偷藏诸如银器和珠宝等某些明亮物体,究竟是出于鉴赏还是出于好奇呢?
吉尔德先生说,某些蜂鸟以“最大的兴趣”来装饰其鸟巢外部,“它们本能地在它上面贴上美丽平坦的地衣块,大者置于中央,小者放在和树枝相连的地方。不时把一根美丽的羽毛缠结在或黏着于鸟巢外面,把羽梗总是放在适当的位置,以使羽毛突出于表面之外”。然而,关于审美力的最好证据,还是前面提到的澳洲造亭鸟三个属所提供的。它们的亭子(见p257,图46)是雌雄二者相聚和进行奇异滑稽表演的场所,其构造各不相同,但同我们的讨论最有关联的乃是几个物种以不同的方式去装饰它们的亭子。萨丁造亭鸟收集色彩华丽的物品,诸如长尾鹦鹉的蓝色尾羽、漂白的骨头和贝壳,把它们插于树枝之间或摆在门口。吉尔德先生在一个亭子里发现了一柄工艺灵巧的石斧和一束蓝色棉花,显然这是从当地土人的一个野营里取来的。这些物体不断地被重新摆设,这些鸟在嬉戏时还把它们带来带去。斑点造亭鸟(spotted bowerbird)的亭子“系用高高的草造成,诸草排列整齐美观,草尖几乎相碰,而且装饰物极其丰富”。圆石子被用来把草梗固定于适当的位置,并用它们铺成一些通往亭子的曲径。石子和贝壳常常是从远方运来的。大王造亭鸟如拉姆齐(Ramsay)先生所描述的,用五六种漂白的陆贝壳以及“蓝的、红的和黑的各种颜色浆果装饰其矮亭,这些浆果的外貌在新鲜时非常漂亮。”此外,还有新拣回来的几片叶子和淡红色的嫩枝用做装饰,整个情况表明“它有一种明确的审美力”,古尔德先生也许说得好:“这等高度装饰起来的聚会大厦必被认为是迄今所发现的鸟类建筑的最奇异事例”;而几个物种的这种审美力,如我们所见到的,肯定有所不同。 (441)
雌鸟对特殊雄鸟的偏爱
在对鸟类的鉴别力和审美力预先作了以上这些记述后,我将把我所知道的有关雌鸟偏爱特殊雄鸟的全部事例列举出来。有一点是肯定的,即鸟类的不同物种在自然状况下会偶然交配并产生杂种。可举出这方面的许多事例:麦克吉利夫雷述说,一只雄乌鸫和一只雌画眉“彼此多么相爱”,并产生了后代。 (442) 关于松鸡和雉之间的杂种,几年前在英国曾记载过18个事例; (443) 但是,大多数这等例子,根据独身雄鸟找不到本种的雌鸟与之相配这一情况,或者可以得到说明。关于其他鸟类,如詹纳·韦尔先生有理由相信的那样,其杂种有时是近巢诸鸟偶尔互相杂交的结果。但这种说法对驯养或家养的异种鸟类的许多见于记载的事例不能适用,它们虽和本种的同类生活在一起,却被异种强烈地吸引住了。例如,沃特顿说 (444) ,有一大群加拿大的白颈雁,共23只,其中一只雌雁和一只独居的伯尼克尔雄雁(Bernicle gander)交配了,尽管它们的外观和大小是那样不同,可是还产生了杂种后代。一只雄赤颈凫(Mareca penelope)虽和同种的雌者生活在一起,据知却和一只针尾鸭(Querquedula acuta)交配了。劳埃德描述过一只雄麻鸭(Tadorna vulpanser)和一只普通母鸭之间的明显相恋。还可进一步举出许多例子;狄克逊牧师说,“凡是把许多异种的鹅养在一起的人都很了解它们彼此之间常常极相依恋,但其原因不明,它们十分愿意和一个显然跟自己最不相同的族(物种)的诸个体交配,并养育其后代,其情况正如和本种交配一样。”
福克斯牧师告诉我说,他同时饲养着一对鸿雁(Anser cygnoides)和一只雄的、三只雌的欧洲普通雁。开始时这两种鹅的界限十分分明,后来一只雄鸿雁竟引诱了一只雌普通雁与之共同生活。尤有甚者,从雌普通雁下的蛋孵出来的小雁只有4只是纯种的,另外18只都证明是杂种;因此这只雄鸿雁的魅力似乎在雄普通雁之上。我只再举一个例子;休伊特先生说,有一只圈养的雌野鸭,“同雄野鸭交配繁育了几年之后,因我把一只雄针尾鸭放入水中,她立刻就把雄野鸭甩掉了。这是一个一见钟情的事例,因为她在新来者的周围游来游去,爱抚备至,尽管雄尖尾鸭对此感到惊奇,并厌恶她主动表示的热情。从此以后,她就把原来的配偶忘掉了。冬季过去之后,到了翌年春天,这时雄针尾鸭对雌野鸭的献媚似乎变得回心转意了,因为它们同巢而居并产生了七八只小鸭”。
在若干这等场合中,除了单纯的新奇之外,还会有什么魅力呢,对此我们甚至无法进行猜测。然而色彩有时会起作用;因为按照贝希斯坦的材料,要使黄雀(Fringilla spinus)和金丝雀产生杂种,最好的办法是选择同样色彩的这两种鸟,把它们放在一起。詹纳·韦尔先生把一只雌金丝雀放进他的鸟舍,那里原来已有雄朱顶雀、雄金燕雀、雄黄雀、雄金翅雀、雄欧洲苍头燕雀以及其他种类的雄鸟,其目的是为了看看她选择何者;毫无疑问,当天她就选定了金翅雀,与之交配并产生了杂种后代。
雌鸟选中同种的某一雄鸟并与之交配的事实,似不及我们刚才看到的异种间所发生的这种情况更容易吸引我们的注意。前一情况最适于在家养的或圈养的鸟类中进行观察;但这些鸟类由于高水平的饲养而吃得过饱,它们的本能有时受到了极度损害。关于后面那种情况,我可举出有关鸽子、尤其是鸡的充分证据,但无法在此述及。上述某些杂种组合也许可用受损害的本能加以说明;但在许多这种场合中,那些鸟类是允许自由地生活于大水塘中的,所以没有理由设想它们会由于高水平的饲养而受到了不自然的刺激。
关于自然状况下的鸟类,每个人最初和最明显的设想是,雌鸟在繁殖季节接受她可能遇到的第一个雄鸟;但是,由于雌鸟几乎总是被许多雄鸟所追求,所以她至少有实行选择的机会。奥杜邦——我们必须记住他曾长期在美国的森林中徘徊,并对鸟类进行观察——并不怀疑雌鸟审慎地选择配偶;例如,当他谈到一只啄木鸟时,说道:这种雌鸟有六只华丽的追求者,它们不断作出奇异的滑稽表演,“直到雌鸟对某只雄鸟表示了明显的偏爱而后已”。红翼椋鸟(Agelaeus phaeniceus)的雌者同样被若干雄者所追求,“等到它们都变得疲乏之后,雌者才落下来接受它们的求爱并迅速作出选择”。他还描述过几只雄夜鹰如何屡屡以惊人的速度从空中急速下降,而后突然旋转,这样便发出一种独特的声响;“但雌鸟一经作出选择,其他雄鸟就全被赶走了”。美国有一种秃鹫(Cathartes aura),其雌雄二者常有8只、10只或更多只在伐倒的木材上聚会,“表示其彼此求悦的最强烈愿望”,几经爱抚之后,每只雄鸟便偕其配偶飞去。奥杜邦同样仔细观察过成群的野生加拿大雁(Anser canadensis)并对其求爱的滑稽表演做过图解描绘;他说,以前有过配偶的雁“早在一月就开始重新进行求偶,而其他未曾有过配偶的雁则每天要花数小时去争斗和献媚,直到所有的鸟似乎对各自的选择都感满意为止,以后,它们虽然仍聚集在一起,但任何人都可容易地看出它们是在小心翼翼地保持其配偶。我也观察过越是年长的鸟其求爱序曲就越短。那些独身的雄者和老处女,无论是处于抱恨之中或是不在意那种喧闹的搅扰,而静静地走到一旁,卧于远离其余诸鸟的地方”。 (445) 这位观察家对其他鸟类所做的相似叙述,尚有许多可以引用。
现在让我们转来看看家养和圈养的鸟类,我将从我了解得不多的有关家鸡求偶的情形开始。我曾收到休伊特先生和特格梅尔先生关于这个问题的长信,并还收到过已故布伦特(Brent)先生一篇将近完成的论文。这几位先生由于他们已发表的著作而闻名于世,每个人都会承认他们是细心而有经验的观察家。他们都不相信雌鸟偏爱某些雄鸟是由于后者羽衣美丽的缘故;但必须对这些鸟类长期被养于人为状态下的情况做些考虑。特格梅尔先生相信一只雄斗鸡虽被刈掉垂肉,拔掉颈羽,以至容貌毁损,但它仍会像一只保持着全部自然装饰的雄者那样容易地被雌者所接受。然而,布伦特先生承认,雄鸟的美丽大概有助于刺激雌鸟;而雌鸟的默认也是必要的。休伊特先生认为雌雄二者的结合决不是单纯碰巧发生的,因为雌者几乎总是挑选精力最旺盛、最好斗而且最勇敢的雄者;因此,正如他说的,“如果一只健康良好而有力的雄斗鸡在那个地点活动,要想进行纯种繁育几乎是无效的,因为,几乎每一只雌鸡当离开鸡棚时,都会前去找那只雄斗鸡相会,即使雄斗鸡实际上可能不把和雌鸡变种相同的雄鸡赶走,也是如此”。布伦特先生向我描述说,在正常情况下家鸡的雄者和雌者似乎依靠某些姿势而达到相互了解。但雌鸡对幼小雄鸡的过分殷勤,乃常避之。老母鸡和性情好斗的母鸡,正如同一位作者告诉我的,不喜欢陌生的雄鸡,而且它在被狠狠打得顺从之前,是决不屈服的。然而,弗格森(Ferguson)描述过一只好争吵的母鸡如何被一只上海雄鸡温存的求爱所征服。 (446)
有理由相信雌鸽和雄鸽都喜欢和同品种的鸽子交配;普通家鸽对所有高度改良的品种都不喜欢。 (447) 哈里逊·韦尔先生最近听一位可信赖的观察家说,他饲养蓝色的鸽,这种鸽把所有其他颜色的变种,诸如白的、红的和黄的,全都赶走;另一位观察家也说过,有一只暗褐色的雌信鸽经过反复试验之后,还是不能同一只黑色雄鸽相配,但同一只暗褐色的雄鸽马上就配上了。此外,特格梅尔先生养过一只雌蓝色浮羽鸽(turbit),它顽固地拒绝和同品种的两只雄鸽交配,这两只雄鸽曾连续和雌鸽共同关在一起达数周之久;但一放出去,雌鸽马上就接受了向它提供的第一只雄蓝色龙鸽(dragon)。由于它是一种有价值的鸽子,所以把雌鸟和一只银色(即很淡的蓝色)的雄鸽在一起关了许多星期,最后它还是和这只雄鸽交配了。尽管如此,就一般规律而言,羽色对于鸽的交配似乎没有多大影响。特格梅尔先生根据我的请求,把他养的一些鸽子染上了洋红,但它们并没有引起其他鸽子的很多注意。
雌鸽偶尔也会对某些雄鸽感到强烈憎恶,而无任何明显原因可言。例如,积有45年以上经验的包依塔和考尔比说,“当一只雌鸽厌恶一只被弄来和它交配的雄鸽时,尽管雄鸽燃起了爱情的全部火焰,尽管喂以白燕米和大麻仁以增加其情欲,尽管把它们关在一起达六个月乃至一年之久,这只雌鸽还是断然拒绝了雄鸽的求爱。它的殷勤、它的挑逗、它的回旋表演、它那温柔的咕咕叫声,所有这一切都不能引起它的喜爱,也不会使它激动;雌鸽气鼓鼓地蜷缩于笼子的一角,除了饮水和进食以及对雄鸽的纠缠不休而狂怒的时候,它总是蹲在那里不动。” (448) 另一方面,哈里逊·韦尔先生亲自观察过而且听几位养鸽人说过:一只雌鸽偶尔会强烈爱上一只特殊的雄鸽并且为着它而抛弃了原来的配偶。另一位富有经验的观察家里德尔(Riedel)说 (449) ,有些雌鸽性情放荡,它们几乎对任何所遇到的雄鸟的喜爱皆胜过对其原有配偶的喜爱。某些好色的雄鸽,被我们英国的鸟类玩赏家称为“花花鸟”(gay birds)的,是从事风流艳事的能手,以致必须把它们关起来以免去捣乱。
按照奥杜邦的材料,美国的雄野火鸡“有时会向家养的雌鸡求爱,一般都会受到她们的欢迎”。因此,在野生雄鸡和家养雄鸡之间母鸡们显然喜欢前者。 (450)
这里还有一个更奇妙的事例。赫伦(R.Heron)爵士曾大量繁育过孔雀,关于它们的习性,他保存有多年的记载。他说,“雌孔雀常常很偏爱一只特殊的雄孔雀。所有它们都非常偏爱一只老的雄斑孔雀,有一年它被关了起来,但仍可以看到,这些雌孔雀经常聚集在这只雄孔雀的铁丝笼之旁,而且不容许一只黑翼雄孔雀去碰它们。到了秋天,这只雄斑孔雀被放出来了,于是最老的雌孔雀马上向它求爱并获得了成功。翌年,这只雄斑孔雀被关进一个马厩,这时,雌孔雀就全向那只雄孔雀的一个竞争对手求爱了”。 (451) 这只竞争对手乃是雄黑翼孔雀,在我们看来,它比普通孔雀更美丽。
利希滕施泰因(Lichtenstein)是一位优秀的观察家,而且有极好的机会在好望角进行观察,他向鲁道菲(Rudolphi)保证说,雄黑羽长尾鸟在生殖季节饰有长尾羽,如果长尾羽脱落后,雌鸟就会同它脱离关系。我想他所观察的这种鸟类一定是圈养的。 (452) 这里还有一个近似的例子;维也纳动物园主任耶格尔博士(Dr.Jaeger)说, (453) 一只雄白鹇(silverpheasant)战胜了所有其他雄鹇而成为雌鹇所接受的爱侣,可是其羽饰被弄坏之后,它的位置马上就被一只竞争对手顶替了,后者占了上风,然后把整个雉群带走了。
已经表明色彩对鸟类的求偶是何等重要,因此下述事实值得注意:博德曼(Boardman)先生,一位多年在美国北部从事鸟类收集和考察工作的著名人士,在其广泛的经历中从未见过一只白变鸟同另一只鸟相配的;虽然他有机会去观察属于若干物种的白变鸟。 (454) 简直不能肯定,白变鸟在自然状况下不能繁育,因为它们在圈养条件下能够极其容易地进行繁殖。因此,看来我们必须把它们没有相配这一事实归因于它们遭到了其正常色彩的同伙所拒绝。
雌鸟不仅实行选择,而且在少数场合中,还追求雄鸟,或者甚至为占有雄鸟而互相争斗。赫伦爵士说,关于孔雀,最先的求爱总是由雌者进行的;按照奥杜邦的材料,野火鸡的年长雌者也是如此。关于雷鸟,当雄者在一个聚会地点昂首阔步行进时,雌者则在其周围飞来飞去,吸引雄者注意。 (455) 我们知道,有一只驯养的野鸭经过长时间的求偶后,终于将一只不情愿的雄针尾鸭勾引上了。巴特利特先生相信虹雉属(Lophophorus)和其他许多鹑鸡类的鸟一样,天生是一雄多雌者,但不能将两只雌者和一只雄者关在同一个笼里,因为这样它们会激烈相斗。下述有关竞争的例子更加令人惊奇,因为这是一个叙述红腹灰雀的例子,而红腹灰雀通常是终身配偶的。詹纳·韦尔先生把一只颜色暗淡而丑陋的雌鸟引进了他的鸟舍,它马上向另一只已有配偶的雌鸟发动了如此无情的攻击,以致不得不把后者隔离开。新来的雌鸟尽其爱之能事,最后获得了成功,因为它和那只雄鸟交配了;但过了一段时间,雌鸟受到了公正的报应,因为,当其好斗性停息后,它就被原来的雌鸟取而代之,于是雄鸟舍弃了新欢而同旧偶重归于好。
在所有正常场合中,雄鸟对雌鸟是如此热切以致它会接受任何一只雌鸟,就我们所能判断的来说,雄鸟不会选来选去;但是,如我们今后将要看到的,在少数某些类群中这一规律显然还有例外。关于家养的鸟类,我听说过的仅有一例表明,雄者对某些雌者有所偏爱,根据休伊特先生高度权威的材料,雄鸡喜爱年轻母鸡胜于喜爱年老母鸡。相反地,凡雄雉和普通母鸡杂交奏效者,休伊特先生相信,雄雉总是选择年老的母鸡。雄鸡似乎丝毫不受雌鸡色彩的任何影响,而“其爱情最反复无常” (456) :由于某种莫名其妙的原因,雄鸡对某些母鸡表示了断然的憎恶,繁育者虽想尽力矫正这种毛病,也是枉然。休伊特先生告诉我说,有些母鸡即使对本种的雄者也毫无魅力,因此,它们可能同几只雄鸡在整个繁殖季节都被关在一起,但所下的四五十个卵竟被证明无一受精者。另一方面,埃克斯特龙(M.Ekstr m)说,“长尾鸭(Harelda glacialis)的某些雌者据说远比其他雌者受到更多的追求。确实可以常常看到一只雌者被六只或八只好色的雄者所包围”。我不清楚这一叙述是否可靠,但当地猎人射杀这些雌者是为了把它们剥制成媒鸟的。 (457)
m)说,“长尾鸭(Harelda glacialis)的某些雌者据说远比其他雌者受到更多的追求。确实可以常常看到一只雌者被六只或八只好色的雄者所包围”。我不清楚这一叙述是否可靠,但当地猎人射杀这些雌者是为了把它们剥制成媒鸟的。 (457)
关于雌鸟对特殊雄鸟感到偏爱,必须记住,我们只能用类比方法来判断雌鸟实行选择。如果有一位另一星球的居民看见了许多年轻的乡下人在一处集市上追求一位漂亮姑娘并为她而争吵,就像鸟类在一处聚会地点所做的那样,那么,他将根据追求者热心地取悦于姑娘和显示他们的华丽服饰来推论这位姑娘有选择的能力。那么对鸟类来说,实行选择的证据是这样的:它们具有敏锐的观察能力,而且对色彩和声音似乎都有某种审美力。有一点是肯定的,即,雌鸟由于未知的原因偶尔会对特殊雄鸟表示最强烈的憎恶和偏爱。如果雌雄二者在颜色或其他装饰上有所差异,除了很少例外,总是雄鸟装饰得更美,无论这等装饰是永久性的或只是在生殖季节暂时表现的,都是一样。它们在雌鸟之前孜孜不倦地炫耀其各种装饰,发出鸣声并进行奇特的滑稽表演。即使武装良好的雄鸟在大多数场合中也是具有高度装饰的,虽然它们的成功完全是按战争的法则来决定的;而且它们获得这些装饰乃是以某些能力的损失为代价的。在其他场合中,装饰物的获得则是以增加来自猛禽和猛兽的危害为代价的。各个物种的许多雌雄个体集合于同一地点,而且它们的求偶是一件费时甚久的事情。由此看来,甚至有理由猜想同一地区内的雄者和雌者在相互取悦和交配方面并非总是成功的。
那么,根据这些事实和考察我们应作出怎样的结论呢?雄鸟以如此浮夸的姿态和以如此激烈竞争的手段来显示其魅力,难道这是毫无目的的吗?我们相信雌鸟会实行选择并接受最使她喜爱的那些雄鸟的求爱,难道是不正确的吗?雌鸟大概不会有意识地进行周密考虑;但那些最美丽的、或最善于鸣叫的。或最会献殷勤的雄鸟最能使雌鸟激动,或最能吸引她。无须设想雌鸟会研究色彩的每一条纹或每一斑点,譬如说,无须设想雌孔雀会赞赏雄孔雀华丽尾巴上的每个细节——雌者所受到的大概只是一般影响而已。尽管如此,当听到雄锦雉多么仔细地显示其优美的初级翼羽并将其具有眼斑的羽饰竖起到恰当位置以达到充分的效果之后;或者当听到雄金翅雀如何交替地显示其金光闪闪的翅膀之后,我们就不应过于肯定地认为雌鸟不会注意到美的每个细节。如上所述,我们只能根据类比方法来判断雌鸟是实行选择的;而且鸟类的心理能力同我们的并无根本差异。根据这种种考察,我们可以断言,鸟类的交配并非完全靠机会;在正常情况下那些被接受的,是以其种种魅力最能取悦和引起雌鸟激动的雄鸟。如果这一点得到承认,那么在理解雄鸟如何逐渐获得其种种装饰方面就没有太多困难了。一切动物都表现有个体差异,而且,正如人类靠着选择那些他认为最美丽的个体就能改变其家养的鸟类那样,雌鸟经常地或者甚至偶尔地偏爱那些魅力较强的雄鸟,几乎肯定也会导致雄鸟的改变;而这种改变只要同物种的存在不相矛盾,则几乎可以随着岁月的推移而扩大到任何程度。
鸟类的变异性尤其是其第二性征的变异性
变异性和遗传性是选择工作的基础。家养鸟类肯定发生了重大变异,而且它们的变异肯定是遗传了的。鸟类在自然状况下发生了变异而成为不同的族,这一点现在已得到了普遍承认。 (458) 变异可分为两类;一类似乎是自然发生的,我们迄今还不能了解其原因,另一类同周围环境有直接关联,因此同一物种的一切或几乎一切个体所发生的变异是相似的。艾伦先生对后一类情况曾进行过仔细观察, (459) 他指出美国鸟类的许多物种愈往南方其色彩愈逐渐加强,愈往西方内陆干旱平原其色彩则愈变淡。其雌雄二者似乎一般都受到了相等的影响,但有时某一性所受到的影响比另一性所受到的为大。这一结果同下述看法并不矛盾,即,鸟类色彩主要是由于在性选择作用下的连续变异的积累;因为,甚至在雌雄两性已发生了重大分化之后,气候还可能对双方产生相等的影响,或者因某种体质差异对某一性产生的影响比对另一性为大。
同一物种诸成员之间在自然状况下所发生的个体差异是每个人都承认的。强烈显著的突然变异则属罕见;如果这等变异是有益的,它们是否会常常通过选择而被保存下来并传递给后代,还是一个疑问。 (460) 尽管如此,举出少数我所能搜集的主要有关色彩的例子还是值得一做的,——单纯的白化(albinism)和暗化(melanism)则除外。古尔德先生承认只有少数变种存在,乃是人所共知的,因为他把很轻微的差异评价为物种的差异;然而他还说, (461) 靠近波哥大,阔嘴蜂鸟属(Cynanthus)的某些蜂鸟分为两三个族或变种,它们彼此的差异在于尾羽的颜色——“有的整个尾羽呈蓝色,而其他只有八支中央尾羽的尖端呈美丽的绿色”。在这个场合以及下述一些场合中,似乎没有观察到中间的级进。有一种澳大利亚长尾鹦鹉(parrakeets),只有某些雄者的“大腿呈猩红色,而其他雄者的大腿则为草绿色”。另外还有一种澳大利亚长尾鹦鹉,“它们某些个体的翼覆羽上有一鲜黄色横带斑,而其他个体的同一部位则为红色”。 (462) 在美国,猩红色的红灰雀 (463) (Tanagra rubra)的某些少数雄者“在较小的翼覆羽上有一条美丽的红光闪闪的横带斑”; (464) 但是,这种变异似乎多少是罕见的,因此它只有在特殊有利环境下才能通过性选择而被保存下来。在孟加拉,蜂鹰(Pernis cristata)或在其头顶上有一个小型的痕迹羽冠,或完全没有:然而,如果不是印度南部的这同一物种具有“由若干渐次变化的羽毛所形成的一个十分显著的后头羽冠”, (465) 那么上述那种非常轻微的差异就不值得注意了。
下述事例在某些方面更为有趣。渡鸟的一个黑白斑变种只限于费罗群岛(FeroeIslands)才有,其头部、胸部、腹部以及翼羽和尾羽的一部分均呈白色。这个变种在该处并不少见,因为格拉伯(Graba)在访问那里期间曾见过8~10只活标本。尽管这个变种的性状不十分稳定,却仍然被几位著名的鸟类学家定为一个独特的物种。这种黑白斑鸟受到岛上其他渡鸟大吵大闹的追求和迫害,这一事实是使布吕尼哈(Brünnich)断定它们是一个独特物种的主要原因;但现在已经知道这是一个错误。 (466) 这个事例同刚刚举出的下述事例似乎是相似的,即,白化的鸟类由于遭到其同伙的拒绝而不能交配。
在北方海域的各个不同部分都发现有普通海雀(Uria troile)的一个显著变种,而在费罗群岛,据格拉伯估计,每五只鸟中就有一只发生了这样的变异。其特征为眼睛周围有一纯白色的圈,从白圈向后伸出一条弯曲的白色窄线条,长达一时半。 (467) 这个显著的特征使几位鸟类学家把这种鸟列为一个独特的物种,命名为U.lacrymans,但现在已弄清它不过是一个变种而已。它常和普通种类交配,然而从未见有中间级进;这也不足为奇,因为那些突然发生的变异,如我在别处所指出的, (468) 往往是不变地传递下去,要不就是完全不传递。于是,我们看到同一物种的两个不同类型可在同一地区共存,我们不能怀疑如果其中某一个类型具有超出另一个类型的任何优势,则它就会迅速地成倍增殖起来而把后者排斥掉。例如,如果黑白斑的雄渡鸟不是受其同伙的迫害,而是能够高度吸引黑色雌渡鸟(像上述黑白斑雄孔雀那样),那么它们的数目就会迅速地增加起来。这大概就是性选择的一例子。
就同一物种一切成员所共有的轻微个体差异而言,不论其程度大小如何,我们有各种理由可以相信这等差异对于选择工作是最重要的。第二性征是显著易于变异的,无论对自然状况下的动物还是对家养状况的动物来说,都是这样。 (469) 还有理由相信,如我们在第八章中所看见的那样,雄者比雌者容易发生变异。所有这等偶然发生的情况都是高度有利于性选择的。这样获得的性状究竟是传递给雌雄中的一性还是传递给雌雄两性,如我们将在下一章看到的,乃决定于遗传形式。
鸟类雌雄二者之间的某些轻微差异,究竟是单纯地由于变异受到了限于性别的遗传,而不借助于性选择;还是这等轻微差异通过性选择的作用而被扩大了,对此难以形成一种见解。我在此没有论及雄鸟显示华丽羽彩和其他装饰物而且雌鸟在这方面也稍有表现的许多事例,因为这几乎肯定是由于最先由雄鸟获得的性状或多或少地传递给雌鸟了。但关于某些鸟类,譬如说,其雌雄二者的眼睛在色彩上有轻微差异者,我们又该怎样来做结论呢? (470) 在某些场合中,雌雄二者的眼睛差异显著;例如黑颈鹳属(Xenorhynchus)属的鹳(stork),其雄者的眼睛为淡黑褐色,而雌者的眼睛则为橙黄色;我听布莱思先生说 (471) ,雄犀鸟的眼睛呈强烈的艳红色,而雌者的眼睛则为白色。关于犀鸟(Buceros bicornis),其雄者的头羽后缘以及喙部突起上的一道条纹均呈黑色,而雌者并不如此。我们可否假设雄鸟的这等黑色标志以及眼睛的艳红色彩系通过性选择而被保存下来或被扩大的呢?这是很有疑问的;因为巴特利特先生在伦敦动物园中向我说明,雄犀鸟嘴的内侧为黑色而雌犀鸟嘴的内侧则为肉色;至于它们的外貌或它们的美并不受这样影响。我在智利见过一只一岁左右的新域鹫(condor),其眼睛虹彩为暗褐色,到了成年,其雄者的眼睛虹彩就变为黄褐色,而雌者的则变为鲜红色。 (472) 这种神鹰的雄者还有一个小而长的铅色肉冠。许多鹑鸡类的肉冠都是高度富有装饰性的,而且在求偶活动中其色彩变得鲜艳;但是,神鹰的铅色肉冠在我们看来一点也没有装饰性,对此我们又该做何解释呢?关于各种其他性状,也可以提出同样的问题,例如鸿雁(Anser cygnoides)喙基上的瘤状物,在雄者就比在雌者大得多。对于这些问题均无法作出确切的回答;但是,我们在假设那些瘤状物以及各种肉质附器对雌者不会有吸引力时,务必要慎重;如果我们想到未开化人的种种可怕的毁形风俗——面部的深刻伤痕使肌肉突出而成为若干肉疙瘩,用细枝或骨头穿透的鼻壁,大大拉开的耳孔和唇孔——全都作为装饰而受到赞赏,我们就知道为什么在作出上述假设时一定要慎重了。
雌雄二者之间无关紧要的差异,诸如上面所举出的那些,不管是否通过性选择而被保存下来,这等差异以及其他所有差异最初一定是由变异法则来决定的。根据相关发育的原理,羽毛常常在身体的不同部位或在全身按照同样的方式发生变异。我们看到家鸡的某些品种在这方面提供了很好的例证。所有这些品种的雄者,其颈部和腰部的羽毛都延长了,因而被称为长绒羽(hackles);那么,当雌雄二者都获得了作为该属一种新性状的顶结时,雄者头部羽毛则变为长羽状,这显然是由于相关原理起的作用所致;而雌者头部羽毛仍保持正常形状。构成雄者顶结的长羽在色彩上也常常同颈部和腰部的长羽相关,例如,我们把金斑和银斑波兰品种的这等羽毛,以及把霍丹鸡(Houdans) (473) 、V-形肉冠鸡(Crève-caeur) (474) 等品种的这等羽毛,加以比较,就可看出上述情形。关于某些生活于自然界的物种,我们可以观察到这等相同的羽毛在色彩上完全一样的相关,例如华丽的金雉和云实树雉的雄者就是如此。
各个单独羽毛的构造一般会致使羽色的任何变化成为对称的;我们在家鸡的花边品种、亮斑品种以及条纹品种中可以看到这种现象;根据相关原理,全身羽毛常常是按照同样方式着色的。因此,我们不必费多大劲就可育出羽色同自然物种一样对称的品种来。花边品种和亮斑品种的羽毛边缘的颜色,其界限是截然分明的;但是,我用一只带有绿色光泽的雄西班牙黑鸡同一只白色雌斗鸡杂交,育成了一个杂种,这个杂种的全身羽毛全是黑中略带微绿,只有每根羽毛的尖端是白中略带微黄;不过在每根羽毛白色顶端和黑色基部之间有一个弯曲而对称的暗褐色区域。在某些事例里,羽轴决定着羽色的分布范围;例如,用同一只雄西班牙黑鸡和雌银斑波兰鸡杂交所育出的一只杂种鸡,其体部羽毛的羽轴及其两侧的一窄条部位全是黑中略带微绿,它又被一个有规则的暗褐色区域所环绕,其边缘则系白中略带微褐。在这些事例中,我们看到羽毛的颜色都是对称的,就像许多自然物种的情形那样,这等对称的颜色使其羽衣增添了无限的华丽。我还注意过普通家鸽的一个变种,其翼部带斑对称地环以三种鲜明色调的羽毛,而不像其亲种那样,这等带斑只是单调地在石板青的底色上呈现出黑色而已。
鸟类有许多类群,其若干物种的羽色虽不相同,但全都保持着一定的点斑、块斑或条斑。一些鸽的品种也有类似的情况,它们通常都保有两条翼带斑,尽管带斑的颜色可以是红的、黄的、白的、黑的或蓝的,而羽毛的其余部分则呈现某种完全不同的色彩。这里还有一个更为奇妙的事例:某些斑记的颜色虽然同自然物种的这等斑记的颜色差不多完全相反,但这等斑记仍被保持着;原鸽有一条蓝色的尾,其中两根外尾羽的外部羽瓣位于末端的那一半呈白色;于是出现了一个亚变种,它的尾部不是青色,而是白色,而且原种呈白色的那一部分显然变为黑色的了。 (475)
鸟类羽衣眼斑的形成及其变异性
装饰物之美没有过于各种鸟类羽毛上的、某些哺乳类毛皮上的、爬行类和鱼类鳞片上的、两栖类皮肤上的、许多鳞翅类和其他昆虫翅膀上的眼斑,因此它们值得特别予以注意。一个眼斑是由一个斑点围以另一种颜色的圆环所构成的,犹如瞳孔位于虹彩之内一样,但其中央的斑点往往被附加的若干同心色带所环绕。孔雀尾覆羽上的眼斑以及孔雀蛱蝶(Vanessa)翅上的眼斑向我们提供了一个人所熟知的例子。垂门先生给过我一份描述一种产于南非的蛾(Gynanisa isis)的材料,它同英国的天蚕蛾有亲缘关系,这种蛾每张后翅的全部表面差不多被一个壮丽的眼斑所占满;这个眼斑含有一黑色中心,其中有一个半透明的新月形斑,其外挨次围以赭黄的、黑的、赭黄的、桃红的、白的、桃红的、褐的以及白的色带。虽然我们还不了解这等异常美丽而复杂的装饰物的发展步骤,但其过程似甚简单,至少对昆虫类来说是这样;因为,正如垂门先生所函告的那样,“在鳞翅类中作为单纯斑记或色彩的诸性状,没有一种像眼斑那样不稳定的,无论其数目还是其大小都是如此。”华莱士先生最先使我注意到这个问题,他给我看过英国普通草地尺蠖(Hipparchia janira)的一套标本,这套标本显示了由一个简单的小黑点到一个色调优美的眼斑之间有大量的级进。在同一科中还有一种产于南非的莉达蝶(Cyllo leda,Linn.)。其眼斑甚至更容易变异。它的一些标本(图53,A)的翅膀上表面大部分作黑色,其中有不规则的白色斑记;从这种状态到一个相当完善的眼斑(A′)之间,可以追踪出一套完整的级进这个完善的眼斑乃是由不规则色斑的收缩而形成的。在另一套标本中,从非常小的一些白斑点环以勉强看得见的黑线这种状态(B)到一个完全对称的大眼斑(B′) (476) 之间也可以找出它们的级进。在与此相似的一些场合中,一个完善眼斑的发展并不需要一个变异和选择的长过程。
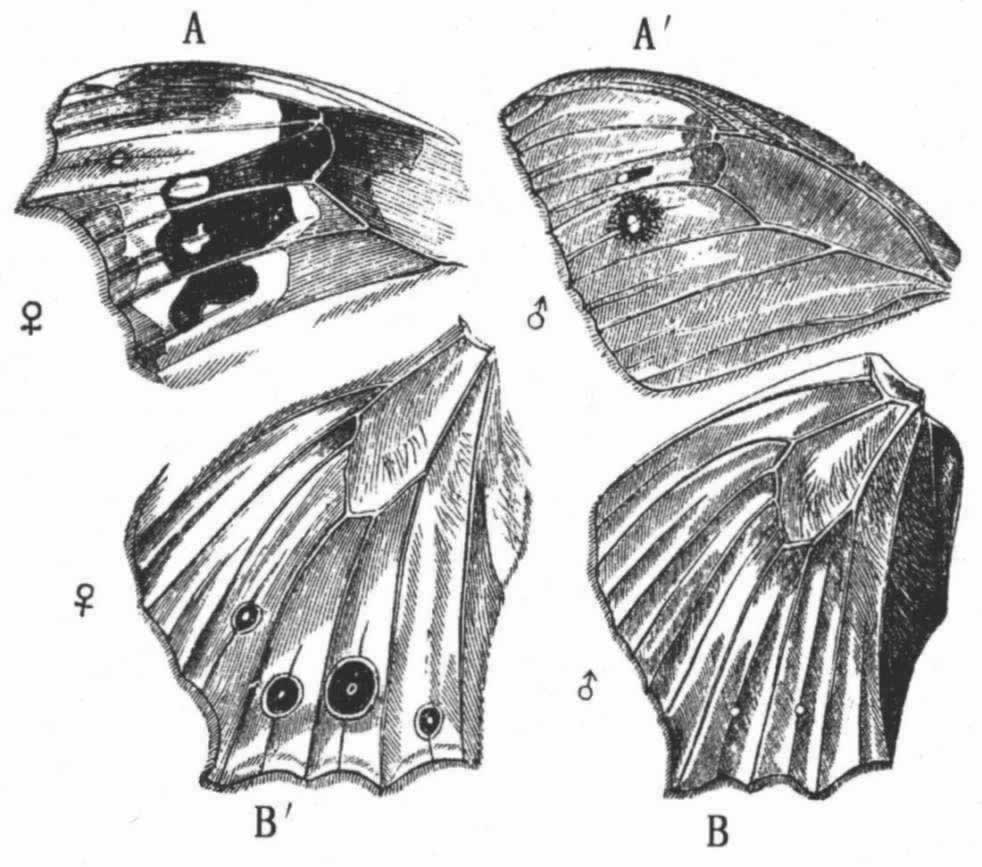
图 53 莉达蝶(Cylloleda,Linn.)
(采自特里门先生的绘图,表明眼斑有极广泛的变异范围)
A.采自毛里求斯的标本, B.采自爪哇的标本,
图示前翅上表面。图示后翅上表面。
A′.采自纳塔尔的标本,同上。 B′.采自毛里求斯的标本,同上。
关于鸟类以及其他许多动物,根据亲缘相近的物种比较的结果,似乎圆斑的产生常常是由于条纹的断裂和收缩。就红胸角雉来说,其雌者身上模糊不清的白线正是相当于雄者身上那些美丽的白斑点; (477) 在锦雉的雌雄二者身上也可观察到多少类似的情况。不论其形成原因如何,它的外观支持了下述信念;即,从一方面来看,一个黑点往往是因有色物质从周围区域向中心点收缩而成,因而其周围区域的颜色因此变淡;从另一方面来看,一个白点则往往是因有色物质从一中心点被驱散而成,因而其周围区域由于有色物质的集聚而加深。无论在哪一种场合中,其结果都会导致一个眼斑的形成。这等有色物质的量大概差不多是固定的,但它既可向心地也可离心地重新分布。普通珍珠鸡(guinea-fowl)的羽毛提供了一个良好的例子,表明白色的斑点环以较暗的色带;凡是在白色斑点既大而彼此接近的地方,则周围的暗色诸带就会融合在一起。在锦雉的同一根翼羽上既可观察到黑色斑点为一淡色带所环绕,又可观察到白色斑点为一暗色带所包围。因此,一个最基本状态的眼斑的形成看来是一件简单的事情。至于进一步还要经过那些步骤才能产生那些更为复杂的、依次环以许多层色带的眼斑,我不敢妄加评说。但是,不同颜色的家鸡所产生的杂种,其羽毛具有色带,并且鳞翅类昆虫的眼斑具有异常大的变异性,这两种情况可以引导我们作出下列结论:眼斑的形成并不是一个复杂的过程,而是决定于相邻组织的性质所发生的某种轻微而逐渐的变化。
第二性征的级进
级进的情况是重要的,因为它向我们表明了高度复杂的装饰物可由一些连续的小步骤而获得。为了发现任何现存鸟类的雄者获得其华丽的色彩或其他装饰物所经过的实际步骤,我们就应该追溯其绝灭的祖先的悠久系谱;但这显然是不可能做到的。然而,一般我们可以用比较同一类群所有物种的方法——如果它是一个大类群的话,找到一点头绪;因为它们之中有些大概还会保存、至少部分地保存其以往性状的痕迹。在各个类群中固然有一些关于级进的显著事例可举,但为了避免讨论那些烦琐的细节,最好的办法似乎是对一两个非常典型的事例加以研究,例如孔雀的事例,看看这样是否可以说明这种鸟的装饰经过了怎样步骤而变得如此华丽。雄孔雀之所以引人注目,主要在于其尾覆羽特别长;而尾羽本身并没有延长多少。几乎沿着尾羽全长的尾枝都是分离的或分解的;但许多物种的羽毛以及家鸡和家鸽的羽毛也是如此。这些羽枝向羽干的末端合拢而形成一个椭圆形的圆盘或眼斑,它肯定是世界上最漂亮物体之一。它包含一个闪光的、深蓝色的锯齿状中心,环以一层鲜绿的色带,其外又环以一层铜褐色的宽色带,在这层宽色带外面又环以五层彼此略有不同的闪光的窄色带。在圆盘上有一种微小的性状值得注意;沿着某一同心环带的羽枝或多或少地都缺少小羽枝,所以圆盘的一部分被一个几乎透明的环带所围绕,使它具有一种非常精致完美的外观。不过我在别处也曾描述过 (478) 雄斗鸡的一个亚变种,其颈部长羽的变异同上述情形完全相似,具有金属光泽的这种长羽顶端,“由一个对称形状的透明环带把它同其下的羽毛隔开,这个透明环带系由羽枝的无毛部分构成的”。眼斑的蓝黑中心的下缘或底部在羽干线上成深锯齿形。周围的色带同样显露了缺刻甚至破裂的痕迹,如图54所示。这些缺刻是印度孔雀(Pavo cristatus)和爪哇绿孔雀(P.muticus)所共有的,鉴于这等缺刻同眼斑的发展可能有关系,似乎值得特别注意;然而长期以来我未能猜出其意义何在。
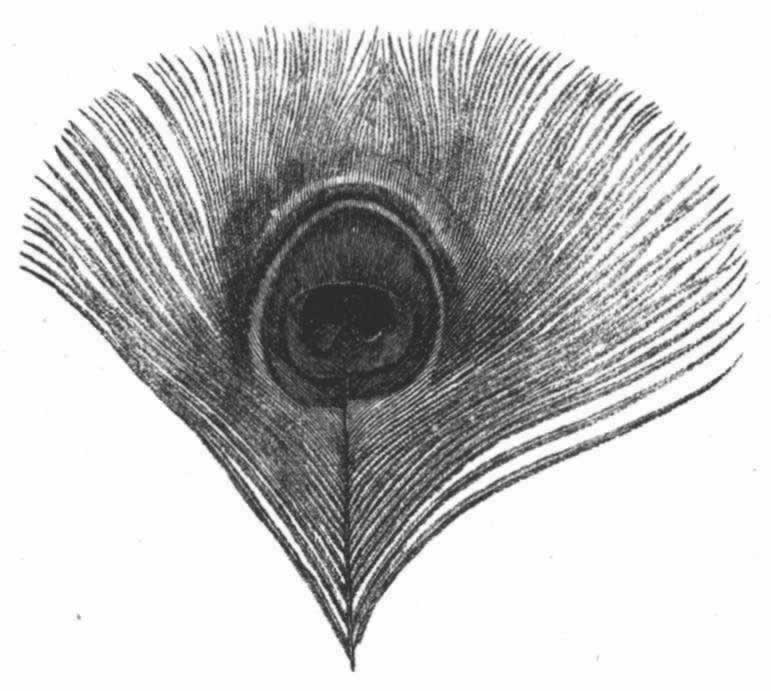
图 54 孔雀的羽毛,福特(Ford)先生绘,
透明的环带以最外面的白色环带代表之,只限于圆盘的上端
如果我们承认逐渐进化的原理,那么在孔雀那特别长的尾覆羽和所有普通鸟类的短尾覆羽之间;还有,在孔雀那壮丽的眼斑和其他鸟类的比较简单的眼斑或仅仅是有色的斑点之间,必定有许多体现了每个连续步骤的物种存在过,至于孔雀的所有其他性状亦复如此。让我们通过亲缘相近的鹑鸡类来看一看今天依然存在的任何级进。团花雉的物种和亚种所栖息的地方同孔雀的原产地相毗邻;它们同孔雀如此相似,以致它们有时也叫做孔雀雉(peacock-pheasants)。巴特利特先生也向我说过,团花雉在鸣声和某些习性方面也同孔雀相似。如上所述,其雄者于春季期间,在色彩相对平淡的雌者之前大摇大摆地走来走去,展开并竖起它们的尾羽和翼羽,其上装饰着大量的眼斑。我请读者再看一下前面那幅团花雉的图(图51)。拿破仑团花孔雀雉(P.napoleonis)身上的眼斑只限于尾羽才有,其背部呈华丽的蓝色,具金属光泽;这个物种在这些方面都接近于爪哇孔雀。哈德团花孔雀雉(P.hardwickii)有一个特殊的顶结,同爪哇孔雀的顶结多少相似。所有物种的翼眼斑和尾眼斑不是圆的就是椭圆的,这种眼斑含有一个闪光的蓝绿色或紫绿色的美丽圆盘,圆盘的周围环以黑色的边缘。这个黑色边缘在成吉思团花孔雀雉(P.chinquis)身上逐渐向外变为褐色,镶着淡黄的边,因此这里的眼斑是由各种不同色调的、但不明亮的同心色带环绕着。团花雉另一个显著的性状是它的尾覆羽特别长;因为在某些物种中其尾覆羽为真尾羽的一半长,在另外一些物种中,其长度为真尾羽的三分之二。其尾覆羽就像孔雀那样地具有眼斑。这样,团花雉的几个物种在其尾覆羽的长度、眼斑的环带以及其他一些性状方面都明显地向着孔雀逐渐接近。
尽管有这种接近,但我检查的第一个团花雉的物种几乎使我放弃这方面的探索;因我不仅发现其真尾羽装饰着眼斑——孔雀的真尾羽则完全没有这种装饰,而且其所有羽毛的眼斑都同孔雀的眼斑有根本差异,在团花雉的同一根羽毛上有两个眼斑,各居羽干的一侧(图55)。因此,我断定孔雀的早期祖先不能和团花雉相似。但随着我的研究继续深入,我观察到某些物种的那两个眼斑彼此挨得很近;在哈德团花雉的尾羽上它们相互接触了;而且这同一物种尾覆羽上的以及马六甲团花孔雀雉(P.malaccense)尾覆羽上的两个眼斑终于实际上融合在一起了(图56)。由于融合起来的只是中央部分,因此在上下两端都有一个缺刻,其周围的色带同样也有缺刻。一个简单的眼斑就这样在每根尾覆羽上形成了,虽然它还明显地表现着两个眼斑的来源。这等融合而成的眼斑和孔雀的单个眼斑之间的差异在于前者的上下两端都有缺刻,而不像后者那样只在下端或底端才有缺刻。然而,这种差异并不难以说明;关于团花雉的某些物种,其同一根羽毛上的两个卵形眼斑彼此平行;其他物种(如成吉思团花孔雀雉)的那两个眼斑则向一端收敛;那么这两个收敛的眼斑的局部融合将会在岔开的一端比在收敛的一端明显地留下一个深得多的缺刻。如果这种收敛极其显著而且融合得完全,那么在收敛一端的缺刻就会趋于消失。
图 55 成吉思团花孔雀雉的尾覆羽局部,其上有两个眼斑,原大

图 56 马六甲团花孔雀雉(Polyplectron malaccense)的尾羽局部,具有两个眼斑,原大,部分地融合在一起
有两个孔雀的物种,其尾羽完全没有眼斑,这显然同它们被长的尾覆羽所掩盖有关系。在这一点它们同团花雉的尾羽显著不同,大多数团花孔雀雉的尾羽上的眼斑都大于尾覆羽上的眼斑。因此,这引导我对若干物种的尾羽进行了仔细检查,其目的在于发现它们的眼斑是否有任何消失的倾向;使我感到很满意的是,情况似乎正是这样。拿破仑团花雉的中央尾羽在羽干的两侧各有一个充分发达的眼斑;但越靠外边的尾羽其内侧的眼斑就越来越不显著,到了最外边的那根尾羽,其内侧眼斑就只剩下了一个暗影或痕迹而已。此外,马六甲团花孔雀雉(P.malaccense)尾覆羽上的那两个眼斑,就像我们已经看到的那样,融合起来了;而且这等尾覆羽特别长,竟达尾羽长度的三分之二,因此在这两方面马六甲团花孔雀雉都同孔雀接近。那么,在马六甲团花孔雀雉中,只有两根中央尾羽有所装饰,每一根中央尾羽有两个色彩明亮的眼斑,所有其他尾羽的内侧眼斑则全消失了。结果,团花雉这个物种的尾覆羽和尾羽在构造和装饰这两方面都很接近于孔雀的相应羽毛。
按照级进原理既然可说明孔雀获得其华丽尾羽所经历的步骤,那么几乎不需要再多谈什么了。如果我们给自己勾画出一个孔雀的祖先,它几乎完全处于一种中间状态,介乎现存孔雀和一种普通鹑鸡类的鸟之间,前者具有大大延长了的并装饰着单个眼斑的尾覆羽,而后者的尾覆羽则是短的,其上仅有某一种颜色的斑点,那么我们将会看到一种同团花雉相似的鸟——这就是说,这种鸟具有能竖起和展开的尾覆羽,其上装饰着两个局部融合起来的眼斑,它的尾覆羽特别长,几乎足可以把尾羽掩藏起来,而尾羽上的眼斑已部分地消失了。两个孔雀种的眼斑中心圆盘的缺刻及其周围色带的缺刻都明显地表明它们同这个观点是吻合的,否则这种缺刻就无法得到解释。团花雉的雄者无疑是美丽的鸟,但从稍远的地方去看,它们的美就无法同孔雀相比了。许多雌孔雀的祖先,在其由来的悠久系统中必定欣赏这等优越性,因为通过对最漂亮雄者的不断选择,它们已无意识地使雄孔雀变成了现存鸟类中的最佼佼者。
锦 雉
另一个可供研究的极好事例乃锦雉翼羽上的眼斑,其色彩浓淡适宜,令人惊异,犹如松松地置于穴中的诸球,因而和普通眼斑有所不同。我想没有人会把这种曾激起许多有经验艺术家赞叹的色调归因于偶然——即有色物质的原子之偶然汇集。如果认为这等装饰物的形成是通过对许多连续变异的选择,而其中没有一种变异打算产生“球与穴”的效果,那么这种说法之不可信,犹如认为拉斐尔(Raphael)所画圣母马利亚像是由于对青年艺术家长期连续的乱涂胡抹所进行的选择的结果,而其中没有一位艺术家曾经最初打算过去画人体像的。为了发现眼斑是如何发展起来的,我们无法追溯其悠久的祖先系统,也无法考察其许多亲缘密切相近的类型,因为它们目前已不复存在了。但幸运的是,某些翼别足可以给我们提供一个解决问题的线索,它们可以证明从一个简单的斑点逐渐发展成一个像“球与穴”那样精致完美的眼斑至少是可能的。
具有眼斑的翼羽布满黑色条纹(图57)或数行黑色斑点(图59),每根条纹或每行斑点皆自羽干外侧斜趋向下而达于一个眼斑。那些斑点一般延长成一条线而横过其所在的那一行。它们往往会合起来,使其所在的那一行连成一线——这时便形成了纵条纹——或是横向会合,即由相邻诸行的斑点会合起来,这时便形成了横条纹。有时一个斑点会分裂为若干小斑点,它们仍位于其固有的位置。
为方便起见,我们先描述一个形似“球与穴”的完整眼斑,这种眼斑包含一个漆黑的圆环,圆环之内的部分着色浓淡非常适宜,使其恰似一个球。这里刊出的图系由福特先生精巧绘制的,而且雕刻甚佳,不过一幅木刻图是无法显示出原来优美色调的。这个圆环差不多总是略有破裂或中断(见图57),中断之处在上半部的某一点,位于球外白影上方略偏右之处;有时圆环也在右侧靠基部处破裂。这等小破裂具有一种重要意义。圆环靠左上角处总是大大变粗,这里的边缘界限模糊不清,这根羽毛是直竖的,其位置如图所示。在变粗的那一部分下面,有一道几乎纯白的倾斜斑记位于球的表面,往下颜色逐渐变淡,先变成一种铅灰色,再变成黄色,而后为褐色,于是朝着球的下部再徐徐地越变越黑。正是这种色调当光线照射到一个凸面时便产生了如此令人赞叹的效果。如果检查一下其中的一个球,就会看到其下部系褐色,它同上部被一条斜曲线所模糊地分开,上部颜色较黄,铅色也较深;这道斜曲线同白色光块的长轴、确实也同所有色调的长轴相垂直;当然这种颜色差异是不能在木刻图上表现出来的,但这种颜色差异一点也不妨碍这个球的完整色调。特别要加以观察的是,每个眼斑都和一根黑条纹、要不就和一纵行黑斑点明显地相连,因为二者都出现于同一根羽毛之上,并无差别,如图57所示,条纹A走向眼斑a;条纹B走向眼斑b;条纹C的上部断裂了,它走向下一个眼斑,但在木刻图上没有表示出来;条纹D又走向更下的一个眼斑,条纹E和F则照此类推。最后,这几个眼斑彼此被一个带有不规则黑色斑记的淡色表面所分开。
图 57 绵雉次级翼羽的一部分,图示两个完整的眼a和b、A、B、C、D等乃暗色条纹,斜趋向下各至一个眼斑(羽干两侧的羽瓣尤其是在羽干左侧者,大部分被去掉了)

图 58 次级翼羽的基部,最靠近身体的部位。
其次我将描述这个系列的另一极端,即一个眼斑的最初痕迹。那些短次级翼羽(图58)最靠近身体的部分就像其他羽毛那样,具有斜走的、纵向的、不甚规则的数行颜色很暗的斑点。其下方五行(最下一行除外)的基部斑点,即距羽干最近的斑点,比同行的其他斑点略大并在横的方向略长。它和其他斑点的差别还表现在其上部边缘是以某种模糊的暗黄色调为界的。但这个斑点在任何方面并不比许多鸟类羽衣上的那些斑点更惹人注目,因而容易被忽略掉。在它上方的那个斑点和同行上部的其他斑点就完全没有差别了。短次级翼羽的这等较大的基部斑点所在的位置正是较长翼羽的完整眼斑所在的相应位置。
通过对依次的另外两三根翼羽的观察,可从刚刚描述的那个基部斑点以及在它上面的那个同一行斑点到一个不能称为眼斑的奇妙装饰物——由于还没有更好的名称,所以我命名它为“椭圆装饰物”,可以追踪出一个绝对不知不觉的级进过程。所有这些都在图59中示明。我们在这里看到具有平常性状的几行斜趋的暗色斑点A、B、C、D等(参阅右侧的文字图解)。每行斑点都下趋至一个椭圆装饰物并与之相连,其方式正同图57所示的每根条纹下趋至一个形似“球与穴”的眼斑并与之相连的情况完全一样。拿任何一行来看,譬如图59的B行,其最下方的斑点b比它上面的那些斑点更粗,而且长得多,其左端变尖并向上弯曲。这个黑斑的上边突然出现一个具有鲜艳色调的宽阔部分,始于一条褐色狭带,然后逐渐变为橙色,由橙色又逐渐变为一种淡铅色,其向羽干的那一端颜色还要淡得多。这等浓淡具备的色彩充满了椭圆装饰物的整个内部。斑记(b)在每个方面都同上一节(图58)所描述的简单羽毛的那个浓淡适宜的基部斑点相当,只不过是发达得更为高度、而且色彩更为鲜明而已。这个斑点的右上方色调鲜明,那里有一个属于同一行的窄而长的黑斑(c),稍微向下弯曲,正好与(b)相对。这个黑斑有时断裂为两个部分。其下部边缘也是窄的,呈暗黄色。c的左上方尚有另一个黑斑(d),亦居于同一倾斜的方向,但总是或多或少地不同于c。这个斑记一般为亚三角形,而且形状不规则,但这个图解中所示的,它异常地窄而长并且是规则的。它显然包含斑点(c)已断裂的横向延长部分,以及与之会合的上面那个斑点已断裂的延长部分;但关于这一点我还不能肯定。这三个斑记b、c和d,以及它们之间的明亮色调一起构成了所谓的“椭圆装饰物”。这些装饰物同羽干平行,其位置明显地同那些形似“球与穴”的眼斑的位置相当。我们无法从图中来欣赏其非常优美的外观,因为橙色和铅色同黑斑之间的衬托如此之美,那是无法从该图显示出来的。

图 59 一根次级翼羽靠近身体的部分,示明所谓的“椭圆装饰物”。右图仅为文字
说明的图解:A、B、C、D等分别代表下趋的诸斑点行列以及形成的椭圆装饰物。
b.为B行最下方的斑点或斑记;c.同一行中的次一斑点或斑记;
d.显然是同一B行的斑点C已经断裂的一个延长部分。
在椭圆装饰物和形似“球与穴”的完善眼斑之间有如此完整的级进,以致几乎不可能决定应该何时使用眼斑这一术语才是。从前者过渡到后者是这样来完成的,即,下面的黑斑(图59,b)伸长并朝上弯曲,尤其是上面的黑斑(c)更为如此,同时那个伸长的亚三角形,即那个狭斑(d)收缩,所以这三个斑记最后会合在一起,形成一个不规则的椭圆环。这个环逐渐变得越来越圆,越来越规则,同时扩大了其直径。我在此提供一幅按天然大小画下来的一个尚未十分完善的眼斑图(图60)。黑环的下部比椭圆装饰物(图59,b)的下部斑记弯曲得多。黑环的上部包含两三处分开的部分;形成白影上方那块黑斑的部位只有一点变粗的痕迹。这白影本身尚未十分集中;而且在其下方的表面比一个形似“球与穴”的完整眼斑的色彩更为明亮。即使在最完善的眼斑中,也可以观察到形成圆环的那三四个伸长的黑斑的接合。这个不规则的亚三角形或狭斑(图59,d),通过它的收缩和均等化,明显地在形似“球与穴”的完善眼斑的那个白影上方形成了圆环的加粗部分。圆环的下部总是比其他部分略粗一些(见图57),这是由于椭圆装饰物的下部黑斑(图59,b)原来就比其上部的黑斑(c)为粗。会合和改变的过程所经历的每个步骤都能被追查出来;围绕圆形眼斑的黑环无疑是由椭圆装饰物的那三个黑斑b、c、d所形成的。相邻眼斑之间的那些不规则的弯曲黑斑显然都是由于椭圆装饰物之间的那些较为规则、但彼此相似的黑斑破裂所成。
图 60 一个中间状态的眼斑,介于椭圆装饰物和形似“球与穴”的完善眼斑之间。
图 61 一根具有形似“球与穴”完善眼斑的次级翼羽近末端的部分
a.具有装饰的上部;b.最顶端的形似“球与穴”的不完善眼斑(眼斑顶端的白色斑记光影在这里显得太暗了点);c.完善的眼斑。
形似“球与穴”的眼斑,其色调形成的连续步骤也同样可以清楚地被追查出来。那些褐色的、橙色的和淡铅色的狭带构成了椭圆装饰物下部黑斑的界线,可以看到它们的颜色变得越来越弱并且逐渐融合起来,上面颜色较明亮的那一部分靠左角愈益变得明亮,以致几乎变成白色,同时也更加收缩。如上所述,甚至在形似“球与穴”的最完善眼斑中,还可以察觉出球的上部和下部之间在色彩上有一种轻微差异,虽然并非色调上的差异;球的上部和下部的分界线是斜的,其倾斜的方向正如椭圆装饰物的具有明亮颜色的光影的方向。于是可以阐明,形似“球与穴”的眼斑在形状和颜色上的几乎每一个细节都是来自椭圆装饰物的逐渐变化;而且从两个差不多是简单的斑点的结合,通过同等的小步骤,可以追踪出椭圆装饰物的发展过程,居于下方的那个斑点的上部边缘呈暗淡的黄褐色。
具有形似“球与穴”的完善眼斑的次级长羽末端都有特别的装饰(图61)。那些斜的纵条纹向上突然中止并相互混合起来;在这个界限之上的整个羽毛上端(a)布满了白色小点,围以黑色小环,位于暗色背景之上。属于最上眼斑(b)的斜条纹仅仅成为一个很短的不规则黑斑,仍具有平常那样的横向弯曲的底部。由于这个条纹是那样突然地断掉了,因此我们根据前面所发生的一切,也许能够理解这个圆环上方的加粗部分是如何在这里消失的;因为,如上所述,这个加粗部分显然同上面那个较高斑点已断裂的一个延长部分存在着某种关系。由于圆环上部加粗部分的缺如,最高的那个眼斑尽管在其他所有方面都是完善的,但其顶端好像斜斜地被削去了一块。我想,任何一个人如果认为锦雉的羽衣一创造出来就像我们现在所看到的那样,那么在他说明最高眼斑的不完善状态时就会感到困惑。我应该再作一点补充,即距离身体最远的那些次级翼羽上的所有眼斑都比其他羽毛上的眼斑为小,而且较不完善,其圆环的上部缺如,恰如刚才提到那种情况一样。这种不完善的情况在此似乎同下述事实有关,即,这种羽毛上的斑点会合成条纹的倾向比通常的情况为小;相反,它们往往断裂成较小的斑点,所以有两三行斑点走向同一个眼斑。
现在留下来的还有一个很奇妙的问题值得注意,这是伍德(Wood)先生首先观察到的。 (479) 伍德先生给过我一张照片,其上为一个进行夸耀自己的人工制作标本,可以看到其垂直举起的羽毛上诸眼斑的白色斑记皆在上端或最远的一端,也就是对着上方,这一白色斑记体现着从一凸面反射出来的光线;当这只鸟在地面上夸耀自己时光线自然是从上面照射下来的。但妙处就在于此:它的外部羽毛保持着几乎水平的状态,其眼斑所处的位置似乎应该适于接受来自上方的光照,因而那个白色斑记应该位于眼斑的上侧;它们的位置果然如此,真是令人不可思议!因此,若干羽毛上的眼斑就光线而言虽处于很不相同的位置,但看来好像光线都是从上方照射的一般,恰如一位美术家给它们涂上了浓淡适宜的色彩一样。尽管如此,它们并非严格地从同一点接受光照,像它们应该表现的那样;因为保持几乎水平状态的羽毛上诸眼斑的白色斑记位置过于接近较远的一端;也就是说它们并非完全横向的。无论如何我们无权期望通过性选择所获得的具有装饰性的那一部分是绝对完善的,正如通过自然选择所获得的具有实际用途的那一部分也不是绝对完善的一样;例如,像人类眼睛那样奇妙的器官亦复如此。我们都知道赫姆霍尔兹(Helmholtz)——关于这个问题的欧洲最高权威——对人类眼睛说了些什么;他说,如果一位光学仪器商卖给他的仪器粗制滥造得像人类眼睛那样,他大概会认为自己有充分的正当理由去退货。 (480)
我们现在已经看到从一个简单斑点到形似“球与穴”的奇妙装饰物之间可以追踪出一个完整的系列。古尔德先生给过我一些这种羽毛,盛情可感,关于这个级进的完整性,他完全同意我的意见。同一只鸟的羽毛所显示的发展阶段显然完全没有必要向我们表明这个物种的绝灭祖先在发展过程中所经历的步骤;但是,它们大概向我们提供了有关这个实际步骤的线索,至少它们证明了渐次的级进是可能的。如果没有忘记雄锦雉如何小心翼翼地在雌鸟之前夸示其羽衣,而且如果没有忘记前此所举的许多事实,证明雌鸟会偏爱更有魅力的雄鸟,那么凡是承认性选择在任何场合中都会发生作用的人,就不会否认一个简单的暗黄褐色斑点通过相邻的两个斑点的接近和变异,再加上颜色的稍微变深,就可以变为一种所谓的椭圆装饰物。曾把这等椭圆装饰物给许多人看过,他们莫不承认它们是美丽的,有的人甚至认为它们比形似“球与穴”的眼斑更美丽。由于次级羽毛通过性选择而变长了,又由于椭圆装饰物的直径加大了,因此它们的颜色显然变得较不鲜明了;于是,势必通过样式和色调的改进而获得羽衣的装饰性;这个过程继续进行不已,直到最后发展为奇妙的形似“球与穴”的眼斑为止。这样我们就能理解——在我看来用别的方法都不能理解——锦雉翼羽装饰物的现在状态及其起源。
根据级进原理所提供的说明——根据我们所知道的变异法则——根据我们许多家养鸟类所发生过的变化——最后,根据幼鸟未成熟的羽衣性状(对此我们以后将会看得更清楚)——我们有时能够以某种程度的自信来示明雄鸟获得其鲜艳羽衣以及各式各样的装饰物所经历的大致步骤;但在许多场合中我们还是完全处于黑暗之中的。古尔德先生若干年前曾向我指明,有一种名叫白尾梢蜂鸟(Urosticte benjamini)的,以雌雄二者之间的奇特差异而著称。其雄者除了有一个华丽的新月形颈饰外,还有黑绿色尾羽,其中四根中央尾羽的尖端为白色;其雌者和大多数亲缘相近的物种一样,其每侧三根外尾羽的尖端为白色,因此这种白色尖端的尾羽,雄者有四根是在中央而雌者有六根是在外侧。使这种情况更加奇妙的是,尽管许多蜂鸟种类的雌雄二者的尾羽颜色有显著差异,但除了白尾梢蜂鸟属(Urosticte)以外,古尔德先生再也不知道有任何物种的雄者具有四根白色尖端的中央尾羽。
阿盖尔(Argyll)的公爵评论这一情况时竟完全忽略了性选择,并且问道,“对这等特殊的变种,自然选择法则能够给予什么解释呢?”他的回答是,“什么都解释不了”; (481) 我完全同意他。但这种看法能够令人信服地用于性选择吗?鉴于蜂鸟类的尾羽有如此多方面的差异,为什么那四根中央尾羽不应该单在这一个物种中发生变异从而获得其白色的尖端呢?这等变异可能是逐渐的或者是突发的,像最近所举出的波哥大附近蜂鸟类的例子就是如此,这个例子表明“中央尾羽尖端呈艳绿色者”仅为某些个体。我注意到白尾梢蜂鸟属的雌者的那四根黑色中央尾羽,其外侧两根有极细小的或残迹的白尖;因此我们在这里便有了关于这个物种羽衣的某种变化迹象。如果我们承认雄者的中央尾羽有变白的可能性,那么关于这等变异系出于性选择,就毫不足怪了。这种白色羽尖以及白色小耳簇毛,正如阿盖尔的公爵所承认的,肯定会增添雄者的美貌;而白色显然是其他鸟类所欣赏的颜色,这从雪白的雄性铃鸟的情况可以推论出来。赫伦爵士所做的叙述不应忘记,他说,如果禁止他的雌孔雀同雄斑孔雀接近,那么前者就不同其他任何雄孔雀交配,因而在那个季节就没有后代产生出来。白尾梢蜂鸟属的尾羽异变乃是专门为了装饰而受到选择,并不是奇怪的事。因为该科中紧挨着的下一个属就是由于它的华丽尾羽而取得了辉尾蜂鸟属(Metallura)这个名称。此外,我们还有良好的证据可以证明蜂鸟特别尽力地夸示其尾羽;贝尔特先生 (482) 描述了白颈蜂鸟(Florisuga mellivora)的美丽之后说道,“我见过停息在一条树枝上的这种雌鸟,而且有两只雄鸟在雌鸟面前夸示其魅力。一只雄鸟像火箭似的向上飞去,然后突然展开其雪白的尾羽,犹如一只倒置的降落伞,徐徐地降到雌鸟的面前,顺序回转,展示其全身的前前后后。……其展开的白色尾部遮盖了身体的下余部分还有余,当进行这种表演时,其容貌显然是壮丽的。在一只雄鸟降落的同时,另一只雄鸟就会向上突飞,展开其尾羽,徐徐下降。这场表演将以两个表演者的相斗而告终;但究竟是最美丽的还是最勇敢的成为被接受的求婚者,我还弄不清楚。”古尔德先生在描述了白尾梢蜂鸟属的特殊羽衣之后,接着说道:“我本人一点也不怀疑其唯一的目的乃是为了装饰和变异。” (483) 如果承认了这一点,我们就能看出原先以最优美和最新奇的方式来装饰自己的雄者在同其他雄者的竞争中,而非在正常的生存斗争中,将会获得一种优势,而且会留下较大数量的后代来继承其新获得的美貌。
第十五章 鸟类的第二性征(续二)
关于为何某些物种只有雄者的色彩是鲜明的而另外一些物种的雌雄二者的色彩都是明亮的这一问题的讨论——限于性别的遗传性,适用于各种构造以及色彩鲜明的羽衣——筑巢同颜色的关系——冬季期间婚羽的消失
我们在本章所探讨的是,为什么许多鸟类的雌者没有获得和雄者一样的装饰;另一方面,为什么其他许多鸟类的雌雄二者都有一样的或几乎一样的装饰?在下一章,我们将探讨有关雌鸟的色彩比雄鸟的更为显著的少数事例。
在我的《物种起源》一书中, (484) 我曾简略地提到过雌孔雀如果具有雄孔雀那样的长尾,在孵卵时大概不会方便,而且雌雷鸟如果具有雄雷鸟那样的显著黑色,在孵卵时大概会招致危险;结果,通过自然选择这些性状从雄者向雌性后代的传递就会受到抑制。我原来认为这种情况还可能在少数事例中出现:但对我所能收集到的全部事实加以深思熟虑之后,我现在倾向于相信,如果雌雄二者有所差异,一般说来其连续变异的传递一开始就只限于首先出现这种变异的那一性别。自从我发表了这个见解以来,华莱士先生在他写的一些很有趣的论文中探讨了这个性别色彩的问题, (485) 他相信几乎在所有场合中,那些连续变异在最初都倾向于相等地传递给雌雄二者,只不过雌鸟通过自然选择避免了获得雄鸟的显著颜色,不然的话,雌鸟在孵卵期间就会因此招来危险。
为了说明这个见解,需要对一个难题进行冗长的讨论,即,最初由雌雄二者所承继的一种性状,此后是否能够通过自然选择只限于向某一性别传递。我们必须记住,正如在最先讨论性选择的第八章中所表明的,那些只限于在某一性别发育的性状在另一性别总是潜伏的。下面这个假想的例证将会最好地帮助我们去理解这个问题的难点:我们假设有一位鸟类玩赏家希望育出一个鸽品种,仅使这个品种的雄鸽具有淡蓝色而让雌鸽保持其原有的石板青色。由于鸽的各种性状通常都是相等地传递给雌雄二者,因此这位玩赏家就必须试着把后面这种遗传形式转变为限于性别的传递。他所能做的全部事情无非是百折不挠地把每只稍微具有淡蓝色的雄鸽选择下来;如果长期坚持进行这种选择,而且,如果这种淡蓝色变异得到强烈的遗传或常常重现,那么这种选择过程的自然结果大概可以使他的整个鸽群都具有一种较淡的蓝色。但我们这位玩赏家将被迫一代又一代地使其淡蓝色雄鸽同石板青色雌鸽进行交配,因为它希望使后者仍保持石板青色。其结果一般是育出许多杂色的杂种,而更可能是淡蓝色迅速而彻底地消失;这些因为原始的石板青色将以优势的力量传递下来。然而,假设在连续的每一个世代中都产生了一些淡蓝色的雄鸽和石板青色的雌鸽,并且总是彼此杂交,那么石板青色的雌鸽,如果我可以用下面方式来表达的话,就会在雌鸽们血管里有大量的蓝色血液,因为它们的父代、祖代等全都是蓝色的鸟。在这种情况下可以想象得到(尽管我不知道有什么显著的事实可以说明这是可能的),石板青色雌鸽所获得的淡蓝色的潜伏倾向是如此强烈,以致不会破坏其雄性后代的淡蓝色,而其雌性后代仍会承继石板青色。果真如此,那么育成一个雌雄二者的色彩永不相同的品种这个所要求的目的就会达到。
上述场合中所要求的性状,即淡蓝色这个性状,虽然在雌鸽方面处于潜伏状态,但它的存在还是极端重要的,毋宁说是必不可少的,这样,雄鸽的淡蓝色就不致恶变,下述情况将使这一点得到最好的说明:铜色雉(soemmerring's pheasant)雄者的尾羽长达37英寸,而雌者尾羽的长度仅为八英寸;普通雄雉的尾羽长约20英寸,而其雌者尾羽的长度为12英寸。那么,如果具有短尾的雌铜色雉同普通雄雉进行杂交,毫无疑问其雄性杂种后代的尾羽将会比普通雉纯种后代的尾羽长得多。另一方面,普通雌雉的尾羽比雌铜色雉的尾羽长得多,如果前者同雄铜色雉进行杂交,那么其雄性杂种后代的尾羽就会比铜色雉纯种后代的尾羽短得多。 (486)
我们的玩赏家为了育成一个这样的新品种:其雄鸽为淡蓝色,其雌鸽保持原色不变,他必须在许多世代中连续对雄鸽进行选择;而且颜色变淡每一阶段都必在雄者方面固定下来,并且使其在雌者方面潜伏下来。这将是一项极其困难的工作,从未有人试过,但却是可能实现的。其主要障碍大概是,由于必须同石板青色雌鸽反复进行杂交,而后者一开始就没有产生淡蓝色后代的任何潜伏倾向,则淡蓝的色调将在早期内完全消失。
另一方面,如果有一两只雄鸽出现了非常轻微的淡蓝色变异,而且这种变异从一开始就只限于传递给雄性一方,那么要育成一个预期类型的新品种的工作大概就会容易了,因为只要简单地选择这种雄鸽并使之同普通雌鸽交配就可以了。实际上有一个类似情况曾经出现过,因为在比利时有些鸽的品种只有其雄者才具有黑色条纹。 (487) 再者,特格梅尔先生最近指出, (488) 龙鸽(dragons)产生了不少银色的鸽,这等鸽几乎都是雌的;他自己就育成了10只这样的雌鸽。反之,如果育成一个银色的雄鸽,那就是一件很特殊的事情了;因此如果愿意的话,没有比育成一个龙鸽品种——其雄者为蓝色、其雌者为银色——更为容易的了。这种倾向的确是非常强烈的,因而当特格梅尔先生最后获得了一只银色雄鸽并使它同一只银色雌鸽进行交配时,他期望获得雌雄二者都是这等色彩的一个品种,但他失望了,因为雄幼鸽复现其祖代的蓝色,只有雌幼鸽呈银色。毫无疑问,只要有耐心,用偶尔出现的银色雄鸽同银色雌鸽进行交配,这样育成的雄鸽,其返祖倾向还是可以排除的,于是雌雄二者的色彩便是同样的了;埃斯奎兰特(Esquilant)先生在银色浮羽鸽的场合中成功地实现了这一过程。
至于家鸡,其传递只限于雄性的那些颜色变异是经常发生的。当这种遗传形式居于优势时,大概常常会发生下述情况:即,有些连续变异也会传递给雌鸡,于是这等雌鸡就会同雄鸡稍微相似,如某些品种实际上所发生的情况那样。此外,大多数的但并非全部的连续变异可能传递给雌雄二者,于是雌者就会同雄者密切相似。雄突胸鸽(pouter)比雌突胸鸽的嗉囊大不了多少并且雄信鸽比雌信鸽的垂肉大不了多少,其原因无可怀疑地正在于此;因为玩赏家们所选择的某一性并不比另一性为多,而且没有任何意图使这等性状在雄者方面比在雌者方面表现得更为强烈,尽管这两个品种的情况正是如此。
如果期望育成一个只是雌者具有某种新色彩的品种,那么就必须遵循同样的过程,而且会遇到同样的困难。
最后,我们的玩赏家也许希望育成一个这样的品种:其雌雄二者彼此不同,而且和亲种也不相同。在这种场合中要想获得成功是极其困难的,如果那些连续变异一开始在雌雄双方都有性别限制,那么大概就不会有什么困难。我们在家鸡的场合中可以看到这种情况;例如条斑汉堡鸡的雌雄二者彼此大不相同,而且同原始祖先原鸡(Gallus bankiva)的雌雄二者也不相同;于是靠着连续选择,其雌雄二者在其优秀标准上都保持了稳定;除非雌雄二者的独特性状在传递上有所限制的话,那么这种连续选择就几乎是不可能的了。
西班牙鸡提供了一个更加奇妙的事例:其雄者有一个巨大的肉冠,这一性状系通过连续变异的积累而获得的,但有些连续变异似乎已传递给了雌者;因为它的肉冠比亲种雌者的肉冠要大许多倍。但雌者的肉冠和雄者的肉冠在某一点上有所不同,因为雌者的肉冠容易垂下;最近玩赏家决定使它永远如此,并迅速获得成功。现在肉冠下垂这一性状在传递上必定有性别限制,否则它就会制止雄者肉冠的完全直立,而每个玩赏家对此都感到厌恶。另一方面,其雄者肉冠的直立也必定同样是有性别限制的一种性状,否则它也会制止雌者肉冠的下垂。
根据以上说明,我们知道即使有几乎不受限制的可以自由支配的时间,通过选择把一种传递方式改变为另一种方式,也是极端困难而复杂的、也许是不可能实现的过程。因此,无论在什么场合中,若无显著的证据,我不愿承认自然物种会完成这一过程。另一方面,凭借一开始在传递上就有性别限制的连续变异,要使一只雄鸟在颜色或其他任何性状方面和雌鸟大不相同,就不会有丝毫困难了;同时雌鸟仍保持不变,或稍有改变,或为了保护自己而发生特殊改变。
由于鲜明的色彩有助于雄鸟同其他雄鸟进行竞争,因此这等色彩无论是否专门向同一性别传递,都会受到选择,结果,可以期望雌鸟往往会程度不同地分享雄鸟的鲜明色彩;很多物种都发生过这种情形。如果所有连续变异都相等地传递给雌雄二者,则雌者和雄者就没有区别;许多鸟类所发生的情况正是如此。然而,如果暗淡的色彩对于雌鸟在孵卵期间的安全是高度重要的话,例如许多地栖鸟类(ground birds)的情况,那么那些色彩变得鲜明的雌鸟,或那些通过遗传从雄鸟方面继承了任何显著鲜明色彩的雌鸟,迟早都不免于毁灭。但是,雄鸟把自身的鲜明色彩传递给其雌性后代已经历了无限长的时期,要想消除这种倾向,就必须通过遗传方式的改变;这,正如我们前面的例证所表明的那样,大概是极端难的。假设同等地向雌雄二者传递的方式占主导地位,那么那些色彩比较鲜明的雌鸟长期不断遭到毁灭的更加可能的结果将是色彩鲜明的雄鸟的减少或覆灭,这是由于雄鸟,同色彩比较暗淡的雌鸟不断进行杂交的缘故。要一一列举所有其他可能的结果会令人感到厌烦;但我愿提醒读者注意一点,如果雌鸟发生了有性别限制的鲜明色彩的变异,即使这等变异并没有给它们带来丝毫损害,因而也没有使它们覆灭,可是它们仍然不会因此受益或受到选择,这是因为雄鸟通常会接受任何一只雌鸟,并不选择更有魅力的个体;结果这等变异就容易消失,而对于这个族的性状并不会发生多大影响;这将有助于我们解释雌鸟的色彩为何普遍都比雄鸟的色彩暗淡。
在第八章曾举出过一些事例,表明诸变异在不同年龄出现者,亦于后代的相应年龄遗传之,在此还可以对此等事例做许多补充。它们还表明在生命晚期发生的变异通常都是传递给最先出现这种变异的那一性别;而在生命早期发生的那些变异则倾向于传递给雌雄双方;并非对受得性别限制的一切传递事例都能这样给予解释。上述事例进一步表明,如果一只雄鸟在幼小时发生了色彩更加鲜明的变异,那么这等变异在达到生殖年龄之前是没有用处的,而到了生殖年龄,雄鸟之间就会发生竞争。但是,生活于地面的鸟类通常都需要暗淡的色彩作为保护,在这种场合中,鲜明的色彩对于没有经验的幼小雄鸟比对于成年雄鸟更加危险得多。因此,那些发生鲜明色彩变异的幼小雄鸟就会受到大量损害并通过自然选择而覆灭;另一方面,那些在接近成熟期发生这样变异的雄鸟,尽管处于更多的危险之中,但仍会活下去,而且由于通过性选择得到了利益,它们的种类就会繁衍起来。由于在变异的时期和传递的方式之间往往存在着一种相互联系,因此,倘若具有鲜明色彩的幼小雄鸟遭到了毁灭而且具有鲜明色彩的成熟雄鸟在求偶方面获得了成功,那么就只有雄鸟会获得鲜明色彩并把这种性状专门传递给其雄性后代。但我丝毫没有坚持认为年龄对传递方式的影响乃是许多鸟类雌雄二者在鲜明颜色方面存在着巨大差异的唯一原因。
当鸟类的雌雄二者在颜色上有所差异时,令人感到有趣的是,决定这等差异究竟是因为只有雄鸟在性选择作用下发生了变异而雌鸟保持不变呢,还是因为雄鸟只是部分地和间接地发生了这种变化呢;或者,是否因为雌鸟在自然选择作用下为了保护自己而发生了特殊变异呢。因此,我将用一定的篇幅来充分讨论这个问题,从其内在的重要性来看,我的讨论也许过于充分,因为只有这样才可以便利地对各个并行的奇妙问题进行考察。
在我们讨论色彩问题、尤其是有关华莱士先生的一些结论之前,用同样观点先讨论一下某些别的性差异,也许是有用处的。以前在德国有一个家鸡品种 (489) ,其雌者有足距,是很好的产卵鸡,但它们的足距对于鸡窝的损坏如此之大,以致不能允许它们去孵自己的卵。因此,有一个时期我以为野生鹑鸡类(Gallinaceae)雌者足距的发展大概通过自然选择而受到了抑制,这恐怕也是由于其足距给鸡窝造成了损坏的缘故。从下述事实看来,这一点就似乎完全可能了,即,由于翼距在孵卵期间不会给鸟窝造成损坏,因此雌鸟翼距往往和雄鸟翼距一样地发达;尽管在不少场合中雄鸟翼距略大。如果雄鸟具有足距,雌鸟几乎总是表现有足距的残迹,——这个残迹物像在原鸡属(Gallus)中那样,有时仅仅是由一个鳞片构成的。因此,有人会争辩说,雌鸡原来都具有十分发达的足距,不过后来由于不使用或自然选择作用而消失了。但是,如果承认这个观点,就得把它引申到其他无数事例,而这意味着现存的有距物种的雌性祖先曾一度受到一种有害附器的拖累。
在少数某些属和物种中,诸如山鹑属(Galloperdix)、团扇雉(Acomus)以及爪哇绿孔雀(Pavo muticus),其雌者和雄者一样都有十分发达的足距。根据这一事实我们是否可以推论说雌鸟们所造的巢不同于其近亲物种所造的巢,这种巢不容易受到它们的足距的损坏,所以它们的足距并没有因此消失呢?或者我们是否可以假设这几个物种的雌者特别需要足距以保卫它们自己呢?一个更为可能的结论是,雌鸟足距的存在和缺如都是占有主导地位的不同遗传规律所造成的结果,而同自然选择无关。关于足距以残迹状态出现的许多雌鸟,我们可以断定,通过连续变异,雄鸟的足距发达了,其中少数连续变异是在雄鸟的生命很早时期发生的,结果传递给了雌鸟。另外有些雌鸟的足距也是充分发达的,关于这非常罕见的事例,我们可以断定全部的连续变异都传递给了雌鸟;而且它们逐渐获得了并遗传了不弄坏其鸟巢的习性。
雌雄二者的发音器官和经过种种改变以便发音的羽毛,以及运用这等器官的固有本能,往往彼此有所差异,但有时是彼此相同的。能否用下述原因来说明这等差异呢?即,雄鸟获得了这些器官和本能,而雌鸟并不遗传它们,以免引起猛禽或猛兽的注意而处于危险之中。每当我们想起大量的鸟类在春天无忧无虑地以其鸣声给乡村带来欢乐时,就觉得这个原因似乎是不可能的。 (490) 较为可靠的一个结论是,由于声乐器官和器乐器官的特别用途在于雄鸟的求偶,所以这些器官是通过性选择并在雄性中不断使用而发展起来的——其连续变异以及使用的效果从一开始就或多或少地只限于传递给雄性后代。
还有许多相似的事例可以引证;诸如雄鸟头上的羽饰一般比雌鸟的长,有时二者的长度相等,偶尔雌鸟头上缺少羽饰,——这几种情形竟在鸟的同一类群中发生。用下述说法很难解释雌雄二者之间的这等差异,即认为雌鸟的冠毛小于雄鸟的冠毛对雌鸟来说是有利的,因而通过自然选择它的冠毛缩小了或完全受到抑制了。但是,我将举一个有关尾羽长度的更好事例。如果雌孔雀具有雄孔雀那样的长尾羽,在其孵卵期间以及伴随幼者期间不仅使它不方便而且会给它招来危险。因此,它的尾羽的发育通过自然选择而受到抑制,在演绎上并非一点也不可能。各种雌雉在其开放的鸟巢中所面临的危险显然同雌孔雀所面临的一样大,但前者仍具有相当长的尾羽。琴鸟(Menura superba)的雌者和雄者一样,也具有长长的尾羽,它们还修造了一种有圆顶的鸟巢,这种鸟巢对于如此大型的一种鸟来说是一种重大的反常现象。雌琴鸟在孵卵期间如何处理它的长尾羽曾使博物学家们感到疑惑;但是,现在已经知道,“入巢时头部在先,然后转过身来,它的尾羽有时弯在背上,但更常见的是弯在身边。一到这样的时候,雌鸟的尾羽就变得十分歪斜,这是一个忍受痛苦的标志,表明这种鸟的孵卵期间很长”。 (491) 一种澳洲翠鸟(Tanysiptera sylvia)的雌雄二者都具有大大变长的中间尾羽,其雌者造巢于穴中;我听夏普先生说,这等尾羽在孵卵期间变得非常弯曲。
在后面这两种场合中,尾羽的巨大长度在某种程度上一定对雌鸟是不方便的;这两个物种的雌鸟的尾羽或多或少地短于雄鸟的尾羽,因此有人也许会争辩说,雌鸟尾羽的充分发展通过自然选择受到了阻止。但是,如果雌孔雀尾羽的发展只是在它变得不方便或危险性增大时才被阻止,那么它保留的尾羽,大概要比它实际有的尾羽长得多;因为,按尾羽和体型大小的比例来说,雌孔雀的尾羽并没有像许多雌雉的尾羽那样长,也不长于雌火鸡的尾羽。还必须记住,按照这个观点来说,一旦雌孔雀的尾羽长度达到招致危险的地步,因而它的发展受到抑制时,它大概就会不断地对其雄性后代发生作用,这样就要阻止雄孔雀获得其现在那样华丽的长尾羽。因此,我们可以推断说,雄孔雀尾羽之长和雌孔雀尾羽之短都是雄孔雀必然发生的变异的结果,而这些变异从一开始就只传递给其雄性后代。
有关雉的各个物种的尾羽长度,也会引导我们作出几乎相似的结论。蓝马鸡(Crossoptilon auritum)雌雄二者的尾羽长度是相等的,即16英寸或17英寸长;雄普通雉的尾羽长约20英寸,雌普通雉的尾羽长为12英寸;雄铜色雉的尾羽长为37英寸,而雌铜色雉的尾羽长仅8英寸;最后,雄中国雉(Reeve's pheasant)尾羽的实际长度有时竟达72英寸,而雌中国雉的尾羽长度仅为16英寸。这样,姑置若干物种的雄鸟尾羽不论,雌鸟尾羽的长度也有巨大差异;在我看来,这种差异可以用遗传法则——即,连续变异从一开始就或多或少紧密地限于传递给雄性一方——得到解释;这比用自然选择的作用——即,其结果是由尾羽长度或多或少地有害于这几个亲缘相近的物种的雌者所引起的——来进行解释更加合理得多。
我们现在可以考察一下华莱士先生有关鸟类的性别色彩问题的论点。他认为雄鸟原先通过性选择所获得的鲜明色彩,在所有的或几乎所有的场合中,都传递给了雌鸟,除非这种传递通过自然选择受到了抑制。我在这里可以提醒读者注意,同这个观点相矛盾的各种事实已在有关爬行类、两栖类、鱼类和鳞翅类的章节中列举过了。如我们将在下一章看到的,华莱士先生将其信念主要建立在而并非专门建立在下列陈述的基础之上, (492) 即,当雌雄二者都具有很显著的色彩时,其鸟巢就具有隐蔽对孵卵之鸟加以隐蔽的性质;但是,如果雌雄二者的色彩存在着显著的对比,雄鸟色彩鲜艳而雌鸟色彩暗淡,那么它们的鸟巢就是开放的,孵卵之鸟一望得见。这种巧合,就其所表现的来说,似乎肯定有利于下述信念,即,在开放鸟巢中孵卵的雌鸟为了保护自己发生了特殊的变异;但我们马上就会知道还有另一种更为合理的解释,即,色彩显著的雌鸟比色彩暗淡的雌鸟获得营造圆顶鸟巢之本能者更加常见。华莱士先生承认,像可以预料到的那样,关于他的那两条规律,有一些例外,但问题在于这些例外是否没有多到可以使这两条规律归于无效的严重程度。
阿盖尔的公爵 (493) 认为一种圆顶大鸟巢比开放的小鸟巢容易为其敌害所见,尤其容易为攀行树间的食肉兽所见,这是非常正确的看法。我们一定不要忘记,营造开放鸟巢的许多鸟类的雄者也会孵卵而且帮助雌者喂养幼鸟:例如美国最华丽的一种鸟(Pyranga aestiva)的情况就是如此, (494) 这种雄鸟呈朱红色,雌鸟则为淡褐绿色。那么鲜艳的色彩对于在开放鸟巢里孵卵的鸟如果是极端危险的话,则雄鸟在这等场合中就会大大受害。然而,为了击败其雄性对手,雄鸟具有鲜艳色彩是最重要的,这是以补偿某种附加的危险而有余。
华莱士先生承认,短尾属(Dicurus)、黄鹂属(Orioles)以及八色鸫科(Pittidae)的雌者都具有显著的色彩,但它们却造开放的巢;然而他极力主张,第一类群的鸟高度勇猛善斗,能够保卫自己;第二类群的鸟则极端谨慎地把其开放的鸟巢隐蔽起来,但实际情况并非永远如此; (495) 至于第三类群的鸟,其雌者的鲜明色彩主要在身体的底面。除这些事例外,鸽子的色彩有时也是鲜明的,而且几乎总是显著的,众所熟知,它容易遭受猛禽的攻击,因而这对华莱士的规律又提供了一个突出的例外,因为鸽子所造的巢几乎总是开放的和外露的。另一大科,即蜂鸟科,其所有物种都营造开放的鸟巢,可是有些最华丽的物种,其雌雄二者的色彩是彼此相似的;从大多数物种来看,尽管雌鸟的华丽色彩不及雄鸟,但它们的色彩仍然是鲜明的。认为一切色彩鲜明的雌蜂鸟由于具有绿的色泽就可逃避察觉也是不对的,因为有些雌蜂鸟的体部上表面呈现红色、蓝色以及其他颜色。 (496)
关于鸟类在穴中做巢或营造圆顶鸟巢,华莱士先生认为,这样除了有荫蔽的好处以外,还有其他好处,诸如可以避雨和避酷热,而且在热带可以防止太阳的照射; (497) 据此,就不能有效地反对他的观点,他认为,许多鸟类的雌雄二者虽然都呈暗淡的色彩,却造荫蔽的巢。 (498) 以印度和非洲的雌犀鸟(Bucerus)为例,它在孵卵期间非常细心地把自己保护起来,它在穴中孵卵,用自己的排泄物封闭穴口,只留一个小孔,以便雄鸟给它进食;于是它在整个孵卵期间成了一个被禁闭的囚徒 (499) ;然而雌犀鸟的色彩并不比那些营造开放鸟巢的其他许多同等大小的鸟类的色彩更为显著。如华莱士先生自己所承认的,同他的观点相矛盾的还有一个更严重的情况,即,某些少数类群的雄鸟色彩鲜艳而雌鸟色彩暗淡,但后者仍然在圆顶鸟巢中孵卵。澳大利亚的鹤类(Grallinae)和同一地方的“超等歌手” (500) (Maluridae)、太阳鸟类(Nectariniae)以及某些澳大利亚吸蜜鸟科(Meliphagidae)的情况都是如此。 (501)
倘若我们观察一下英国的鸟类,就会知道在雌鸟的色彩和它所营造的鸟巢的性质之间并没有密切而普遍的关联。大约有40种英国的鸟类(那些能保卫自己的大型鸟类除外)把巢做在河岸的穴、岩石的穴或树穴中,或者营造有圆顶的巢。我们若将金丝雀、红腹灰雀或乌鸫的雌者色彩作为显著程度的一个标准,因为这等色彩对孵卵的雌鸟并没有高度的危险性,那么在上述40种鸟类中只有12种雌鸟色彩的显著程度可以认为达到危险的地步,其余28种的色彩可视为不显著。 (502) 在同一属中,雌雄二者色彩十分显著的差异和其所造的鸟巢的性质之间并无任何密切关系。例如,雄家麻雀(Passer domesticus)和雌家麻雀差异很大,而雄树麻雀(P.montanus)和雌树麻雀几乎没有差异,可是二者都营造十分荫蔽的巢。普通食虫的鹟(Muscicapa grisola)的雌雄二者几乎没有区别,而斑色鹟(M.luctuosa)的雌雄二者差异颇大,可是这两个物种都做巢于穴中,即将其鸟巢荫蔽起来。雌乌鸫(Turdus merula)和雄乌鸫差异很大,雌环纹黑鸫(T.torquatus)和雄环纹黑鸫差异较小,而普通雌鸫(T.musicus)和雄鸫几乎完全没有差异;可是它们做的巢全都是开放的。反之,同上述鸟类亲缘关系不很远的河鸟(Cinclus aquaticus)则营造一种圆顶鸟巢,其雌雄二者之间的差异程度却和环形黑鸫的情形一样。黑松鸡和红松鸡(Tetrao tetrix和T.scoticus)在同等十分荫蔽的地点营造开放的巢,但其中一个物种的雌雄二者差异很大,而另一物种的雌雄二者差异却很小。
尽管有上述同华莱士先生的论点相矛盾的事实,但读了他那篇优秀的论文之后,我还是不能怀疑:从全世界的鸟类来看,确有大多数物种,其雌鸟色彩显著者(在这种场合中,除很少例外,其雄鸟的色彩也同等显著),为了保护自己而营造荫蔽的巢。华莱士先生列举了长长的一系列类群合乎这一规律; (503) 但在这里,举出一些比较驰名的类群,如翠鸟,鵎鵼(toucans),咬鹃(trogons),须鴷科(Capitonidae),蕉鹃(Musophaga),啄木鸟以及鹦鹉,就足够了。华莱士先生相信,在这些类群中,由于雄鸟通过性选择逐渐获得了其鲜艳色彩,这些鲜艳色彩遂传递给雌鸟,因为雌鸟们的造巢方式已经使它们得到了保护,所以没有因自然选择而消灭。按照这个观点,它们现在的造巢方式的获得先于它们现在的色彩的获得。但是,在我看来,远远更加可能的是,在大多数场合中,由于雌鸟分享了雄鸟的色彩而逐渐变得越来越鲜艳,所以导致它们逐渐改变了其本能(假定它们原来造的是开放的巢),而营造有圆顶的或荫蔽的巢,以寻求保护。关于美国北部和南部的同一物种的鸟巢差异,奥杜邦做过报道, (504) 譬如说,凡是读过这篇报道文章的人,就不会感到任何重大困难去承认:鸟类,或通过其习性的改变(按其严格的字义来讲),或通过本能的所谓自发变异的自然选择,大概会容易地被引导去改变其造巢方式。
关于雌性鸟类的鲜明色彩和其造巢方式之间的关系,这种观察方法,就其所能适用的范围来看,可以从撒哈拉沙漠所发生的某些事例得到某种支持。在那里,就像在其他大多数沙漠那样,各种鸟类以及其他许多动物的色彩以可惊的方式同周围地面的色泽相适应。尽管如此,正如特里斯特拉姆(Tristram)牧师告诉我的,关于这一规律还有一些奇妙的例外;如矶鸫(Monticola cyanea)的雄者因其鲜明的蓝色而惹人注目,其雌者因其褐色和白色相杂的羽衣几乎同等地惹人注目;白尾岩鸫(Dromolaea)有两个物种,其雌雄二者都有一种黑色光泽;因此,这三个物种因其色彩而远远不能得到保护,但它们还是能够生存下来了,这是因为它们已经获得了在洞穴中或岩石裂缝中躲避危险的习性。
关于雌鸟既有显著色彩又造荫蔽鸟巢的上述类群,不需要假设每个物种的造巢本能分别地发生过特殊的改变;只不过是每个类群的早期祖先逐渐被引导去建造圆顶的或荫蔽的鸟巢,而且,后来又把这种本能以及鲜明的色彩一起传递给其变异了的后代。就所能相信的来说,下述结论是有趣的,即,性选择以及雌雄二者相等的或几乎相等的遗传性,曾经间接地决定了整个鸟类的造巢方式。
有些类群的雌鸟由于在孵卵期间受到了圆顶鸟巢的保护,所以其鲜明色彩未曾通过自然选择而被消除,按照华莱士先生的意见,即使在这种场合中,雄鸟和雌鸟往往还有轻微的差异,有时有很大程度的差异。这是一个有重要意义的事实,因为对于这等色彩的差异一定要以雄者的某些差异从一开始就限于只向同一性别传递来进行解释;必须做如此解释的原因在于,简直不能认为这等差异可以用于保护雌者;如果这等差异轻微就尤其如此。例如,在三宝鸟这一华丽类群中,所有物种都在穴中造巢;古尔德先生提供了其25个物种的雌雄二者的绘图, (505) 其中除有一个物种表现了局部的例外,全部物种的雌雄二者的色彩有时差异轻微,有时差异显著——尽管这等雌鸟也同样好看,但雄鸟总是比雌鸟更漂亮。翠鸟的所有物种都在穴中造巢,而且大多数物种的雌雄二者的色彩都同等鲜艳,因此华莱士先生的规律在此颇为适用;但在某些澳大利亚物种中,雌鸟的色彩比雄鸟的稍欠鲜艳;还有一个色彩华丽的物种,其雌雄二者的差异如此之大,以致最初一看会把它们当做不同的物种。 (506) 对这个类群进行过特别研究的夏普先生给我看过一些美国的物种鱼狗(Ceryle),其雄者胸部有黑色带斑。此外,还有一种翠鸟(Carcineutes),其雌雄二者差异显著:雄鸟上表面呈暗蓝色,具有黑色带斑,下表面局部呈淡黄褐色,而头部甚红,雌鸟上表面呈红褐色,具有黑色带斑,下表面呈白色,并有黑色斑纹。有一个有趣的事实,表明雌雄色彩的同一独特样式往往构成了亲缘相近的诸类型的共同特征,在鴗属(Dacelo)的三个物种中,雄鸟不同于雌鸟之处仅在于前者的尾羽呈暗蓝色并具黑色带斑,而雌鸟的尾羽则为褐色并具微黑的条纹;因此,在这里雌雄二者尾羽色彩的差异恰如一种翠鸟(Carcineutes)雌雄二者整个上表面色彩的差异一样。
关于同样在穴中造巢的鹦鹉类,我们发现有相似的事例:大多数物种的雌雄二者的色彩都是鲜艳的,而且难以区分,但也有不少的物种,其雄色的色彩比雌者的更加鲜艳,甚至和雌者的很不相同。例如,除了其他强烈显著的差异之外,雄性澳洲猩猩鹦鹉(Aprosmictus scapulatus)的整个下表面均呈猩红色,而雌者的喉部和胸部则为绿色,带有红的色调。另一种鹦鹉(Euphema splendida)也有类似的差异,其雌者的脸部和翼覆羽的蓝色都比雄者的为淡。 (507) 在营造荫蔽巢的山雀(Paridae)这一科中,英国普通蓝山雀(Parus caeruleus)的雌者在色彩上远不及雄者的鲜明。而印度的华丽的苏丹黄山雀(Sultan yellow tit)其雌雄二者的差异还要大。 (508)
此外,在啄木鸟这个大类群 (509) 中,其雌雄二者一般差不多是相像的,但是,大型绿啄木鸟(Megapicus validus)雄者的头部、颈部和胸部呈艳红色,而雌者的所有这等部分则为淡褐色。由于几种啄木鸟的雄者头部呈鲜明的艳红色而雌者头部的色彩是平淡的,所以在我看来,如果雌者具有雄者头部的那种色彩,只要它把头伸出鸟巢洞口之外,就可能有惹起注目的危险,因而按照华莱士先生的信念,雌者头上的这种色彩被消除了。马勒布(Malherbe)关于印度啄木鸟(Indopicus carlotta)的叙述加强了这个观点;他说,幼小的雌啄木鸟和幼小的雄啄木鸟一样它们的头部稍呈艳红色,但在成熟的雌啄木鸟,头部这种色彩就消失了,相反,在成熟的雄啄木鸟头部这种色彩却加强了。尽管如此,下述考察还是使这个观点显得极其可疑:雄啄木鸟在孵卵期间也承担了相当的责任, (510) 这就使得它们几乎相等地暴露于危险之中;有许多物种,其雌雄二者的头部具有同等鲜明的艳红 色;另外有些物种,其雌雄二者的猩红色差异程度如此轻微,以致几乎不能在招致危险方面形成任何可以觉察的差别;最后,雌雄二者头部的色彩也往往在其他方面有轻微的差异。
在雌雄二者按照一般规律彼此相似的那些类群中,凡是雌雄之间在色彩上有轻微的和级进的差异者,迄今所举的事例,都与营造圆顶鸟巢或荫蔽鸟巢的物种有关系。但是,有些类群的雌雄二者按照一般规律是彼此相似的,但营造开放的鸟巢,在这等场合中同样可以观察到相似的级进差异。
由于我在前面曾以澳洲鹦鹉为例,因此我愿在这里以澳大利亚鸽类为例, (511) 但不举任何细节。值得特别注意的是,在所有这些场合中,雌雄二者羽衣的轻微差异的一般性质同偶尔出现的较大差异的一般性质是相同的。关于这个事实,那些翠鸟已经提供了良好的例证,其雌雄二者或仅在尾羽或在羽衣的整个上表面都以同样的方式而有所差异,相似的情况也可在鹦鹉类和鸽类中观察到。同一物种的雌雄二者之间在色彩上的一般差异性质也和同一类群的不同物种之间在色彩上的一般差异性质相同。这是因为在雌雄二者通常彼此相似的类群中,如果雄者相当不同于雌者,那么雄者色彩的风格并不见得是全新的。因此我们可以推断说,在同一类群中,雌雄二者彼此相似时的特殊色彩,以及雌雄者二者稍微不同、甚至相当不同时的雄者色彩,在大多数场合中,都是由相同的一般原因所决定的;这个原因就是性选择。
正如已经说过的,雌雄二者之间的色彩差异如果很轻微,这种差异对于保护雌鸟大概就不会有什么作用。然而,假定这等差异有这种作用,那大概就会认为它们是处于过渡状态;但我们没有理由相信许多物种都同时发生变化。因此我们简直无法承认在色彩上和其雄鸟差异很轻微的大量雌鸟为了保护自己现在一齐开始变得色彩暗淡了。即使我们考察多少更为显著的雌雄差异,例如雌欧洲苍头燕雀的头部,——雌红腹灰雀胸部的艳红色彩——雌金翅雀(greenfinch)的绿色——雌金冠鹪鹩(golden-crested wren)的冠羽,是否可能全是为了保护自己而在缓慢的选择过程中变得较不鲜明吗?我不能认为是这样;尤以营造荫蔽鸟巢的那些鸟类的雌雄之间差异轻微者,更不是这样。反之,雌雄之间的色彩差异,不论大小,都可以在相当大的程度上根据连续变异的原理得到说明,即,雄鸟通过性选择所获得的连续变异,从一开始就或多或少地限于只传递给雌性一方。在同一类群的不同物种中,这种传递的限制程度大概也是不同的,凡是研究过遗传法则的任何人对此都不会感到惊奇,因为这等法则是如此复杂,而且由于我们的无知,在我们看来,其作用好像是彷徨不定的。 (512)
就我所能发现的情况来说,在鸟类中只有少数的大类群,其所有物种的雌雄二者都彼此相似,而且都具有鲜明的色彩,但我听斯克莱特先生说,蕉鹃似乎就是如此。我也不相信有任何这样的大类群存在,即其全部物种的雌雄二者在色彩上会有天壤之别:华莱士先生告诉我说,南美洲的伞鸟科(Cotingidae)在这方面提供了一个最好的例子;在其中有些物种,其雄鸟的胸部呈美艳的红色,而雌鸟的胸部也稍微呈现一点红色;其他物种的雌鸟也表现了雄鸟所具有的绿色和其他色彩的痕迹。尽管如此,我们还有一些近似的事例表明,在几个类群中其全部物种的雌雄二者都是密切相似的,或者是不相似的。而这一点,从刚才听说的遗传的彷徨性质来看,乃是一个多少令人奇怪的情况。但是,相同的法则可以广泛地通用于亲缘相近的动物,却不足为奇。家鸡已经产生了大量的品种和亚品种,它们的雌雄二者的羽衣一般都有差异;所以当某些亚品种出现了雌雄二者彼此相似的现象时就会被看做是一种异常的情况。另一方面,家鸽也同样产生了一大批不同的品种和亚品种,除了罕见的例外,它们的雌雄二者都是完全相似的。
因此,鸡属和鸽属的其他物种如果经过家养和变异,那么我们预言由遗传形式所决定的雌雄相似和不相似的同样规律在这两种场合都可适用,并不算轻率。和上述一样,在自然状况下,同样的传递方式一般也通用于相同的整个类群,尽管对这一规律还有一些显著的例外。这样,在同一科里,甚至在同一属里,其雌雄二者的色彩或完全相同,或差异很大。关于同一属的这等事例早已举出过了,诸如麻雀、鹟科食虫鸟、画眉和松鸡的例子。在雉科中,几乎所有物种的雌雄二者都非常不相似,但马鸡(Crossoptilon)的雌雄则完全相似。鹅的一个属,即白雁属(Chloephaga),其两个物种的雄者除体型大小外,在其他方面都无法同雌者区别;而另外两个物种,其雌雄二者如此不相似,以致容易地把它们误作不同的物种。 (513)
只有用遗传规律才能说明下述情况,即,雌者在其生命后期获得了雄者所特有的某些性状,最终会或多或少地同雄者完全相似。“保护”在这里几乎不起作用了。布莱思先生告诉我说,黑头黄鹂(Oriolus melanocephalus)以及某些亲缘相近的物种,其雌者当成熟到足以生殖时,它的羽衣同成熟雄者的羽衣差异很大;但在第二次或第三次换羽后,雌鸟们和雄者的差异仅仅在于其鸟喙有一种淡绿色彩而已。在矮小的鳽属(Ardetta)中,根据这位权威人士的意见,“其雄者在第一次换羽时就获得了其最后羽衣,而雌者在第三或第四次换羽之后才获得其最后羽衣,这时它的羽衣呈现一种中间的色彩,最终乃换成与雄者一样的羽衣”。还有游隼(Falco peregrinus)的雌者也是这样,它获得蓝色羽衣比雄者较慢。斯温赫先生说,有一种黑卷尾(Dicrurus macrocercus),其雄者当几乎还是未离巢的雏鸟时,就换掉柔软的褐色羽衣,而变成均匀的富有光泽的绿黑色了;但是,其雌者却长期在腋羽上保持着白色条纹和斑点;而且历时三年后才呈现雄者那样的均匀黑色。这位杰出的观察家说,中国的雌篦鹭(Platalea)在第二年春天才同第一年的雄者相像,显然,不到第三年春季它不会获得雄者在早得多的年龄所具有的那样成年羽衣。加罗林太平鸟(Bombycilla carolinensis)的雌者和雄者的差异很小,但其翼羽上有一种火红色念珠般的装饰物, (514) 这种附器在雌者身上发育较晚而在雄者生命的很早的时期就发育了。有一种印度的长尾鹦哥(Palaeornis javanicus),其雄者的上喙在最早的幼年期即呈珊瑚红色,但其雌者的上喙,如布莱思先生就笼养的和野生的这种鸟所观察到的,最初是黑色,至少在一岁以后才变为红色,这时雌雄二者在所有方面都彼此相似了。野火鸡雌雄二者的胸部最终都会具有一簇刺毛,但在两岁时,雄野火鸡的这簇刺毛约为四英寸长,而雌野火鸡这簇刺毛,还不十分显露;然而,当雌野火鸡到四岁时,这簇刺毛即长达四至五英寸。 (515)
千万不要把这等事例同下述情况混淆起来,即,有病的或年老的雌者会反常地呈现雄性特征,或者,能育的雌者在幼小时通过变异或某种未知的原因而获得雄者特征。 (516) 但所有这等事例都有一个密切的共同点,即,根据泛生论的假说,它们是由来自雄者身体各个部分的芽球(gemmules)所决定的,这等芽球在雌者身上也是存在的,却处于潜伏状态;芽球的发育乃是由雌者构造组织的选择亲和力的轻微变化所左右的。
还必须对羽衣的季节性变化稍作补充。根据以前所举出的那些理由,关于白鹭、苍鹭以及其他许多鸟类只在夏季发育和保持的华丽羽饰、下垂的长羽、冠毛等系用于装饰和婚配的目的,并无多大可疑之处,虽说这等羽毛为雌雄二者所共有。这样就会使雌鸟的色彩在孵卵期间比在冬季更显著;但是,像苍鹭和白鹭这等鸟类大概是能够保卫自己的。然而由于羽饰在冬季大概会带来不方便而且确实没有用处,所以可能通过自然选择而逐渐获得了一年换羽两次的习性,这是为了到冬季去掉其不方便的装饰物的缘故。但这个观点不能引申到许多涉禽类,它们的夏羽和冬羽在色彩上差别很小。关于不能自卫的物种,凡雌雄二者或只有雄者的色彩,在生殖季节变得极其显著者,——雄者在这个季节获得那样长的以致妨碍其飞翔的翼羽或尾羽者,如非洲小型夜鹰(Cosmetornis)和黑羽长尾鸟(Vidua),最初看来,肯定高度可能的是,第二次换羽习性的获得乃是专门为了去掉这等装饰物。然而,我们必须记住,有许多鸟类,如某些极乐鸟、锦雉和孔雀,在冬季并不脱落其羽饰;而且几乎不能认为这些鸟类的体质,至少是鹑鸡类的体质,使它们不可能有两次换羽的习性,因为雷鸟就一年换羽三次。 (517) 因此,在冬季脱换其羽饰或失去其鲜明色彩的许多物种,是否为了免除不便或危险、不然就要受害而获得了这种习性,仍属疑问。
所以我的结论是,获得一年换羽两次的习性在大多数或所有场合中首先是为了某种不同的目的,也许是为了得到比较暖和的冬装;而羽衣在夏季发生的那些变异则是通过性选择而被积累起来的,并在每年的同一季节传递给其后代;这等变异或被雌雄双方所承继,或只被雄性一方所承继,视何种遗传形式占主导地位而定。这一结论比下述见解似乎更为合理,即,物种在所有场合中原本都倾向于在冬季保持其装饰性的羽衣,但由于它带来了不便或危险,结果通过自然选择而摆脱了这种倾向。
有人认为,武器,鲜明色彩以及各种装饰物现在只限于雄者所有,乃是由于通过性选择这等性状向雌雄双方同等传递转变为只向雄性一方传递的缘故,我在本章曾试图阐明支持这一观点的论证是不可信赖的。许多鸟类雌者的色彩是否由于为了保护自己而把最初只限于向雌性一方传递的那些变异保存了下来,也是可疑的。但是,对这个问题的进一步探讨,留待下一章我讨论幼鸟和老鸟的羽衣差异时再进行,将是方便的。
第十六章 鸟类的第二性征(续完)
雌雄二者成熟时的羽衣性状同未成熟时羽衣的关系——六大类别的事例——亲缘密切相近的物种或典型的物种的雄鸟之间的性差异——雌鸟呈现雄鸟的性状——幼鸟羽衣同成鸟夏羽和冬羽的关系——关于全世界鸟类美丽的增加——保护色——色彩显著的鸟类——对新奇的欣赏——有关鸟类四章的提要
我们现在必须考察同性选择有关的受到年龄限制的性状传递。关于在相应年龄的遗传原理的真实性和重要性没有必要在此多赘,因为对这个问题已经进行过足够的讨论了。在列举我所知道的有关幼鸟和老鸟羽衣差异的几项相当复杂规律或几类事例之前,最好先稍作绪论如下。
在所有种类的动物中,如果成年动物的色彩异于幼年动物,且幼年动物的色彩,就我们所能知道的来说,并没有任何特殊的用途,则幼年动物的色彩正如各种胚胎的构造那样,乃是以往性状的保留。但是,只有当几个物种的幼年动物密切相似同时和属于同一类群的其他成年物种密切相似时,才能有把握地坚持这一观点,因为后者乃是这种状况在以往可能存在过的活证据。幼狮以及幼美洲狮(Puma)都有不明显的条纹或成行的斑点,而且像许多亲缘相近的物种那样,无论幼者或老者都有相似的斑记,凡是相信进化论的人都不会怀疑狮和美洲狮的祖先都是一种具有条纹的动物,而且其幼者就像黑色小猫那样,保持了这等条纹的残迹,可是黑色小猫一长大就没有一点这种条纹的残迹了。许多鹿的物种在成熟时没有斑点,而在幼小时都布满了白色斑点,就像某些少数物种在成长状态时那样。再者,还有整个猪科(Suidae)的幼者以及某些亲缘关系相当远的动物、如貘(tapir)的幼者,都有暗色纵条纹;但我们在这里所看到的显然是来自一个绝灭祖先的性状,而现在只被幼者所保存。在所有这等场合中,老年动物的色彩随着岁月的推移而发生改变,但幼年动物仍保持原样,很少改变,这是通过在相应年龄遗传的原理来实现的。
这同一原理可以应用于各个类群的许多鸟类,它们的幼者彼此密切相似而同各自的成年父母差异很大。几乎所有鹑鸡类的幼者以及某些远亲的鸟类、如鸵鸟类的幼者,都被有纵条纹的绒毛;但这种性状反映了如此遥远的事物状况,以致简直同我们没有什么关系。幼小嘴雀(Loxia)最初具有直喙,同其他燕雀类的喙相似,它们未成熟时的具有条纹的羽衣则同成熟红雀(redpol)和成熟雌金雀的羽衣相类似,也同金翅雀、绿鷽以及其他一些亲缘相近的物种的幼者羽衣相类似。有许多种类的鹀(Emberiza),其幼者彼此相似,也同普通鹀(E.miliaria)的成年状态相类似。在鸫的差不多整个大类群中,其幼者的胸部都具有斑点——这是许多物种终生保持的一种性状,但其他物种却完全失去了这种性状,如候鸫(Turdus rnigratorius)即是。再者,许多鸫类的背部羽毛在第一次换羽前都是杂色的,这是某些东方物种终生保持的一种性状。伯劳属(Lanius)许多物种的幼者,一些啄木鸟和一种印度的绿背金鸠(Chalcophaps indicus)的幼者,其体部底面具有横条纹;而某些亲缘相近的物种或整个属当成年时也有相似的斑记。在一些亲缘相近、色彩灿烂的印度金色杜鹃(Chrysococcyx)中,成熟的物种彼此在色彩上差异相当大,然而它们的幼者则无法区别。一种印度瘤鸭(Sarkidiornis melanonotus)的幼者在羽衣方面同一个亲缘相近的属、即树鸭(Dendrocygna)的成熟者密切相似。 (518) 以后还要列举有关某些苍鹭类的相似事实。黑琴鸡(Tetrao tetrix)的幼者同某些其他物种、如红松鸡(T.scoticus)的幼者及其老者相类似。最后,正如精密研究过这个问题的布莱思先生所恰当说过的,许多物种的未成熟羽衣最好地表现了它们的自然亲缘关系;由于所有生物的真正亲缘关系都是由它们来自一个共同祖先这一事实所决定的,因此布莱思先生的这个意见有力地证实了一个信念:即,未成熟的羽衣大致向我们表明了该物种以前的或祖化的状态。
各个不同科的许多幼鸟虽然这样隐约地闪现了其遥远祖先的羽衣状况,但还有其他许多鸟类,无论色彩暗淡的或色彩鲜明的,其幼者却和它们的父母密切类似。在这种场合中,不同物种幼者的彼此相似就不能比它们和各自父母的相似更为密切;当它们成长时也不能和亲缘相近的类型显著相似。它们只不过使我们稍微知道一点其祖先的羽衣状况而已,除非整个类群的所有物种的幼鸟和老鸟都具有同样的一般色彩,否则其祖先大概不会具有相似的色彩。
现在我们要对各类的事例进行考察,以便把雌雄双方或单独一方的幼鸟和老鸟在羽衣方面的差异和相似加以分类。这等规律首先是由居维叶提出的;但随着知识的进展,需要对它们做某种修改和补充。在这个问题的极端复杂性所允许的范围内我曾试图根据从不同方面得到的材料来完成这一工作;但由一位有才华的鸟类学家就这个问题写出一篇内容充实的论文还是非常需要的。为了证实每个规律可以通用到什么程度,我把四种巨著所举出的事实列成了表,这些著作是麦克吉利夫雷关于英国鸟类的著作,奥杜邦关于北美鸟类的著作,杰尔登关于印度鸟类的著作以及古尔德关于澳大利亚鸟类的著作。我在这里可以先谈谈,第一,有若干事例或规律在逐渐互相转化,第二,当提到幼鸟同其父母相似时,并不是说它们是完全相像的,因为幼鸟的色彩几乎永远不及父母的鲜艳,而且其羽毛比较柔软,羽毛形状也往往不同。
规律或事例的分类
(一)如果成年雄鸟比成年雌鸟更为美丽或更为显著,则雌雄幼鸟在第一次羽衣方面同成年雌鸟密切相似,普通家鸡和孔雀就是如此;或者,像偶尔发生的情况那样,它们同成年雌鸟的相似远比同成年雄鸟的相似更为密切得多。
(二)如果成年雌鸟比成年雄鸟的色彩更为显著,则雌雄幼鸟在第一次羽衣方面同成年雄鸟相似,这种情况有时发生,但不多见。
(三)如果成年雄鸟同成年雌鸟相似,则雌雄幼鸟具有特殊的羽衣,如欧鸲的情况就是如此。
(四)如果成年雄鸟同成年雌鸟相似,则其雌雄幼鸟在第一次羽衣方面同成鸟相似,如翠鸟、多种鹦鹉、乌鸦以及篱莺(hedge-warblers)即是。
(五)如果雌雄成鸟的冬羽和夏羽不一样,不论雄鸟和雌鸟是否有所不同,其幼鸟在冬羽方面同雌雄成鸟相似;或者,其幼鸟在夏羽方面同雌雄成鸟相似,但这种情形要罕见得多;或者,幼鸟同雌鸟相似。或者,幼鸟可能具有一种中间的性状;再不然,它们同成鸟的冬羽和夏羽都迥然不同。
(六)在少数场合中,雌雄幼鸟的第一次羽衣彼此相异;幼年雄鸟多少同成年雄鸟密切相似,而幼年雌鸟多少同成年雌鸟密切相似。
第一类 在这一类中,幼年的雌雄二者同成年的雌鸟多少密切相似,而成年的雄鸟同成年的雌鸟则有差异,而且其差异往往极为显著。在所有的“目”中可以举出无数这类例子;只要想一想普通雉、鸭和家雀的例子就足够了。这类事例逐渐进入别类。这样,其雌雄二者在成年时的差异如此轻微,而且幼鸟同成鸟的差异也如此轻微,以致令人怀疑这等事例究竟应该归入现在这一类还是应该归入第三类或第四类。此外,幼年的雌雄二者不但不十分相似,反而可能有轻微程度的差异,如第六类所示。然而,这些过渡性的事例还是少数,或者说,同那些可以严格归入现在这一类的事例比较起来,至少不是非常显著的。
按照一般规律雌雄二者同幼鸟全都相似的那些类群,完善地阐明了现在这法则的确切意义;因为在这等类群中,如果雄鸟和雌鸟确有差异,如某些鹦鹉、翠鸟、鸽子等的情况,那么幼年的雌雄二者就会同成年的雌鸟相似。 (519) 我们看到在某些异常的场合中同样的事实表现得更加明显;例如黑耳仙蜂鸟(Heliothrix auriculata),其雄者显著不同于雌者,因为雄者具有美丽的新月形颈饰和耳簇毛,而雌者的尾羽由于比雄者尾羽长得多而著称;那么,幼年的雌雄二者(除胸部有青铜色斑点外)同成年的雌鸟在所有其他方面都相似,包括尾羽的长度在内,所以雄鸟到了成熟时,其尾羽实际上是变短了,这是一种极不寻常的情况。 (520) 再者,雄秋沙鸭(Mergus merganser)的羽衣色彩要比雌者的更显著,肩羽和次级翼羽都长得多;但成年雄鸟的冠羽,就我所知,同其他任何鸟类的都不同,虽比雌鸟的宽,却相当地短,其长度只有1英寸刚出头;而雌鸟的冠羽竟有2.5英寸长。于是,幼年的雌雄二者同成年的雌鸟完全相似,所以幼鸟的冠羽实际上长度较大,虽然比成年雄鸟的冠羽较窄。 (521)
如果幼鸟同雌鸟密切相似,而且二者都同雄鸟相异,则最明显的结论是只有雄鸟发生了变异。即使在黑耳仙蜂鸟和秋沙鸭那样异常的场合中,可能是其中一个物种的成年雌雄二者最初具有大大加长了的尾羽,另一个物种的成年雌雄二者具有大大加长了的冠羽——此后由于某种无法解释的原因,成年雄鸟部分地失去了这等性状,并以减弱的状态只把这等性状传递给在相应成熟年龄的雄性后代。在本类的事例中只有雄鸟发生改变,这一信念,就雄鸟和雌鸟及其幼鸟之间的差异而言,得到了布莱思先生所记载的一些显著事实的有力支持, (522) 这些事实是关于不同地方的可以互相代表的近亲物种的情况。因为,若干这等代表性的物种,其成年雄鸟发生了一定程度的变异,而且是可以区别的;来自不同地方的雌鸟和幼鸟却是无法区别的,因此它们绝对没有发生过变化。某些印度鸣禽类(Thamnobia)、某些花蜜鸟(Nectarinia)、林鵙(Tephrodornis)、某些翠鸟类(Tanysiptera)、卡利雉 (523) (Gallophasis)以及树鹧鸪(Arboricola)的情况都是如此。
在某些类似的场合中,即夏羽和冬羽相异而雌雄相似的鸟类,其某些亲缘密切相近的物种可以容易地从其夏羽或婚羽加以区别,然而从其冬羽以及未成熟的羽衣方面则无法加以区别。某些亲缘密切相近的印度鹡鸰的情况就是如此。斯温赫先生告诉我说, (524) 苍鹭有一个属叫做池鹭属(Ardeola),它的三个物种分居于各大陆,它们的夏羽,“彼此差异极为显著”,但到了冬季它们简直完全没有区别。这三个物种的幼鸟的未成熟羽衣同成鸟的冬羽密切相似。这一情况越发有趣,因为池鹭属另外两个物种的雌雄二者的冬羽和夏羽同上述三个物种的冬羽及其未成熟羽衣差不多是一样的;几个不同物种在不同年龄和不同季节中所共有的这种羽衣大概向我们阐明了该属的祖先具有怎样的色彩。在所有这些场合中,婚羽已发生了变异,而冬羽和未成熟羽衣则保持不变;我们可以假定婚羽原本是由成年雄鸟在生殖季节中获得的,并且在相应的季节传递给成年的雌雄二者。
问题自然由此产生:在后面这些场合中雌雄二者的冬季羽衣,并且在前面那些场合中成年雌鸟的羽衣,以及幼鸟的未成熟羽衣,何以完全没有受到影响呢?那些在不同地方互为代表的物种几乎总是处于多少不同的条件下,但我们简直不能把只有雄鸟羽衣发生变异的这种情况归因于这一作用,因为雌鸟和幼鸟虽然也处于同样的条件下,却没有受到影响。几乎没有任何事实比许多鸟类雌雄二者的惊人差异更清楚地向我们阐明,同无限变异通过选择的积累相比,生活条件直接作用的重要性就显得非常次要了;因为雌雄二者都吃相同的食物并处于相同的气候中。尽管如此,我们并不排除下述信念,即,随着岁月的推移,新的条件可能对雌雄双方都发生某种直接作用,或者,由于雌雄之间的体质差异,只对某一性别发生作用。我们看到的只是同选择的积累结果相比,其重要性就是次要的了。然而,根据广泛的类推来判断,当一个物种迁入一处新地方时(这必定发生在形成代表性物种之前),它们几乎总要处于改变了的外界条件之中,这等外界条件将致使它们发生一定程度的彷徨变异。在这种场合中,受一种易变因素所左右的性选择——雌者的审美力或鉴赏力——将会对新的色调或其他差异发生作用并把它们积累起来;由于性选择经常在起作用,所以,根据我们所知道的人类对家养动物所进行的无意识选择的结果来源,如果居住在隔离地区的、因而决不能够杂交并把它们新获得的性状混合起来的那些动物,经过了充分的时间以后还没有发生不同的变异,那将是令人惊奇的事。这些意见同样可以应用于婚羽或夏羽,不论它们为雄鸟所专有,或为雌雄二者所共有,都是一样。
虽然上述亲缘密切相近的或代表性的物种的雌鸟及其幼鸟彼此几乎完全没有区别,所以可以区别的只有它们的雄鸟,可是同一属中大多数物种的雌鸟显然还是有区别的。然而雌鸟之间的差异程度像雄鸟之间差异那么大的,则属罕见。我们在整个的鹑鸡科中清楚地看到了这一点:例如,普通雉和日本雉的雌者,特别是金雉和云实树雉的雌者——银雉和野鸡的雌者——彼此在色彩上都很近似,而其雄者之间的差异都达到了异常的程度。伞鸟科、燕雀科以及其他许多科的大多数物种的雌鸟都是如此。按照一般规律,雌鸟的改变不及雄鸟那样大,确无疑问。然而,少数某些鸟类提供了一个异常而费解的例外;例如极乐鸟(Paradisea apoda)和巴布亚极乐鸟(P.papuana)的雌者之间的差异大于它们同各自雄鸟之间的差异 (525) ;后一物种的雌者的体部底面为纯白色,而极乐鸟的雌者的体部底面则为深褐色。再者,我听牛顿教授说,伯劳类(Oxynotus)有两个在毛里求斯岛和波旁岛互为代表的物种,其雄鸟彼此只在色彩上稍有差别,而雌鸟彼此差异甚大。 (526) 在波旁岛的那个物种的雌鸟似乎部分保持了羽衣的未成熟状态,因为乍一看来,雌鸟“会被当做毛里求斯的那个物种的幼鸟”。这等差异可同那些和人工选择无关的某些斗鸡亚品种的差异相比拟,这些亚品种雌鸡彼此很不相同,而雄鸡则几乎无法加以区别。 (527)
关于亲缘相近的物种,其雄鸟之间的差异,我既然用性选择进行了大量的说明,那么对所有普通场合中的雌鸟之间的差异,又该如何解释呢?我们没有必要在这里去考察不同属的物种;因为对这些物种来说,对不同生活习性的适应以及其他动因都会发生作用。关于同属的雌鸟之间的差异,在我观察了各个大类群之后,几乎可以肯定其主要动因乃是雄鸟所获得的性状通过性选择或多或少地传递给了雌鸟。在若干英国鷽类中,其雌雄二者之间的差异或很轻微或很显著;如果我们把绿鷽、苍头燕雀、金翅雀、红腹灰雀、交喙鸟、麻雀等的雌者加以比较,我们就会看出它们之间的差异之点主要在于它们同各自雄鸟局部相似的那些部分;而雄鸟的色彩可以稳妥地归因于性选择。关于许多鹑鸡类的物种,雌雄之间的差异已经达到了极度,如孔雀、雉以及家鸡的情况就是如此,另外还有一些物种,其雄鸟的性状部分地或者甚至全部地传递给了雌鸟。因花雉类若干物种的雌者以一种模糊的状态表现了其雄者所具有的华丽眼斑,尤以尾部为甚。雌鹧鸪和雄鹧鸪的差异之处仅仅在于雌者胸部的红斑较小;而雌野火鸡和雄野火鸡的差异之处则在于雌者的色彩要暗淡得多。珠鸡(guinea-fowl)的雌雄二者则彼此无法区分。这种鸟的色彩平淡、但具有特殊斑点的羽衣最先由雄鸟获得、然后传递给雌雄双方,并非是不可能的;因为它们的羽衣同雄红胸角雉所专有的那种美丽得多的斑点羽衣并无本质的区别。
从某些例子里应该看到从雄鸟向雌鸟的性状传递显然是在一个遥远的时期就已经完成了,此后雄鸟又经历了巨大变化,而它后来所获得的任何性状却没有传递给雌鸟。例如,黑色琴鸡(Tetrao tetrix)的雌者和幼者同红色松鸡(T.scoticus)的雌雄二者及其幼者相当密切类似;因此,我们可以推断说,黑色松鸡系起源于其雌雄色彩几乎和红色松鸡色彩一样的某一古老物种。由于后一物种的雌雄二者所具有的带斑在生殖季节比在其他任何时期更加显著,又由于雄鸟以其更强烈显著的红色和褐色而稍异于雌鸟, (528) 因此我们可以断定,它的羽衣至少在某种程度上受到了性选择的影响。倘情况果真如此,我们可以进一步推断说,雌黑色琴鸡的差不多相同的羽衣是在以前的某个时期同样这么产生出来的。但是从那个时期以后,雄黑色琴鸡便获得了其优美的黑色羽衣,具有分叉而向外卷曲的尾羽;但几乎没有任何这等性状传给了雌鸟,除了雌鸟的尾羽现了卷曲分叉的一点痕迹。
所以我们可以得出结论说,那些亲缘相近但彼此不同的物种的雌鸟,其羽衣之所以往往或多或少地有所不同,乃是由于雄鸟在远期或近期通过性选择所获得的性状不同程度地传递给了雌鸟。但值得特别注意的是,鲜艳色彩的传递要比其他色彩罕见得多。例如,红喉蓝胸的瑞典蓝雀(Cyanecula suecica)的雄者具有艳蓝色的胸,其上有一个亚三角形的红斑;现在几乎相同形状的斑记已传给了雌鸟,但斑记的中央呈暗黄色而非红色,而且其周围的羽毛为杂色而非蓝色。鹑鸡科提供了许多相似的事例;因为诸如在鹧鸪、鹌鹑、珍珠鸡等物种中,其雄鸟的羽衣色彩已大量传给了雌鸟,这等物种没有一个是色彩鲜艳的。雉类是这方面的良好例证,雄雉一般都比雌雉鲜艳得多;但蓝马鸡(Crossoptilon auritum)和欢乐雉(Phasianus wallichii)的雌雄二者彼此密切类似,而且它们的色彩都是暗淡的。我们甚至可以相信:如果这两种雉的雄者羽衣的任何部分已变得色彩鲜艳,这等色彩大概不会传给雌鸟。这些事实强有力地支持了华莱士先生的观点,即,关于那些在孵卵期间暴露在大量危险之中的鸟类,其鲜明色彩从雄鸟向雌鸟的传递已经通过自然选择而受到了抑制。然而,我们一定不要忘记,上述有另一种解释还是可能的;那就是,当雄鸟在幼小和无经验的时期发生了变异而色彩鲜明,它们大概也会暴露在大量危险之中,而且一般会遭到毁灭;反之,年龄较老和富有警惕性的雄鸟,如果以同样的方式发生了变异,它们恐怕不仅能够生存下来,而且在同其他雄性对手的竞争中还会处于有利的地位。那么,在生命晚期发生的变异有专门向同一性别传递的倾向,所以在这种场合中,极端鲜明的色彩大概不会向雌鸟传递的。另一方面,一种较不显著的装饰物,诸如角雉和欢乐雉所具有的,大概不会有什么危险,如果这等装饰物是在幼年早期出现的,一船会传递给雌雄双方。
亲缘密切相近的物种,其雌鸟之间的某些差异,除了由于雄鸟向雌鸟部分传递其性状这一效果之外,还可归因于生活条件的直接或一定的作用 (529) 对雄者来说,任何这类作用一般都会被通过性选择所获得的鲜艳色彩所掩盖;但对于雌者来说,就不是这样了。我们在家禽中所看到的羽衣每个无止境的变化当然都是某种一定原因所造成的结果;而在自然的和更一致的条件下,假定某种色彩决无害处,几乎肯定它迟早会占优势。属于同一物种的许多个体的自由杂交最终会使这样诱发起来的任何色彩变化在性状上成为一致的。
没有人会怀疑许多鸟类雌雄二者的色彩适于保护之目的;也可能有些物种,仅是其雌鸟为了这一目的而发生了改变。正如上一章所阐明的,通过选择把一种传递形式转变为另一种形式,虽然是一个困难的、也许是不可能的过程,但是,通过一开始就限于传递给雌鸟的那些变异的积累,使雌鸟的色彩——与雄鸟无关的色彩——适应于周围的物体,却没有一点困难。如果这等变异不是受到这样的性别限制,那么雄鸟的鲜明色彩就会退化或遭到破坏。许多物种是否只有雌鸟才有这样特殊的改变,在目前来说还很有疑问。但愿我能充分领会华莱士先生的见解;因为承认他的见解就可解决一些难题。任何变异几对雌鸟的保护无所裨益者即被消除,而不单是由于没有被选择、或由于自由杂交而被取消,也不是由于传递给雄者的变异在任何方面都有害于它而被消除。这样,雌鸟的羽衣性状就会保持稳定。如果我们能承认许多鸟类的雌雄二者获得和保持其暗淡色彩是为了保护自己,那么这在解决难题上大概也是一种帮助——例如,岩鹨(Accentor modularis)和普通鹪鹩(Troglodytes vulgaris)的暗淡色彩就是如此,关于它们的暗淡色彩,我们还没有掌握性选择作用的充分证据,然而,当我们断定在我们看来是暗淡的色彩就会对某些物种的雌鸟没有魅力时,应该小心;我们应该记住普通家雀那样的情况,它们的雄鸟和雌鸟差异很大,但没有表现任何鲜明色调。在开阔地面上生活的许多鹑鸡类的鸟为了保护自己,已经获得现有色彩,至少是部分地获得这种色彩,对此大概已没有任何争论。我们知道它们借此被隐蔽得多么好;我们知道羽脚松鸡类当从冬羽换成夏羽时,虽则冬羽和夏羽都具有保护性的色彩,仍受猛禽类为害甚大。但是,我们能够相信黑色松鸡和红色松鸡的雌者在色彩以及斑记上的很轻微差异是用于保护的吗?鹧鸪现在的色彩是否比它们同鹌鹑的色彩相类似可以受到更好的保护吗?普通雉、日本雉和金雉的雌者之间的轻微差异是作为保护之用的吗?或者,它们的羽衣是否可以彼此交换而不受害吗?根据华莱士先生对东方某些鹑鸡类习性所做的观察,他认为这种轻微差异是有益的。至于我自己,我要说的只是:我不信。
以前解释雌鸟的色彩比较暗淡时,我倾向于把重点放在保护作用上,那时我以为雌雄二者及其幼鸟的色彩可能原来都是同等鲜明的;但后来雌鸟由于在孵卵期间招致了危险以及幼鸟由于缺少经验,所以它们的色彩都变得暗淡以作为保护。但这一观点没有得到任何证据的支持,而且是不可能有的事;因为,如果我们这样来设想,那就是使雌鸟及其幼鸟在过去都暴露于危险之中,因此保护其变异了的后裔,此后便成为必要的了。通过逐渐的选择过程,我们还势必使雌鸟和幼鸟具有几乎完全相同的色彩和斑记而且势必使它们在相应的生命时期把这等性状传递给相应的性别。如果假定雌鸟和幼鸟在变异过程的每一阶段中共同都倾向于变得像雄鸟那样的鲜明色彩,那么雌鸟不会变得色彩暗淡,倘幼鸟不参与这种变化,这也是一件多少奇怪的事;因为就我所能发现的来说,还没有一个事例表明雌鸟色彩暗淡而幼鸟色彩鲜明的物种。然而,某些啄木鸟的幼者提供了一个局部的例外,因为它们“头的整个上部都呈红色”,而雌雄二者以后到达成年时,它就减弱为仅仅一圈红线,或者雌鸟到达成年时,它就完全消失。 (530)
最后,关于我们眼前这一类事实最可能的解释似乎是:只有在雄鸟生命相当晚的时期在鲜明色彩或其他装饰性状方面发生的连续变异才被保存了下来;而这等变异的大部分或全部,由于它们出现的时期是在生命晚期,因而从一开始就只向成年的雄性后代传递。雌鸟或幼鸟所发生的任何鲜明色彩的变异,对它们都没有用处,因而不会被选择;若有危险的话,甚至会被消除掉。这样,雌鸟和幼鸟或保持不变,或通过传递从雄者那里接受某些连续变异而发生局部改变(这更是常见得多)。雌雄二者恐怕都受到了它们长期暴露于其中的生活条件的直接作用:但雌鸟将会更好地表现任何这等效果,因为除此之外别无他法可使它大大改变。这些变化以及所有其他变化由于许多个体之间的自由杂交将会保持一致。在某些场合中,特别是在地栖鸟类的场合中,雌鸟和幼鸟为了保护自己,可能发生与雄鸟无关的变异,结果是获得了同样的暗淡色彩的羽衣。
第二类 凡是成年雌鸟比成年雄鸟的色彩更显著的,则雌雄幼鸟的第一次羽衣同成年雄鸟的相似。——这一类事实同上一类恰恰相反,因为在这里雌鸟的色彩比雄鸟的更鲜明或更显著;而其幼鸟,就已知情况来看,是同成年雄鸟相似而不是同成年雌鸟相似。但是,雌雄之间的差异决不像第一类许多鸟类的雌雄差异那么大,而且这一类情况比较罕见。华莱士先生最先注意到雄鸟较不鲜明的色彩和它们承担孵卵义务之间存在着一种独特关系,他非常强调这一点, (531) 认为这是一个决定性的考察,可以证明在孵卵期间暗淡色彩的获得乃是为了保护自己。还有一个不同的观点,在我看来,似乎更为合理。由于这些事实奇特而且为数不多,我将把我所能找到的全部事实简要列举如下。
三趾鹑属(Turnix)有一个组(section),形似鹌鹑,其雌鸟永远大于雄鸟(有一个澳大利亚的物种,差不多要大两倍),对鹑鸡类来说,这却是一种异常的情况。在该属大多数物种中,雌鸟的色彩比雄鸟的更显著而且更鲜明, (532) 但在少数一些物种中雌雄二者则彼此相似。印度三趾鹑(Turnix taigoor)的雄者,其“喉部和颈部缺少黑色,其羽衣的整个色调比雌鸟的较淡而且较不鲜明”。雌鸟看来比雄鸟更爱吵闹,肯定比雄鸟更加好斗得多;因此,当地人民常常养雌鸟而不是养雄鸟,使它们相斗,就像养斗雄鸡那样。英国捕鸟人用雄鸟为媒鸟,置于陷网的近旁,以激起其他雄鸟的竞争心而捕获之,在印度则用三趾鹑的雌者作为媒鸟。当雌鸟被这样用做媒鸟时,它们很快就开始“咕噜咕噜地高声鸣叫,这种叫声在远处还能听到,任何雌鸟听到叫声时就会迅速奔往该地,并开始同放在那里的笼中鸟相斗。”用此方法仅在一天之内就可以捕到12~20只鸟,全都是正在生殖期的雌鸟。当地人民断言,雌鸟在下完卵以后就集合成群并且留下雄鸟去孵卵。没有任何理由可以怀疑这个断言的真实性,它得到了斯温赫先生在中国所作的一些考察材料的支持。 (533) 布莱思先生相信其幼年的雌雄二者同成年雄鸟相似。
彩鹬(Rhynchaea,图62)有三个物种,其雌鸟“不仅比雄鸟大,而且色彩华丽得多”。 (534) 凡是雌雄二者气管构造有所不同的一切其他鸟类,都是雄鸟的气管比雌鸟的更为发达而且更为复杂;但在南方彩鹬(Rhynchaea australis)来说,则雄鸟的气管构造简单,而雌鸟的气管要经过四道显著的盘旋才进入肺部。 (535) 因而这个物种的雌鸟已获得了一种显著的雄性特征。布莱思先生在检查了许多标本之后,查明孟加拉彩鹬(R.bengalensis)的雌雄二者的气管也都不盘绕,这个物种同南方彩鹬如此相似,以致除了脚趾较短以外,简直没有其他区别。亲缘密切相近的类型的次级性征往往差异很大,关于这一法则,上述事实又是一个显著的例证;但这等差异同雌有关时,那便是一种很罕见的情况了。孟加拉彩鹬雌雄幼鸟的第一次羽衣据说同成熟雄鸟的相似。 (536) 也有理由相信其雄鸟承担了孵卵的义务,因为斯温赫先生 (537) 发现其雌鸟在夏末以前就集合成群,正如雌三趾鹑所发生的情况一样。

图 62 彩鹬(Rhynchaea capensis)(采自布雷姆)
灰瓣蹼鹬(Phalaropus fulicarius)和红领瓣蹼鹬(P.hyperboreus)的雌者都比雄者大,它们的夏羽也“装束得更华丽”。但是,雌雄二者在色彩上的差异则决不是显著的,按照斯廷斯特拉普(Steenstrup)教授的材料,灰瓣蹼鹬只有雄鸟担任孵卵的义务;雄鸟在生殖季节中的胸羽也表明了这一点。斑点鸻(Eudromias morinellus)的雌者比雄者大,其体部底面具有红和黑的色彩,胸部新月形白斑以及眼睛上面的条纹都更强烈显著。其雄鸟至少也参与孵卵;但雌鸟同样照看幼鸟。 (538) 我没有能够弄清楚这些物种的幼鸟同成年雄鸟的相似是否甚于同成年雌鸟的相似;因为作出这种比较或多或少是困难的,这是由于两次换羽的缘故。
现在来谈谈鸵鸟目:普通鹤鸵(Casuarius galeatus)的雄鸟由于其体型较小,由于其附器和头部裸皮的颜色远不如雌鸟的那样鲜明,所以任何人都会把它当做雌鸟;并且巴特利特先生告诉我说,在动物园里肯定只有其雄鸟孵卵并照看幼鸟。 (539) 伍德先生说 (540) ,其雌鸟在生殖季节表现了一种最好斗的性情;这时其垂肉变大而且色彩更鲜艳。此外,斑鸸鹋(Dromaeus irroratus)的雌鸟比雄鸟大得多,它具有一个微小的顶结,除此之外,在羽衣其他方面则无法加以区别。然而,它“在愤怒或受到其他刺激时,似乎比雄鸟的力气更大,颈部和胸部的羽毛皆竖起,就像雄火鸡那样。它通常更勇敢而且更好斗。尤其在夜间它会发出一种空洞深沉的隆隆喉音,就像一面小锣的响声一般。其雄鸟的骨骼较纤细,而且较驯顺,愤怒时只发出一种压抑的咝咝声,或一种嘎嘎声”。它不仅承担了孵卵的整个义务,而且还要保护幼鸟不受它们母亲的危害;“因为母亲一看见其后代就变得非常激动,不顾父亲的抵抗,似乎要尽最大努力去毁灭幼鸟。数月之后,把双亲放在一起,还是不安全的,激烈的争吵乃是不可避免的结果,在这场争吵中雌鸟一般都是胜利者。” (541) 因此,关于鸸鹋,我们看到了一种完全颠倒的情况,不但父母的和孵卵的本能颠倒了,而且雌雄二者的正常精神品质也颠倒了;其雌鸟凶猛,好争吵而且喧闹,而雄鸟则温和而且良善。非洲鸵鸟的情况远非如此,因为其雄鸟比雌鸟多少要大些,而且具有比较美观的羽饰,其色彩对比更为强烈,尽管如此,雄鸟还是担负着整个孵卵的义务。 (542)
我将列举我所知道的其他少数事例来作说明,这些事例表明其雌鸟比雄鸟的色彩更为显著,虽然对其孵卵方式毫无所知。关于福克兰群岛(Falkland Islands)上的一种食腐肉的鸢(Milvago leucurus),我在解剖时非常惊奇地发现,那些全身色彩强烈显著的个体——蜡膜和腿部呈橙色,都是成年的雌鸟;而那些羽衣色彩比较暗淡的、腿部呈灰色的个体,都是雄鸟或幼鸟。澳大利亚有一种红喉短嘴旋木鸟(Climacteris erythrops),其雌鸟异于雄鸟之处在于“喉部有美丽发亮的赤褐色斑记,而雄鸟这一部分的色彩则十分平淡”。最后,澳大利亚有一种欧夜鹰,“其雌鸟体型总是大于雄鸟,色彩之鲜艳也超过雄鸟,另一方面,雄鸟初级飞羽上的两个白斑点比雌鸟的更为显著”。 (543)
于是我们看到,雌鸟的色彩比雄鸟的更显著,幼鸟的未成熟羽衣同成年雄鸟的羽衣相似,而不是像前章所述的那样同成年雌鸟的羽衣相似,这些事例虽见于各个“目”,但为数不多。雌雄二者之间的差异量比前一类常常发生的差异量小到无法相比的程度;所以差异的原因,不论它可能是什么,在这里对雌鸟发生的作用,不及对第一类雄鸟发生的作用那样有力或那样持久。华莱士先生认为雄鸟的色彩在孵卵期间为了保护自己而变得较不显著;但是几乎所有上述事例都表明雌雄二者之间的差异并不够大,似不足据以稳妥地接受这一观点。在一些这类场合中,雌鸟较为鲜明的色彩几乎都限于体部底面,而雄鸟体部底面的色彩如果鲜明,它们在孵卵时大概也不会暴露在危险之中。还应该记住,雄鸟不仅在色彩显著程度上稍逊于雌鸟,而且比雌鸟小而弱。此外,它们不仅获得了母性孵卵的本能,而且还不如雌鸟那样好斗和大声喧叫,有一个例子表明它们的发音器官也比较简单。这样,雌雄二者之间的本能、习性、性情、色彩、大小以及某些构造之点便完成了几乎完全的倒置。
现在如果我们可以假定本类的雄鸟已经失去了它们这一性通常具有的一些热情,因而不再急切地寻求雌鸟;或者,如果我们可以假定其雌鸟的数量比雄鸟的多得多——在印度三趾鹑的场合中据说其雌鸟“远比雄鸟更为常见” (544) ——那么这导致雌鸟追求雄鸟而不是被雄鸟所追求,就不见得不可能了。在一定程度上某些鸟类的情况确系如此,我们在雌孔雀、野火鸡以及某些种类的松鸡中所看到的情况就是这样。如果把大多数雄鸟的习性作为指针,则三趾鹑和鸸鹋的雌鸟的较大体型和体力以及异常的好斗性,都必定意味着它们为了占有雄鸟而尽力把竞争的雌性对手赶跑;从这个观点来看,全部事实就变得一清二楚了;因为,雌鸟由于具有鲜明色彩、其他装饰物以及发音能力,所以最能吸引雄鸟,而雄鸟大概最容易受这等雌鸟的媚惑和刺激。于是,性选择就发生作用,不断地给雌鸟增添魅力;而雄鸟和幼鸟则完全不变,或改变甚少。
第三类 凡成年雄鸟和成年雌鸟相似的,则雌雄幼鸟就具有自己特殊的第一次羽衣。在这一类中,雌雄成鸟彼此相似并异于幼鸟。许多种类的鸟所发生的情况就是如此。雄欧鸲和雌欧鸲几乎无法区别,但其幼鸟则大有差异,它具有暗橄榄色和褐色的斑点羽衣。华丽的猩红色鹮鸟的雄者和雌者彼此相似,而其幼鸟却呈褐色;至于这种猩红色虽为雌雄所共有,但显然是一种性的特征,因为在圈养的条件下,这种性征在任何一性都不会充分发育;当鲜艳的雄鸟被圈养时,这种色彩往往就会消失。有许多苍鹭的物种,其幼鸟和成鸟彼此差异很大;成鸟的夏羽虽为雌雄所共有,却清楚地具有婚羽的特征。幼天鹅呈鼠色,而成熟的天鹅却为纯白色;不过再举一些这方面的例子大概就是多余的了。幼鸟和老鸟之间的这等差异,正如上述两类情况那样,显然在于幼鸟保持了既往的古老状态的羽衣,而雌雄老鸟则获得了新的羽衣。如果成鸟具有鲜明的色彩,我们根据刚才对猩红色鹮鸟和许多种苍鹭所做的记述,以及根据第一类诸物种所做的类推,可以得出结论说,这等色彩是由接近成熟的雄鸟通过性选择而获得的;但不同于上述两类情况的是,这种性状的传递虽然限于相同的年龄,却不限于相同的性别。结果,雌雄二者在成熟时彼此相似而异于幼鸟。
第四类 凡成年雄鸟同成年雌鸟相似的,则雌雄幼鸟的第一次羽衣同成鸟的羽衣相似。——在这一类中,幼年的和成年的雌雄二者,不论其色彩鲜艳或暗淡,都彼此相似。我想,这一类情况比上一类情况更为常见。在英国可举之例有:翠鸟、某些啄木鸟、松鸦、喜鹊、乌鸦以及许多色彩暗淡的小型鸟类,如篱莺或普通鹪鹩。但幼鸟和老鸟之间在羽衣方面的相似决不完全,而渐次变为不相似。例如,鱼狗科某些成员的幼鸟不仅在色彩上不及成鸟鲜艳,而且其体部底面的许多羽毛具有褐色的边, (545) 这大概是其已往羽衣状态的痕迹。在这种鸟的同一类群中,甚至在同一属中,例如在锦鹉(Platycercus)的一个澳大利亚的属中,某些物种的幼鸟同其彼此相似的双亲密切相似,而其他物种的幼鸟则同其彼此相似的双亲相当不同。 (546) 普通 鸟的雌雄二者及其幼鸟都彼此相似;但加拿大噪鸦(Perisoreus canadensis)的幼者同其双亲的差异如此之大,以致过去曾把它们描述为不同的物种 (547) 。
鸟的雌雄二者及其幼鸟都彼此相似;但加拿大噪鸦(Perisoreus canadensis)的幼者同其双亲的差异如此之大,以致过去曾把它们描述为不同的物种 (547) 。
在继续往下讨论之前,我愿指出,在这一类以及下述两类的场合中,事实是如此复杂,而且结论是如此可疑,因而对这个问题不感特别兴趣的任何人还是略而不谈为好。
这一类的许多种鸟都是以其鲜艳的或显著的色彩作为特征,这等色彩很少有或者决不会有保护作用;因此大概是雄鸟通过性选择获得了这等色彩,而后传递给了雌鸟和幼鸟。然而,有可能雄鸟选择更有魅力的雌鸟,如果这些雄鸟将其性状传递给雌雄双方的后代,则其结果同雌鸟选择更有魅力的雄鸟所产生的结果是一样的。但有证据可以证明,在雌雄二者一般彼此相似的任何鸟的类群中,这种偶然情况即便曾经有过,也是很少发生的;因为这等连续变异未能传递给雌雄二者的甚至只是少数,则雌鸟也会在美观方面稍微超过雄鸟。在自然状态下发生的情况恰好相反;因为,在雌雄二者一般彼此相似的差不多每一个大类群中,某些少数物种的雄鸟色彩的鲜明程度略胜于雌鸟。也有可能雌鸟会选择比较美丽的雄鸟,而这等雄者反过来也会选择比较漂亮的雌鸟;但这种双重选择过程是否可能发生,还是有疑问的,这是因为其中一性的热情要大于另一性,再者,这种双重选择过程是否比只有一方的选择更为有效,也是有疑问的。因此最合理的观点是,按照动物界的一般规律,就所涉及的装饰性状而言,性选择曾经对雄鸟发生过作用,而且这些雄鸟把它们渐次获得的色彩或是相等地或是几乎相等地传给了其雌雄二者的后代。雄鸟最初发生的连续变异究竟在其接近成熟之后,抑在其十分幼小的时候,是一个更大的疑问,在任何上述一种场合中,只要雄鸟为了占有雌鸟势必同其对手进行竞争时性选择就一定会对雄鸟发生作用;在那两种场合中这样获得的性状就会传递给雌雄双方,而且这种传递系在所有年龄中进行的。但是,这等性状如果是由成年雄鸟获得的,那么就只传递给成鸟,并且在此后的某个时期再传递给幼鸟。因为已经知道,在相应年龄遗传的规律一旦失效,其后代承继那些性状的时期,往往要早于双亲最初出现那些性状的时期。 (548) 在处于自然状况下的鸟类中可以明显地观察到这等情况。例如,布莱思先生见过红色伯劳(Lanius rufus)和冰雪群岛鷉(Colymbus glacialis)的标本,它们在幼小的时候便十分反常地呈现了其双亲的成年羽衣。 (549) 再者,普通天鹅(Cygnus olor)的幼者要到18个月或两岁才脱掉其暗色羽毛而变成白色的;但福勒尔(F.Forel)博士描述过三只精力旺盛的幼天鹅,它们这一窝一共有4只,而这3只一生下来就是纯白色的。这些幼鸟并非白化体(albinoes),因为它们的喙和腿的颜色和成鸟的同一部分接近相似。 (550)
本类雌雄二者以及幼鸟可能按照上述三种方式而彼此相似,用麻雀属的奇妙事例来说明这三种方式,这是值得一试的。 (551) 家雀的雄者同其雌者以及幼者差异很大。其幼者和雌者彼此相似,并在很大程度上同巴勒斯坦麻雀(P.brachydactylus)的以及亲缘关系密切相近的物种的雌雄二者和幼者相似。因此我们可以假定家雀的雌者和幼者大致向我们表明了该属祖先的羽衣状况。且说,山麻雀的雌雄二者和幼者都同家雀的雄者密切相似,所以说,它们全都按照同样的方式发生变异,而且全都背离了它们早期祖先的典型色彩。这可能是由山麻雀的一个早期雄性祖先来实现的,它发生变异,第一可能是在接近成熟的时期,第二可能在它十分幼小的时期,而且无论在上述哪一种场合中,都把改变了的羽衣传递给了雌鸟和幼鸟;或者第三,它也许在成年发生变异,并把其羽衣传递给了成年的雌雄二者,并且,由于在相应年龄遗传的法则失效,在以后某个时期传递给了幼鸟。
这三种方式在这一类情况中何者属于主导地位,仍是无法确定的。最可能的是,雄鸟在幼年发生变异,并把它的变异传递给了其后代的雌雄二者。我可以在此作点补充,我曾查阅各种著作,试图确定鸟类的变异时期对于性状传递给一性或传递给两性究竟可以决定到什么程度,但很少成功。常常提到的那两条规律(即,在生命晚期发生的变异只传递给相同性别的一方,而在生命早期发生的变异则传递给雌雄双方),明显地适用于第一类 (552) 、第二类以及第四类的情况;但对第三类,往往还对第五类 (553) ,而且对小小的第六类就不适用了。然而,就我所能判断的来说,它们可以应用于相当多数的物种;我们千万不要忘记马歇尔博士对鸟类头部突起所作的惊人概括。这两条规律是否一般都能适用,我们根据第八章所举出的事实可以断言,变异的时期在决定传递形式方面是一个重要的因素。
关于鸟类,我们应该用什么标准去判断变异时期的迟或早,是根据同生命期限有关的年龄,还是根据生殖能力,要不根据物种所通过的换羽次数,都是难以决定的。鸟类的换羽,即使在同一科里,有时没有任何可指出的原因也彼此大不相同。有一些鸟类那么早就换羽,以致在其初级翼羽充分成长之前其体部的羽毛就几乎全脱光了;我们不能相信这是事物的原始状态。如果换羽时期提前了,则其成熟羽衣颜色最先发育的年龄会使我们错误地认为它比实际的发育年龄为早。有些鸟类玩赏家所用的鉴定性别的方法可以说明这一点,他们从尚未离巢的红腹灰雀的胸部,从幼小金雉的头部或颈部,拔掉少许羽毛,以确定它们的性别;因为,若是雄鸟,则在拔掉的那些羽毛原处立即会长出有色的羽毛。 (554) 我们只知道少数鸟类的生命期限,所以我们几乎无法用这个标准来作出判断。至于获得生殖能力的时期,有一个值得注意的事实,即,各种鸟类在保持其未成熟的羽衣时就偶尔进行繁育。 (555)
鸟类在保持其未成熟羽衣时就进行繁育的事实似乎同以下信念相反,即,如我所相信的,性选择在使雄者获得装饰性的色彩、羽饰等并通过同等传递把这些性状传递给许多物种的雌者等方面起了重要的作用。如果年龄较小和装饰较差的雄者能像年龄较大和外观较美的雄者那样成功地赢得雌者和繁殖其种类,那么这种相反的情况就是有确实根据的了。可是我们没有理由假定情况确系如此。奥杜邦曾把一种彩鹳(Ibis tantalus)的未成熟雄者进行繁殖当做一种稀罕的事来说,斯温赫先生也提到过黄鹂属(Oriolus)的未成熟雄者发生过这种情况。 (556) 如果任何物种的幼鸟在羽衣尚未成熟的状况下能够比成鸟更成功地赢得配偶,则成年的羽衣大概很快就会消失,因为那些最长久保持未成熟羽衣雄鸟大概居于优势,这样,物种的性状最终要发生变异。 (557) 反之,如果幼鸟在获得雌鸟方面从未成功,那么早期生殖的习性恐怕迟早要归于消灭,因为这一定要消耗体力,但无此必要。
某些鸟类的羽衣在充分成熟后的许多年里还不断增添其美丽;孔雀和一些极乐鸟的尾羽,以及几种苍鹭(如Ardea ludovicana)的羽冠和羽饰就是如此。 (558) 但这种羽毛的不断发育究竟是对连续的有利变异的选择的结果(尽管这个观点对极乐鸟来说是最合理的)还是单纯的不断生长的结果,尚有疑问。大多数鱼类在它们健康和食物丰富的期间,其大小还在不断增长;鸟类的羽饰可能也受一种与此多少相似的法则所支配。
第五类 凡成年的雌雄二者具有不同冬羽和夏羽的,无论其雄鸟是否异于雌鸟,其幼鸟的冬羽同成年雌雄二者的冬羽相似,或同它们的夏羽相似,但这种情况要罕见得多;或者幼鸟仅同雌鸟相似。或者,幼鸟可能具有一种中间的性状;还有幼鸟同雄鸟的冬羽和夏羽都迥然不同。——这一类情况异常复杂;这也不奇怪,因为它们决定于遗传,而这种遗传不同程度地受到三个方面、即性别、年龄以及季节性的限制。在某些场合中同一物种的一些个体至少要经过五种不同的羽衣状态。关于雄鸟只在夏季或更为罕见地在冬夏两季异于雌鸟的物种, (559) 其幼鸟一般同雌鸟相似——所谓北美的金翅雀、显然还有鲜艳的澳洲莫鲁里鸟(Maluri)都是如此。 (560) 关于雌雄二者在夏季和冬季都彼此相像的物种,其幼鸟同成鸟的相似在于:第一,它们的冬羽;第二,它们的夏羽,但这种情况要罕见得多,第三,介于以上两种状态之间;第四,幼鸟可能在所有季节里都和成鸟迥然不同。这四种情况的第一种有一种印度白鹭(Buphus coromandus)为例,其幼鸟和成年的雌雄二者在冬季均呈白色,到夏季,成鸟就变为浅金黄色。关于印度的懒钳嘴鸭(Anastomus oscitans),其情况与此相似,但其色彩正好相反:因为在冬季其幼鸟和成年的雌雄二者均呈灰色和黑色,到夏季其成鸟就变为白色。 (561) 作为第二种情况的一个例子,如剃刀喙海雀(Alca torda,Linn.),其幼鸟的早期羽衣在色彩上同成鸟的夏季羽衣相像;而北美白冠燕雀(Fringilla leucophrys)的幼鸟,一会飞时,头上就有优美的白色条纹,到冬季幼鸟和老鸟的这等性状都消失了。 (562) 关于第三种情况,即幼鸟具有介于成年的夏羽和冬羽之间中间性状,雅列尔 (563) 坚决认为许多涉禽类都发生过这种情况。最后,关于幼鸟和雌雄二者的成年夏羽和成年冬羽都迥然不同的情况,在北美和印度的一些苍鹭以及白鹭中均有发生——只有它们的幼鸟呈白色。
关于这些复杂的情况我仅稍作陈述。凡幼鸟和雌鸟的夏羽相似或同成年雌雄二者的冬羽相似的,其情况同第一类和第三类情况的不同之点仅在于雄鸟在生殖季节最先获得的并限于在相应季节传递的那些性状。凡成鸟具有不同的夏羽和冬羽,而且其幼鸟异于雌雄二者的,其情况就比较费解了。我们可以承认幼鸟大概保持了一种古老的羽衣状态;我们根据性选择能够说明成鸟的夏羽或婚羽,可是我们如何说明其不同的冬羽呢?如果我们能承认这种羽衣在所有场合中都是作为保护之用的,那么它的获得就是一件简单的事了;但似乎没有恰当的理由来承认这一点。或可提出这样的意见:冬季和夏季迥然不同的生活条件对这种羽衣发生了直接作用;这也许有一些影响,但我没有多大信心来承认我们所看到的这两种羽衣之间的如此重大差异是这样引起的。比较合理的一种解释是,通过夏羽某些性状的传递古老的羽衣样式发生了部分变异,而这种古老的羽衣样式在冬季还为成鸟所保持。最后,属于这一类的所有情况,显然都是由成年雄鸟获得的性状所支配的,这些性状的传递受到了年龄、季节和性别的各种不同限制;但要试图进一步探明这些复杂的关系也许是不值得。
第六类 凡幼鸟的第一次羽衣按不同性别而彼此相异的,则幼小的雄鸟同成年的雄鸟多少密切相似,而幼小的雌鸟同成年的雌鸟多少密切相似。属于这一类的情况,虽出现于各种类群,但为数不多;幼鸟一开始就应该或多或少地和词一性别的成鸟相似,而且逐渐变得越来越相似,好像是最自然不过的事了。黑冠莺(Sylvia atricapilla)的成年雄鸟的头部为黑色,而雌鸟的头部则呈红褐色;布莱思先生告诉我说,其雌雄幼鸟甚至在还没有离巢的时候也能根据这等性状加以区分。在鸫这一科中,有异常多的相似事例已被注意到了;例如雄乌鸫(Turdus merula)的雏鸟就能同雌乌鸫的雏鸟相区别。饶舌鸫(Turdus polyglottus,Linn.)的雌雄二者彼此差异很小,可是其雄鸟在年龄很小时由于呈现更纯的白色而能容易地同雌鸟区分开来。 (564) 树鸫(Orocetes erythrogastra)和岩鸫(Petrocincla cyanea)的雄鸟有很多羽毛呈现一种优美的蓝色,其雌鸟则呈褐色;这两个物种的雄性雏鸟其主要的翼羽和尾羽都有蓝色的边,而雌鸟则具有褐色的边。 (565) 幼小乌鸫的翼羽表现了成熟的性状,而且在其他羽毛之后变黑;反之,刚才提到的那两个物种的翼羽则在其他羽毛之前变蓝。关于这一类情况,最合理的观点是,和第一类情况有所不同,即,雄鸟把这种色彩传递给雄性后代的年龄早于它们最初获得这种色彩的年龄;因为,如果雄鸟是在十分幼小时发生变异的,那么其性状大概就会传递给雌雄二者。 (566)
有一种蜂鸟,叫亮羽蜂鸟(Aithurus polytmus),其雄鸟具有黑和绿的灿烂色彩,而且有两条大小延长了的尾羽;雌鸟只有一条平常的尾羽而且色彩也不显著;于是其幼小雄鸟不是按照普通的规律同成年雌鸟相似,而是以一开始就呈现这种性别所固有的那种色彩而且其尾羽也迅速变长了。我感谢古尔德先生提供这个材料,他还向我提供下述更惊人的而且迄今尚未发表的事例。有两种属于Eustephanus的蜂鸟均具美丽的色彩,栖居于胡安·费尔南德斯的小岛上,而且永远被划为不同的物种。但最近已经证实,其中呈鲜艳栗褐色、头部为金红色的一种乃是雄鸟,而呈绿色和白色斑驳的优美色彩、头部为金绿色的另一种则是雌鸟。于是,其幼鸟在最初就同相应性别的成鸟或多或少地相似了,以后这种相似逐渐变得越来越完全。
如果我们像以前那样地把幼鸟的羽衣作为我们的指针,那么雌雄二者似乎彼此无关地各自变得美丽;而不是某一性把其美丽部分地传递给了另一性。雄鸟显然是通过性选择获得了鲜明的色彩,其方式正如第一类场合中的孔雀和雉那样;而雌鸟则是按照第二类场合中彩鹬属或三趾鹑属的雌鸟那样方式获得其显著色彩的。但是要理解同一物种雌雄二者为何能够同时实现这一过程那就困难得多了。沙尔文先生说,正如我们在第八章所见到的,关于某些蜂鸟,其雄鸟在数量上大大超过了雌鸟,而栖居在同一地方的其他物种则是雌鸟的数量大大超过了雄鸟。于是,如果我们可以假定在以前某个长时期里胡安·费尔南德斯物种的雄鸟在数量上大大超过了雌鸟,而在另一个长时期里又是雌鸟的数量远远超过了雄鸟,那么我们就能理解如何通过对任何一性的色彩鲜明的诸个体的选择在某一个时期使雄鸟、又在另一个时期使雌鸟增添了美丽;雌雄二者在比平常相当早的年龄把它们的性状传递给了幼鸟。这种解释正确与否,我不想妄加评说;但这个事例极其值得注意,以致不能把它忽略过去。
我们现在已于所有这六大类中看到幼鸟羽衣和雌雄两性的或某一性别的成鸟羽衣之间所存在的密切关系。这些关系可根据下述原理得到相当确切的解释,即,某一性别——在大多数场合中这是雄性——通过变异和性选择最先获得了鲜明色彩或其他装饰物,然后按照那些公认的遗传规律以不同方式把它们传递下去。为什么有时即使是属于同一类群的物种,它们的变异也会发生于生命的不同时期,我们还不清楚,但就传递的形式而言,一个重要的决定性原因似乎是变异最先出现时的年龄。
根据在相应年龄遗传的原理,并且根据雄者早期发生的任何颜色变异,在那时都不受到选择的原理——反之往往由于有危险而被淘汰——尽管在生殖期前后发生的相似变异都被保存了下来,幼鸟的羽衣还往往会保持不变或变得很少。这样我们就可略窥现存物种的祖先是什么颜色了。在这六类情况中的五类有大量物种其某一性别或雌雄两性的成鸟是色彩鲜明的,至少在生殖季节期间是如此,而同时幼鸟的色彩总不如成鸟的鲜明,或是色彩十分暗淡;因为,就我所能发现的来说,没有一个事例表明色彩暗淡的物种其幼鸟呈鲜明色彩的,或色彩鲜明的物种其幼鸟比其父母更鲜艳。然而,在第四类中,幼鸟和老鸟彼此相似,有许多物种(尽管决非全部物种)其幼鸟呈鲜明的色彩,既然古老类群是由这些物种组成的,所以我们可以推论其早期祖先也同样是鲜明的。除了这一例外,如果我们从全世界鸟类去看,自从它们的未成熟羽衣给我们作了部分记录的那个时期以来,它们的美似乎大大增加了。
同保护有关的羽衣色彩
已经看到我不能追随华莱士先生去相信暗淡的色彩,当限于雌鸟所专有时,在大多数场合中乃是特别为了保护而被获得的。然而,毫无疑问,如上所述,许多鸟类雌雄二者的色彩都发生了变异,以便逃避其敌害的注意;或者在某些事例中,乃是为了接近其捕食对象时不会被发觉,正如猫头鹰具有柔软的羽毛乃是为了在飞行时不闻其声。华莱士先生说,“只有在热带不落叶的丛林里我们才会找得到其主要色彩是绿色的整群鸟类。” (567) 凡曾试过的,每个人都会承认要把鹦鹉从一株长满绿叶的树识别出来有多么困难。尽管如此,我们还必须记住许多鹦鹉都装饰了深红的、蓝的和橙黄的色彩,而这些色彩几乎是没有保护作用的。啄木鸟显然是树栖的,但除了绿色物种以外,还有许多黑的以及黑白相间的种类——所有这些物种都明显地暴露在几乎相同的危险之中。因此,大概是出没于树间的鸟类通过性选择获得了强烈显著的色彩,但由于额外的保护利益,故绿色的获得往往多于任何其他颜色。
关于在地面上生活的鸟类,每个人都承认它们的色彩是模拟其周围地面的。要看见一只伏在地上的小鹑、沙锥、丘鹬(Woodcock)、某些鸻类、云雀和欧夜鹰是多大地困难。在沙漠居住的动物提供了最惊人的事例,因为那里的地面是裸露的,没有藏身之处,几乎所有较小的四足兽类、爬行类以及鸟类的安全都依靠其体色。特里斯特拉姆先生说,所有撒哈拉的居住者都是以其“淡黄色或沙色”来保护自己。 (568) 当我想起南美的沙漠鸟类以及大不列颠的大多数地栖鸟类,在我看来其雌雄色彩一般差不多是一样的。因此,我向特里斯特拉姆先生请教有关撒哈拉沙漠鸟类的情况,他好心地向我提供了下述材料。有15个属的26个物种,都明显具有保护色彩的羽衣;这种保护色越发惊人的是,大多数这等鸟类的色彩都异于其同种的鸟类。在这26个物种中有13个其雌雄二者的色彩是一样的;但这些物种都是属于通常受这一规律所支配的属,因此,关于沙漠鸟类雌雄二者具有一样的保护色,这些物种并没有向我们说明什么。至于其他13个物种,其中有3个是属于其雌雄二者平常有差异的属,而在这里它们都彼此相似。其余的10个物种,其雄鸟异于雌鸟,但这种差异主要局限于羽衣的底面,当这等鸟伏于地面时就把这一部分掩盖住了;其雌雄二者的头部和背部也具有同样的沙色。因此,这10个物种的雌雄二者的上表面为了保护之故在自然选择的作用下而彼此相像;同时为了装饰之故只有雄鸟的体部底面通过性选择而多样化了。在这里,既然雌雄二者都相等地得到了妥当的保护,我们可以清楚看到自然选择并没有阻止雌鸟继承其父方的色彩;因此我们必须求助于受性别限制的传递法则了。
在世界所有部分的许多软嘴鸟类,特别是那些常常出没于芦苇或苔草中的鸟类,其雌雄二者均呈暗淡的色彩。毫无疑问,如果其色彩是鲜明的,惹起其敌害的注目就要容易得多;可是它们的暗淡色彩是否特别为了保护自己而获得的,就我所能判断的来说,似乎还有相当疑问。至于获得这种暗淡色彩是否会为了装饰之故,就更加可疑了。然而我们必须记住,雄鸟尽管是色彩暗淡的,但往往还同雌鸟有很大差异(像普通麻雀那样),这就使我们相信为了吸引雄鸟这等色彩是通过性选择而被获得的。许多软嘴鸟类都是鸣禽;不应忘记前章的那一段讨论,它表明装饰着鲜明色彩的鸣禽是罕见的。如此看来,作为一般的规律,选择配偶的好像是雌鸟,它们或取雄鸟甜蜜的鸣声或取雄鸟漂亮的色彩,而并非二者兼取。有一些物种的色彩显然是为了保护自己,诸如姬鹬(jacks-nipe)、丘鹬以及欧夜鹰即是,但它们的斑纹和色调按照我们的审美标准来看,照样是极其优美的。在这等场合中我们可以作出结论说,自然选择和性选择为了保护和装饰而共同发挥作用。是否有任何这样一种鸟存在,它并不具有某种用来诱惑异性的特殊魅力,实属可疑。如果雌雄色彩是如此暗淡,以致不应轻率地去假定性选择可以发生作用,而且如果不能提出直接的证据来表明这等色彩系作为保护之用,那么最好还是承认我们对其原因一无所知,或者,几乎是一回事,把其结果归因于生活条件的直接作用。
许多鸟类雌雄二者的色彩虽不鲜艳,却是触目的,诸如数目众多的黑的、白的或黑白相间的物种即是;这些色彩大概都是性选择的结果。普通翅鸫、雷鸟、黑色公松鸡、黑凫(Oidemia)、甚至还有一种黑极乐鸟(Lophorina atra),只有其雄鸟是黑色的,而雌鸟则呈褐色或杂色;这种黑色乃是一种性选择的性状,几乎是无可怀疑的。所以像乌鸦、某些美冠鹦鹉、鹳和天鹅,以及许多种海鸟那样的一些鸟类,其雌雄二者的全局部的黑色同样是性选择的结果,并伴以向雌雄双方的同等传递,在某种程度上是可能的;因为黑色在任何场合中几乎都不能作为保护之用。有若干鸟类,仅是其雄鸟呈黑色,另有一些鸟类,其雌雄二者均呈黑色,而它们的喙或头皮则呈鲜明的色彩,由此产生的颜色对比大大增添了它们的美;从雄翅鸫的鲜明的黄喙,从黑色公松鸡和雷鸟眼睛上方的艳红色皮肤,从雄黑凫的各种鲜明颜色的喙,从黄嘴山鸦(Corvus graculus,Linn.)、黑天鹅以及黑鹳的红喙,我们看到了这种美。这个情况使我注意到巨嘴鸟的巨型鸟喙也许是性选择的结果,这样来显示装饰在其巨喙上的各式各样色彩鲜艳的条纹。我看这并不是不可信的。 (569) 鸟喙基部和眼睛周围的裸皮往往也同样是色彩灿烂的;古尔德先生在提到某一个物种时说道,其喙色“在支配期间无疑是最优美和最灿烂的”。 (570) 巨嘴鸟以其巨喙来显示其优美的色彩(在我们看来,误以为不重要),巨喙的松质的构造虽然尽可能使它变轻,但也给它带来了不便,正如雄锦雉以及某些其他鸟类的羽饰妨害了它们的飞行,给它们带来了不便一样,前一情况的可能性未必小于后一情况。
如上所述,各个物种只有雄鸟呈黑色,而雌鸟则呈暗色,同这种情况一样,在少数场合中只有雄鸟全部或局部呈白色,而雌鸟则呈褐色或暗淡的杂色,如南美的几种铃鸟(Chasmorh ynchus)、南极黑雁(Bernicla antarctica)、银雉等即是。因此,按照上述同样的原理,许多鸟类的雌雄二者通过性选择获得了其多少完全白色的羽衣是可能的,诸如鹮、若干种有漂亮羽饰的白鹭、某些鹮类、鸥类(gulls)、燕鸥类(terns)等就是这样。在某些这种场合中,只有到了成熟期羽衣才变为白色。若干种鲣鸟(gannets)、热带鸟类等,还有雪雁(Anser hyperboreus)的情况均系如此。后者既然是在尚未被雪覆盖的“不毛地面上”产卵繁育,并且在冬季向南迁居,因此毫无理由假设其雪白的成年羽衣是作为保护之用的。关于懒钳嘴鸭,我们有更好的证据可以证明这种白色羽衣乃是一种婚羽的性状,因为它只在夏季发育;而不成熟的幼鸟以及身着冬装的成鸟都呈灰色和黑色。至于有许多种类的鸥(Larus),它们的头部和颈部在夏季变为纯白色,在冬季以及在其幼小状态则呈灰色或杂色。反之,较小的鸥鸟、或潜鸟(Gavia)以及某些种燕鸥(Sterna),所发生的情况正好相反;因为其幼鸟在头一年,以及成鸟在冬季,它们的头部或呈纯白色或比在生殖季节的色彩暗淡得多。后面这些情况提供了另一种事例,表明性选择好像往往以不定的方式发生作用。 (571)
水栖鸟类获得白色羽衣者远比陆栖鸟类常见得多,这大概决定于水栖鸟类的巨大体型和强大飞行能力,所以它们能容易地防卫自己或逃避猛禽类,此外它们和猛禽类也不常遇。结果,性选择在这里并没有受到保护作用的干扰或支配。毫无疑问,在广阔海洋上方翱翔的鸟类,当因其全白色或深黑色而容易得见时,其雄鸟和雌鸟的彼此觅得就容易得多;因此这等色彩可能就像许多陆栖鸟类呼唤鸣叫那样地用于同一目的。 (572) 当一只白色的或黑色的鸟发现一具在海上漂动或冲上海滩的尸体并向它飞下时,在很远的地方就可看到这只鸟。而且它会引导同一物种的和其他物种的鸟飞向那具尸体,但是,这对第一个发现者是不利的,所以那些最白的或最黑的个体所获得的食物不会多于色彩较不强烈的个体。因此不能为此目的通过自然选择而逐步获得这种显著色彩。
既然性选择受到像审美那样一种彷徨不定的因素所支配,于是,我们就能理解在具有几乎相同习性的鸟的同一类群中,何以会存在白的或接近白的物种,而且存在黑的或接近黑的物种——例如,既有白的又有黑的美冠鹦鹉、鹳、鹮、天鹅、燕鸥和海燕。同样地,黑白斑物种有时和黑的以及白的物种一起在同一类群中出现;如黑颈天鹅,某些燕鸥和普通喜鹊皆是。我们在经过彻底调查任何大量采集品之后可以断言,鸟类对强烈对比的色彩是喜爱的,因为雌雄二者的差异往往是,就淡白色和纯白色的对比而言,雄鸟的白色比雌鸟的更纯,就各种暗色同较深的暗色对比而言,雄鸟的暗色比雌鸟的更深。
甚至还会出现这种情形:单纯的新颖,即为了改变而发生的轻微变化,有时也会作为一种魅力对雌鸟发生作用,就像风尚的改变对我们发生作用一样。例如,有些鹦鹉的雄者几乎不能说比其雌者更美丽,至少按我们的审美标准来看是如此,但它们有这样几点是不同的,如具有一条玫瑰色的颈圈,而不是“一条鲜明的翡翠般的绿色狭颈圈”,或者雄鹦鹉具有一条黑颈圈,而不是“位于颈部前方的黄色半颈圈”,而且其头部呈浅玫瑰色而不是梅青色。 (573) 既然有那么多雄鸟以其延长的尾羽或冠羽作为它们主要的装饰,所以上述雄蜂鸟的缩短尾羽以及雄秋沙鸭的缩短冠羽,大概就像我们所欣赏的许多时装改变中的一种改变那样。
苍鹭科的一些成员提供了一个更为奇妙的例子,表明新颖的颜色似乎为了新颖而受得赞赏。灰鹭(Ardea asha)的幼鸟呈白色,而其成鸟则呈暗鼠色;亲缘相近的一种印度白鹭(Buphus coromandus)不仅其幼鸟而且其成鸟的冬羽均呈白色,这种白色到生殖季节就变为鲜艳的淡金黄色。要说这两个物种的以及同一科某些其他成员的幼鸟由于任何特殊目的而变为纯白色 (574) 并因而引起其敌害的注目,乃是不可置信的,或者说这两个物种之一,其成鸟在一个冬季从不下雪的地方特别变为白色,也是不可置信的。反之,我们有良好的理由来相信许多鸟类获得这种白色乃是作为性的装饰的。因此,我们可以作出结论说,灰鹭和印度白鹭的某一早期祖先为了婚配的目的而获得了白色羽衣,并把这种颜色传递给了幼鸟;因而其幼鸟和老鸟就像某些现存的白鹭那样变成了白色;此后这种白色由幼鸟保存了下来,同时成鸟以更强烈显著的色彩代替了这种白色。但是,如果我们能够进一步追溯这两个物种的早期祖先,我们大概可以看到,其成鸟是色彩暗淡的。根据幼鸟呈暗色、成鸟呈白色的许多其他鸟类的类比,我可以推论出情况大概就是这样的;根据另一种喉鹭(Ardea gularis)的情况来类比,问题就清楚了,这种鹭的色彩同灰鹭的色彩正相反,其幼鸟呈暗色,而成鸟呈白色,幼鸟保持了往昔的羽衣状态。因此,在悠久的生物由来的系统中,灰鹭、印度白鹭及其某些近亲的成年祖先似曾经历了下述变化:第一,暗淡的色调;第二,纯白色;第三,由于风尚(如果我可这样表达的话)的另一种变化,它们表现了现今的鼠色、微红色或淡金黄色。只有根据鸟类本身崇尚新颖这一原理才可以解释这等连续的变化。
某些作者设想雌性动物和未开化女人对某些色彩和其他装饰物的审美不会在许多代里保持固定不变;它们最初赞美这一种颜色,后来又赞美另一种颜色,结果就不能产生持久的效果;他们以此来反对性选择的整个理论。我们可以承认审美是彷徨不定的,但并不是完全随心所欲的。它多半是由习性所决定的,我们在人类所看到的就是如此;我们可以推论这也适用于鸟类和其他动物。即使我们的服装也长期保留了其一般的特征,其变化在某种程度上还是逐渐的。在后面一章的两个地方将举出大量的证据,表明许多种族的未开化人若干代以来都赞美同样的皮肤疮痕、同样丑陋的穿孔的嘴唇、鼻孔、或耳朵,变形的头部等等;而这种毁形同各种动物的天然装饰物表现了某种类似。尽管如此,对未开化人来说,这种风尚并不会永久保持下去,因为我们可以从同一个大陆上亲缘相近的部落之间在这方面的差异推论出来。再者,珍奇动物的饲养者许多代以来肯定赞美一些同样的品种,而且现今还在赞美这些品种;他们热切期望出现一些轻微的变化,轻微的变化被视为改进,而任何重大的或突然的变化则被视为最大的瑕疵。关于自然状况下的鸟类,我们没有理由设想它们会赞美一种样式全新的色彩,即使重大的或突然的变异常常发生也是如此,而在自然状况下的变异决不会这样。我们知道,普通家鸽不愿意同各种色彩的珍奇品种合伙;白化的鸟平常找不到配偶;法罗群岛(Feroe Islands)的黑渡鸟把其黑白斑的弟兄赶走。但是,对突然变异的这种厌恶并不妨碍它们对轻微变化的欣赏,这种情况不会有任何超出人类好恶的地方。因此,审美是受许多因素所支配的,但它部分决定于习性,部分决定于对新颖的爱好,关于审美,动物既很长时期地赞美,装饰物或其他吸引物的同样的一般样式,还欣赏颜色、形状或声音的轻微变化,这似乎不是没有可能性的。
关于鸟类四章的提要
大多数雄鸟在生殖季节是高度好斗的,而且有的还具有适于同其竞争对手进行战斗的武器。但最好斗和武装得最好的雄鸟很少或从来不是单单依靠赶跑或杀死其竞争对手的能力而取得成功的,它们还有媚惑雌鸟的特殊手段。这等手段有些是鸣唱的能力,或发出奇怪的叫声,或为器乐的演奏,结果雄鸟在发音器官、或某些羽毛构造方面就要同雌鸟有所差异。根据产生各种声响的多种多样的奇特手段,我们深刻意识到这种求偶手段的重要性。许多鸟类在地面或天空、有时还在事先预备好的地方进行爱情舞蹈或作滑稽表演以尽力媚惑雌鸟。但许多种类装饰物,最鲜艳的色彩,鸡冠和垂肉,漂亮的羽饰,延长的羽毛,顶结等等,乃是最常见的手段。在某些场合中单是新颖似乎也起了一种媚惑作用。雄鸟的一些装饰物对它们来说一定是高度重要的,因为在并非少数的场合中,获得这等装饰物是以增加来自敌害的危险为代价,甚至以损害同其竞争对手进行战斗的某种能力为代价的。很多物种的雄鸟不到成熟时不会披上其装饰性的装束,或只在生殖季节才披上这等装束,或者其色彩到这时才变得更为鲜艳。某些装饰性的附器在求偶行为的期间增大了,饱满了,而且色彩鲜明了。雄鸟精心地夸示其魅力,以达到最佳的效果;这一切都是在雌鸟的面前进行的。求偶有时是一件冗长的事,而且许多的雄鸟和雌鸟集合于一个预定的地点。要是假定雌鸟不欣赏雄鸟的美丽,就无异于承认它们那些灿烂的装饰、它们所有的盛大仪式和夸示魅力都是无用的;而这种假定是不可置信的。鸟类具有敏锐的识别能力,在少数场合中,可以表明它们具有一种审美力。还有,据了解雌鸟对某些雄性个体偶尔显露出一种显著的偏爱或厌恶。
如果承认雌鸟喜爱比较漂亮的雄鸟,或无意识地受到这等雄鸟的刺激,那么雄鸟通过性选择大概就会缓慢地、但肯定地变得越来越富有魅力。主要发生变异的乃是雄鸟,从下述事实我们可以推论出这一点,即:在雌雄相异的几乎每个属中,雄鸟彼此之间的差异远比雌鸟彼此之间的差异大得多;这种情况又可从下述事实得到充分阐明,即:在某些亲缘密切接近的诸代表性物种中,其雌鸟几乎无法区别,而其雄鸟则十分不同。自然状况下的鸟类所提供的个体差异可以充分满足性选择工作的需要;不过我们已经看到它们偶尔会表现更强烈的显著变异,而这等变异如此经常重现,以致它们如果可以诱惑雌鸟就会马上被固定下来。变异的法则必然决定最初变化的性质并将大大影响其最后结果。在亲缘相近物种的雄鸟之间可以观察到的级进指明了它们所通过的那些步骤的性质。它们还以最有趣的方式说明了某些性状是如何发生的,诸如孔雀尾羽上的齿状眼斑,以及锦雉翼羽上的球与穴眼斑。许多雄鸟的灿烂色彩、顶结、优美的羽饰等等不会是作为保护手段而被获得的;它们的确有时会招致危险。我们可以肯定这等性状的产生并不是由于生活条件之直接而一定的作用,因为雌鸟也暴露在相同的条件之下,却往往同雄鸟极度不同。虽然变化了的外界条件的长期作用在某些场合中可能对雌雄两性、有时对某一性别产生一定的效果,但更重要的结果将是一种变异倾向的加强或产生更强烈显著的个体差异;而这种差异将为性选择提供最好的基础。
雄鸟为了装饰自己、为了产生各种声响以及为了彼此相斗而获得的那些性状,永久地或者在一年的某些季节定期地只传递给雄性一方还是传递给雌雄双方,皆由遗传法则来决定,而与选择无关。为什么各种性状有时会按某一种方式传递,有时又按另一种方式传递,在大多数场合中都是我们所不知的;但变异的时期似乎常常是决定性的原因。当雌雄二者共同遗传了所有性状时,它们必然彼此相似;但是,由于连续变异的传递方式不同,因此,甚至在同一属中,从雌雄二者彼此最密切相似到最广泛不相似之间,可以找到每一个可能的级进。关于遵循差不多相同生活习性的亲缘密切相近的物种,其雄鸟彼此之间的差异主要是由性选择作用造成的,而雌鸟彼此之间的差异则主要是由于或多或少地分享了雄鸟这样获得的那些性状。加之,由于强烈显著的色彩和其他装饰物通过性选择而被积累起来,生活条件一定作用的效果在雄鸟中便被掩盖,而在雌鸟中则不然。雌雄二者的诸个体虽受这样影响,但由于许多个体间的自由杂交,在各个相继的时期内仍能保持接近一致。
关于雌雄色彩相异的物种,有些连续变异可能是常常倾向于相等地传递给雌雄双方的;但是,当这种情况发生时,雌鸟由于在孵卵期间所遭到的毁灭,它获得雄鸟的鲜明色彩就会受到阻止。没有任何证据可以证明通过自然选择把一种传递形式转变为另一种是可能的。但是,通过一开始就限于传递给同一性别的连续变异的选择,使一只雌鸟呈暗淡色彩而雄鸟仍保持其鲜明色彩,并没有丝毫困难。许多物种的雌鸟是否实际上发生了这样的改变,目前一定还有疑问。通过性状相等地向雌雄双方传递的法则,当雌鸟的色彩变得和雄鸟一样显著时,它们的本能似乎也常常发生了改变,所以它们被引导去建造有圆顶的或荫蔽的鸟巢。
一小类奇妙的事例表明,雌雄二者的性状和习性正好完全颠倒,因为雌鸟比雄鸟更大,更强壮,更好斗,而且色彩更鲜明。它们还变得如此爱争吵,以致为了占有雄鸟而常常互相搏斗,就像其他好斗物种的雄鸟为了占有雌鸟而相斗一样。如果这类雌鸟惯常地把其竞争对手赶走,并且靠显示其鲜明色彩或其他魅力以尽力吸引雄鸟的话——看来这是很可能的,那么我们就能理解它们如何通过性选择和受性别限制的遗传而逐渐变得比雄鸟更美丽——而后者则保持不变或只有轻微的改变。
无论何时,只要在相应年龄遗传的法则起支配作用,而不是受性别限制的,遗传法则起支配作用,那么,如果其双亲发生变异的时候是在生命晚期——我们知道,我们的家鸡偶尔也有其他鸟类所发生的情况永远如此——则其幼鸟将不受影响,而其成鸟的雌雄二者将会发生改变。如果这两个遗传法则都起支配作用,而且无论哪一性别的变异均发生在生命晚期,那么就只有那一性别才发生改变,而另一性别和幼鸟都不受影响。当鲜明色彩或其他显著性状的变异都发生在生命早期时,无疑就像常常发生的情况那样,不到生殖时期性选择不会对它们发生作用;因而,如果它们对幼鸟有危险的话,就会通过自然选择而被淘汰。这样,我们就能理解那些发生在生命晚期的变异何以被保存下来,作为雄鸟的装饰;而雌鸟和幼鸟则几乎不曾受到影响,所以彼此相似。关于具有不同的夏羽和冬羽的物种,其雄鸟在冬夏两季或只在夏季不是和雌鸟相似就是和雌鸟有差异,幼鸟和老鸟彼此相似的程度和性质都是极其复杂的;而这等复杂性是由雄鸟最初获得的性状来决定的,这等性状以各种不同的方式,如在年龄、性别和季节的限制下传递下去。
既然有那么多物种的幼鸟在色彩和其他装饰方面只发生很小改变,所以这使我们能够对其早期祖先的羽衣作出某种判断;如就全类情况来看,我们就可推论出我们的现存物种自从那个时期以来已大大增加了其美丽,而幼鸟不成熟的羽衣向我们指明了有关那个时期的间接记录。许多鸟类,尤其是那些多半生活在地面上的鸟类无疑是为了保护自己而呈现暗淡色彩的。在某些事例中,其雌雄二者羽衣暴露在上方表面的均呈暗淡色彩,同时只有雄鸟的底面通过性选择才装饰着各式各样的色彩。最后,根据这四章所列举的事实,我们可以作出结论说,战斗的武器、发声的器官、许多种类的装饰物、鲜明而显著的色彩,一般都是由雄鸟通过变异和性选择而获得的,并且按照几项遗传法则以各种不同的方式传递下去——雌鸟和幼鸟则相对地改变很小。 (575)
第十七章 哺乳类的第二性征
斗争的法则——限于雄者所有的特殊武器——雌者不具武器的原因——为雌雄二者所共有的、而最初由雄者所获得的武器——这等武器的其他用途——它们的高度重要性——雄者的较大身体——防御的手段——四足兽雌雄任何一方对配偶的选择
关于哺乳类动物,其雄者赢得雌者似乎是通过斗争,而不是通过魅力的夸耀。最怯懦的、不具有任何特殊斗争武器的动物,在求爱季节也进行殊死的冲突。两只雄野兔据知相斗到其中一只死去为止,雄鼹鼠常常相斗,有时会造成致命的结果;雄松鼠屡屡争斗,“彼此皆负重伤”,雄河狸也是如此,因而“几乎没有一张皮不是有伤痕的”。 (576) 我在巴塔戈尼亚看到美洲羊驼(guanacoes)的皮也是伤痕累累;有一次几只美洲羊驼如此精神贯注地进行争斗,以致冲到我身旁也无所畏惧。利文斯通说,南非许多雄性动物差不多都显示有在以往争斗中所负的伤痕。
水栖哺乳动物也受斗争法则的支配,与陆栖哺乳动物无异。众所周知,雄海豹在繁殖季节如何用牙和爪拼命地进行争斗;它们的皮同样也是伤痕累累。雄抹香鲸在繁殖季节是很嫉妒的;在斗争中“它们的颚往往咬在一起,扭来扭去”,所以它们的下颚常常被弄歪。 (577)
众所熟知,具有特殊战斗武器的一切雄性动物都进行猛烈斗争。关于雄鹿的勇敢及其殊死的争斗,常见于记述;世界各地都曾发现过它们的骨骼,双方的角紧紧扭在一起而不可解,表明争斗双方同归于尽。 (578) 世界上没有一种动物比求偶时的象更为危险的了。坦克维尔(Tankerville)勋爵给过我一份有关奇玲根(chillingham)狩猎公园中公野牛相斗的图解,它们是巨大原牛(Bos primigenius)的后裔;虽在身体大小上退化了,但勇气依然如旧。1861年有数牛争霸;人们看到有两头比较年轻的公牛合伙向一头老的带头公牛进行攻击,把它打倒,使其丧失战斗力,所以狩猎公园管理人以为这头老公牛已经受到致命伤而倒在附近的树林中了。但是,数日之后当其中一头幼公牛单独走近那片树林时,这位“狩猎地之王”激起了复仇的火焰,跑出树林,很快就把它的敌对者弄死了。于是这头老公牛悠然地回到牛群,长期保持了其无可争辩的统治。沙利文海军上将告诉我说,当他住在马尔维纳斯群岛时,他曾输入一匹英国幼种马,它常同八匹母马往来于威廉港(Port William)附近的山中。在这座山里还有两匹野公马,各领一小群母马;“这些公马一相遇就要发生争斗。这两匹野公马都曾试图单独地同那匹英国种马争斗并把它的母马赶走,但都失败了。有一天,这两匹野公马一齐来了,并对英国种马进行攻击。管理马群的队长看到这种情况,乘马驱至该处,发现其中一匹野公马同英国种马争斗,另一匹则驱赶母马,而且已经赶走了四匹。于是那位队长把整个马群赶入畜栏,问题才告解决,否则那两匹野公马是不会舍母马而去的。
雄性动物凡具有用于普通生活目的之切断齿或撕裂齿者,如食肉类、食虫类和啮齿类,很少再有另外同其竞争对手进行战斗的特殊武器。至于许多其他动物的雄者,其情况就很不相同了。我们看到鹿和某些种类的羚羊就是如此,它们的雄者有角,而雌者无角。有许多动物,其雄者的上颚犬齿、或下颚犬齿、或上下颚双方犬齿都远比雌者的大得多,也许雌者完全没有这等犬齿,有时仅留有一个隐蔽的残迹。某些羚羊、麝、骆驼、马、野猪、各种猿类、海豹、海象均为可举之例。雌海象有时完全不具獠牙。 (579) 印度的雄象以及儒艮(dugong)的上颚切齿乃是攻击性的武器。 (580) 雄独角鲸(narwhale)唯有左侧犬齿非常发达,呈螺旋状,有时长达9~10英尺,所谓角者即是。人们相信雄独齿鲸用这种角相斗;因为“很少找到一只没有损坏的角,偶尔在损坏处还会发现另一个齿尖”。 (581) 雄独角鲸的左侧犬齿仅是一个残迹,长约10英寸,埋藏于颚中;但是,有时左右两侧的犬齿也同等发达,虽然这是罕见的。雌独角鲸的左右两侧犬齿永远是残迹的。雄抹香鲸的头大于雌者,在水战中大的头无疑是有助益的。最后,成年的雄鸭嘴兽(Ornithorhynchus)具有一种奇器,即前腿上的距(spur),同毒蛇的毒牙密切相似;但是,按照哈廷(Harting)的说法,这个腺体的分泌物并无毒;而且在雌鸭嘴兽的腿上有一个凹陷,显然为承受那个距之用。 (582)
如果雌者没有雄者所具有的那样武器,则这等武器系用于同其他雄者进行争斗就毫无疑问了;这等武器是通过性选择获得的,而且只传递给雄者。要说雌者由于武器对它们无用、多余或在某一方面有害而被阻止去获得这等武器,乃是不可能的。相反,既然雄者常常把它们用于各种不同的目的,尤其是用于防御其敌手,所以它们在许多动物的雌者身上如此发育不良,或完全缺如,却是一件可怪的事。关于雌鹿,如果在每年的一定季节内有大型的枝角发育,关于雌象,如果有巨大獠牙发育,假定它们对雌者没有任何用处,那么这大概会造成生命力的重大浪费。结果,倘连续变异的传递仅限于雌者,则这等角和牙通过自然选择在雌者方面就会倾向于消失;因为倘不如此,则雄者的武器大概就要受到有害的影响,而且这会造成较大的恶果。从全面来看,并且根据对下列事实来考虑,可能的情况似乎是,如果各种武器在雌雄两方面有所差异,那么这种差异一般决定于通行的传递方式。
在整个鹿科中雌者具角的只有驯鹿这一个物种,但雌鹿们的角比雄驯鹿的角稍小、稍细而且分枝略少,因此,自然会认为这种角对于雌者有某种特殊用途,至少在这一场合中是如此。雌驯鹿的角充分发育时系在九月,从那时起,经过整个冬季,直到四月或五月雌鹿产小鹿时为止,都保持着角。克罗契(Croteh)先生曾在挪威特别为我调查过此事,看来雌驯鹿在这个季节为了生产小鹿似乎要隐匿两周之久,然后又再现,那时一般已经没有角了。然而我听里科斯先生说,在新斯科夏(Nova Scotia)雌驯鹿保持的角的期间有时要长些。另一方面,雄驯鹿角的脱落时期要早得多,约在十一月末。雌雄驯鹿皆有同样的需要,遵循同样的生活习性,而且雄驯鹿在冬季无角,因此这等角在冬季对雌驯鹿来说不见得会有任何特殊用途,冬季占其具角时期的大部分。雌驯鹿的角也未必是由鹿科的某一古代祖先遗传而来的,这是因为地球上所有地方的如此众多的物种的雌者均不具角,所以我们可以作出结论说,这是该类群的原始性状。 (583)
驯鹿的角在极其幼小的时候就发育了;但其原因是什么,现在还弄不清楚。显然是角向雌雄双方传递起了作用。我们应该记住,角永远是通过雌者向下传递的,而且它有一种发育角的潜在能力,我们从老年的或患病的雌者可以看到这种情形。 (584) 再者,鹿的其他一些物种的雌者正常地或偶尔地表现有角的残迹;例如,雌羌鹿(Cervulus moschatus)“具有硬而短的毛簇,其先端形成一个瘤状物,以代替角”;大多数雌美洲赤鹿(Cervus canadensis)的标本表明,“在角的位置上生有尖锐的骨质突起”。 (585) 根据这几种考察结果,我们可以作出结论说,雌驯鹿具有十分发育良好的角,乃是由于雄者最初获得了角作为同其他雄者进行争斗的武器;其次由于某种未知原因,它们在雄者年龄异常小的时候就发育了,结果遂传递给雌雄二者。
现在转来谈谈鞘角反刍动物:关于羚羊,可以形成一个级进的系列,从雌者完全不具角的物种开始——进而到雌者的角小至几乎成为残迹的那些物种[例如叉角羚羊(Antilocapra americana),这个物种在四只或五只中仅有一只具角者 (586) ],再进而到一些物种具有相当发达的角,但显然比雄者的角较小、较细,而且有时角的形状也不同, (587) 最后到达雌雄二者具有相等的角的那些物种为止。对驯鹿来说,同样地对羚羊来说,如上所述,在角的发育时期和角向某一性传递或向雌雄两性传递之间存在着一种关系;所以某些物种的雌者有角或无角以及其他物种的雌者具有较完善状态的角或较不完善的角,并不决定于它们有任何特殊用途,而是简单地决定于遗传。下述情况同这种观点相符合,即,甚至在同一个属中,有些物种的雌雄二者均具角,而另外一些物种唯独雄者具角。还有一个值得注意的事实:印度黑羚(Antilope bezoartica)的雌者通常不具角,但布莱思先生曾看到具角的雌者不少于三只,而且没有理由来假定它们是老的或患病的。
山羊和绵羊的一切野生种,其雄者的角都比雌者的角为大,而且雌者常常完全无角。 (588) 这两种动物的几个家养品种,唯独其雄者具角,还有一些品种,例如北威尔士(North Wales)绵羊,虽然雌雄二者正常都具角,但母羊很容易变得无角。有一位可信赖的目击者在产羔季节有目的地对一群这种羊进行过检查,他告诉我说,羊羔初生时,其雄者的角一般比雌者的角发育得更充分。皮尔(J.Peel)先生曾用雌雄二者永远都具角的隆克(Lonk)绵羊同无角的莱斯特(Leicester)绵羊以及无角的希罗普郡绒毛绵羊(Shropshire Downs)进行杂交;结果是,其雄性后代的角相当地缩小了,同时雌性后代则完全无角。这几个事实表明,母绵羊的角远远不像公绵羊的角那样地是一个十分稳定的性状;这就引导我们相信绵羊的角最初起源于雄者。
关于成年的麝牛,其雄者的角大于雌者的角,而且雌者的角基不相接触。 (589) 至于普通牛,布赖茨先生指出:“在大多数野生牛类中,公牛的角比母牛的牛既大且粗,母爪哇牛(Bos sondaicus) (590) 的角显著地小,而且非常向后倾斜。”关于牛的家养族,无论是隆背的还是不隆背的类型,其公牛的角既短且粗,而母牛和阉牛的牛则比较长而细;关于印度水牛,也是公牛的角既短且粗,而母牛的角比较长而细。关于印度野牛(Bos gaurus),其雄者的角大都比雌者的角既长又粗。 (591) 福塞思·梅杰(Forsyth Major)博士也告诉我说,在瓦尔达诺(Val d'Arno)发现过一个头骨化石,据信这是属于狂野牛(Bos estruscus)这种母牛的,它完全没有角。我再补充一点,雌白独角犀(Rhinoceros simus)的角一般大于雄白犀的角,但不及后者那样有力;另外有些犀牛的物种,据说其雌者的角较短。 (592) 根据这几个事实我们可以推论说,所有种类的角,甚至在雌雄双方同等发育时,大概也是最初由雄者获得,以便战胜其他雄者,而且或多或少完全地传递给了雌者的。
去势的效果值得注意,因为它对上述同一问题的解决投射了光明。公鹿在去势之后,永不重新生角。但雄驯鹿必须除外,因为它在去势后仍然重新生角。这一事实以及雌雄二者均具角的情况,最初一看似乎证明了在这一物种中角并不构成性的特征; (593) 但是,它们是在雌雄体质尚无差异的很幼小年龄中发育的,所以它们不应受到去势的影响,这并没有什么奇怪,即使它们最初是由雄者获得的,也是如此。关于绵羊,雌雄二者正常均具角;我听说雄威尔契绵羊(Welch sheep)的角由于去势而相当地缩小了;但缩小的程度大部分视其实行去势的年龄而定,其他动物的情况也是如此。公美利奴羊具有大角,而母美利奴羊则“一般没有角”;去势对这个品种所产生的作用多少要大些,所以倘在早期实行去势,它们的角“就几乎保持不发育的状态”。 (594) 在几内亚海岸有一个品种,它们的雌羊决不具角,温伍德·里德先生告诉我说,其公羊在去势之后就完全没有角。公牛在去势之后,它们的角就发生很大改变,不再短而粗,而是比母牛的角更长,在其他方面则同母牛的角相似。印度黑羚提供了多少相似的情况:其雄者具有长而直的螺旋形角,二角接近平行,并且向后倾斜;其雌者偶尔具角,但是当这等角出现时,其形状却很不相同,因为它们不是螺旋形的,而且彼此相距甚远,弯曲而角尖向前。那么,正如布赖茨先生告诉我的,在去势的雄性动物中,它们的角就像雌性动物的角那样地具有同样特殊的形状,不过比较长而粗罢了,这是一个值得注意的事实。如果我们可以根据类推法来判断,那么雌者在牛和羚羊这两种场合中大概向我们表明了各个物种的某一早期祖先所具之角的往昔状态。但去势为什么会导致对角的早期状态的重视,目前还不能肯定地加以说明。尽管如此,下述说明似乎还是近理的,即:正如两个不同物种或两个不同族之间的杂交在后代中造成体质的扰乱,因而会导致长久亡失性状的重现, (595) 同样地,由去势在个体体质中所引起的扰乱,也会产生同样的效果。
不同物种或不同族的象的獠牙,依性别不同而有所差异,其情况同反刍类几乎一样。在印度或马六甲,只是雄象具有十分发达的獠牙。大多数博物学者认为锡兰象是一个不同的族,但有些博物学者则认为它们是一个不同的物种,“在100头中未曾发现1头具有獠牙,少数具有獠牙者也都是雄象”。 (596) 非洲象无疑是不同的,它们的雌者具有大而充分发达的獠牙,虽然它们不及雄象的獠牙那样大。
象的几个族和几个物种的獠牙差异——鹿角的巨大变异性,这在野生的驯鹿中表现得尤其显著——黑印度羚(Antilope Bezoartica)的雌者偶尔具角,以及叉角羚羊(Antilocapra americana)的常常不具角——少数一些雄独角鲸具有两个獠牙——有些雌海象完全没有獠牙——都是有关次级性征极端变异性的事例,也是次级性征在亲缘关系密切接近的诸类型中易于出现差异的事例。
虽然獠牙和角在所有场合中似乎是最初作为性武器而发达起来的,但它们常常用于其他目的。象用它的獠牙向虎进攻;按照布鲁斯(Bruce)的材料,它用牙刻截树干,直到容易把它弄倒时为止,它还会用牙把棕榈树的含淀粉的树心取出;非洲象常常使用一只獠牙,而且永远使用这一只,去探查地面是否能承当它的重量。普通公牛用角来保卫其牛群;按照劳埃德的材料,瑞典的驼鹿(elk)用它的大角一下就可以把狼击死。还可以举出许多相似的事实。动物角的第二种最奇妙用途,曾为赫登(Hutton)上尉所见, (597) 即:喜马拉雅角 (Capra aegagrus)的雄者如果不慎自高处跌落,就把头向内弯,以其巨角触地,减轻震荡,据说北山羊(ibex)也会如此。母山羊的角较小,不能做此用,但是,由于母羊的性情比较温和,并不那样迫切需要这种奇怪的防护。
(Capra aegagrus)的雄者如果不慎自高处跌落,就把头向内弯,以其巨角触地,减轻震荡,据说北山羊(ibex)也会如此。母山羊的角较小,不能做此用,但是,由于母羊的性情比较温和,并不那样迫切需要这种奇怪的防护。
每一种雄性动物各以特有的方式来使用它的武器。普通公羊的角基猛撞之力如此强大,以致我曾见到一个强壮的汉子犹如儿童那样被撞翻在地,山羊以及绵羊的某些物种,如阿富汗的圆角盘羊(Ovis cycloceros) (598) ,使其后腿立起,然后不仅猛烈顶撞,“而且以其弯刀形双角的有棱顶尖向下刺入,再猛然向上一拉,就像一把马刀一般。当阿富汗圆角盘羊向一只以好斗闻名的大型家养公羊进攻时,采取了一种全然新奇的战斗方法而获胜,它总是立即接近其敌手,对准其面鼻,用头猛撞,然后在反击来到之前即飘然逸去”。在彭布罗克希尔郡(Pembrokeshire)有一只公山羊,它是一群羊的头羊,这群羊野化已有数代之久,据知它仅在一次战斗中就把几只公羊杀死了;这只公羊具有巨大的角,全长足有39英寸。众所周知,普通公牛用角抵撞和掀挑其敌手;但是,据说意大利水牛从来不用它们的角,而是用它们的凸额对其敌手进行猛击,当后者倒翻在地后,更以膝盖践踏之——这是普通公牛所不具有的一种本能。 (599) 因此,一只狗如果被水牛的鼻子按住,就会立刻被碾成齑粉。然而,我们必须记住,意大利水牛是长期家养的,其野生亲类型肯定不会具有相似的角。巴特利特先生告诉我说,如果把一头母好望角水牛(Bubalus caffer)放进围栏,和一头同种的公水牛生活在一起,这头母水牛就要对公水牛进行攻击,而后者则猛烈地把母水牛推来推去,以为回敬。但是,巴特利特先生明了,如果不是这头公水牛表现了高贵的克制,只要用它的巨角从侧面一击,就可以容易地把那头母水牛杀死。雄长颈鹿的角比雌长颈鹿的角稍长,前者以奇妙的方式使用它的带有茸毛的短角;它向两边摇摆头部,几乎是由上而下,其力至大,我曾看到一块坚硬的木板在它一击之下就出现了深深的刻痕。
羚羊角的形状甚为奇特,其如何使用,有时难以想象;例如南非跳羚(Ant.euchore)的角相当短而直,角尖锐利,向内弯曲,几成直角,彼此相对;巴特利特先生不知道这等角如何使用,但他认为它们可以重创敌手面部的两侧。阿拉伯大羚羊(Oryx leucoryx,图63)的角微向后方弯曲,极长,角尖超出背部的中点,在背部上方几乎成平行线。这样,它们似乎特别不适于相斗;但巴特利特先生告诉我说,当两个这种动物准备相斗时,它们先跪下,置其头于两条前腿之间,当做这种姿势时,它们的角差不多同地面平行,甚为接近,角尖向前直指,稍微向上。于是这两只格斗者彼此逐渐接近,每一只都力图把朝上翘的角尖插入对方的身下;如果有一只成功地做到这一点,它就突然跃起,同时高耸其头,这样,它就会使其敌手负伤,甚至把它戳穿。双方总是跪下,尽可能提防对方的暗算。记载表明,有一只这种羚羊甚至用它的角有效地敌住了一头狮子;然而,为了使角尖指向前方,它不得不把头置于前腿之间,这样,当受到任何其他动物攻击时,一般就要处于非常劣势。因此,它们的角变为现在这样的巨大长度及其特殊位置,大概不是为了保护自己以防御猛兽之用的。可是,我们可以知道,一旦阿拉伯大羚羊的某一古代雄性祖先获得了适度的角长时,它大概就会在同其雄性对手的战斗中被迫把头略微向下朝内弯曲,就像某些雄鹿现在的情形那样;于是它大概最初偶尔获得了跪下的习性,以后便成为一种固定的习性,这并非是不可能的。在这种场合中,几乎肯定的是,双角最长的雄者比双角较短的雄者占有巨大优势;于是它们的角通过性选择就会逐渐地变得越来越长,终于获得它们现在这样的异常长度和位置。
图 63 阿拉伯大羚羊(Oryx leucoryx),雄鹿
(引自Knowsley Menagerie)
许多种类的雄鹿具有分枝的角,这是一个难以解释的奇特事例;因为单独一个直角尖肯定远比几个分歧的角尖更能造成严重的创伤。在菲利浦·埃格顿(Philip Egerton)爵士的博物馆内陈列着一具马鹿的角,长30英寸,其上“不少于15个分枝”;在莫里茨堡(Moritzburg)仍然保存有一对马鹿的角,是弗雷德里克一世(Frederick I.)于1699年射杀的,其中一只角的分枝数令人吃惊,竟达33个,另一只角的分枝为27个,二者合计为60个分枝。理查森绘制过野生驯鹿的一对角,共有29个角尖。 (600) 根据鹿角的分枝形式,特别是根据诸鹿相斗偶尔用前足相踢的情况, (601) 贝利(M.Bailly)实际上作出的结论不是说鹿角害多于利吗?但是,这位作者忽略了竞争的雄鹿所进行的猛烈战斗。关于分枝角的用途或利益,我感到十分困惑,于是我向科隆塞(Colonsay)的麦克尼尔(MacNeill)请教,他曾长期细心地观察过赤鹿的习性,他告诉我说,他从来没有见过鹿角的分枝有什么用途,不过额前的分枝由于向下倾斜,对于保护前额大有裨益,其角尖同样也可用于攻击。菲利浦·埃格顿爵士也告诉我说,马鹿和粘鹿(fallow-deer)在争斗时彼此突然猛撞,以角尽力抵住对方的身体,拼命相斗。当一方被迫屈服并后退时,胜利者便尽力用它的额前分枝角刺人被击败的对手。这样,上部的分枝角似乎主要地或完全地用于相推或相刺。尽管如此,还有几个物种,其上部分枝角是作为进攻武器之用的;在卡顿的渥太华猎园中,有一人受到加拿大马鹿(Wapitideer,cervus canadensis)的攻击,当数人前往救援时,那头雄鹿“决不从地面上把头抬起,事实上其面部几与地平,其鼻差不多处于二前足之间,但是当它窥测新的冲刺方向时,就把头转向一边。”当做这种姿势时,角尖便直对敌方。“它必须把头稍微抬起,才能转动它,因为它的角特长,如果不把头在一边抬起,就无法转动,同时在另一边它的角已触及地面。”这只公鹿用这种方法把一群前来救援的人逐渐赶到150~200英尺以外,而受攻击的那个人终于被弄死。 (602)

图 64 库杜捻角羚(Strepsiceros kudu)
(引自安德鲁·史密斯爵士的《南非洲动物学》)
鹿角虽是有效的武器,但我以为单独一个角尖无疑要比分枝角危险得多;对鹿类具有丰富经验的卡顿完全同意这个结论。分枝角对于防御其他竞争的雄鹿虽是高度重要的手段,但它容易纠结难分,看来也并不十分完善地适于这种目的。于是我猜想它们的角也许部分地作为装饰之用。鹿的分枝角以及某些羚羊的优美竖琴状的角呈双重弯曲(图64),在我们眼里都具有装饰性,任何人对这一点都不会有争论。如果它们的角有如古代骑士的华丽装备,可以增添鹿和羚羊的高贵风采,它们可能部分地为此目的而发生变异,虽然其实际用途主要还在于争斗;不过我没有掌握有利于这一见解的证据。
最近发表的一个有趣事例表明,在美国的某一地方有一只鹿,它的角目前正在通过性选择和自然选择进行变异。一位作者在一份最优秀的美国杂志 (603) 上写道,晚近21年来他都在阿迪隆达克斯(Adirondacks)行猎,那里盛产弗吉尼亚鹿(Cervus virginianus)。约在十四年前,他最初听说有一种钉状角的雄鹿(spike-horn bucks)。这种鹿逐年增多;约在五年前,他射得一只,以后又射得一只,而现在射得的就很多了。“钉状角同弗吉尼亚鹿的普通角大不相同。它是一个单独的钉状物,比分枝角为细,长不及分枝角的一半,自额部突向前方,末端锐利。具有这种角的雄鹿比普通雄鹿占有相当的优势。这种钉状角可以使鹿更迅速地穿过茂密的森林和矮树丛(每一个猎人都知道雌鹿和一周岁的雄鹿远比具有笨重分枝角的大型雄鹿跑得快得多),除此之外,钉状角同普通角相比,还是一种更有效的武器。具有钉状角的雄鹿由于占有这种优势,就会胜过普通雄鹿,总有一天前者在阿迪隆达克斯可以完全取代后者。毫无疑问,具有钉状角的雄鹿的最初出现,不过是一种偶然的反常现象而已。但这种钉状角给予它一种优势,而且使它可以传续这种特性。其后裔具有同样的优势,而且以稳定的增长率来传续这种特性,终于它们会慢慢地把具有分枝角的鹿排挤出它们所栖息的地域之外。”一位批评家对这种说法提出十分有力的异议,问道:如果说单角现今如此有利,那么祖代类型的分枝角为何能够发达?对此我的回答只能是,利用新武器实行新式攻击大概是最有利的,上述阿富汗圆角绵羊的例子阐明了这一点,它就是这样战胜了一只以其战斗力闻名的家养公羊。如果一只公鹿只是和同一种类的其他公鹿相斗,那么它的分枝角虽然十分适于同其竞争对手相斗,而且慢慢获得长而分枝的角虽然对叉角变种有利,但决不能因此就说分枝角最适于战胜具有不同武装的敌手。上述瞪羚的例子表明,如果它只和同一种类的竞争对手相斗,它的角越长大概就越有利,但若遇到一种短角羚羊而无须跪下者,则胜利几乎肯定要归于这种羚羊。
具有獠牙的雄性四足兽,以各种方式使用它们,正如角的使用情况一样。公野猪用其獠牙进行侧击和向上挑;麝 (604) 以其獠牙向下刺,均可给其敌手以重创。 (605) 海象的颈部虽很短,体部虽很笨拙,却能同等敏捷地从上方、下方以及侧面进击。 (606) 已故的福尔克纳博士(Falconer)告诉我说,印度象按其獠牙的位置和曲度而采取不同的争斗方式。如果它的獠牙直插前方而且向上,它就能把一只虎抛掷甚远——据说可至30英尺;如果它的獠牙短而且向下,它就会突然地尽力把虎压在地上,这样,对乘象人是危险的,因为容易把他掷出象轿(howdah)之外。 (607)
很少四足兽具有特别适于同雄性对手进行战斗的两种不同武器。然而雄吠麂(Cervulus,muntjac deer) (608) 提供了一个例外,因为它既有角,又有突出的犬齿。但我们可以从下述事实推论一种类型的武器随着岁月的推移,可以代替另一种类型的武器。关于反刍动物,角的发达甚至同中等发达的犬齿一般处于相反的关系。例如,骆驼、红褐色美洲羊驼(guanacoes)、鼷鹿(chevrotains)和麝均无角,但有有效的犬齿;“雌者的这等犬齿永远小于雄者的”。骆驼科(Camelidae)除了具有真正的犬齿以外,在上颚还有一对犬齿形状的切齿。 (609) 另一方面,雄鹿和雄羚羊都有角,它们却很少有犬齿;如有犬齿,也总是小型的,所以它们在战斗中究竟有何作用是可以怀疑的。在蒙大拿山羚羊(Antilope montana)中,其幼小雄者只有犬牙的残迹,当它成长以后,犬齿即行消失;而所有年龄的雌者都不具犬齿;但某些其他种类的羚羊和鹿据知偶尔也有犬齿的残迹。 (610) 公马有小型的犬齿,母马完全没有犬齿或仅有其残迹;但这等犬齿似乎并不用于战斗,因为公马用切牙咬啮,而且不像骆驼和红褐色美洲羊驼那样地可以把嘴张大。如果成年雄者具有犬齿,现已无效,同时雌者没有犬齿或仅具其残迹,我们就可断言,这个物种的早期雄性祖先具有有效的犬齿,而且部分地传递给了雌者。雄者的这等犬齿的缩小,似乎是由于其战斗方式发生了某种变化所致(但马的情况并非如此),而这种变化乃是由新武器的发达所引起的。
獠牙和角对其拥有者显然具有高度重要性,因为它们的发育要消耗大量的有机物质。亚洲象——一个绝灭的多毛物种的一个獠牙以及非洲象的一个獠牙据知各重150、160和180磅;有些作者所记载的重量还要大。 (611) 鹿的角定期地更新,这在体质消耗上一定更大;驼鹿(moose)的角重达50~60磅,还有一种绝灭的爱尔兰驼鹿,它们的角重达60~70磅——而后者的头骨平均仅重5.25磅。绵羊的角虽不定期地更新,但许多农学家们认为它们的发达会给饲养主造成明显的损失。再者,公鹿当逃避猛兽的追击时,其角重有碍它的奔驰,而且大大减弱其穿过树林的速度。例如,驼鹿角两个顶端之距为5.5英尺,虽然它们在安步行走时,能够如此灵巧地运用它们的角,以致不会碰到或折断一个树枝,但当迅速逃避一群狼时,就不能那样灵巧地适用它们的角了。“当它前进时,高举其鼻,以便把角向后放在水平的位置;而做这种姿势时,就无法清楚地看到地面了。” (612) 大型爱尔兰驼鹿的两个角端相距实际上竟达8英尺!当角上被以茸毛时,这在赤鹿要持续20周左右,它们极易受伤,所以在德国这时公鹿的习性多少有些变化,它们避开茂密的森林,往来于幼树和低矮灌木之间。 (613) 这些事实会使我们想起,雄鸟获得装饰性的羽毛是以飞翔受到阻碍为代价的,而获得其他装饰物则以损害它同雄性对手相斗的力量为代价的。
关于哺乳动物,正如情况所常常表明的那样,雌雄二者大小不同,雄者几乎永远比雌者大而强。古尔德先生告诉我说,澳大利亚有袋类的这种情况也是显著的,它们的雄者直到异常老的年龄还继续生长不已。但是,一个最特殊的例子还是由一种海狗(Callorhinus ursinus) (614) 提供的,其充分成长的雌者在重量上小于充分成长的雄者六分之一。 (615) 吉尔博士说,众所周知,多配偶的海狗,其雄者彼此相斗非常剧烈,而且雌雄二者在体型上差别很大;单配偶的物种则差别很小。鲸类也提供了证据,表明雄者的好斗性同其体型有一定关系,好斗鲸类的雄者在体型上大于雌者;雄露脊鲸(right-whales)彼此不相斗,它们不但不大于雌者,反而较小;相反,雄巨头鲸彼此激烈相斗,它们的身体“往往有其对手牙齿所造成的伤痕”,其雄者的体型为雌者的两倍。雄者的强大力气,正如亨特很久以前所指出的 (616) ,永远表现在同其他雄性对手战斗时所使用的那些身体部分——例如公牛的粗壮颈部。雄性四足兽也比雌者更为勇敢、更为好斗。毫无疑问,这等特性的获得,一部分是通过性选择,这是由于较强的和较勇敢的雄者对较弱的雄者取得了一系列的胜利所致,一部分则是通过使用的遗传效果。在体力、大小以及勇气方面的连续变异无论是起于单纯的变异性,还是起于使用的效果,雄性四足兽都是借着连续变异的积累而获得了在生命晚期出现的这等特性,因而这等特性在很大程度上大概只限于传递给同一性别。
根据上述若干考察,我急于得到有关苏格兰猎鹿狗的材料,因为其雌雄二者在体型上的差异大于任何其他品种(虽然嗅血猎狗 (617) 的雌雄差异也相当大),也大于我所知道的任何野生犬种。因此,我向卡波勒斯先生请教,他以成功地驯养这个品种而闻名于世,他曾对自己养的那些狗进行过称重和度量,蒙他盛情相助,为我从各种来源收集了下述事实。优良的公苏格兰猎鹿狗,其肩高从低者28英寸至33英寸、甚至34英寸;其重量从轻者80磅至120磅或更多。母苏格兰猎鹿狗的高度从23英寸至27英寸,甚至28英寸;其重量从50磅至70磅、甚至80磅。 (618) 卡波勒斯断言,公猎鹿狗的重量从95至100磅,母猎鹿狗的重量70磅,大概是一个可靠的平均数;但有理由相信,雌雄二者在往昔都曾达到过更大的重量。卡波勒斯先生曾对降生后两周的小狗进行过称重,一胎中四只小公狗的平均重量超出两只小母狗的平均重量6.5盎司;在另一胎中四只小公狗的平均重量超出一只小母狗的重量不到1盎司;长到三周时,这等小公狗的平均重量超出那只小母狗7.5盎司,长到六周时,差不多超出14盎司。赖特先生在给卡波勒斯先生的一封信中写道:“我曾对许多胎小狗的大小和重量做过记录,就我经验所得,按照一般规律,小公狗和小母狗的重量在5~6个月之前差异很小;此后小公狗即开始增大,无论在大小方面或重量方面都超过小母狗。在降生时或降生后数周之内,小母狗偶尔大于小公狗,但最终一定要被小公狗所超过。”科隆塞的麦克尼尔(McNeill)先生断言,“公狗不超出两岁不会达到充分成长的状态,虽然母狗达到这种状态要早些。”按照卡波勒斯(Cupples)先生的经验,公狗直到12~18个月的时候还在身高方面继续增长,在重量方面直到18~24个月还继续增长;而母狗一到9~14个月或15个月的时候在身高方面就停止生长,在重量方面则到12~15个月的时候停止增长。根据这几种记载,苏格兰猎鹿狗(Scotch deerhound)不到生命的相当晚期,其雌雄二者在大小方面显然不会获得充分的差异。用于追猎的几乎完全是公狗,因为,正如麦克尼尔告诉我的,母狗没有足够的体力和重量来推倒一只充分成长的鹿。我听卡波勒斯先生说,在远古时代的传说中,公狗最负盛名,而母狗仅作为有名公狗的母亲而被提及。因此,许多世代以来在体力、大小、速度以及勇气方面受到测验的主要是公狗,而且选其最优良者为传种之用。由于公狗不到生命的相当晚期不会达到其充分大小,所以按照常常提出的那条规律来看,它们倾向于把其特性只传给雄性后代;这样,苏格兰猎鹿狗雌雄之间的大小极不相等大概就可以得到解释了。
有些少数四足类,其雄者具有专为对付其他雄者进攻的器官或部分。某些种类的鹿,就像我们已经见到的那样,主要地或完全地使用它们角的上部分枝来防卫自己;正如巴特利特先生告诉我的,瞪羚用其微微弯曲的长角非常巧妙地进行防卫;但是这等角同样地也可作为攻击器官来使用。同一位观察家说道,犀类在相斗时彼此用它们的角挡开对方的侧击,咔嗒咔嗒地作响,其声甚高,就像公野猪使用獠牙时的情况那样。虽然公野猪彼此拼命相斗,但是,按照布雷姆的材料,它们很少负重伤,这是由于彼此的打击皆落在獠牙之上,或者落在那层遮盖肩部的软骨般的皮肤之上,德国猎人把这块皮肤叫做盾;在这里我们看到了一个身体部分专门为了防卫而发生了改变。公野猪在壮年时期的下颚獠牙是用于战斗的(参阅图65),但是,正如布雷姆所述,到了老年,这等獠牙向内和向上弯曲得如此厉害,甚至高过鼻部,所以不再能用于战斗了。然而,它们仍然可能作为防御的手段,甚至更为有效。为了补偿下颚獠牙不能再作武器的损失,一向从两侧稍微向外突出的上颚獠牙在年老时便大大增加了其长度,并且向上弯曲得很厉害,因而它们也可用于攻击。尽管如此,一头老龄的公野猪对人来说,就不像六七岁的公野猪那样危险了。 (619)

图 65 壮年期公野猪的头
图 66 东南亚疣猪(Babirusa)的头骨
(采自华莱士的《马来群岛》)
西里伯斯产的充分成长的雄东南亚疣猪(Babirusa pig,图66),其下颚獠牙是可怕的武器,就像欧洲野猪在壮年时期的獠牙那样,然而,其上颚獠牙如此之长并牙尖向内弯曲如此之甚,有时甚至弯及额部,以致完全不能作为进攻武器之用。与其说它们是牙,倒不如说它们很像角,它们显然不能作为牙用,所以从前设想这种动物是把头部挂在树枝上面来休息的!如果把头部稍微侧向一方,上颚獠牙的凸面大概可以作为最好的防御器;因此,老龄东南亚疣猪的这等獠牙“一般都是折断的,好像就是由于争斗所致”。 (620) 于是在这里我们看到了一个奇妙的事例:东南亚疣猪的上颚獠牙在壮年时期所正常呈现的构造显然只适于用做防御;而欧洲公野猪的下部獠牙只在老年时期具有差不多同样的形状,唯其程度较轻,这时它们才以同样的方式用做防御。
雄疣猪(Phacochoerus aethiopicus,图67)在壮年时期的上颚獠牙向上弯曲,而且尖锐,是一种可怕的武器。其下颚獠牙比上颚獠牙尤其锐利,但很短,所以它几乎不能用做攻击的武器。然而它们同上颚獠牙的根部密切相合,作为它们的基础,所以它们一定可以大大加强上颚獠牙的力量。无论上颚獠牙或下颚獠牙似乎都没有为了防卫而发生特别变异,虽然它们在一定程度上无疑是用于这一目的的。但是,疣猪并不缺少其他特别的防御手段,它在面部两侧的眼睛下方各有一块软骨性的椭圆形护垫,与其说它是坚硬的,不如说是韧性的,而且向外突出二三英寸;当看到这种活的动物时,巴特利特先生和我都以为,当其敌手用獠牙从下方进行攻击时,这等护垫大概就会向上翻起,于是就可以极好地保护其多少突出的眼睛。根据巴特利特先生的权威材料,我还可以补充一点:这等公野猪在战斗时直接面对面而立。
图 67 母疣猪的头部
(引自《动物学会会报》(1869年);图示具有公疣猪的同样性状,虽然其程度较差)
附注:当此图最初刻成时,我还以为它是雄的。
最后,非洲河猪(Potomochoerus penicillatus)在面部两侧的眼睛下方各有一个软骨性的硬瘤,同疣猪的韧性护垫相当;它在上颚还有两个骨质突起,位于鼻部的上方。在伦敦动物园里,一只这种公野猪最近弄坏了疣猪的围栏,躜人其中。它们彻夜相斗,至晨双方疲惫不堪,但均未负重伤。上述护垫和瘤状物满布血迹,其上有非常严重的戳伤和擦伤;这是一个重要的事实,因为它阐明了这等护垫和瘤状物的用途。
虽然猪科很多成员的雄者具有武器,而且像我们刚才看到的,还具有防御手段,但这等武器似乎在较晚的地质时期内才获得的。福赛思·梅杰博士 (621) 列举了几个中新世的物种,其中没有一个物种的雄者的獠牙似乎是非常发达的;卢特迈耶教授以前也曾被这一事实所打动。
雄狮的鬃毛对敌对雄狮的攻击是一种良好的防御,这是它容易遇到的一种危险,因为,正如史密斯爵士告诉我的,雄狮之间进行极其猛烈的争斗,幼狮不敢接近老狮。1857年,布拉米奇(Bromwich)的一只虎弄坏了一只狮子的围栏,躜入其中,于是一个可怕的场面出现了:“狮子的颈部和头部由于受到鬃毛的保护,未受重伤,但那只虎终于把狮子的腹部撕裂,数分钟后即行死去。” (622) 加拿大山猫(Felis canadensis)喉部和颏部周围的丛毛,在雄者要比在雌者长得多;但这种丛毛是否用于防御,我不知道。众所熟知,雄海豹(seals)彼此拼命相斗,其某些种类如鬃海狗 (623) (Otaria jubata) (624) 的雄者具有长鬃,而雌者的鬃却很短或根本没有。好望角的雄鼯猴(Cynocephalus porcarius)的鬃和犬齿要比雌者的大得多;其鬃大概作为保护之用,因为,我曾问过伦敦动物园的管理员,为了不暴露我提问的目的,我说,是否有任何种类的猴专向颈背攻击,我得到的答复是,除了上述狒狒之外,都不如此。爱伦堡(Ehrenberg)曾把成长的雄阿拉伯狒狒(Hamadryas baboon)的鬃比做幼狮的鬃,幼小的雌雄狒狒以及成长的雌狒狒几乎都没有鬃。
雄美洲野牛(bison)的羊毛状的巨鬃,长几及地,而且雄者的鬃远比雌者的发达,我以为这种鬃大概是在剧烈的战斗中作为保护之用的;但一位有经验的猎人告诉卡顿说,他从来没有见过任何可以支持这种信念的事实。雄马的鬃比雌马的茂密;我曾特别询问过两位经验丰富的驯马人和养马人,他们都曾管理过许多马群,皆确言“它们永远力图咬住对方的颈部”。然而,这并不是说,当颈毛作为保护之用时,在最初就是为了这个目的而发达起来的,虽然在某些场合中可能是这样,如在狮子的场合中就是如此。麦克尼尔告诉我说,雄马鹿(Cervus elaphus)喉部的长毛当被猎逐时,可以起重大的保护作用;但这等长毛并不见得是专门为了这个目的而发达起来的;否则,幼者和雌者也会有同等的保护。
四足兽的雌雄任何一方对配偶的选择
在下章对雌雄二者在发声、气味以及装饰物等方面所表现的差异进行讨论之前,在这里先考察一下雌雄二者当结合时是否实行选择,将会有某些方便。在雄者争夺雄性霸权之前或在此之后,雌者是否会挑选任何特殊的雄者;或者,雄者如果不是多配偶的,是否会选择任何特殊的雌者?育种家的一般印象似乎是,雄者可以接受任何雌者,这是由于雄者对雌者的热切追求,在大多数场合中,大概确是如此。作为一般规律,雌者是否毫无差别地接受任何雄者,则是一个大得多的疑问。在第十四章中已经指出,关于鸟类,有大量的直接和间接的证据阐明了雌者对其配偶是实行选择的;那么,位于较高等级并且具有较高心理能力的雌四足兽如果不一般地、或者至少常常地实行某种选择,那大概是一种奇怪的反常现象。在大多数场合中,如果一个不能取悦于雌者或者不能使它激动的雄者来求偶时,这个雌者就要逃去;如果一个雌者受到几个雄者追求,如普通发生的情形那样,当雄者彼此争斗时,这个雌者往往有机会同一个雄者逃走,或者至少同其临时交配。在苏格兰常常可以看到雌赤鹿有后面这种情形,这是菲利普·埃格顿(Philip Egerton)爵士以及其他人士向我说过的。 (625)
关于自然状况下的雌四足兽在婚配时是否实行任何选择,几乎不可能知道的很多。勃兰特上尉有充分的机会对海狗(Callorhinus ursinus)进行观察,下述有关一只海狗求偶的奇妙细节,就是根据这位人士的权威材料。 (626) 他说,“许多雌海狗当到达其进行繁育的岛屿时,好像渴望依附某一特定的雄者,爬上外围的岩石,眺望全群,发出呼叫,并似乎倾听那熟悉的声音。于是更换另一地点,重复同样的动作……当有一只雌者一到海岸,最相近的一只雄者就从上方下来同雌者相会,同时发出一种喧嚣声,就像母鸡呼唤其雏鸡一般。雄者向它点头弯腰,进行哄诱,直到它处于雌者和水之间,所以它无法再避开它。于是雄者的态度为之一变,厉声吼叫,把它赶到其‘妻妾’所在的地方。然后又继续这样进行,直到其‘妻妾’所在之地差不多充满时为止。于是诸雄者登上较高处所,窥伺时机,当其更为幸运的邻居疏于防范时,即行窃取其‘妻妾’。当进行窃取时,它们把雌者叼在嘴中,高高举起,超出其他雌者的头部之上,小心谨慎地把它们放置在自己的‘妻妾,之间,就像老猫携带小猫那样。居于更高处的雄者也按同法为之,直到整个场所充满雌者而后已。为了占有同一只雌者,两只雄者之间屡屡发生争斗,双方同时咬住这只雌者,以致撕裂为二,或者由于咬啮而受到重伤。当整个场所充满雌者时,老年雄者洋洋得意地巡阅其家族,申斥那些拥挤或打扰其他雌者的分子,而且凶猛地把一切入侵者赶跑。进行这样监视经常使自己忙碌不堪。”
关于自然状况下的动物的求偶,所知者非常之少,因此我曾努力发现家养四足兽在交配时所实行的选择会达到怎样程度。犬类提供了最好的观察机会,因为它们受到了细心的照顾,而且对它们可以充分理解。许多育种家对这一问题表示了强有力的意见。例如,梅休(Mayhew)先生说:“母狗能够施给爱情,温柔的回忆对它有强烈的影响,就像我们所知道的比它更为高等的动物在其他场合里所表现的那样。母狗在爱情方面并非总是那么持重,而是容易委身于低等的杂种狗。若把母狗同外貌卑劣者同育一处,则在这一对配偶间常常会发生热爱,此后就永远不能制止。这种热爱是真实的,并非浪漫主义的,所以能够持久。”梅休先生所观察的主要是小型品种,他相信大型公狗对小型母狗有强烈的吸引力。 (627) 著名的兽医布莱恩(Blaine)说道 (628) ,他自己养的一条母哈巴狗(pug)非常热情地爱上了一只长毛垂耳狗(spaniel),还有一只母谍犬(setter)也非常热情地爱上了一只杂种狗,以致经过几个星期之后,它们才同自己的品种交配。我曾收到同样的而且可以信赖的两项记载,表明一只母拾物猎狗(retrieverr)和一只母长毛垂耳狗都爱上了 (terrier)。 (629)
(terrier)。 (629)
卡波勒斯先生告诉我说,他可以亲自保证下述更为显著的事例是确实的,即,一只贵重的、异常聪明的雌 爱上了邻居的一只长毛垂耳狗,它竟爱到这样程度,以致势必常常把它拖走,才能离开那只公狗。把它永久隔离之后,它的乳头虽然屡现乳汁,但决不接受任何其他公狗的求爱,因而终生没有生仔,它的主人对此甚为遗憾。卡波勒斯先生还说,在他的狗窝中有一只母猎鹿狗(deerhound),1868年曾三次产仔,同窝还有四只公猎鹿狗,每次母狗都对其中一只身体最大的、长相最漂亮的公狗表现了最显著的爱好,所有这四只公狗均在壮年。卡波勒斯先生观察到,母狗一般喜爱和它有过交往而相识的公狗;母狗的腼腆和怯懦最初使其倾向于拒绝陌生的公狗。相反,公狗却似乎倾向于陌生的母狗。公狗拒绝任何特定的母狗,大概是罕见的,但是,一位著名的狗育种家——耶尔德斯雷俱乐部(Yeldersley House)的赖特(Wright)先生告诉我说,他知道有关这方面的一些事例;他举出自己饲养的一只公猎鹿狗为例,它对任何特定的母獒(mastiff)都不屑一顾,所以势必使用另一只公猎鹿狗才行。再举我所知道的其他事例大概是多余的,我只补充一点;巴尔(Barr)先生细心地繁育过许多嗅血猎狗,他说,几乎在每一个事例中,公狗或母狗的特定个体都彼此表现了一种明显的爱好。最后,卡波勒斯先生又一年研究了这个问题之后,写信向我说,“我可以充分证明我以往的叙述,即,狗在繁育时彼此均表现有明显的爱好,这往往受到体型大小、毛色鲜明以及个性的影响,同时也受到以往彼此熟识程度的影响。”
爱上了邻居的一只长毛垂耳狗,它竟爱到这样程度,以致势必常常把它拖走,才能离开那只公狗。把它永久隔离之后,它的乳头虽然屡现乳汁,但决不接受任何其他公狗的求爱,因而终生没有生仔,它的主人对此甚为遗憾。卡波勒斯先生还说,在他的狗窝中有一只母猎鹿狗(deerhound),1868年曾三次产仔,同窝还有四只公猎鹿狗,每次母狗都对其中一只身体最大的、长相最漂亮的公狗表现了最显著的爱好,所有这四只公狗均在壮年。卡波勒斯先生观察到,母狗一般喜爱和它有过交往而相识的公狗;母狗的腼腆和怯懦最初使其倾向于拒绝陌生的公狗。相反,公狗却似乎倾向于陌生的母狗。公狗拒绝任何特定的母狗,大概是罕见的,但是,一位著名的狗育种家——耶尔德斯雷俱乐部(Yeldersley House)的赖特(Wright)先生告诉我说,他知道有关这方面的一些事例;他举出自己饲养的一只公猎鹿狗为例,它对任何特定的母獒(mastiff)都不屑一顾,所以势必使用另一只公猎鹿狗才行。再举我所知道的其他事例大概是多余的,我只补充一点;巴尔(Barr)先生细心地繁育过许多嗅血猎狗,他说,几乎在每一个事例中,公狗或母狗的特定个体都彼此表现了一种明显的爱好。最后,卡波勒斯先生又一年研究了这个问题之后,写信向我说,“我可以充分证明我以往的叙述,即,狗在繁育时彼此均表现有明显的爱好,这往往受到体型大小、毛色鲜明以及个性的影响,同时也受到以往彼此熟识程度的影响。”
就马来说,世界上最伟大的竞赛跑马育种家布伦基隆(Blenkiron)先生告诉我说,种马在其选择上如此屡屡反复无常,没有任何明显的原因,就拒绝某一母马而就另一母马,以致必须惯常地对它施用各种诡计才行。例如,著名的公“大王”马(Monarque)决不会有意识地对母“斗士”马(Gladiateur)看上一眼,因而势必施以诡计。对贵重的种竞塞马的需求如此之大,以致会把它弄得筋疲力尽,我们知道这就是这类种马为什么在其选择上那样苛求的部分原因。布伦基隆先生从来不知道母马会拒绝公马;但在赖特先生的马厩中就曾发生过这种情形,所以势必对这匹母马进行欺骗。卢卡斯 (630) 引用过各种法国权威人士的论述,他说,“确有公马特别选取一定的母马,而对其他一切母马一概拒绝”。他根据贝伦(Ba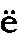 len)的权威材料举出过有关公牛的相似事实;里克斯(H.Reeks)先生向我保证说,他父亲有一头公短角牛,“永远拒绝同一头黑母牛交配”。霍夫勃格在描述家养的驯鹿时说道:“母鹿似乎很喜爱大型而强壮的公鹿,避开幼小的以及壮年的鹿,于是那只公鹿将把诸幼鹿驱散。” (631) 有一位传教士繁育过许多猪,他断言母猪往往拒绝某一头公猪,而立即接受另一头公猪。
len)的权威材料举出过有关公牛的相似事实;里克斯(H.Reeks)先生向我保证说,他父亲有一头公短角牛,“永远拒绝同一头黑母牛交配”。霍夫勃格在描述家养的驯鹿时说道:“母鹿似乎很喜爱大型而强壮的公鹿,避开幼小的以及壮年的鹿,于是那只公鹿将把诸幼鹿驱散。” (631) 有一位传教士繁育过许多猪,他断言母猪往往拒绝某一头公猪,而立即接受另一头公猪。
根据上述事实可以断言,家养四足兽常常表现有强烈的独特反感和独特爱好,在这方面雌者甚于雄者更为常见。既然如此,则处于自然状况下的四足兽的交配大概不可能仅仅委于偶然。远为可能的是,雌者被特殊雄者所诱惑或引起雌者的激情,这等雄者比其他雄者具有某种较高程度的特性;但这等特性是什么,我们很少能够或者永远不能够确切地发现。
第十八章 哺乳类的第二性征(续)
声音——海豹的显著雌雄特性——气味——毛的发达——毛和皮肤的颜色——雌类比雄类装饰得更美的反常事例——由于性选择而发生的体色和装饰物——为了保护的目的而获得的体色——虽为雌雄二者所共有的体色,也往往是由于性选择而发生的——关于成年四足兽的斑点和条纹的消失——关于四手目的体色和装饰物——提要
四足兽使用它们的声音有各种不同目的,或作为危险的信号,或作为兽群中某一成员对另一成员的呼唤,或系母兽对亡失仔兽的召唤,或系仔兽呼唤母兽来保护自己;但对这等用途均无须在此讨论。我们现在所要讨论的仅仅是雌雄二者之间的声音差异,例如,雄狮和雌狮、公牛和母牛的声音差异。几乎所有雄性动物在发情季节远比在任何其他时期更多使用它们的声音;有些动物,例如长颈鹿和豪猪 (632) 除了在发情季节以外据说是完全哑的。比如喉部(即喉头和甲状腺 (633) )在繁殖季节开始时定期地肥大,因而可以设想它们强有力的声音由于某种未知原因对它们一定是高度重要的;但这一点非常可疑。根据二位有经验的观察家麦克尼尔先生和埃格顿爵士向我提供的材料,三岁以下的幼鹿似乎并不鸣叫;老鹿在繁殖季节开始时才开始鸣叫,当它们到处不停地漫游去寻求雌鹿时,最初仅偶尔一鸣,其声低沉。当雄鹿进行战斗之前,则大声鸣叫,而且声音拖长,但在实际冲突中,则毫不出声。惯于使用声音的所有种类的动物当处于任何强烈的感情之中时都会发出各种不同的噪声,例如当激怒和准备战斗时就会如此;但这可能只是神经兴奋的结果,于是引起身体的几乎所有肌肉的痉挛收缩,例如当一个人在愤怒时的咬牙切齿和紧握双拳就是如此。毫无疑问,雄鹿以其鸣叫挑起彼此进行殊死的战斗;但是,具有比较强有力声音的那些雄鹿,除非同时也是更强壮的,具有更好武装的,而且更加勇敢的,否则就不会比其敌对者占有任何优势。
狮子的吼叫在威吓其敌对者方面可能对它有某种帮助;因为当它发怒时,同时也把其鬃毛竖起,这样就本能地使自己尽量显得可怕。但是,几乎不能假定,公鹿的鸣叫即使在这方面有所帮助,就会导致其喉部的定期扩大。有些作者提出,雄鹿的鸣叫系用于召唤雌鹿;但上述两位富有经验的观察家告诉我说,虽然雄鹿热切地寻求雌鹿,但雌鹿并不寻求雄鹿,根据我们所知道的其他四足兽习性来说,可以预料情况确系如此。另一方面,雌鹿的鸣声可以把一头或更多的雄鹿很快引到自己的身旁, (634) 猎人们清楚地晓得这一点,所以他们在野外模仿雌鹿的鸣声。如果我们能够相信雄鹿有用声音使雌鹿激动或媚惑雌鹿们的能力,那么根据性选择原理以及受到同一性别和季节所限制的遗传原理,雄鹿发音器官的定期肥大就是可以理解的了;但我们还没有证据来支持这一观点。按照其情况来看,雄鹿在繁殖季节的高声鸣叫,无论在求偶期间,或在战斗期间,或在任何其他方面,对它来说似乎都没有任何特殊用途。但是,在强烈的爱情、嫉妒以及愤怒中屡屡使用其声音,并连续许多世代如此为之,我们能够不相信这对于雄鹿乃至对于其他雄性动物的发音器官最终会产生一种遗传的效果吗?在我们现今的知识状况下,我以为这是最近理的一种观点。
成年雄大猩猩的叫声非常洪大,它具有一种喉囊,就像雄猩猩那样。 (635) 长臂猿为猿类中的最喧嚣者,而且苏门答腊合趾长臂猿(Hylobates syndactylus)也具有气囊;但是,曾有机会对其进行过观察的布莱恩先生并不相信其雄者比雌者更为喧嚣。因此,合趾长臂猿的叫声大概用做相互召唤,有些四足兽类,例如河狸,肯定如此。 (636) 敏捷长臂猿(H.agilis)之所以著名,是由于它有发出完全而准确的八度音阶的能力, (637) 我们可以合理地推测这是用于性的媚惑;但在下一章,我势必还要谈到这个问题。美洲卡拉亚吼猴(Mycetes caraya),其雄者的发音器官大于雌者的三分之一,而且非常强有力。这种猴在温暖的天气里使树林朝夕充满着其压倒一切的叫声。雄猴开始其可怕的合唱,常常延续许多小时,雌猴有时也参加,但其吼叫的力量较小。一位优秀的观察家伦格尔(Rengger) (638) 未能发现这是由任何特殊原因所激起的,他以为,就像许多鸟类那样,它们也喜欢自己的音乐,而且彼此争胜。上述大多数猿类之所以获得其强有力的叫声,是否为了击败其敌对者并向雌者献媚——或者,这等发音器官通过长期连续使用的遗传效果而被加强和增大,是否没有因此而获得任何特殊利益——我不敢说;但上述观点,至少在敏捷长臂猿的场合中似乎是最近理的。
我愿提一提海豹类所具有的两种很奇妙的雌雄特性,因为有些作者设想它们对声音有影响。雄象海豹(Macrorhinus proboscideus)的鼻子大大增长,并且能够竖起。在这种状态下有时长达一英尺。雌象海豹在生命的任何时期都不如此。雄象海豹发出一种狂热的、嘶哑的和咯咯的叫声,可闻于很远的地方,据说这种声是由其长鼻增强的;雌象海豹的叫声则有所不同。莱生(Lesson)把这种鼻的竖立比做鹑鸡类在向雌者求偶时的垂肉膨胀。另一种亲缘相近的冠海豹(Cystophora cristata),其头上冠以巨大的兜帽、即囊状物。这是由鼻隔所支持的,鼻隔向后伸长甚远,且于鼻内隆起,高达7英寸。这种兜帽被有短毛,而且是肌肉质的,膨胀时可以超出整个的头部!雄冠海豹当发情时在冰上进行剧烈的争斗,它们的吼声“据说有时如此之高,以致可闻于四英里之外”。当受到攻击时,它们同样地吼叫;当激怒时其头部囊状物即行膨胀而颤动。有些博物学者相信它们的声音就是这样增强的,但也有人举出这种异常构造还有各种其他用途。布朗先生以为它有保护作用,以防止所有种类的意外事故;但这是不可能的,因为,曾经杀过600头这种海豹的拉蒙特先生向我确言,其雌者的兜帽是残迹的,而且雄者的兜帽在幼小时不发达。 (639)
气 味
有些动物,譬如说著名的美洲臭鼬(skunk),它们发出的那种压倒一切的气味好像是完全作为防御之用的。鼩鼱(Sorex)的雌雄二者均具腹部臭腺,毫无疑问,从鸷鸟和猛兽拒食它们的身体来看,这种气味是保护性的;尽管如此,其雄者的这种腺体在繁殖季节还是增大了。在许多其他动物中,雌雄二者的这种腺体是同等大小的, (640) 但它们的用途还不清楚。另外有些物种,这种腺体只限于雄者才有,或者雄者的腺体比雌者的更为发达;它们的作用几乎总是在发情季节变得更强。雄象面部两侧的腺体在这期间变大,并且分泌一种具有强烈麝香气的分泌物。许多种类的蝙蝠,其雄者在身体各个不同部位具有腺体和可以突出的囊袋;据信这等囊袋是有臭味的。
雄山羊放出的恶臭气是众所周知的,某种雄鹿的恶臭气也非常强烈而且持久。在普拉塔河岸边距离一群平原鹿(Cervus campestris)半英里下风处,我就觉察到那里的空气沾染了这种雄羊的气味,我曾用丝手帕包了一块这种羊皮回家,虽然常用常洗,在一年零七个月中,最初一把这块手帕打开,还可闻到它保持着的这种气味痕迹。这种动物在生育以后上才会散发它的强烈气味,如果在幼小时进行阉割,就永远不会散发这种气味。 (641) 除了某些反刍动物(如麝牛,Bos moschatus)的全身在繁殖季节弥漫着一般气味外,许多种类的鹿、羚羊、绵羊和山羊在身体各种不同部位、特别是在面部都具有臭腺。所谓泪囊或眶下窝(suborbital pits)也可以归入这一部分。这等腺体分泌一种半流体的恶臭物质,有时如此大量泌出,以致污及整个面部,我曾亲自看到一只羚羊就是如此。“雄者的这等腺体通常比雌者的为大,而且其发育受到去势的抑制。” (642) 按照德马雷(Desmarest)的材料,红斑羚羊(Antilope subgutturosa)的雌者完全缺少这种腺体。因此,这种腺体无疑同生殖机能有密切关系。在亲缘密切相近的诸类型中,这种腺体也是有时存在、有时不存在。成年的雄麝(Moschus moschiferus),其尾部周围的裸皮湿漉漉地沾满了芳香液体,而成年的雌麝以及未满两岁的雄鹿,其尾部周围具毛,而且不散发香气。这种鹿所特有的麝香囊从其部位来看,必然限于雄者所有,而且形成了一种附加的芳香器官。奇怪的是,这种腺体所分泌的物质,按照帕拉斯的说法,在发情季节,浓度不变,数量也不增加;尽管如此,这位博物学者还承认这种腺体的存在在某一方面是同生殖行为有关联的。但是,他对其用途只提供了一种推测的而且不能令人满意的解释。 (643)
在大多数场合中,如果只有雄者在繁殖季节散发强烈的气味,那么这大概是用以刺激或魅惑雌者。我们千万不要以我们的嗜好来判断这个问题,因为,众所熟知,鼠喜好某种香料油,猫喜好缬草,而我们却非常讨厌这些东西,狗虽然不吃死尸,却用鼻子嗅它们,并且在它们上面打滚。根据以上讨论公鹿鸣唤时所举出的理由,我们可以拒绝接受气味乃用以从远方招致雌者来就雄者的概念。积极的和长期连续的使用在这里不能发生作用,就像发音器官的情形那样。散发的气味对雄者来说一定是相当重要的,因为在某些场合中,大而复杂的腺体发达了,它们具有肌肉以便把囊袋翻开,并且启闭囊孔。如果气味最盛的雄者在赢得雌者方面是最成功的,而且所留下的后代遗传了它们逐渐完善化的腺体和气味,那么这等器官的发展就可以依据性选择得到解释了。
毛的发育
我们已经看到,雄四足兽在颈部和肩部的毛常常比雌者的发达得多;此外还可以举出许多有关事例。这种毛在雄者进行争斗时对它有保护作用,但在大多数场合中其发育是否特别为了这一目的,还很有疑问。我们差不多可以肯定的是,当沿着背部仅有一条稀而狭的脊毛时,情况并非如此;因为这种脊毛几乎不能提供任何保护,何况脊背也不是容易受到伤害的地方;尽管如此,这等脊毛有时也仅限于雄者所有,或者在雄者身上比在雌者身上更为发达得多。有两种羚羊,林羚(Tragelaphus scriptus参阅图70) (644) 和大羚羊(Portax picta),可作为例子。当马鹿以及雄野山羊被激怒和惊恐时,这等脊毛即行竖起 (645) ;但不能设想它们的发达仅是为了威吓其敌手。上述大羚羊(Portax picta)的喉部具有一大块界限分明的黑毛丛,雄者的这种毛丛比雌者的大得多。北非的鬣羊(Ammotragus tragelaphus)为绵羊科(sheep-family)的一个成员,悬挂在其颈部和前腿上半部特别长的毛差不多把它的前腿都遮盖住了;但巴特利特先生不相信这等毛盖对雄者有什么用处,而雄者的这种毛盖比雌者的要发达得多。
许多种类的雄四足兽同雌者的差别在于前者面部的某些部位具有较多的毛或不同特性的毛。例如,只是公牛在其前额具有卷毛。 (646) 在山羊科(goat family)中有三个亲缘密切近似的亚属,只是其雄者具有颔毛,有时且甚长;还有另外两个亚属,其雌雄二者均具颔毛,但普通山羊的某些家养品种则没有颔毛;塔尔羊(Hemitragus)的雌雄二者也都没有颔毛。北山羊(ibex)的颔毛在夏季不发达,在其他时期也非常之小,以致可以称为残迹。 (647) 某些猿猴,如猩猩,仅限于雄者才有颔毛;或者,雄者的颔毛比雌者的大得多,例如卡拉亚吼猴和僧面猴(Pithecia satanas,图68)就是如此。猕猴某些物种的颊毛是这样, (648) 狒狒某些物种的鬃毛也是这样。但大多数种类的猴,其雌雄二者面部和头部的各种毛丛都是一样的。

图 68 僧面猴(Pithecia satans),雄性
(引自布雷姆)
牛科(Bovidae)以及某些羚羊类各个成员的雄者均具颈部垂肉,即大型皮褶,而在雌者方面其发达程度就差得多。
关于这样的性差异,我们必须作出的结论是什么呢?大概谁也不敢说某些雄山羊的颔毛、公牛的颈部垂肉或某些雄羚羊沿着背部的脊毛在其普通习性方面对它们有任何用途。雄僧面猴(Pithecia)的巨大颔毛、雄猩猩的长颔毛在它们进行战斗时可能保护其喉部;因为伦敦动物园的管理员告诉我说,许多猴类彼此攻击对方的喉部;但是,要说颔毛的发达,其意义不同于颊毛、触须以及面部的其他毛丛,似乎是不可能的;而且谁也不会设想它们可用于保护。我们必须把毛和皮的所有这等附器仅仅归因于雄者无目的的变异性吗?不能否认这是可能的;因为,许多家养动物的某些性状,显然不是通过返祖从任何野生的祖先类型那里传下来的,这些性状仅限于雄者才有,或者在雄者比在雌者更为发达——例如,印度雄瘤牛(zebu-cattle)的隆肉,公肥尾羊的尾巴,几个绵羊品种的雄者前额的弓形轮廓,最后,雄伯布拉(Berbura)山羊 (649) 的鬃毛、后腿长毛以及颈部垂肉,都是如此。有一个非洲的绵羊品种,只是公羊有鬃毛,这是一种真正的次级性征,因为我听温伍德·里德(Winwood Read)说,如果对这种公羊施行去势,其鬃毛就不发育。正如在我的著作《动物和植物在家养下的变异》中所阐明的,要断言任何性状,甚至由半开化人所养的那些动物的性状,没有受过人类的选择并因而有所扩大,应该极其小心,但在刚才举出的那些例子中,却不可能如此,尤其是仅限于雄者所有的那些性状,或在雄者方面比在雌者方面更为强烈发达的那些性状,更不可能如此。如果确知上述非洲公羊同其他绵羊品种均属于同一个原始祖先的后裔,如果具有鬃毛、颈部垂肉的公伯布拉山羊同其他山羊均出自同一个祖先,而且假定选择未曾应用于这等性状,那么它们的发生必定是由于单纯的变异性以及限于性别的遗传性。
因此,把这个观点引申到生活在自然状况下的动物的所有相似事例看来是合理的。尽管如此,我还不敢相信这可适用于一切情况,如雄鬣羊(Ammotragus)喉部和前腿异常发达的毛以及雄狐尾猴的巨大颔毛。根据我对自然界所能做的研究,我相信高度发达的部分或器官是在某一时期为着一种特殊目的而获得的。有些羚羊,其成年雄者的色彩比雌者的色彩表现得更为强烈,有些猴类,其面部的毛排列优雅,颜色殊异,那些其毛冠和毛簇就可能是作为装饰物而被获得的;据我所知,这正是有些博物学者的见解。如果这是正确的,则它们是通过性选择而被获得的、至少是通过性选择而变异的,就很少疑问了;但这种观点对哺乳动物来说究竟能引申到何等地步却很难说。
毛和裸皮的颜色
首先我要大致谈谈我所知道的有关雄四足兽的色彩不同于雌者的所有事例。关于有袋类,正如古尔德先生告诉我的,其雌雄二者在这方面很少差异;但红色大袋鼠(kangaroo)提供了一个显著的例外,“雌者呈现优雅蓝色的那些部分,在雄者则为红色”。 (650) 卡宴(Cayenne) (651) 的负鼠(Didephis opossum)雌者的颜色据说比雄者的稍红。关于啮齿类,格雷博士说,“非洲松鼠,尤其是热带地方的松鼠,其毛皮在每年的某些季节比在其他季节更为鲜艳,而且其雄者的毛皮一般比雌者的更为鲜明”。 (652) 格雷博士告诉我说,他之所以举出非洲松鼠为例,是因为它们异常鲜明的颜色最好地表示了这种差异。俄国巢鼠(Mus minutus)雌者的色泽比雄者的较浅而且较暗。大多数雄蝙蝠的皮毛比雌蝙蝠的鲜明 (653) 。关于这种动物,多布森博士说道:“其差异部分地或者完全地决定于雄者皮毛色彩远为鲜艳者,或以不同斑纹或以某些皮毛部分较长为区别者,在任何可以觉察的范围内,仅见于视觉发达良好的、以果实为食的蝙蝠。”这一叙述值得注意,因为它同鲜明颜色是否由于是装饰性的而对雄性动物有所帮助这一问题有关。关于树懒(sloths)的一个属,格雷博士说,现已证实“其雄者在装饰上和雌者有所不同——这就是说,雄者在两肩之间生有一片柔软的短毛,一般或多或少地呈橘黄色,有一个物种则呈白色。相反,雌者却缺少这种标志。”
陆栖的食肉类和食虫类很少表现任何种类的性差异,包括体色在内。然而豹猫(Felis pardalis)是一个例外,其雌者的颜色如同雄者相比,“不及后者鲜明,而且雌者的灰褐色部分较暗,白色部分较不纯,斑纹较狭,斑点较小”。 (654) 同豹猫亲缘相近的线斑猫(Felis mitis),其雌雄二者也有差异,但程度较轻;雌者的一般颜色比雄者的颇淡,而且斑点的黑色也较差。另一方面,海栖食肉类或海豹类有时在颜色上的差异相当大,正如我们已经看到的,它们还有其他显著的性差异。例如南半球的褐海狗(()taria nigrescens),其雄者的体部上面呈浓艳的褐色;而雌者的体部上面则呈暗灰色,不过雌者获得其成年的色泽比雄者为早,雌雄二者的幼仔均呈深巧克力色。格陵兰海豹(Phoca groenlandica)的雄者呈茶灰色,在背部有一块奇特的马鞍形暗色斑纹;其雌者的身材要小得多,并且具有很不相同的外貌,“呈暗淡的白色或草黄色,背部则呈茶色”;“幼仔最初是纯白色的,同冰丘和雪几乎无法区别,这样它们的颜色便起着保护作用。” (655)
反刍类在颜色方面的性差异比在其他目中更常常发生。条纹羚羊(Strepsicerene antelopes)的这种差异是普遍的,例如雄大羚羊(Portax picta)呈青灰色,远比雌者的颜色为深,而且喉部有一个正方形白色斑块,蹄后上部毛丛有白色斑纹,双耳有黑色斑点,所有这些都比雌者的显著。我们已经看到,这个物种的毛冠和毛丛同样地在雄者方面比在无角的雌者方面更为发达。布莱思先生告诉我说,其雄者不换毛,而定期地在繁育期间其毛色变得较深。幼小的雄者在出生后12个月以前同幼小的雌者并无差别;如果在此时期以前对雄者进行去势,按照这位权威的材料,它的颜色就永远不会改变。这个事实是重要的,它可以证明大羚羊的颜色来源于性别,当我们听到 (656) 弗吉尼亚鹿(Virginian deer)的红色夏毛和青色冬毛完全不受去势影响时,这个事实的重要性就显而易见了。林羚属(Tragelaphus)的大多数或全部高度装饰的物种,其雄者的颜色都比无角雌者的为深,而且其毛冠也更加充分发达。华丽的德比大角斑羚(Derbyan eland),其雄者比雌者的体部较红,整个颈部较黑,间隔这两种颜色的白色带斑较宽。好望角大角斑羚(Cape eland),其雄者也比雌者的颜色稍深。 (657)
公黑印度羚(A.bezoartica)属于羚羊的另一个族(tribe),其雄者的颜色很暗,几乎是黑色的;而无角的雌者则呈浅黄褐色。布莱思先生告诉我说,关于这个物种,我们遇到了一系列同大羚羊(Portax picta)完全相似的事实,即:其雄者定期地在繁育季节改变颜色,去势对这种颜色改变的影响,雌雄二者的幼仔彼此无法区别。另一种黑印度羚(Antilope niger)的雄者是黑色的,而雌者以及雌雄二者的幼仔都是褐色的;水草地印度羚(A.sing-sing)的雄者比无角雌者的颜色要鲜明得多,而且前者的胸部和腹部的黑色也较深;卡玛印度羚(A.caama)的雄者,其身体各个部位的块斑和条纹都是黑色的,而在雌者方面这些则是褐色的;斑纹角马(A.gorgon)的雄者的颜色“几乎同雌者的颜色一样,只是较深而且较鲜明而已”。 (658) 还可举出一些相似的事例。
马来群岛的公爪哇牛(Bos sondaicus)几乎是黑色的,四腿和臀部则呈白色;母牛具有鲜明的暗褐色,牛犊在三岁之前也如是,此后便迅速改变颜色。去势的公牛则重视母牛的颜色。母克马斯山羊(Kemas goat)的颜色较浅,据说这些雌者们以及母喜马拉雅野山羊(Capra aegagrus)均较其雄者的颜色更为均匀。鹿的颜色很少呈现任何性差异。然而卡顿告诉我说,雄美洲赤鹿(Cervus canadensis)的颈部、腹部以及四腿的颜色均远比雌者的为深;但在冬季这种较深的颜色即逐渐褪去以至消失。我在这里可以提一下卡顿的园囿,那里有弗吉尼亚鹿的三个族,其体色彼此稍有差异,但这种差异几乎完全限于青色的冬季毛皮,即繁育时期的毛皮;所以这个例子可以同前章所说的鸟类的亲缘密切近似种、即代表种相比拟,它们的羽衣只在繁育时期才呈现差异 (659) 。南美沼地鹿(Cervus paludosus)的雌者以及雌雄幼鹿在鼻部不具黑色条纹,在胸部不具黑褐色线纹,而这些却都是成年雄者的特征 (660) 。最后,正如布莱思先生告诉我的,颜色美丽和具有斑点的南亚斑鹿(axis deer)的成熟雄者在体色方面比雌者深得多,但去势后的雄者永远不会获得这种颜色。
我们需要讨论的最后一个目为灵长目(Primates)。黑狐猴(Lemur macaco)的雄者一般呈煤黑色,而雌者呈褐色。 (661) 在新世界的四手目中,卡拉亚吼猴的雌者和幼者均呈灰黄色,而且彼此相似;当两岁时,雄性幼者变为红褐色;三岁时,除去腹部外均呈黑色,而到四岁或五岁时,腹部也变得十分黑了。赤吼猴(Mycetes seniculus)和白喉卷尾猴(Cebus capucinus)雌雄之间差异也非常显著;前一个物种,我相信还有后一个物种的幼者均同雌者相似。白头僧面猴(Pithecia leucocephala)的幼者也是同雌者相似的,其上部呈黑褐色,下部呈锈红色。蛛猴(Ateles marginatus)面部周围的毛丛在雄者呈黄色,在雌者则呈白色。再来看看旧大陆的情况:白眉长臂猿(Hylobates hoolock)的雄者除去眉的上方有一条白色带斑外,通身都是黑色的,而雌者则由白褐色到杂以黑色的暗色,但决没有完全黑色的。 (662) 关于美丽的白须长尾猴(Cercopithecus diana)其成年雄者的头部呈深黑色,而雌者的头部则呈暗灰色;前者大腿之间的皮毛为优雅的浅黄褐色,而后者的这一部分的颜色则较淡。关于美丽而稀有的髭长尾猴(Cercopithecus cephus),雌雄之间的唯一差异为雄者的尾巴呈栗色,而雌者的尾巴则呈灰色;但巴特利特先生告诉我说,当雄者到达成年时,其全身颜色都变得更为显著,而雌者则仍保持幼小时的颜色。按照所罗门·米勒(Solomon Müller)的彩色绘图,金黑瘦猴(Semnopithecus chrysomelas)差不多是黑色的,而雌者则是淡褐色的。关于狗尾猴(Cercopithecus cynosurus)和灰绿长尾猴(C.griseoviridis),其雄者身体的一部分呈最鲜艳的青色或绿色,同其臀部的鲜红色裸皮形成了显著的对照。
最后,在狒狒科(baboon family)中,埃塞俄比亚鼯猴(Cynocephalus hamadryas)的成年雄者不仅在其巨大鬃毛方面同雌者有所差别,而且在毛和胼胝的颜色方面也和雌者稍有不同。山魈(C.leucophaeus)的雌者和幼者均比成年雄者的颜色较淡,而且比后者的绿色较浅。在整个哺乳动物纲中,没有一个成员像西非山魈(C.mormon)的成年雄者那样颜色特殊的。其面部到成年时即变成优雅的青色,鼻梁和鼻尖则呈最鲜艳的红色。按照有些作者的材料,其面部还有苍白的条纹,而且部分地略现黑色,不过这等颜色似乎是变异的。其前额有一毛冠,而且在下巴上有黄须。“其股之上方及臀部大片裸皮呈极其强烈的红色,并杂以优雅的青色,这就使其颜色越发鲜艳活泼。” (663) 当西非山魈激动起来时,所有无毛部分的颜色都变得更为鲜艳得多。有几位作者以最热烈的词句来描写这等灿烂的颜色,他们把这等颜色同最美丽的鸟类的颜色相比拟。另一个值得注意的特性是,当大犬齿充分发育时,在其双颊便形成了巨大的骨质突起,这骨质突起有纵向的深沟,在它上面的裸皮呈鲜艳的颜色,就像以上所叙述的那样(图69)。在其雌者和雌雄幼者方面几乎看不见有这种骨质突起,无毛部分的颜色也远不及雄者那样鲜明,而且它们的面部差不多是黑色的,带有青的色调。然而成年雌者的鼻子到一定的时期却变成红色。
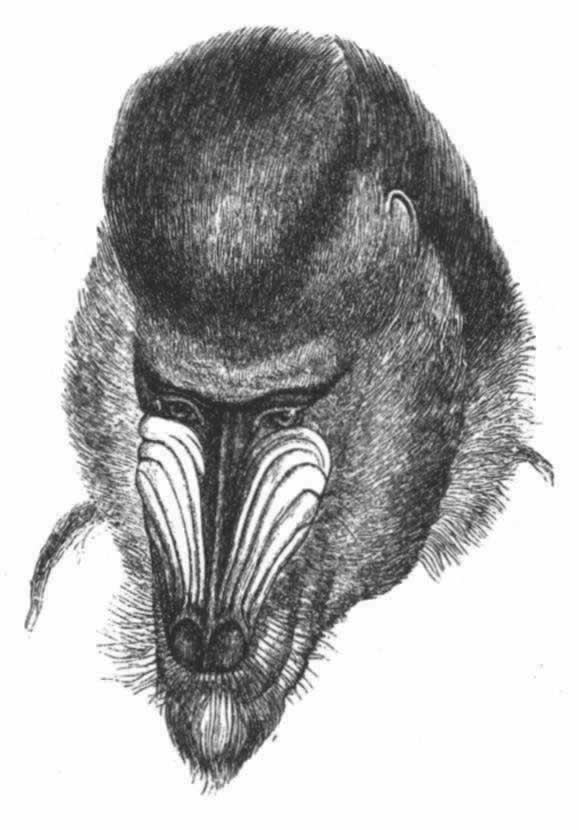
图 69 雄西非山魈(mandrill)的面部
(引自热尔韦的《哺乳动物志》)
迄今所举的一切事例都表明雄者比雌者的颜色更为强烈或更为鲜明,而且雄者的颜色同雌雄幼者的都不相同。但是,就像某些少数鸟类那样,雌者的颜色比雄者的更为鲜明,恒河猴(Macacus rhesus)也是如此,其雌者尾部周围的裸皮面积甚大,呈一种鲜艳的胭脂红色,伦敦动物园的管理员向我确言,这种颜色定期地表现得更为鲜艳活泼,它的面部也是浅红的。另一方面,恒河猴的成年雄者以及雌雄幼者(我曾在伦敦动物园中见到)不论其臀部裸皮或面部一点也没有红色的痕迹。但是,根据有些发表的记载看来,其雄者的确偶尔地或在某些季节表现有一些红色的痕迹。虽然在装饰方面不及雌者,但是雄者的手较大,犬齿较长,颊须较发达眉脊较突出,在这些方面还是遵循雄者胜过雌者的普遍规律。
有关哺乳动物雌雄二者之间的颜色差异,现在我已经举出我所知道的一切事例。有些这等差异可能是变异的结果,而这种变异只限于同一性别并向同一性别传递,并不因此获得任何利益,所以不借助于选择。关于我们的家养动物就有这样的事例,例如某些猫类的雄者是锈红色的,而其雌者却呈龟甲色。在自然界中也有近似的例子:巴特利特先生曾见到美洲豹(jaguar)、豹、袋貂(Vulpine phalanger) (664) 和毛鼻袋熊(wombat) (665) 的许多变种是黑色的,他肯定所有或者几乎所有这等动物都是雄性的。另一方面,狼、狐而且显然还有美洲松鼠,其雌雄二者偶尔生下来就是黑色的。因此,十分可能的是,某些哺乳动物的雌雄二者在颜色上的差异,并不借助于选择,而单纯地为一种或一种以上变异的结果,且这等变异的传递从一开始就专限于某一性别。尽管如此,某些四足兽、例如上述猿猴类和羚羊类的各种各样的、鲜艳的和对照鲜明的色彩还是不可能因此得到解释。我们应该记住,这等颜色并不是在雄者降生时出现的,而仅仅是在成熟期或接近成熟期才出现的;而且这和普通变异有所不同,如果对雄者施行去势,这等颜色就消失了。总之,雄性四足兽的强烈显著的颜色以及其他装饰性状,在它们同其他雄者竞争时大概是有利的,因而是通过性选择而获得的。根据上述各点可以推断,这一观点由于以下的情况而被加强,即:雌雄二者在颜色上的差异几乎完全是发生于表现有其他强烈显著第二性征的那些哺乳动物类群或亚类群;这等第二性征也是性选择的结果。
四足兽对颜色显然是注意的。贝克爵士屡屡见到非洲的象和犀特别愤怒地对白色或灰色的马进行攻击。我在他处曾阐明, (666) 半野生的马显然喜爱那些颜色和自己相同的马,颜色不同的黇鹿群,虽在一起生活,却长期保持界限分明。还有一项更有意义的事实:一匹母斑马不接受一头公驴的追求,可是当把这头驴涂饰成斑马的模样时,正如约翰·亨特所说的,“母斑马就欣然同意那头公驴了”。“从这一奇妙的事实,我们看到了仅仅由颜色所激发起来的本能,这一本能的作用如此之强,以致胜过了任何其他本能。但雄者并不需要这种本能,只要雌者同他自己的颜色稍为相似,就足以使他激动起来。” (667)
在以前的一章我们曾看到,高等动物的心理能力同人类的、尤其是同低等野蛮种族的相应能力,虽在程度上有重大差别,但在性质上并无不同;看来甚至人类的审美感同四手目的审美感也没有广泛的差异。非洲黑人把面部肌肉弄成平行的隆起条纹,“即疤痕,高出颜面的本来表面,这种丑陋的毁形却被视为个人容貌的巨大魅力”; (668) 世界许多地方的黑人和未开化人在他们的面部画上红的、青的、白的和黑的带斑;与此相似,西非狒狒也获得了它们的具有深刻凹痕和色彩绚丽的面部,以吸引雌者。毫无疑问,臀部的颜色为了装饰之故甚至比面部更为鲜艳,这在我们看来,是极其滑稽可笑的;但这比许多鸟类尾羽具有特别装饰,并不会使人感到更为奇怪。
关于哺乳动物,目前我们还没有掌握任何证据可以证明雄者尽力在雌者面前夸示其魅力;而雄鸟和其他雄性动物以其精心设计的方式来进行这样的表演,乃是最强有力的论点来支持如下的信念:雌者赞赏在其面前展示的装饰物和颜色,或受到这等装饰物和颜色的刺激而兴奋起来。可是哺乳类和鸟类在它们的一切第二性征方面还是有显著的平行现象,即,在它们所具有的同其雄性对手进行争斗的武器方面,在它们所具有的装饰性附器方面,在它们的颜色方面,均有平行现象。在这两个纲的动物中,如果雄者和雌者有所不同,雌雄二者的幼仔却几乎总是彼此相似,而且在大多数场合中,它们的幼仔也同成年的雌者相似。在这两个纲的动物中,雄者在繁殖龄期不久之前会表现出这一性别所特有的性状;如果在早期施行去势,这等性状就要消失。在这两个纲的动物中,颜色的改变时常是季节性的,而且无毛部分的色泽时常在求偶的行为中变得更为鲜艳活泼。在这两个纲的动物中,雄者几乎总是比雌者的颜色更为鲜艳活泼,或者更为强烈,而且雄者装饰有较大的冠毛或羽冠以及其他这类附器。在少数例外的场合中,这两个纲的雌者比雄者的装饰更为高级。有许多种哺乳动物,至少有一种鸟,其雄者比雌者所散发的香气为甚。在这两个纲的动物中,雄者比雌者的声音更加强有力。鉴于这种平行现象,毫无疑问,有一个同样的原因,不管它是什么,曾对哺乳类和鸟类发生作用;仅就装饰的性状来说,在我看来,其结果可以归因于某一性别的个体对异性某些个体长期连续的喜爱,而且结合着它们在遗留大量后代以承继其优越的魅力方面获得成功。
装饰性状对雌雄二者的同等传递
由类似之理推之,可信许多鸟类的装饰物最初是由雄者获得的,然后同等地或几乎同等地传递给雌雄二者;那么我们可以问,这一观点对哺乳动物究竟能应用到怎样程度。相当多的物种,尤其是较小的种类,其雌雄二者的颜色是为了保护自己而获得的,同性选择并无关系;但就我所能判断的来说,这样的事例不及在大多数较低等诸纲中那样多,而且其表现方式也不那样显著。奥杜邦说道,当麝鼠(musk-rat) (669) 蹲在浑浊河流的岸边时,他常常误认它为一块泥土,其形酷似。山兔(hare)当跑向兔穴时凭借颜色而隐蔽起来的事例是众所熟知的;但这一原理对一个亲缘密切近似的物种——家兔(rabbit)就部分地不适用了,因为当它跑向兔穴时,它那向上翻卷的白尾就会引起猎人、无疑也会引起一切猛兽的注意。谁都不会怀疑栖息在白雪覆盖地方的四足兽变为白色,乃是为了保护自己免受敌方的危害,或者有利于它们潜近所要捕食的动物。在容易化雪的地方,白色的毛皮将会有害;因而在世界上较热的地区,白色物种是极其罕见的。值得注意的是,栖息在不甚寒冷地方的许多四足兽虽然没有白色的冬季毛皮,但在这一季节其毛皮颜色却变得较淡;显然这是它们长期暴露于其中的外界条件所造成的直接结果。帕拉斯述说 (670) ,在西伯利亚,狼、鼬属(Mustela)的两个物种,家马、野驴(Equus hemionus) (671) 、家牛,印度羚的两个物种、麝、狍(roe)、驼鹿、驯鹿都会发生这种性质的颜色变化。例如狍其夏季皮毛是红色的,而冬季皮毛则是灰白色的,当这种动物漫游于那些点缀着白雪和严霜的无叶灌木丛中时,那种灰白色对它们也许可以起一种保护作用。如果上述动物扩展其栖息范围而达到永久覆盖冰雪的地方,那么通过自然选择它们的淡色冬季皮毛大概会变得愈来愈白,直到白得似雪为止。
瑞克斯(Reeks)先生给我的一个奇妙事例表明,一种动物由于具有独特颜色而得到了利益。他在一个有围墙大园内养了五六十只褐白杂色的家兔;同时在他的家里还养着一些同样颜色的猫。正如我常常注意到的那样,这种猫在日间是很显眼的,但它们惯于在黄昏时刻卧守于兔穴之口,而那些家兔显然不能把它们同其杂色弟兄分别开来。其结果便是,每一只这样杂色的家兔在18个月内全被灭绝;而且有证据表明这都是那些猫干的。颜色对另一种动物——臭鼬似乎也是有助益的,其方式正如其他动物纲中许多事例所表现的那样。当这种动物激怒时,便会散发出可怕的气味,所以没有一种动物会自愿地攻击它;但在黄昏时就不容易把它辨识出来,因而可能受到猛兽的攻击。这样,正如贝尔特(Belt)先生 (672) 所相信的那样,臭鼬便有一条白色的蓬松大尾巴,作为一种容易引起注意的警告。

图 70 林羚(Tragelaphus scriptus)的雄者
(引自诺斯雷动物园,Knowsley Menagerie)
虽然我们必须承认许多四足兽获得它们现在那样的颜色,或是为了保护自己,或是为了有助于捕食其他动物,但是,还有大量的物种,其色彩太显眼了,而且颜色的排列也太奇特了,以致不能允许我们去设想它们可用于这等目的。我们可以用某些羚羊的情况来做例证;当我们看到喉部的正方形白色块斑、蹄部后上方丛毛的白色标志以及双耳的黑色圆形点斑,在大羚羊(Portax picta)的雄者方面均比在其雌者方面更为明显——当我们看到德比大角斑羚(Oreas derbyanus)的雄者比雌者的颜色更为鲜艳活泼,而且肋部的狭白线和肩部的宽白斑更为明显——当我们看到具有奇妙装饰的一种林羚(图70)雌雄二者之间的相似差异,——我们无法相信这种种差异对雌雄任何一方在其日常生活习性方面有什么用处。更可能的结论似乎是:这各种不同的斑纹最初是由雄者获得的,其颜色通过性选择而被加强,然后部分地传递给雌者。如果承认这一观点,毫无疑问的是,许多其他羚羊的同等奇特的颜色和斑纹,虽为雌雄双方所共有,却是按照同样的方式而被获得和传递的。例如,南非捻角羚羊(Strepsiceros kudu,图64)雌雄二者在其后肋均有狭窄的垂直白线,而且在其前额均有优雅的角形白色斑纹。南非达玛利斯羚属(Damalis)雌雄二者的颜色都很奇特;白臀达玛利斯羚(D.pygarga)的背部和颈部呈紫白色,到两肋逐渐变为黑色;这等颜色同其白腹和臀部的一大块白斑截然分明;其头部的颜色还更奇特,有一块镶着黑边的椭圆形白斑遮盖面部直达双眼(图71),在前额还有三条白纹,双耳也有白色的标志。这个物种的幼羚通身均呈淡黄褐色。白耳达玛利斯羚(Damalis albifrons)的头部同前一个物的头部有所不同,前者头部只有一条白纹,而不是三条白纹,并且它的双耳几乎是完全白色的。 (673) 在我尽力研究了所有各纲动物的性差异之后,我不能不作出如下结论:许多羚羊的排列奇妙的颜色,虽为雌雄双方所共有,却是最初应用于雄者的性选择之结果。
同一结论也许可以引申到虎,这是世界上最美丽的动物,但其雌雄二者无法从颜色方面加以区别,即使野兽商人也不能做到这一点。华莱士先生相信, (674) 虎的条纹皮毛“同竹子的挺直茎干如此谐调一致,以致可以帮助它们隐蔽起来去接近所要捕食的动物”。但是,在我看来,这一观点恐难令人满意。我们有某种微小的证据可以证明虎的颜色可能是由性选择所致,因为猫属(Felis)有两个物种,其彼此相似的斑纹和颜色在雄者方面均比在雌者方面更为鲜明。斑马的条纹是容易引起注目的,而且在开阔的南非平原上这等条纹不能提供任何保护。伯切尔(Burchell) (675) 在描述斑马群时说道:“它们柔滑发亮的肋部在日光中闪耀,它们鲜明的、整齐的条纹皮毛呈现了一幅非常美丽的图画,大概没有任何四足兽可以胜过它们。”但在整个马科(Equidae)中,雌雄二者的颜色是完全一致的,所以在这里我们还没有掌握性选择的证据。尽管如此,这位曾把各种羚羊肋部的白色和暗色垂直条纹归因于性选择作用的人,大概还会把同样的观点引申到兽中之王——虎以及美丽斑马的。
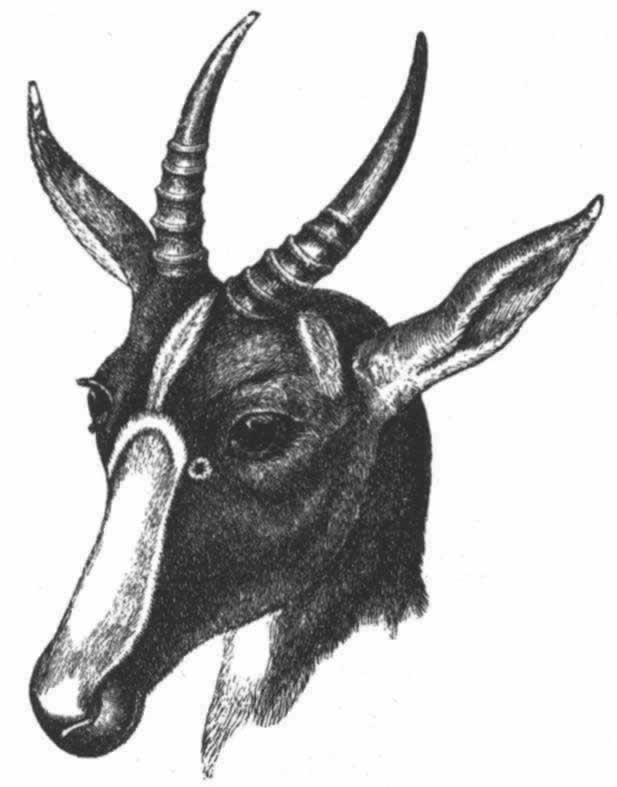
图 71 白臀南非达玛利斯羚(Damalis pygarga),雄者
(采自诺斯雷动物园)
我们在前一章已经看到,属于任何纲的幼小动物所遵循的生活习性如果同其双亲的生活习性差不多相同,但其颜色却有差异,那么可以推论这等幼小动物保持了某一绝灭了的古老祖先的颜色。在猪族以及貘族中,幼仔具有纵条纹,这同这两个类群的一切现存的成年物种均有差异。许多种类的幼鹿具有优雅的白色斑点,而其双亲却一点没有表现这种痕迹。从斑鹿——其雌雄二者在一切年龄和一切季节都具有美丽斑点(雄者比雌者的颜色更强烈),到无论老鹿和幼鹿均不具有斑点的物种可以找出一条逐渐的系列。我将举出这个系列中的一些等级。满洲鹿(Cervus mantchuricus)全年均具白色斑点,但我在伦敦动物园里看到,在夏季其斑点就比在冬季淡得多,其一般的夏季皮毛颜色也较淡,而一般的冬季皮毛颜色就较深,而且双角也充分发达。豚鹿(Hyelaphus porcinus)的斑点在夏季极其显著,那时它的皮毛呈赤褐色,但在冬季它的斑点就完全消失,那时它的皮毛则呈褐色。 (676) 这两个物种的幼鹿均具斑点。幼弗吉尼亚鹿同样也有斑点,卡顿告诉我说,在他的园囿中约有百分之五的这种成年鹿当红色夏季皮毛由带蓝色的冬季皮毛所代替时,在其双肋便暂时地各现一行斑点,这两行斑点的清晰度虽有变异,但其数目却永远是一样的。从这种状态到成年鹿在一切季节中均不具斑点者,相距不过很小的一步;最后则到达在一切年龄和一切季节中均不具有斑点的状态,有如某些物种所发生的那样。根据这一完整系列的存在,尤其是根据如此众多物种的幼鹿均具斑点,我们便可断言鹿科现存的成员乃是某一古代物种的后裔,这个物种就像轴鹿那样地在一切年龄和一切季节中均具斑点。它们更早的一个古代祖先大概同西非鹿(Hyomoschus aquaticus)多少相似,因为这种动物具有斑点,而且无角雄者具有可以发挥作用的大型犬齿,现在还有少数真正的鹿保持着犬齿残迹。西非鹿也是一个类型把两个类群联结在一起的有趣事例之一,因为它在某些骨骼性状上介乎厚皮类(pachyderms)和反刍类之间,而以前却认为这两类动物是截然相异的。 (677)
于是在这里产生了一个奇难的问题。如果我们承认有色的斑点最初是作为装饰物而被获得的,那么如此众多的现存鹿——本来具有斑点的动物之后裔以及猪和貘的所有物种——本来具有条纹的动物之后裔,怎么会在成熟状况下失去了其以往的装饰物?我还不能令人满意地回答这个问题。我们几乎可以肯定的是,现存物种的祖先在成熟期或接近成熟期失去了斑点和条纹,所以幼仔依然保持着它们;而且,由于在相应年龄遗传的法则,这等斑点和条纹传递给此后各代的幼仔。由于狮和美洲狮的生息地是开阔的,所以条纹的消失可能对它们有重大利益,这样便可不易为猎物所见;如果达到了这一目的的连续变异发生于生命的较晚时期,那么幼仔大概还会保持其条纹,现在的情况正是如此。弗里茨·米勒(Fritz Müller)向我提出,关于鹿、猪和貘,通过自然选择而去掉其斑点或条纹,它们大概就不易被敌者所见;并且食肉动物在第三纪体格增大,数量增多的时候,它们大概特需要这种保护。这也许是正确的解释,但幼仔没有受到这样的保护就颇为奇怪了,而且更加奇怪的是,有些物种的成年动物却在每年的一部分时期局部地或者完全地保持了它们的斑点。我们知道,当家驴发生变异并且变为赤褐色、灰色或黑色时,其肩部、甚至脊部的条纹常常消失,虽然我们还不能说明其原因。在身体任何部分均具条纹的马,除暗褐色者以外,为数很少,然而我们有良好的理由可以相信原始马在其腿部、脊部、大概也在肩部均具条纹。 (678) 因此,在现存的成年的鹿、猪和貘中,其斑点和条纹的消失可能是由于一般的皮毛颜色发生了变化;但这种变化究竟是由于性选择或自然选择的作用,还是由于生活条件的直接作用,抑或由于某种其他未知的原因,还不可能决定。斯克莱特先生所做的一个观察很好地示明了我们对于那些支配条纹的出现和消失的法则乃一无所知;栖息在亚洲大陆的驴属(Asinus)一些物种均不具条纹,甚至连肩部的横条纹也没有,而栖息在非洲的那些物种则具有显著的条纹,但纹驴(A.taeniopus)这个物种是一个局部的例外,它只有肩部横条纹,一般在腿部还有一些模糊不清的带斑;这个物种所栖息的地带几乎介于上埃及(Upper Egypt)和埃塞俄比亚之间。 (679)

图 72 赤瘦猴(Semnopithecus rubicundus)
[本图及以下各图(引自热尔韦)示明头毛奇特的排列和发育。]
四手类
当作出结论之前,先稍微谈一谈猿猴类的装饰物将是有益的。在大多数物种中,其雌雄二者在颜色方面彼此相似,但在某些物种中,就像我们已经看到的那样,雄者同雌者有所差异,尤其是在皮肤无毛部分的颜色方面、在颔毛、颊须和鬃毛方面更加如此。许多物种的颜色如此特殊或者如此美丽,而且具有如此奇妙而漂亮的冠毛,以致我们不能不把这等性状的获得视为用做装饰。附图(图72至图76)示明几个物种的面毛和头毛的排列情形。要说这等冠毛以及毛和皮的对照强烈的颜色仅仅是变异的结果,而没有选择作用的帮助,几乎是不可想象的;而且,要说这等性状在任何日常生活方面对这等动物有什么用处也是不可想象的。如果这样说法不错,那么这等性状大概是通过性选择而获得的,虽然它们同等地或者几乎同等地传递给了雌雄双方。关于许多四手类动物,我们还有另外的证据可以证明性选择在以下各方面的作用:如雄者比雌者的体格较大、体力较强而且犬齿较发达。
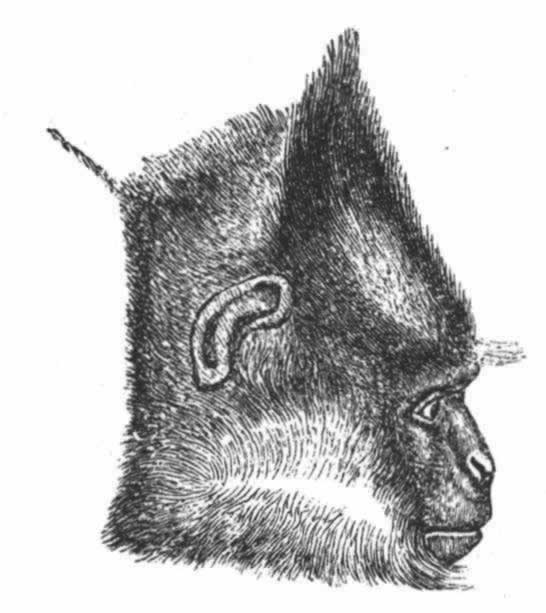
图 73 怒发瘦猴(Semnopithecus comatus)的头部

图 74 白喉卷尾猴(Cebus capucinus)的头部

图 75 蛛猴(Ateles marginatus)的头部

图 76 长毛卷尾猴(Cebus vellerosus)的头部
举出少数事例就足以说明有些物种雌雄二者的奇异颜色以及其他物种的美丽。长尾猴(Cercopithecus petaurista,图77)的面部是黑色的,颊须和颔毛是白色的,它的鼻部具有一个界限分明的圆形白色斑点,其上蔽以白色短毛,使得这种动物的外貌差不多是滑稽可笑的。额斑猴(Semnopithecus frontatus)的面部也是稍带黑色的,而且还有黑色的长颔毛,在其青白色前额上有一个无毛的大斑点。多毛猕猴(Macacus lasiotus)的面部呈不鲜明的肉色,而且在其双颊各有一个界限分明的红色斑点。埃及南部白眉猿(Cercocebus aethiops)的外貌是滑稽可笑的,它具有黑色的面部、白色的颊须和颈毛以及栗色的头部,在两个眼睑上各有一个无毛的大型白色斑点。很多物种的颔毛、颊须以及面部周围的冠毛同其余头部的颜色是不同的,如果不同,它们的色泽总较淡, (680) 常常是白色的,有时是亮黄的或者是微红的。南美短尾猴(Brachyurus calvus)的整个面部均呈灿烂的猩红色,但不到这种动物接近成熟时,不会出现这种颜色。 (681) 各个不同物种的面部裸皮在颜色上差异非常之大。它常常呈褐色或肉色,局部呈白色,而且也常常呈黑色,有如最黑的黑人面色一般。秃顶猴的猩红色面部比含羞的高加索少女的面部更为鲜艳。有的面部呈橘黄色,比任何蒙古人的面部黄色更加明显,有几个物种的面部呈青色,亦有呈紫罗兰色或灰色者。在巴特利特先生所知道的一切物种中;凡是成年雌雄二者的面部呈浓色时,在其早年的幼小时期,这等颜色则是暗淡的,或竟不具颜色。西非狒狒和恒河猴的情况也是如此,其面部和臀部具有鲜艳颜色者只是雌雄中的一方。在后述这等场合中,我们有理由相信这等颜色是通过性选择而被获得的;我们自然地便被引导把同样的观点引申到上述物种,虽然其雌雄二者在成年时的面部具有同样的颜色。

图 77 长尾猴(Cercopithecus petaurista)
(采自布雷姆)

图 78 白须长尾猴(Gereopithecus diana)
(引自布雷姆)
虽然许多种类的猴按照我们的趣味来看,远远不是美丽的,但另外有些物种由于它们的漂亮外貌和鲜艳颜色普遍地受到了称赞。眉线瘦猴(Semnopithecus nemaeus)的颜色虽然奇特,却被描写得非常之美;其橘黄色面部绕以具有光泽的白色长颊须,在双眉之上各有一条栗红色的线;背部皮毛呈雅致的灰色,腰部各有一块方斑,尾部和前臂是纯白的;胸部覆盖着栗色的皮毛;大腿为黑色,小腿为栗红色。我只再谈一谈另外两种猴的美;我之所以选用它们是因为它们在颜色方面表现了轻微的性差异,这就在某种程度上可能说明其雌雄二者的漂亮外貌是由于性选择所致。髭猴皮毛的一般颜色为带有斑驳的微绿色,喉部为白色;其雄者的尾端为栗色,但其面部则系最富装饰的部分,面皮主要呈微带青色的灰色,至双眼下方逐渐变为微黑色,上唇呈优雅的青色,下唇边有一条稀疏的黑髭,颊须为橘黄色,其上部则呈黑色,向后延伸到双耳,形成一条带形物,双耳则覆被着微白色的毛。在动物学会的动物园里,我常常无意中听到游客们赞美另一种猴,它可以名副其实地称为白须长尾猴(图78);其皮毛的一般颜色为灰色;胸部和前腿内面呈白色;背的后部有一个界限分明的大三角形块斑,呈鲜艳的栗色;雄者大腿的内面及其腹部为优雅的浅黄褐色,而且头顶呈黑色;面部和双耳为浓黑色;同其双眉上方的横向丛毛和白色长颔毛形成了优美的对照,颔毛的基部则呈黑色。 (682)
在这等猴以及许多其他猴中,其颜色之美及其奇特的排列,尤其是头部冠毛以及簇毛的各式各样的优雅排列,迫使我不得不相信这等性状完全是作为装饰物,通过性选择而获得的。
提 要
为了占有雌者而进行战斗的这一法则,看来是通行于整个的巨大哺乳纲的。大多数博物学者们都承认雄性动物的较大的体格、体力、勇气以及好斗性,它特有的进攻武器以及特有的防御手段,都是通过我称为性选择的那种方式而获得或变异的。这并不决定于一般生存竞争中的任何优越性,而是决定于某一性别的某些个体、一般是雄者的某些个体成功地战胜其他雄者,并比成功较小的雄者留下较大数量的、遗传其优越性的后代。
还有另一种比较和平的竞争,在这种竞争中雄者尽力以各种不同的魅力去刺激或引诱雌者。在某些场合中这种竞争大概是由雄者在繁殖季节散发出强烈的气味来进行的;散发气味的腺体是通过性选择而获得的。与此相同的观点是否可以引申到声音,尚有疑问,因为雄者发音器官的加强,一定是由于雄者在成熟时期受到了爱情、嫉妒或愤怒的强烈刺激而使用这等器官的缘故,因而这种特性只向雄者传递。各种不同的冠毛、簇毛和鬃毛或者仅限于雄者所有,或者在雄者方面比在雌者方面更为发达,它们在大多数场合中似乎仅仅是为了装饰,虽然有时也用做防御雄性竞争对手的手段。甚至有理由来设想公鹿的枝角以及某些羚羊的漂亮双角,虽然用做进攻的或防御的武器,大概也是部分地为了装饰而发生变异的。
当雄者在颜色上不同于雌者时,雄者的色泽一般表现得较深而且对照较强烈。在哺乳纲中我们所看到的华丽的红色、青色、黄色和绿色并不像在雄性鸟类和许多其他动物中那样普遍。无论如何,某些四手类动物的无毛部分必须除外,因为这等部分的位置往往是奇特的,而且在某些物种中,其颜色是鲜艳的。在其他场合中,雄者的颜色可能是单纯地由于变异,而不借选择之助。但是,如果其颜色是丰富多彩而且强烈显著的,如果它们不接近成熟时不会发展,而且,如果它们在去势以后就会消失,那么我们简直不得不作出如下结论:它们是为了装饰通过性选择而获得的,而且完全地或者几乎完全地只向同一性别传递。如果雌雄二者具有同样的颜色,而且这等颜色是显著的或者排列奇妙的,却一点没有作为保护的明显用途,尤其是如果它们和各种不同的其他装饰性附器结合在一起,那么我们便可用类推方法作出同样的结论,即:它们是通过性选择而获得的,虽然它们是向雌雄双方传递的。显著而丰富多彩的颜色,不论是仅限于雄者所有或为雌雄二者所共有,按照一般规律,在同一类群或同一亚类群中都是和用于战争或装饰的其他次级性征结合在一起的,如果我们回顾一下本章和前一章中所举的各种不同事例,便可知道情况确系如此。
向雌雄双方同等传递性状的法则,仅就体色和其他装饰物而言,通行于哺乳类远比通行于鸟类更加广泛;但是,诸如角、獠牙那样的武器往往专向雄者传递,或者传递给雄者远比传递给雌者更加完全。这是使人惊奇的,因为雄者使用其武器来防御一切种类的敌对者,所以这等武器对于雌者大概也是有用的。仅就我们所能知道的来看,雌者缺少这等武器只能根据通行的那种遗传形式才能得到解释。最后,在四足兽中同一性别的诸个体之间所进行的争斗,无论是和平的还是流血的,除了极罕见的例外,仅限于雄者才进行之;所以雄者通过性选择发生变异者远比雌者更加普遍,无论是在彼此之间的战争方面,还是在向异性进行引诱方面,均系如此。

雄蜂鸟之美几乎可与极乐鸟相匹敌,凡是见过古尔德先生的佳作或其丰富采集品的人都会承认这一点。

▲这幅漫画想说明:根据达尔文的进化理论,艺术家是由刷子和颜料罐进化而来的(讽刺画,后期着色木版画,1879年)。

▲维多利亚女王时代的中层社会都认为,女王是一位完美的女子,她是王朝的天使,伦理的捍卫者和王朝一切美好生活的源泉。虽然猿猴在很多方面与人类有惊人的相似,但是这些人不敢想象,美丽的女子与猴的后代有何关联?正如图中所示,把她与多毛的猿猴联想起来是多么不可能!

▲这是英国收藏家霍金斯所画,表现了达尔文《物种起源》中的论述“人类最早的祖先是外表丑陋、毫无魅力的哺乳动物,并且身体矮小,由于经常被风吹日晒,皮肤变成为难看的暗棕色。全身大部分皮肤都覆着长而粗糙的毛发。”
达尔文在《人类的由来及性选择》一书中阐明了他在《物种起源》中已经形成的观念,即物种起源的一般理论也完全适用于人这样一个自然的物种。他进一步认为,人类的智力、人类社会道德和感情的心理基础等特性也像人体结构的起源那样,可以追溯到较低等的动物。

▲《人类的由来及性选择》英文版。该书在1871年问世之后的3年里,就连续重印数次。1874年第二版时,达尔文又进行了一些改正。本中文版即是根据其英文第二版译得。

▲《人类的由来及性选择》现代法文版的封面。书中,达尔文通过相当多的研究证据,来支持人类源于动物的思想。他指出,人耳上残存的耳郭,其中有些肌肉显然是用于移动耳朵的。他还提到,脊柱底部的尾骨,是痕迹器官的又一例证,显然它是人种演变过程中的早期遗留产物。
▲当年的一份拉丁文报纸详细地阐述了达尔文关于人类进化的理论。
▲根据达尔文的理论而绘制的人类进化图。达尔文在本书结论里谈到:人类起源于某种低等的生物类型,将会使许多人感到非常厌恶。但几乎无可怀疑的是,我们乃是未开化人的后裔。
本书的第一部分,主要是论述人类起源于低等生物类型,为此,达尔文引用大量的事实证据来证明自己的观点。他认为:“人类是按照其他哺乳动物同样的一般形式或模型构成的。人类骨骼中的一切骨可以同猴的、蝙蝠的或海豹的对应骨相比拟。人类的肌肉、神经、血管以及内脏也如此。”
▲赫胥黎通过解剖手段证明,人脑也遵循达尔文提出的这一法则。图为赫胥黎《人类在自然界的位置》一书的插图,比较了从猿到人的骨骼结构。

▲脑部结构的比较图。19世纪末以后,陆续发现了很多古猿和古人类化石,为研究人类起源提供了直接证据。

▲达尔文在本书中提到:“美洲蛛猴,当喝醉酒之后的第二天早上,非常易怒而忧郁,用双手抱住疼痛的脑袋,作最可怜的表情,当再给它们啤酒或果子酒时,它们就厌恶地躲开,但对柠檬汁却喝得津津有味……这些证明了人类和猴类的味觉神经是多么相似,而且他们的全部神经系统所受到的影响又是多么相似。”图为一只美洲蛛猴。
达尔文在本书第二部分,用性别选择理论对某些物种作了说明:物种个体(往往是雄性个体)之间的争斗往往是为了有限的性伙伴,而不是为了有限的环境资源。

▲雄性鸥鸽。达尔文说:“我养的一种鸟,当它膨胀起颈项,喙就埋进了脖子。雄性鸥鸽尤为突出,一受刺激,喉咙就鼓得比雌性更厉害,而且,还因自己有能力展现这个本事而洋洋得意。”

▲求偶过程中的雄性孔雀为了夸示自己,吸引雌孔雀的注意,便展开并树立其尾羽,尾羽上有美丽的眼斑。

▲雄性麝香鸭,在夏季月份会散发出一种麝香气味,用于魅惑或刺激雌性,有些个体可以把这个气味保持全年。野生的雄性麝香鸭,在繁殖季节常会发生血腥战斗。

▲达尔文认为,在庞大的昆虫纲中,雌雄的差异有时表现在运动器官上,但往往是表现在感觉器官上,如许多物种的雄虫所具有的节状触角和美丽的羽状触角即是。但他关心的主要是使某只雄者在战斗中或求偶中凭其体力、好斗性、装饰,或音乐去战胜其他雄者的那些构造。比如,达尔文观察到:有一种蜻蜓(Anax junius),其雄者的腹部呈鲜艳的青蓝色,而雌者的则呈草绿色。
达尔文在本书的第三部分,详细讨论了人类的第二性征,并明确地得出本书的结论:人类起源于某种低等的生物类型。

▲达尔文认为:不同种族的、甚至同一种族不同部落或家族的男人,在胡须以及体毛的发育方面均有显著差异。一般以为,印度以东的各个种族,胡须稀疏,尽管如此,在日本列岛最北方居住的虾夷人(Ainos)(如图)却是世界上最多毛的人。

▲达尔文在谈到“美对决定人类婚姻的影响”时,列举了世界各地对于美丑的不同标准。他认为,完全的美意味着许多性状都以一种特殊方式发生改变,这在每一个种族中大概都是奇迹。

▲达尔文认为,导致人类种族之间在外貌上有所差别的所有原因,以及人类和低于人类的动物之间在某种程度上有所差别的所有原因,其中最有效的乃是性选择。

▲达尔文的表弟弗朗西斯·高尔顿(1822—1911),一生都执著地热衷于有关杰出思想家的遗传谱系统计研究。图为他建立的“人体测量实验室”。当达尔文在人类学研究方面保持沉默期间,高尔顿建立了偏离达尔文主义的第二大支流,创立了所谓的“优生学”,但他对达尔文理论诠释得面目全非,所以,终在1871年被达尔文否决。
▲鱼、火蜥蜴、龟、鸡、猪、牛、兔、人类的胚胎发育比较。从图中可见,人类和许多低等动物的早期胚胎相似。

▲今天,在实验室里,对“进化论”的研究仍然在继续。随着科技的进步,达尔文曾推断出所有生物体是“从某一个原型细胞遗传下来的”,这种尝试性的结论得到了有效论证。
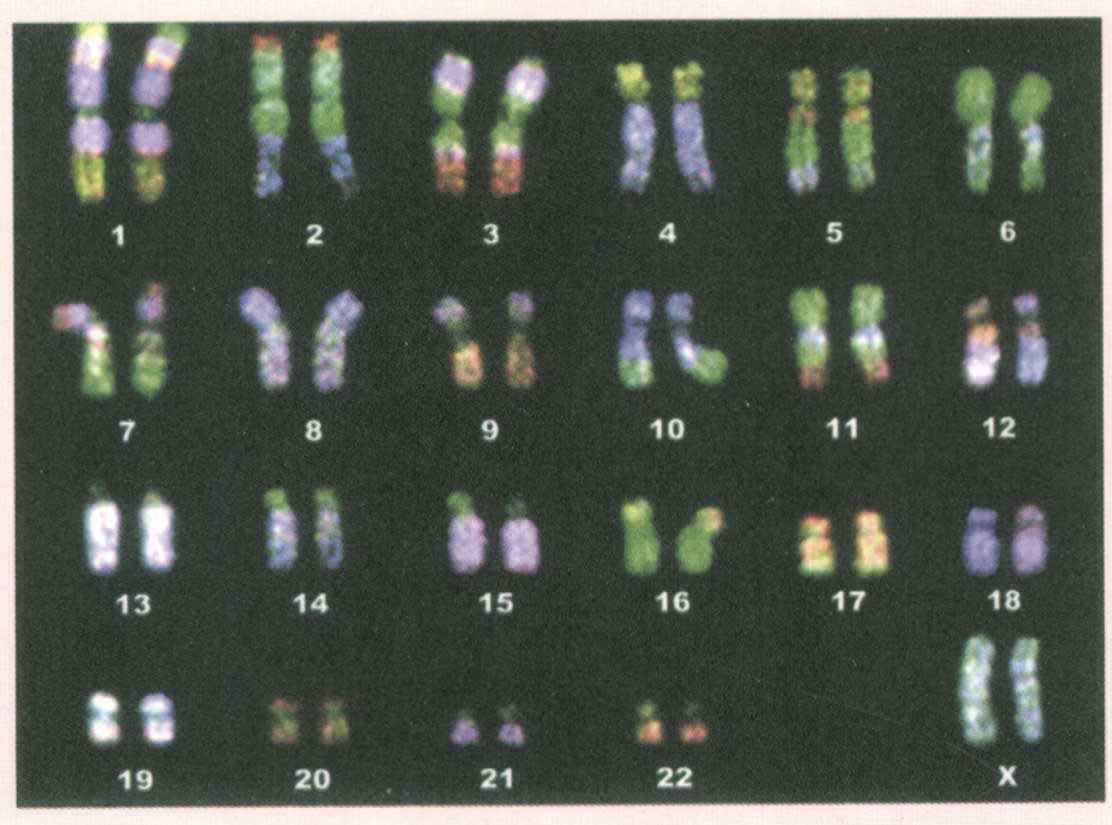
▲人类染色体图 研究发现,人类与黑猩猩的同源性超过了95%,在基因数、基因结构与功能、染色体与基因组构造上,人类和黑猩猩几乎相同。但在漫长的进化过程中,这不到5%的差别,决定了人类和黑猩猩的根本区别。
进化论没有停滞不前,20世纪和21世纪的许多生物学家在达尔文和华莱士的原始理论基础上,又增加了有关突变和遗传机制方面的新知识。正在兴起的地球历史理论,对现代进化论也有贡献,例如古生物学家古尔德(Stephen Jay Gould,1941—2002)和艾尔德里奇(Niles Eldredge,1943一)。

▲达尔文港 早在1839年达尔文随“贝格尔号”航行到澳大利亚一个无名港湾时,船长将之命名为“达尔文港”,1869年这个名字得到了官方认可,图为今日的达尔文港。

▲1964年,为了纪念达尔文家族而在剑桥大学建成的达尔文村。

▲▲2009年是达尔文诞辰200周年,《物种起源》发表150周年,为此,世界各地都在举办达尔文诞辰庆祝活动。就连那些反对达尔文的人也承认:达尔文是一名伟人,他显然很重要,他的思想改变了世界。图为著名的《柳叶刀》杂志以Darwin’s Gifts为主题的封面和插画。

▲自2006年起,以达尔文为主题的展览就先后在纽约、波士顿、多伦多相继举办,2009年伦敦自然历史博物馆(Natural History Museum)将此展览作为全世界纪念达尔文诞辰200周年的活动之一。在达尔文的故乡,更是以“Darwin’s Shrewsbury 2009 Festival”庆祝。

▲2009年,英国为纪念达尔文诞辰及《物种起源》发表而铸造的2英镑钱币。
————————————————————
(1) 韦斯特伍德,《昆虫的近代分类》,第2卷,1840年,541页,下面关于Tanais的描述得到了弗里茨·米勒(Fritz Müller)先生的协助。
(2) 柯尔比与斯彭斯(Kirby and Spence),《昆虫学导论》,1826年,309页。
(3) 《新西兰的鸟类》(Birds of New Zealand),1872年,66页。
(4) 佩里埃(M.Perrier)提出这个例子[见《科学评论》(Revue Scienti fique),1873年2月1日,865页]以为这是对性选择信念有决定性的,因为他猜想我把全部性差异都归因于性选择的作用。因此这位著名的博物学者,像许多其他法国人一样,即使对性选择最初的那些原则也很容易理解。有一位英国博物学者坚持认为某些雄性动物的抱握器不是通过雌者选择而发展起来的。如果我没有遇到佩里埃这个论述,我想任何人读了本章之后,都不会认为我所主张的雌者选择对雄者抱握器官的发展有所帮助是可能的。
(5) 艾伦,《佛罗里达的哺乳类和冬鸟类》(Mammals and Winter Birds of Florida),见《比较动物学学报》,哈佛大学出版,268页。
(6) 即使那些雌雄异株的植物,其雄花一般也比雌花早熟。正如斯普林格尔(C.K.Sprengel)最先指出的,许多雌雄同株的植物都是异花授粉的,这就是说,其雄性器官和雌性器官不在同一时间成熟,因此这些植物不可能自花授粉。现在这种花的花粉一般比柱头先成熟,虽然也有雌性器官先成熟的例外。
(7) 下面是关于后代性状的有力证据。一位有经验的鸟类学家艾伦先生(见《佛罗里达的哺乳类和冬鸟类》,229页)讲到在最先孵出的雏鸟遭到了意外毁灭之后才晚期出生的同窝雏鸟时说道,它们“比在该季节较早孵出的雏鸟,个子比较小,色彩也较暗淡。假如每年都要孵出若干窝的雏鸟来,那么一般说那较早成窝孵出的雏鸟在所有方面都显得最完善和精力充沛”。
(8) 赫尔曼·米勒(Hermann Müller)关于那些每年最先羽化的雌蜂,也得出相同的结论。参阅他的著名论文《达尔文学说在蜜蜂中的应用》(Anwendung den Darwin'schen Lehre auf Bienen),见《动物比较解剖学年刊》(Verh.d.V.Jahrg.),第29卷,45页。
(9) 关于家鸡,我曾收到有关这一效果的报告,将在以后举出。甚至关于鸟类,像终生配偶的鸽子,我听詹纳.韦尔先生说,当雄鸽受伤或衰弱了之后,雌鸽也会把它遗弃而去。
(10) 属阔鼻类,卷尾猴科。形小,拇指发达,面部肉色,头部与四肢皆呈黑褐色。尾长,末端卷曲,尾善缠绕,栖于美洲森林中。悲哀时,发一种泣声,故又名“泣猴”(weeper capuchin)。体有麝香气,亦名“麝猴”(musk monkey)。——译者注
(11) 关于大猩猩,参阅萨维奇(Savage)和怀曼的文章,见《波士顿博物学杂志》,第5卷,1845—1847年,423页。关于鼯猴(Cynocephalus),参阅布雷姆的《动物生活图解》(Illust.Thierieben),第1卷,1864年,77页。关于吼猴,参阅伦格尔的《巴拉圭哺乳动物志》,1830年,14,20页。关于卷毛猴,参阅布雷姆的著作,同前书,108页。
(12) 帕拉斯,《动物学专论》(Spicilegia Zoolog.,Fasc.),第12卷,1777年,29页。安德鲁·史密斯爵士,《南非动物学图解》(Illustrations of the Zoology of S.Africa),1849年,29页。欧文在其《脊椎动物解剖学》中(第3卷,1868年,633页)列举了一个表,附带指出羚羊的哪一个种是营群居生活的。
(13) 坎贝尔博士,《动物学会会报》,1869年,138页。再参阅海军上尉约翰斯东(Johnstone)所写的有趣论文,见《孟加拉亚洲学会会报》(Proc.Asiatic Soc.of Bengal),1868年5月。
(14) 格雷,《博物学年刊杂志》(Annals and Mag.of Nat.Hist.),1871年,302页。
(15) 参阅多布森博士(Dr.Dobson)的出色文章,见《动物学会会报》,1873年,241页。
(16) 见他所写的有关海豹(The Eared Seals)一文,《美国博物学者》,第4卷,1871年1月。
(17) 即Turdus merula L.,多产于欧洲及北美。——译者注
(18) 即Pyrrhula pyrrhula。——译者注
(19) 关于普罗戈尼黑羽长尾鸟,参阅《彩鹳》(TheIbis),第3卷,1861年,133页;关于另一种黑羽长尾鸟(Vidua axillaris),参阅同一著作,第2卷,1860年,211页;关于雷鸡和硕鸨,参阅劳埃德(L.Lloyd)的《瑞典的猎鸟》(Game Birds of Sweden),1867年,19,182页。蒙塔古(Montagu)和塞尔比(Selby)说,黑松鸡是多配性的,而红松鸡则是单配性的。
(20) 诺埃尔·汉弗莱斯(Noel Humphreys),《水上公园》(River Gardens),1857年。
(21) 柯尔比与斯彭斯,《昆虫学导论》,第3卷,1826年,342页。
(22) 一种寄生的膜翅目昆虫(韦斯特伍德,《昆虫的近代分类》,第2卷,160页)是这个规律的例外,其雄虫具有残迹的翅,从来不离开它出生的那个小穴,而雌虫却有发育良好的翅,奥杜安认为这个物种的雌虫是由出生在同一小穴的雄虫来授精的,但远为可能的是雌虫跑到别的小穴以避免近亲交配。我们以后还会在各个动物网中碰到少数例外,即追求者和求偶者是雌者,却不是雄者。
(23) 欧文(Owen)编,《观测报告和论文集》(Essays and Observations),第l卷,1861年,194页。
(24) 萨克斯(Sachs)教授《植物学理论》(Lehrbuch der Botanik,1870,633页)在谈到雄性和雌性的生殖细胞时说道:“其一在结合时是自动的,另一在结合时似为被动的。”
(25) 《关于家畜饲养的报告》(Vortrage über Viehzucht),1872年,63页。
(26) 《诺瓦拉游记》(Reise der Novara):人类学部分,1867年,216—269页。魏斯巴赫(Weisbach)博士根据希和译和许瓦茨两位博士的观测材料计算出来的结果。关于雄性家养动物具有较大的变异性,参阅我的《动物和植物在家养下的变异》,第2卷,1868年,75页。
(27) 《皇家学会会报》,第16卷,1868年7月,519,524页。
(28) 《爱尔兰皇家学会会报》,第10卷,1868年,123页。
(29) 《马塞诸塞医学会会刊》,第2卷,第3期,1868年,9页。
(30) 《病理学、解剖学和生理学文献集》(Archiv für Path.Anat.und Phys.),1871年,488页。
(31) 斯托克顿·霍夫(J.Stockton Hough)博士最近关于人类体温所做的结论见《通俗科学评论》,1874年1月1日,97页。
(32) 曼特加沙(Mantegazza)教授认为[《致达尔文先生的一封信》,见《人类学文献集》(Archivio per l'Anthropologia),1871年,306页],许多雄性动物常见的鲜艳色彩是由于它们产生并保存了精液的缘故,但这情况简直是不可能的,因为许多公鸟如小公雉在它们出生第一年的秋季其色彩就变鲜艳了。
(33) 关于人类,参阅霍夫博士的结论,见《通俗科学评论》,1874年,97页。参阅吉拉德(Girard)关于鳞翅目的观察材料,见《动物学记录》(Zoological Record),1869年,347页。
(34) 《佛罗里达的哺乳类和鸟类》,234,280,295页。
(35) H.米勒,《达尔文学说在蜜蜂中的应用》,见《动物比较解剖学年刊》,第29卷,42页。
(36) 《动物和植物在家养下的变异》,第2卷,1868年,75页。曾对上述泛生论的假说做过充分解说。
(37) 这些事实是根据一位伟大饲养家蒂贝(Teebay)先生的材料提出来的,见特格梅尔(Tegetmeier)的《家禽手册》(Poultry Book),1868年,158页。以下一节所提到的有关不同品种小鸡的性状以及鸽子的品种,参阅《动物和植物在家养下的变异》,第1卷,160,249页;第2卷,77页。
(38) 《关于四足兽的新种》(Novae species Quadrupedum e Glirium ordine),1778年,7页。关于马的毛色的传递,参阅《动物和植物在家养下的变异》,第1卷,51页。第2卷,71页上有关于“限于性别的遗传性”的一般探讨。
(39) 夏普伊(Chapuis)博士,《比利时信鸽》(Le Pigeon Voyageur Belge),1865年,87页。布瓦塔尔和科尔比(Boitard et Corbié),《家鸽》(Les Pigeons de Volière),1824年,173页。关于摩德纳地方某些品种的同样差异,再参阅保罗·旁尼兹(Paolo Bonizzi)著,《家鸽的变异》(Le variazioni dei Colombi domestici),1873年。
(40) 自从本书第一版问世以后,我感到高度满意的是见到了特格梅尔(Tegetmeier)先生这么有经验的育种家的下述见解(《田野》(Field),1872年9月)。他描述的一些奇妙事例表明,有些鸽子的羽色传递只限于一性,并且形成了一个具有这种性状的亚品种,然后他又说:“达尔文先生提出了用人工选择的方法改变鸟类性别的颜色的可能性,当他这么做的时候,并不知道我所提到的这些事实,然而值得注意的是他提出了多么符合实际程序的正确方法。”
(41) 参阅我著的《动物和植物在家养下的变异》,第2卷,72页。
(42) 即Rangifer tarandus,属反刍偶蹄类,鹿科。产于亚、美、欧三洲的北极地,栖地愈北者体躯愈大。雌雄皆有角,雄者角大,长达五尺左右,角顶生枝,枝端扁平如锹。雌者角较小,分枝亦少。原系野生,性易驯,北极人民多养之,教以曳橇,行进甚速。驯鹿的化石多产于欧洲洪积层,角多分枝,其断面为椭圆形。——译者注
(43) 我很感激卡波勒斯(Cupples)先生为我向布雷多尔本侯爵夫人(Marquis of Breadalbane)手下的有经验的首席林务官罗伯逊(Robertson)先生作了有关苏格兰的公獐和马鹿(red deer)的调查。我感谢艾顿(Eyton)先生以及其他人士为我提供了有关黇鹿(fallow-deer)的材料。关于北美的一种驼鹿(Cervus alces),见《陆地与水》(Land and Water),1868年,221,254页;有关该大陆的C.Virginianus和C.Strongyloceros.参阅凯顿(J.D.Caton)的文章,见《渥太华学院自然科学学会会报》,1868年,13页。关于勃固(Pegu)的Cervus Eldi,参阅比万(Beavan)中尉文章,见《动物学会会报》,1867年,762页。
(44) 学名为Antilocapra Americana.我感谢坎菲尔德(Canfield)博士提供的有关母鹿的角的材料,另参阅他在《动物学会会报》发表的文章(1866年,109页)。还有再参阅欧文的《脊椎动物解剖学》,第3卷,627页。
(45) 我确信北威尔士绵羊的角在出生时完全可以觉察到,有时竞长达一英寸。尤雅特(Youatt)说(《牛》,1834年,277页)牛在出生时其额骨突起已穿入表皮,角质物很快在其上形成。
(46) 我非常感激维克托·卡鲁斯(Victor Carus)教授为我向最高权威人士作了有关萨克森的美利奴绵羊的调查。然而非洲几内亚海岸有一个绵羊品种,同美利奴绵羊一样,只有公羊才生角;而温伍德·里德先生告诉我说,他看到过的一个例子表明,一只小公羊产于2月10日,在3月6日就初现羊角,此例同上述规律相符,其角的出现晚于雌雄二者皆生角的威尔士绵羊。
(47) 《鸟类头骨的骨质突起》(Ueber die kn chernen Sch
chernen Sch delh
delh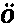 cker der V
cker der V gel),见《荷兰的动物学文献》(Niederlandischen Archiv für Zoologie),第1卷,第2册,1872年。
gel),见《荷兰的动物学文献》(Niederlandischen Archiv für Zoologie),第1卷,第2册,1872年。
(48) 普通孔雀(Pavo cristatus)只有公的有距,而爪哇绿孔雀(P.muticus)提供了一个反常事例,即公的母的都有距。因此我完全预料到后面这个物种的距在发育时期上要早于普通孔雀;但阿姆斯特丹的赫格特(M.Hegt)告诉我说,1869年4月23日曾对上一年出生的这两个物种的雏鸟作了比较,它们距的发育方面没有差别。然而它们的距至今只表现为小瘤或突起。我认为以后如果观察到发育速率有什么不同,我一定会得到这方面的报告。
(49) 在鸭科的一些其他物种中,公鸭和母鸭羽毛灿点的差别更大;但我未能发现这些物种的公鸭羽毛灿点的充分发育时期是否晚于普通野公鸭,按照我们的规律,应该如此。然而,有一个亲缘关系相近的物种秋沙鸭(Mergus cucullatus)的情况就是这样,雌雄二者不仅在一般羽衣上差异显著而且在羽毛灿点上也有相当大的差异,公鸭的羽毛灿点是全白色的,母鸭的羽毛灿点则是灰白色的。小公鸭最初同母鸭完全相似,其羽毛灿点也是灰白色的,这些灿点变为纯白色的时期要在成年公鸭获得其他更加强烈显著的性差异之前,参阅奥杜旁(Audubon)的《鸟类志》(Ornithological Biography),第3卷,1835年,249—250页。
(50) 《鸽类饲养通论》(Das Ganze der Taubenzucht),1837年,21,24页。关于有条纹的鸽子,参阅夏普伊的《比利时信鸽》,1865年,87页。
(51) 关于某些鸡的品种的细节以及有关所有这方面的参考材料,请参阅《动物和植物在家养下的变异》,第1卷,250,256页。关于高等动物,由家养引起的性差异,均以各个物种为题在该书中有所描述。
(52) 在《中央注册处1866年29号年报》第七部分有一张特别的十年统计表。
(53) 关于挪威与俄国的材料,见费伊教授研究报告的摘要,刊于《英国和外国外科学评论》(British and Foreign Medico-Chirurg.Review),1867年4月,343,345页。关于法国的材料,见《男女年龄年鉴》(Annuaire pour l'An),1867年,213页。关于费城的材料,见斯托克顿·霍夫博士文章,登于《社会科学协会》,1874年。关于好望角的材料,祖特文(H.H.Zouteveen)博士在这本书的荷译本中引用了奎特列特(Quetelet)的文章,其中提供了很多有关性别比例的材料。
(54) 关于犹太人的材料,见蒂里(M.Thury)的文章,登于《男女出生数的规律》(La Loi de Production des Sexes),1863年,25页。
(55) 《英国和外国外科学评论》,1867年4月,343页。斯塔克(Stark)博士也注意到(《苏格兰关于出生、死亡等的第十号年报》,1867年,第28部分)这个情况,即“这些例子足以表明几乎在生命的每个阶段,苏格兰的男性都更易死亡,死亡率均比女性高。男女两性在衣、食及一般福利都一样的情况下,上述特性在婴儿时期表现最强烈,此事实似乎证明男性死亡率高是给人以深刻印象的限于性别的一种天然固有特性”。
(56) 《约克郡西部行政区疯人院报告》,第1卷,1871年,8页。辛普森(J.Simpson)爵士已证明男婴的头周长要超过女婴1/3~1/8英寸。奎特利特已证明女人生下来要比男人小,见邓肯(Duncan)博士文章,登于《生殖力、多产与不孕》,1871年,382页。
(57) 根据精确的阿扎拉(Azara)的材料见《南美航游记》(Voyages dans l'Amérique merid第2卷,1809年,60,179页),巴拉圭未开化的瓜拉尼人(Guaranys)的男女比例为13∶14。
(58) 巴贝季(Babbage),见《爱丁堡科学杂志》(Edinburgh Journal of Science),第1卷,1829年,88页;有关死产的婴儿,见90页。关于英国私生子,见《中央注册处1866年报告》绪言,15页。
(59) 洛伊卡特(Leuekart),瓦格纳(Wagner)在《物理学手册》(Handw rterbuch der Phys.)中引用,第4卷,1853年,774页。
rterbuch der Phys.)中引用,第4卷,1853年,774页。
(60) 见《费城社会学学会》(Social Science Assoc.of Philadelphia),1874年。
(61) 《人类学评论》,1870年4月,108页。
(62) 有一份关于十一年间不孕母马和早期流产母马数字的记录,此事值得注意,因为它表明了这些营养极良的和近亲交配的动物已经变得何等不育,以致有将近三分之一的母马不能生产活驹。例如,在1866年,生下了809匹公驹,816匹母驹,并且有743匹母马不育。1867年,生下了836匹公驹,902匹母驹,并且有794匹母马不育。
(63) 我很感激卡波勒斯先生为我取得上述苏格兰的统计材料,以及下述有关牛的统计材料。莱伍德(Laighwood)的埃利奥特先生最先使我注意到雄者的早期夭折,——后来艾奇逊(Aitchison)先生以及其他人士证实了这一叙述。我感谢艾奇逊先生和佩安(Payan)先生,他们向我提供了有关绵羊的大量统计材料。
(64) 贝尔,《英国四足兽史》(History of British Quadrupeds),100页。
(65) 见《南非动物学图解》(Illustrations of the Zoology of S.Africa),1849年,第29图。
(66) 布雷姆得出相同的结论,见《动物生活图解》(Illust.Thierleben),第4卷,990页。
(67) 根据劳埃德的权威意见,见《瑞典的猎鸟》,1867年,12,132页。
(68) 《塞尔旁的博物学》(Nat.History of Selborne),第1卷,第29版,1825年,139页。
(69) 詹纳·韦尔先生于次年进行调查,也收到了类似报告。为了举出活捉的苍头燕雀数字,我愿提一下有两位能手在1869年搞了一次比赛,其中一位每天捉到62只公苍头燕雀,另一位捉到40只。有一个人一天捉到公苍头燕雀的最大数量是70只。
(70) 《彩鹳》,第2卷,260页,高尔得在其《蜂鸟科》(Trochilidae)一书(1861年,52页)中曾加以引用。关于上述的比例数,我感谢沙尔文先生提供的一张有关他研究结果中的一张表。
(71) 《彩鹳》,1860年,137页;以及1867年,369页。
(72) 《彩鹳》,1862年,187页。
(73) 洛伊卡特引用布洛克(Bloch)的材料(瓦格纳,《物理学手册》,第4卷,1853年,775页),表明鱼类的雄者为雌者的二倍。
(74) 在《农夫》杂志中引用,1869年3月18 日,369页。
(75) 《斯托蒙特菲尔德的养鱼实验》(The Stormontfield Piscicultural Experiments),1866年,23页。《田野新闻》,1867年6月29日。
(76) 《陆地与水》,1868年,41页。
(77) 雅列尔(Yarrell),《英国鱼类志》(Hist.British Fishes),第1卷,1826年,307页。关于鲤鱼(Cyprinus carpio)和丁 鱼(Tinca vulgaris),见331页。关于欧鳊(Abramis brama),见336页。关于鱥雅罗鱼(Leuciscus phoxinus),参阅《伦敦博物学杂志》,第5卷,1832年,682页。
鱼(Tinca vulgaris),见331页。关于欧鳊(Abramis brama),见336页。关于鱥雅罗鱼(Leuciscus phoxinus),参阅《伦敦博物学杂志》,第5卷,1832年,682页。
(78) 留卡特引用迈内克(Meinecke)的材料(瓦格纳,《物理手册》,第4卷,1853年,775页),表明蝴蝶的雄虫数量为雌者的三四倍。
(79) 《亚马孙河上的博物学者》,第2卷,1863年,228,347页。
(80) 特里门先生在其《南非的锤角虫亚目》(Rhopalocera Africa Australis)一书中举出了这些例子中的四个。
(81) 特里门引用的材料,见《昆虫学会会报》,第5卷,第四部分,1866年,330页。
(82) 《林奈学会会报》,第25卷,37页。
(83) 《昆虫学会会报》,1868年2月17日。
(84) 华莱士博士引用材料,见《昆虫学会会报》,第5卷,第3辑,1867年,487页。
(85) 布朗夏尔(Blanchard),《昆虫的变态及其习性》(Métamorphoses,Maeurs des Insectes),1868年,225—226页。
(86) 《鳞翅目价目表》(Lepidopteren-Doubletten Liste),柏林,第10号,1866年。
(87) 这位热心的博物学家送给我一些数年前的研究结果,其中表明雌者数量似乎超过雄者;但由于好多数字都是估计的,因此我觉得不可能将它们列制成表。
(88) 冈瑟的《动物学文献记录》(Record of Zoological Literature),1867年,260页。关于雌锹甲虫占有数量优势,见同书,250页。关于英格兰的雄锹甲虫,参阅韦斯特伍德的文章,见《昆虫的近代分类》,第1卷,187页。关于扁蟞,见同书,172页。
(89) 沃尔什,《美国昆虫学家》,第1卷,1869年,103页。史密斯,《动物学文献记录》,1867年,328页。
(90) 《农业昆虫》(Farm Insects),45—46页。
(91) 《达尔文学说在蜜蜂中的应用》,第24卷。
(92) 《飞蝗的飞迁线》(Die Strich,Zug oder Wanderheuschrecke),1828年,20页。
(93) 海根(H.Hagen)和沃尔什著,《对北美脉翅目昆虫的观察》(Observationson N.American Neuroptera),见《费城昆虫学会会报》,1863年10月,168,223,239页。
(94) 《伦敦昆虫学会会报》,1868年2月17日。
(95) 关于这个纲的另一位大权威,乌普萨拉的托列尔(Thorell)教授说[《关于欧洲蜘蛛》(On European Spiders),第一部分,1869—1870年,205页],雌蜘蛛好像一般要比雄者更常见。
(96) 关于这个问题,参阅坎布里奇(O.P.Cambridge)先生的材料,在《科学季刊》引用,1868年,429页。
(97) 《有关孤雌生殖文献》(Beitr ge zur Parthenogenesis),174页。
ge zur Parthenogenesis),174页。
(98) 《托达人》,1873年,100,111,194,196页。
(99) 尼尔吉里(Nilgiri)山地的游牧部落。——译者注
(100) 《政府报告,新西兰的原始居民》(Aboriginal Inhabitants of New Zealand;Government Report),1859年,36页。
(101) 同上,30页。
(102) 同上,26页。
(103) 见《夏威夷游记》(Narrative of a Tour through Hawaii),1826年,298页。
(104) 《桑威奇群岛史》(History of the Sandwich Islands),1843年,93页。
(105) 这项材料引自祁佛牧师的《在桑威奇群岛的生活》(Life in the Sandwich Islands)一书,1851年,277页。
(106) 库尔特(Coulter)博士在描述(《皇家地理学会学报》,第5卷,1835年,67页)加利福尼亚州1830年左右的情况时说道,当地土著居民虽然受到良好待遇,也没有从故乡被撵走,并且禁止他们饮酒,但在西班牙教士的教化下,几乎全部濒于灭亡或正在灭亡。他把这个现象主要归因于男人数量远远超过女人这一无可怀疑的事实;但他不知道这是否由于缺少女性后代还是由于女婴在幼年时期死亡较多。后一假定按照所有推论都是极不可能的。他接着说,“人们所恰当地称之为杀婴的事虽不流行,但流产却为常见”。如果库尔特博士关于杀婴问题的说法是正确的,那么这个例子就不能支持马歇尔上校的观点。从被教化的土著人口急剧减少的情况来看,我们可以猜想这和刚才举出的例子一样,他们生育力的降低是由于生活习惯的改变。
关于这个问题,我曾希望从狗的繁育中找到一点说明:因为在大多数狗的品种中,也许细躯猎狗是例外,被杀死的小母狗比小公狗的数量多得多,托达人的幼婴情况也恰恰如此。卡波勒斯先生向我确言,苏格兰猎鹿狗的情况通常就是这样。不幸的是,除了细躯猎狗外,我对其他任何品种的雌雄比例均毫无所知,而细躯猎狗生出的公狗和母狗之比为110.1∶100。现在根据向许多养狗人所做的调查表明,母狗似乎在某些方面更受重视,虽然在其他方面不受欢迎;最优良品种小母狗有计划的被杀死看来并不比小公狗为多,虽然有时确乎在有限制的范围内这样实行过。因此能否根据上述原则来解释细躯猎狗生出的公狗数超过母狗,我还无法决定。另一方面,我们已经知道,关于马、牛和绵羊,由于幼畜的任何一种性别的价值都很高,所以没有随便宰杀的现象,如果其比例有什么差别的话,那就是雌者稍微超过了雄者。
(107) 《动物学实验文献集》(Archives de Zoolog.Expér.),1872年10月,563页。
(108) 《关于纲之下的物种》(De l'Espèce et de la Class.)&c.,1869年,106页。
(109) 例如,参阅我的有关记载,见我著的《研究日志》(Journal of Researches),1845年,7页。
(110) 我举出过一个奇妙例子(《关于火山岛的地质考察》,1844年,53页)示明光线对一种在阿森松岛海岸沉积的叶状外壳颜色所产生的影响,这种沉积是由于拍岸浪向海岸岩石的冲击,并由磨碎了的海贝壳的溶液而形成的。
(111) 莫尔斯(Morse)博士最近对这个问题进行了讨论,见他写的《关于软体动物的适应性的色彩》的论文,载于《波士顿博物学会学报》,第14卷,1871年4月。
(112) 参阅他的专题著作《英国环节动物类》(British Annelids),第一部分,1873年,3页。
(113) 参阅佩里埃(M.Perrier)的《达尔文以后有关人类起源的讨论》(l'Origine de l'Homme d'après Darwin),见《科学评论》,1873年2月,866页。
(114) 《支持达尔文的事实和论据》(Facts and Arguments for Darwin),英译本,1869年,20页。参阅上述关于嗅觉线状体的讨论。萨斯(Sars)曾描述过一个多少类似的例子,是关于一种挪威甲壳动物的,即Pontoporeia af finis(《自然》,1870年,455页引用)。
(115) 参阅卢伯克爵士的文章,见《博物学年刊杂志》,第11卷,1853年,1、10图;第12卷,1853年,7图。再参阅卢伯克的文章,见《昆虫学会会报》,新刊第4卷,1856—1858年,第8页。至于下面提到的锯齿形触角,参阅弗里茨·米勒的《支持达尔文的事实和论据》,1869年,40页,脚注。
(116) 参阅斯彭斯·巴特先生一篇附有图解的论文,见《动物学会会报》,1868年,363页;以及关于属的系统命名法一文,同前刊,585页,我以上几乎全部有关高等甲壳动物的螯的论述,均承斯彭斯·巴特先生的大力协助。
(117) 《甲壳动物志》(Hist.Nat.des Crust.),第2卷,1837年,50页。
(118) 一名“巢螺”第一对肢形大为钳状,其在右方者常比左方大,全体赤色或苍黑,栖于海滨,寄居螺类遗壳之中,故名。其右方的大螯用以步行和采食,当退居螺壳中时并用它掩蔽螺壳之口。——译者注
(119) 斯彭斯·巴特先生著文,见《大不列颠协会,关于南部德文郡动物区系的第四个报告》。
(120) 弗里茨·米勒,《支持达尔文的事实和论据》,1869年,25—28页。
(121) 短尾类为甲壳类的一个亚目,腹部短而曲屈,密着于头胸部下,第一对肢大而具螯,次四对小而有爪,或最后一对肢端广阔,变为游泳器,如蟹等属之。——译者注
(122) 为甲壳类的一个亚纲,体型小,当蜕皮为成虫时,有肢三对,为感觉、取食和游泳之用,如水蚤等属之。——译者注
(123) 《巴西腹地纪游》(Travels in the Interior of Brazil),1846年,111页。在我的《研究日志》的第463页有关于椰子蟹习性的记述。
(124) 参阅弗雷泽(Ch.Fraser)先生的文章,见《动物学会会报》,1869年,3页。承巴特先生给我提供了鲍尔博士的记述。
(125) 属于节肢动物,甲壳类,金鱼虫、红虫属之。——译者注
(126) 克劳斯(Claus),《关于桡足类的游动生活》(Diefreilebenden Copepoden),1863年,35页。
(127) 《支持达尔文的事实和论据》,79页。
(128) 下述事实见《大不列颠蜘蛛志》(A History of the Spiders of Great Britain),1861—1864年,77,88,102页。
(129) 这位作者最近发表了一篇《关于蛛形纲的次级性征》的有价值论文,见《帕多瓦博物学的威尼托-特兰提诺协会会报》(Atti della Soc.Veneto-Trentina di Sc.Nat.Padova),第1卷,第3册,1873年。
(130) 奥古斯特·万松(Aug.Vinson)在《岛屿蜘蛛类》(Aranéides des Iles de la Réunion),第六幅插图的图1和图2,书中提供了一个良好例子来说明雄黑蜘蛛(Epeira nigra)的小型身体。我可以补充一点,这个物种的雄者是黄褐色的,而雌者是黑色的并且腿上具有红色带状花纹。根据记载,关于雌雄二者大小的不相等,甚至还有更显著的事例(《科学季刊》,1868年7月,429页),但我没有见到原始材料。
(131) 柯尔比和斯彭斯,《昆虫学导论》,第1卷,1818年,280页。
(132) 《动物学会会报》,1871年,621页。
(133) 球腹蛛类,锯齿形球腹蛛,四星球腹蛛,斑点球腹蛛:参阅韦斯特林的著文,见克罗耶尔(Kroyer)的Naturhist.Tidskrift,第4卷,1842—1843年,349页;以及第2卷,1846—1849年,342页。关于其他物种,再参阅《瑞典蜘蛛类》(Araneae Suecicae),184页。
(134) 祖特文博士在这本著作的荷兰文译本中收集了一些事例(第1卷,444页)。
(135) 然而,希尔根道夫(Hilgendorf)最近引起人们注意某些高等甲壳动物的相似构造,该构造似乎适于发出声音:见《动物学记录》,1869年,603页。
(136) 瓦尔克纳和热尔威,《昆虫志:无翅目》(Hist.Nat.des Insectes:Apteres),第4卷,1847年,17,19,68页。
(137) 卢伯克爵士,《林奈学会会报》,第25卷,1866年,484页。关于蚁蜂科,参阅韦斯特伍德的《昆虫的近代分类》,第2卷,213页。
(138) 在亲缘关系密切接近的物种中,雄虫的这等器官往往不一样,并呈现最显著的物种性状。但从功能的观点来看,其重要性正如麦克拉克伦先生对我说过的,大概被估计过高了。有人提出这等器官的轻微差异就足以阻止十分显著的品种、即端始物种之间的杂交,这就有助于它们的发展。但实际情况简直不会如此,根据不同物种也可结合的许多观察记录[例如,参阅勃龙的《自然史》(Geschichte der Natur),第2卷,1843年,164页;韦斯特伍德的文章,《昆虫学会会报》,第3卷,1842年,195页],我们可以这样推论。麦克拉克伦先生告诉我[《昆虫报合订本》(Stett.Ent.Zeitung),1867年,155页],迈耶博士曾把这种差异非常显著的石蛾科(Phryganidae)的某些物种关在一起,它们互相交配了,而且其中一对还产下了受精卵。
(139) 《实际昆虫学家》(The Practical Entomologist),美国,费城,第2卷,1867年5月,88页。
(140) 同上杂志,107页,参阅沃尔什先生的文章。
(141) 见《昆虫的近代分类》,第2卷,1840年,205,206页。沃尔什先生唤起我对颚之双重用途的注意,他说他反复观察到这类事实。
(142) 我们在这里见到一种奇特而费解的二态现象,因为龙虱属四个欧洲物种和Hydroporus的某些物种的一些雌虫具有光滑的鞘翅,在有槽或有刻点的鞘翅和十分光滑的鞘翅之间并没有见过中间的级进类型。参阅绍姆(H.Schaum)博士的材料,《动物学家》(Zoologist),第5—6卷,1847—1848年,1896页,曾被引用。此外,参阅柯尔比和斯彭斯所著《昆虫学导论》(Introduction to Entomology),第3卷,1826年,305页。
(143) 见韦斯特伍德,《昆虫的近代分类》,第2卷,193页。以下关于霉蛰(Penthe)的叙述以及其他引文均采自沃尔什先生的材料,见费城出版的《实际的昆虫学家》,第3卷,88页。
(144) 见柯尔比和斯彭斯,《昆虫学导论》,第3卷,332—336页。
(145) 《马德拉昆虫》(Insecta Maderensia),1854年,20页。
(146) 道布尔戴《博物学年刊杂志》,第1卷,1848年,379页。我应再补充一点:某些膜翅目昆虫的翅脉因性别而异。参阅沙卡德(Shuckard)的《掘土膜翅目昆虫》(Fossorial Hymenop.),1837年,39—43页。
(147) 见贝茨,《林奈学会会报杂志》(Journal of Proc.Linn.Soc.),第6卷,1862年,74页。翁弗尔(Wonfor)先生的观察材料在《通俗科学评论》中引用,1868年,343页。
(148) 见《博物学者在尼加拉瓜》,1874年,316—320页。关于有荧光的卵,见《博物学年刊杂志》,1871年11月,372页。
(149) 罗比内特(Robinet),《关于蚕丝》(Vers à Soie),1848年,207页。
(150) 《昆虫学会会报》,第5卷,第3辑,486页。
(151) 见《昆虫学会会报杂志》(Journal of Proc.Ent.Soc.),1867年2月4日,71页。
(152) 为现有体型最大的甲虫,属金龟子科,鳃角类,雄者头部生有巨大的角,有时胸部也生有一个或一个以上的角,产于美国的南部和西部。俗名犀甲虫(rhinoc eros beetle)。——译者注
(153) 属金龟子科,体长达5英寸,头部生有向上弯曲的叉形大角,产于中美。俗名象甲虫(elephant beetle)。——译者注
(154) 关于雌雄大小的这一叙述以及其他叙述,见柯尔比和斯彭斯的《昆虫学导论》,第3卷,300页;关于昆虫的寿命,见344页。
(155) 见《林奈学会会报》,第26卷,1868年,296页。
(156) 《马来群岛》(The Malay Archipelago),第2卷,1869年,313页。
(157) 参阅《昆虫的近代分类》,第2卷,1840年,526页。
(158) 参阅《动物比较解剖学年刊》,第29卷,80页。迈耶,《美国的博物学家》(American Naturalist),1874年,236页。
(159) 参阅洛恩先生有趣的著作《关于绿头苍蝇的解剖学》(On the Anatomy of the Blow-fly,Musca vomitoria),1870年,14页。他说(33页)“被捉住的蝇子发出一种特殊的哀鸣,这声音使其他蝇子都跑掉了”。
(160) 引自韦斯特伍德,《昆虫的近代分类》,第2卷,473页。
(161) 这些细节引自韦斯特伍德的《昆虫的近代分类》,第2卷,1840年,422页。关于白蜡虫科,参阅柯尔比和斯彭斯的《昆虫学导论》,第2卷,401页。
(162) 《动物科学杂志》(Zeitschrift für wissenschaft Zoolog.),第17卷,1867年,152—158页。
(163) 见《新西兰科学院院报》(Transact.New Zealand Institute),第5卷,1873年,286页。
(164) 承蒙沃尔什先生把哈特曼博士的《关于十七年蝉行为》(Journal of the Doings of Cicada septemdecim)的摘要送给我,对此谨表感激之意。
(165) 吉尔丁(L.Guilding),《林奈学会会报》,第15卷,154页。
(166) 我的这一叙述是根据克彭(K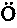 ppen)的《飞越俄国南部的蝗虫》(Ueber die Heuschrecken in Südrussland),1866年,32页,因我极力想弄到库尔特的原著,但未能如愿。
ppen)的《飞越俄国南部的蝗虫》(Ueber die Heuschrecken in Südrussland),1866年,32页,因我极力想弄到库尔特的原著,但未能如愿。
(167) 吉尔伯特·怀特(Gilbert White),《塞尔包内博物学》(Nat,Hist.of Selborne),第2卷,1825年,262页。
(168) 哈里斯,《新英格兰的昆虫》(Insects of New England),1842年,128页。
(169) 《亚马孙河上的博物学者》,第1卷,1863年,252页。贝茨先生对这三个科音乐器官的级进变化作了很有趣的讨论。再参阅韦斯特伍德的《昆虫的近代分类》,第2卷,445,453页。
(170) 见《波士顿博物学会学报》,第11卷,1868年4月。
(171) ⑺《新比较解剖学概论》(Nouveau Manuel d'Anat.Comp.),法译本,第1卷,1850年,567页。
(172) 《动物科学杂志》,第17卷,1867年,117页。
(173) 见韦斯特伍德的《昆虫的近代分类》,第1卷,440页。
(174) 《关于螽斯科的发音装置,达尔文主义的贡献》(Ueber der Tonapparat der Locustiden,ein Beitrag zum Darwinismus),《动物科学杂志》,第22卷,1872年,100页。
(175) 韦斯特伍德,《昆虫的近代分类》,第1卷,453页。
(176) 兰多依斯,《动物科学杂志》,第17卷,1867年,121,122页。
(177) 沃尔什先生还告诉我说,他曾注视过穴居扁叶 (Platyphyllum concavum)的雌虫“被捉住时它们的鞘翅就会在一起摩来摩去,发出一种微弱的摩擦声”。
(Platyphyllum concavum)的雌虫“被捉住时它们的鞘翅就会在一起摩来摩去,发出一种微弱的摩擦声”。
(178) 兰多依斯,《动物科学杂志》,第17卷,1867年,113页。
(179) 《新英格兰的昆虫》,1842年,133页。
(180) 韦斯特伍德,《昆虫的近代分类》,第1卷,462页。
(181) 兰多依斯最近发现了某些直翅目昆虫的残迹构造和同翅目昆虫的发音器官非常近似,这是一个令人吃惊的事实。参阅《动物科学杂志》,第22卷,第3册,1871年,348页。
(182) 《昆虫学会会报》,第2卷,第3辑(《会报杂志》,117页)。
(183) 见韦斯特伍德,《昆虫的近代分类》,第1卷,427页,有关蟋蟀的资料,见445页。
(184) 霍恩(Ch.Horne)先生,见《昆虫学会会报》,1869年5月,12页。
(185) 即雪白树蜂(OEcanthus nivalis),《新英格兰的昆虫》,1842年,124页。我听维克托·卡拉斯(Victor Carus)说,欧洲透明树蜂(OE.pellucidus)的雄虫和雌虫的差异也几乎一样。
(186) 即扁叶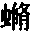 (Platyblemnus),见韦斯特伍德的《昆虫的近代分类》,第1卷,447页。
(Platyblemnus),见韦斯特伍德的《昆虫的近代分类》,第1卷,447页。
(187) ⑺沃尔什,《伊利诺伊的拟脉翅类昆虫》(Pseudoneuroptera of Illinois),见《费城昆虫学会会报》(Proc.Ent.Soc.of Philadelphia),1862年,361页。
(188) 《昆虫的近代分类》,第2卷,37页。
(189) 沃尔什,同上书,381页。以下有关Hetaerina,Anax和Gomphus的材料,是由这个博物学家提供的,谨此致谢。
(190) 《昆虫学会会报》,第1卷,1836年,81页。
(191) 参阅1867年《动物学纪录》的摘要,450页。
(192) 柯尔比和斯彭斯,《昆虫学导论》,第2卷,1818年,35页。
(193) 乌佐(Houzeau),《论智力》,第1卷,104页。
(194) 参阅一篇有趣的文章,《法布尔的著作》(The Writings of Fabre),见《博物学评论》,1862年4月,122页。
(195) 《昆虫学会会报杂志》,1863年9月7日,169页。
(196) 胡贝尔(Huber),《蚁类习性的研究》,1810年,150,165页。
(197) 《费城昆虫学会会报》,1866年,238—239页。
(198) ⑺《达尔文学说在蜜蜂中的应用》,见《动物比较解剖学年刊》,第29卷。
(199) 佩里埃,《达尔文以后的性选择研究》(la Sélection sexuelle d'après Darwin)(见《科学评论》,1873年2月,868页),他在这篇文章中显然未经深思熟虑就提出异议,认为既然社会性的雄蜂是公认由未受精卵产出来的,它们就不会把新性状传递给其雄性后代。这真是一个不同寻常的异议。一只雄蜂如果具有便于两性结合的某种性状或者具有更能吸引雌蜂的某种性状,那么,被它受精的雌蜂所产下的卵将只育出雌蜂;但这些新育成的雌蜂在第二年还会生育出雄蜂,能说这等雄蜂不会承继其祖父雄蜂的性状吗?让我们从普通动物中尽可能举出一个近似的例子:如果有一只雌白色四足兽或鸟同一只雄黑色品种杂交,杂种后代的雌雄二者又互交,能说这个杂种第二代不会把它们祖父的黑色倾向承继下来吗?不育的工蜂获得新性状是一个难得多的问题,但在我的《物种起源》一书中,我尽力阐明了这些不育的生物是如何被自然选择力量所左右的。
(200) 韦斯特伍德引用,见他写的《昆虫的近代分类》,第2卷,214页。
(201) 即Entimus imperialis。——译者注
(202) 贝茨先生在《昆虫学会会报》(1869年)第50页曾描述过锯天牛(Pyrodes pulcherrimus)雌雄二者的明显差异。我将举出少数我听到的例子来说明甲虫雌雄二者在颜色上的差异。柯尔比和斯彭斯(《昆虫学导论》,第3卷,301页)提到一种花萤(Cantharis)、油芫青(Meloe)、Rhagium和砖色天牛(Leptura testacea),后者雄虫呈砖瓦色,胸部黑色,而雌虫全身暗红色。后面这两种甲虫都属于天牛科。垂门和小沃特豪斯(Waterhouse.jun),二位先生向我说过两种鳃角组甲虫的情况,一种是缘甲类(Peritrichia),一种是斑金龟(Trichius),后者雄虫颜色比雌虫晦暗。长形郭公虫(Tillus elongatus)雄虫呈黑色,雌虫据说呈暗蓝色,胸部为红色。沃尔什先生说有一种负泥虫(Orsodacna atra),其雄虫也呈黑色,而雌虫(所谓O.ruficollis)的胸部则呈赤褐色。
(203) 《费城昆虫学会会报》,1864年,228页。
(204) 柯尔比和斯彭斯,《昆虫学导论》,第3卷,300页。
(205) 柯尔比和斯彭斯,《昆虫学导论》,第3卷,329页。
(206) 《昆虫的近代分类》,第1卷,172页;扁鳖,172页。在大不列颠博物馆我见过一只扁鳖的雄虫标本处于中间类型,因此这二态现象不是严格的。
(207) 《马来群岛》,第2卷,1869年,276页。赖利(Riley),《关于密苏里州昆虫的报告》(Report on Insects of Missouri),1874年,115页。
(208) 见《昆虫学杂志》(Entomological Magazine),第1卷,1833年,82页。关于这个物种的冲突问题,参阅柯尔比和斯彭斯的《昆虫学导论》,第3卷,314页,韦斯特伍德的《昆虫的近代分类》,第1卷,187页。
(209) 引自费舍尔(Fischer),见《博物分类学辞典》(Dict.Class.d'Hist.Nat.),第10卷,324页。
(210) 《法国昆虫学会年报》(Ann.Soc.Entomolog.France),1866年,默里(A.Murray)在其《旅行记》(1868年,135页)一书中曾加以引用。
(211) 韦斯特伍德,《昆虫的近代分类》,第1卷,184页。
(212) 沃拉斯顿(Wollaston),《关于某些鸣叫悦耳的象虫科》(Oncertain Musical Curculionide)。见《博物学杂志年刊》,第6卷,1860年,14页。
(213) 此类甲虫见鼠等小动物的尸体,则掘尸旁之土为穴,使之陷入土中而食之,故名。——译者注
(214) 兰多依斯,《动物科学杂志》,第17卷,1867年,127页。
(215) 克罗契(Crotch)先生曾送给我属于这三个科以及其他科的各种甲虫的许多制成标本和有价值的资料,对此我非常感激。他认为锯角甲虫的摩擦发音能力以前没有发现过。我也非常感激詹森(E.W.Janson)先生提供的资料和标本。我要补充一点,鼠形皮蠢(Dermestes murinus)会摩擦发音,是我的儿子F.达尔文先生发现的,但没有找到发音器官。查普曼(Chapman)博士最近描述过棘胫小蠹虫(Scolytus)是一种能摩擦鸣叫的昆虫,见《昆虫学家月刊》(Entomologist's Monthly Magazine),第6卷,130页。
(216) 引自席厄德(Schi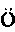 dte),其译文见《博物学年刊杂志》,第20卷,1867年,37页。
dte),其译文见《博物学年刊杂志》,第20卷,1867年,37页。
(217) 韦斯特林描述过[克罗耶尔,《博物学文集》(Naturhist.Tidskrift),第2卷,1848—1849年,334页]这两科以及其他科的摩擦发音器官。在步行虫科中,我检查了克罗契先生送给我的黏滑斑蝥(Elaphrus uliginosus)和Blethisa multipunctata。根据我所能作出的判断来说,Blethisa腹节绉边上的横向隆起线并没有刮拨鞘翅音锉的作用。
(218) 我感谢伊利诺斯州的沃尔什先生,蒙他赠我拉康德的《昆虫学导论》,101,143页的摘要。
(219) 布律勒里(M.P.de la Brulerie),默里的《旅行记》(第1卷,1868年,135页)曾加引用。
(220) 据道布尔戴先生说,“昆虫所发出的声音是靠着用后腿尽可能地把自己抬高,然后用胸部向坐在其下的东西连续五六次急速拍打”。关于这问题的参考资料,参阅兰多依斯的文章,见《动物科学杂志》,第17卷,131页。奥利维尔(Olivier)说(柯尔比和斯彭斯在其《昆虫学导论》,第2卷,395页,曾加引用)Pimelia striata的雌虫用其腹部向任何坚硬的东西拍打都可发出相当高的声音,于是雄虫“按照这种召唤,很快前来伴随着它,然后交配了”。
(221) 大彩虹蝶(Apatura Iris),见《昆虫学家周刊》,1859年,139页。关于婆罗洲蝶类,见科林伍德的《一个博物学家的漫谈》,1868年,183页。
(222) 参阅我的《研究日志》,1845年,33页。道布尔戴先生已发觉[见《昆虫学会会报》,1845年3月3日,123页]在前翅基部有一特别的膜质囊,大概同发音有关。关于黾蛾,见《动物学纪录》,1869年,401页。关于布坎南·怀特先生的观察材料,见《苏格兰博物学家》,1872年7月,214页。
(223) 参阅《苏格兰博物学家》,1872年7月,213页。
(224) 参阅《动物学记录》,1869年,347页。
(225) 再参阅贝茨先生的论文,见《费城昆虫学会会报》,1865年,206页。关于同样的问题,华莱士先生还对冠冕蠃(Diadema)进行过讨论,见《伦敦昆虫学会会报》,1869年,278页。
(226) 参阅《亚马孙河流域的博物学家》,第1卷,1863年,19页。
(227) 参阅《威斯敏特评论》,1867年7月,第10页上的一篇有趣文章。华莱士先生在《哈德威克的科学随笔》(Hardwicke's Science Gossip)一书上提供了一幅木叶蝶的木刻图,1867年9月,196页。
(228) 弗雷泽先生,《自然界》,1871年4月,489页。
(229) 《隔离对种类形成的影响》(Einfluss der Isolirung auf die Artbildung),1872年,58页。
(230) 参阅伍德先生写的、有趣的观察报告,见《学生》(The Student),1868年9月,第81页。
(231) 华莱士先生,《哈德威克的科学随笔》,1867年9月,193页。
(232) 关于这个问题再参阅韦尔先生的论文,见《昆虫学会会报》,1869年,23页。
(233) 《威斯敏特评论》,1867年7月,16页。
(234) 例如虎蛄蟖(Lithosia),但韦斯特伍德教授(见《昆虫的现代分类》,第2卷,390页)对这例子似乎感到惊奇。关于昼出和夜出的鳞翅目昆虫的相对色彩,见《昆虫的现代分类》,第2卷,333,392页;再参阅哈里斯著作《关于新英格兰昆虫类的论文》(Treatise on the Insects of New England),1842年,315页。
(235) 关于几个凤蝶种的翅膀上下表面的这等差别,见华莱士先生的《关于马来亚地区凤蝶科的研究报告》(Memoir on the Papilionidae of the Malayan Region)一文中的图版,载于《林纳学会会报》,第25卷,第一部分,1865年。
(236) 参阅沃莫尔德(Wormald)先生讨论这种蛾的文章,见《昆虫学会会报》,1868年3月2日。
(237) 再参阅有关南美洲的一个属Erateina(尺蠖蛾Geometrae类的一种)的记载,见《昆虫学会会报》,新辑,第5卷,15,16页。
(238) 参阅《伦敦昆虫学会会报》,1868年7月6日,27页。
(239) 哈里斯,《关于新英格兰昆虫类的论文》,弗林特校,1862年,395页。
(240) 例如,我在我儿子的标本箱内见到栎树枯叶蛾(Lasiocampa quercus)、吸饮枯叶蛾(Odonestis potatoria)、Hypogymna dispar、苹红尾毒蛾(Dasychira pudibunda)以及平纹细布蛾(Cycnia mendica)的雄蛾都比雌蛾的颜色更暗淡。后一物种雌雄之间在颜色上差异显著;华莱士先生告诉我说,他相信我们在这里见到一个事例表明保护性的模拟只限于一种性别,这在后面还要更充分地加以说明。平纹细布蛾的白色雌蛾同很普通的具斑灯蛾(Spilosoma menthrasti)相似,而后者雌雄都呈白色;斯坦登先生观察到后面这种蛾子为整窝小火鸡极端厌恶地所拒食,而它们却爱吃其他蛾类;因此若英国鸟类普通把平纹细布蛾误认作樱蛾,它们就可避免被吃掉,这样,其伪装的白色就是高度有利的。
值得注意的是,在设得兰群岛这种蛾的雄者不但同雌者没有很大差别,反而在色彩上同雌者密切相似(参阅麦克拉克伦先生的文章,《昆虫学会会报》,第2卷,1866年,459页)。弗雷泽先生提出(《自然》,1871年4月,489页)当蝙蝠蛾出现在这北方群岛的季节,月色微明,为了使雌蛾易于见到,雄蛾的白色就不是必要的了。
(241) 《一个博物学家在中国诸海漫笔》(Rambles of a Naturalist in the Chinese Seas),1868年,182页。
(242) 《自然》,1871年4月27日,508页。米尔多拉(Meldola)先生引用了唐塞尔(Donzel)关于蝴蝶交配飞行的资料,见《法国昆虫学会》,1837年,77页。再参阅弗雷泽先生关于若干英国蝴蝶的性别差异的文章,见《自然》,1871年4月20日,489页。
(243) 见华莱士关于马来西亚地区凤蝶科的著作,见《林奈学会会报》,第25卷,1865年,8,36页。华莱士先生提出过一个稀有变种的显著事例,这个稀有变种严格介于其他两个特征显著的雌蝶变种之间。再参阅贝茨先生的文章,见《昆虫学会会报》,1866年11月19日,40页。
(244) 热心的贝茨先生曾在昆虫学会上提出这个问题,关于这种效果,我收到过几位昆虫学家的答复。
(245) 贝茨,《亚马孙河流域的博物学家》,第2卷,1863年,228页。再参阅华莱士的文章,见《林奈学会会报》,第25卷,1865年,10页。
(246) 关于这整个问题,参阅《动物和植物在家养下的变异》,1868年,第2卷,23章。
(247) 《动物和植物在家养下的变异》,第2卷,12章,17页。
(248) 《林奈学会会报》,第23卷,1862年,495页。
(249) 《昆虫学会会报》,1866年,12月3日,45页。
(250) 华莱士,《林奈学会会报》,第25卷,1865年,1页;以及《昆虫学会会报》,第4卷(第3辑),1867年,301页。特里门,《林奈学会会报》,第26卷,1869年,497页。赖利,《关于密苏里州害虫类第三次年报》(Third Annual Report on the Noxious Insects of Missouri),1871年,163—168页。后面这篇文章是有价值的,因赖利先生在这里讨论了所有反对贝茨先生理论的意见。
(251) 见《博物学家在尼加拉瓜》(The Naturalist in Nicaragua),1874年,385页。
(252) 《昆虫学会会报》,1866年12月3日,45页;1867年3月4日。
(253) 参阅詹纳·韦尔先生关于昆虫类和食虫鸟类的论文,见《昆虫学会会报》,1869年,21页;以及巴特勒先生的论文,同前会报,27页。赖利先生也举出过类似的事实,见《关于密苏里州害虫类第三次年报》,1871年,148页。然而,华莱士博士和道威尔(M.H.d'Orville)提出过一些相反的事例,见《动物学纪录》,1869年,349页。
(254) 雅列尔(Yarrell),《英国鱼类志》(Hist.of British Fishes),第2卷,1836年,417,425,436页。京特博士告诉我,刺背鱼的刺只限于雌者才有。
(255) 《美国博物学家》(The American Naturalist),1871年4月,119页。
(256) 参阅韦林顿(Warington)先生的有趣文章,见《博物学年刊杂志》,1852年10月和1855年11月。
(257) 诺埃尔·汉弗莱斯(Noel Humphreys),《水上公园》,1857年。
(258) 劳登编辑的《博物学杂志》,第3卷,1830年,331页。
(259) 《田野新闻》(The Field),1867年6月29日。关于肖先生的叙述,参阅《爱丁堡评论》(Edinburgh Review),1843年。另一位有经验的观察家说道[斯克罗普(Scrope)的《在捕鲑鱼的日子里》(Days of Salmon Fishing);60页]雄鲑鱼就像公鹿那样,如它能够做到的话,就会把其他雄鱼全赶走。
(260) 雅列尔,《英国鱼类史》,第2卷,1836年,10页。
(261) 《博物学家在温哥华岛上》(The Naturalist in Vancourer's Island),第1卷,1866年,54页。
(262) ⑺《斯堪的纳维亚探险记》(Scandinavian Adventures),第1卷,1854年,100,104页。
(263) 雅列尔,《英国鱼类志》,第2卷,1836年,416页,关于鳐鱼类的说明,附精美插图,再参阅422,432页。
(264) 为《农夫》(The Farmer)一书所引用,1868年,369页。
(265) 我的这一描述系根据雅列尔的《英国鱼类志》,第1卷,1836年,261,266页。
(266) 《自然》,1873年7月,264页。
(267) 《大英博物馆棘鱼(Acanth)鱼类目录》,1861年,138—151页。
(268) 《瑞典的猎鸟》,1867年,466页等。
(269) 有关这个和以下的物种,我感谢京特博士所提供的材料;另参阅他写的论文,《中美洲的鱼类》,见《动物学会会报》,第6卷,1868年,485页。
(270) 这是京特博士的论述,见《大英博物馆鱼类目录》,第3卷,1861年,141页。
(271) 参阅京特博士关于这个属的文章,《动物学会会报》,1868年,232页。
(272) 巴克兰,见《陆和水》(Land and Water)杂志,1868年7月,377页,附插图。有关雄者的特殊构造还可补充许多其他情况,它们的用处均属不明。
(273) 京特博士,《鱼类目录》,第3卷,221,240页。
(274) 另参阅阿加西斯教授及其夫人合著的《巴西纪游》(A Journey in Brazil),1868年,220页。
(275) 雅列尔,《英国鱼类志》,第2卷,1836年,10,12,35页。
(276) 汤普孙(W.Thompson),《博物学年刊杂志》,第6卷,1841年,440页。
(277) 《美国农学家》(The American Agriculturist),1868年,100页。
(278) 《博物学年刊杂志》,1852年10月。
(279) 见《自然》,1873年5月,25页。
(280) 《驯化协会会报》(Bull.de la Soc.d'Acclimat),巴黎,1869年7月和1870年1月。
(281) 圣懋尚(Saint Vincent),《博物分类学辞典》(Dict.Class,dHist.Nat.),第9卷,1826年,151页。
(282) 由于我在《动物和植物在家养下的变异》一书中有关于这一问题的一些论述,因而迈耶斯(W.F.Mayers)先生[见《关于中国的笔记和质疑》(Chinese Notes and Queries),1868年8月,123页]查阅了中国古代百科全书,他发现金鱼最早是在宋朝于圈养中培育出来的,该朝始于公元960年。到公元1129年这等金鱼已遍及各地。该书另一处宣称,公元1548年以来,“在杭州产生了一个变种,以其浓烈的红色而称之为火鱼。它受到了普遍的赞赏,乃至没有一家不养它,而且以其颜色互相竞赛,并把它作为一种赢利的来源”。
(283) 《威斯敏特评论》,1867年7月,7页。
(284) 麦克莱兰先生的《印度的鲤科》(Indian Cyprinidae)一文,见《亚细亚的研究》(Asiatic Researches),第19卷,第二部分,1839年,230页。
(285) 普歇,《研究》(L'Institut.),Nov.1,1871年,134页。
(286) 《动物学会会报》,1865年,327页,图14和15。
(287) 雅列尔,《英国鱼类志》,第2卷,11页。
(288) 根据格贝(M.Gerbe)的观察;参阅京特的《动物学文献著录》(Record of Zoolog.Literature),1865年,194页。
(289) 居维叶,《动物界》(Règne Animal),第2卷,1829年,242页。
(290) 参阅沃林顿先生对光尾刺鱼习性的最有趣的描述,《博物学年刊杂志》,1855年11月。
(291) 怀曼教授,见《波士顿博物学会会报》,1857年9月15日,特纳教授,见《解剖学和生理学杂志》,1866年11月1日,78页。同样地,京特博士也描述过其他事例。
(292) 雅列尔,《英国鱼类志》,第2卷,1836年,329,338页。
(293) 自从记述这个物种的《桑给巴尔的鱼类》(The Fishes of Zanzibar)一书出版后(普莱费尔上校著,1866年,137页),京特博士又重新检查了这些标本,并给我提供了上述材料。
(294) 《法兰西科学院院报》(Comptes Rendus),1858年,第46卷,353页;1858年,第57卷,916页;1862年,第54卷,393页。荫鱼类(鹰石首鱼Sciaena aquila)所发出的声音,据一些作者说,与其说像鼓声,莫如说更像笛声或风琴声:祖特文博士在这本书的荷文译本(第2卷,36页)中进一步举出了有关鱼类发声的一些细节。
(295) 一 (fathom)等于6英尺。——译者注
(fathom)等于6英尺。——译者注
(296) 金斯利(C.Kingsley)牧师,《自然》,1870年5月,40页。
(297) Triton即Triturus。——译者注
(298) 贝尔,《英国爬行类志》(History of British Reptiles),第2版,1849年,156—159页。
(299) 贝尔,《英国爬行类志》,第2版,1849年,146,151页。
(300) 《“贝格尔”号舰航海中的动物学研究》,1843年,贝尔,同上书,49页。
(301) 《博物学者在尼加拉瓜》(The Naturalist in Nicaragua),1874年,321页。
(302) 有一种锡金蟾蜍(Bufo sikimmensis),只是其雄者的胸部有两块碟状老茧皮,脚趾上也有某些皱纹,这也许和上述的突起物一样有助于达到同一目的(安德森博士,《动物学会会报》,1871年,204页)。
(303) 贝尔,《英国爬行类志》,1849年,93页。
(304) 比肖波,《托德编的解剖学和生理学全书》,第4卷,1503。
(305) 贝尔,同前书,112—114页。
(306) 梅纳德(C.J.Maynard),《美国博物学者》(The American Naturalist),1869年12月,555页。
(307) 参阅我的《“贝格尔”号舰航海调查日记》,1845年,384页。
(308) 京特博士,《英属印度的爬行类》(Reptiles of British India),1864年,7页。
(309) 《卡罗利纳游记》(Travels through Carolina),1791年,128页。
(310) 欧文,《脊椎动物解剖学》,第1卷,1866年,615页。
(311) 安德鲁·史密斯爵士,《南非动物学:爬行纲》,1849年,图10。
(312) 京特博士,《英属印度的爬行类》,雷蒙特协会(RaySoc.),1864年,304,308页。
(313) 斯托里兹卡(Stoliczka)博士,《孟加拉亚细亚协会杂志》,第39卷,1870年,205,211页。
(314) 欧文,《脊椎动物解剖学》,第1卷,1866年,615页。
(315) 见《锡兰漫笔》(Rambles in Ceylon),见《博物学年刊杂志》,第9卷,第2辑,1852年,333页。
(316) 京特博士,《英属印度的爬行类》,1864年,340页。
(317) 《威斯敏特评论》,1867年7月1日,32页。
(318) 安德森博士,《动物学会会报》,1871年,196页。
(319) 《美国博物学者》,1873年,85页。
(320) 奥斯汀先生把这些动物养活了相当长的一段时间,见《陆和水》,1867年7月,9页。
(321) 斯托里兹卡,《孟加拉亚细亚协会杂志》,第35卷,1870年,166页。
(322) 以上有关聋蜥属、赛塔蜥蜴和飞蜥属的全部叙述和引文,以及下述有关角蜥属(Ceratophora)和避役属的事实,都引自京特博士本人的述说或其辉煌的著作《英属印度的爬行类》,雷蒙特协会,1864年,122,130,135页。
(323) 斯温赫先生,《动物学会会报》,1870年,240页。
(324) 布肖尔茨(Bucholz)博士,《普鲁士科学院月刊》(Monatsbericht K.Preuss.Akad.),1874年1月,78页。
(325) 贝尔,《英国爬行类志》,第2版,1849年,40页。
(326) 关于瘦蜥蜴,见《“贝格尔”号舰航海中的动物学研究:爬行类》,贝尔先生著,8页。关于南非蜥蜴类,见《南非动物学:爬行类》,安德鲁·史密斯爵士著,25,39页。关于印度树蜥属,见《英属印度的爬行类》,京特博士著,143页。
(327) 京特,《动物学会会报》,1870年,778页,附彩色插图。
(328) 位于南美阿根廷南部。——译者注。
(329) 《彩鹳》(Ibis),第3卷(新辑),1867年,414页。
(330) 古尔德,《澳大利亚鸟类手册》(Handbook to the Birds of Australia),第2卷,1865年,383页。
(331) 古尔德先生引用,见《蜂鸟科导论》(Introduction to the Trochilidae),1861年,29页。
(332) 古尔德,同上著作,52页。
(333) 汤普孙,《爱尔兰博物志:鸟类》(Nat.Hist.of Ireland:Birds),第2卷,1850年,327页。
(334) 杰尔登,《印度鸟类》(Birds of India),第2卷,1863年,96页。
(335) 麦克吉利夫雷,《英国鸟类志》(Hist.Brit.Birds),第4卷,1852年,177—181页。
(336) 肖姆勃克爵士,《皇家地理学会会报》(Journal of R.Geograph.Soc.),第13卷,1843年,31页。
(337) 《鸟类志》(Ornithological Biography),第l卷,191页。关于鹈鹕类和鹬类,见第3卷,138,477页。
(338) 古尔德,《澳大利亚鸟类手册》,第1卷,395页;第2卷,383页。
(339) 引自休伊特(Hewitt)先生,见《特戈梅尔的家禽手册》(Poultry Book by Tegetmeier),1866年,137页。
(340) 莱亚德,《博物学年刊杂志》,第14卷,1854年,63页。
(341) 杰尔登,《印度鸟类》,第3卷,574页。
(342) 布雷姆,《动物生活图解》,第4卷,1867年,351页。上述有些引自劳埃德,《瑞典的猎鸟》,1867年,79页。
(343) 杰尔登,《印度鸟类》,关于血雉属(Ithaginis),见第3卷,523页;关于鹑鸡属(Galloperdix),见541页。
(344) 关于埃及鹅,参阅麦克吉利夫雷的《英国鸟类志》,第4卷,639页。关于距翅鹅(Plectropterus),见《利文斯通游记》(Livingstone's Travels),254页。关于叫鸟见布雷姆的《动物生活图解》,第4卷,740页。关于这种鸟,再参阅阿扎拉的《南美游记》,第4卷,1809年,179,253页。
(345) 关于英国的凤头麦鸡,参阅卡尔(R.Carr)先生著述,见《陆和水》,1868年8月8日,46页。关于跳凫,见杰尔登的《印度鸟类》,第3卷,647页,以及古尔德的《澳大利亚鸟类手册》,第2卷,220页。关于Holopterus,参阅艾伦先生著述,见《彩鹳》,第5卷,1863年,156页。
(346) 奥杜邦,《鸟类志》,第2卷,492页;第1卷,4—13页。
(347) 布莱思先生,《陆和水》,1867年,212页。
(348) 见理查森(Richardson)关于伞松鸡(Tetrao umbellus)的著述,见《美国边境地区动物志:鸟类》(Fauna Bor.Amer:Birds),1831年,343页。关于雷鸟和雄黑松鸡,见劳埃德的《瑞典猎鸟》,1867年,22,79页。然而,布雷姆认为(《动物生活图解》等,第4卷,352页)德国的雌灰色松鸡一般不光临雄黑色松鸡的巴尔兹舞场,但这是普遍规律的一个例外;可能雌松鸡隐藏于周围灌木丛中,如同斯堪的纳维亚的雌灰色松鸡的情况以及北美洲其他物种的情况那样。
(349) 《鸟类志》,第2卷,275页。
(350) 布雷姆,《动物生活图解》,第4卷,1867年,990页;奥杜邦,《鸟类志》,第2卷,492页。
(351) 《陆和水》,1868年,7月25日,14页。
(352) 奥杜邦,《鸟类志》,关于狂热松鸡,见第2卷,492页;关于椋鸟属(Sturnus),见第2卷,219页。
(353) 《鸟类志》,第5卷,601页。
(354) 戴恩斯·巴林顿,《自然科学学报》,1773年,252页。
(355) 《鸟类学辞典》(Ornithological Dictionary),1833年,475页。
(356) 《笼鸟志》(Naturgeschichte der Stubenv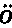 gel),1840年,4页。哈里森·韦尔先生同样写信对我说:“我听说养在同一鸟舍里的最善于鸣唱的雄鸟,一般会首先获得一个配偶。”
gel),1840年,4页。哈里森·韦尔先生同样写信对我说:“我听说养在同一鸟舍里的最善于鸣唱的雄鸟,一般会首先获得一个配偶。”
(357) 英国以往的金币名,合现在21先令。——译者注
(358) 《自然科学学报》,1773年,263页。怀特的《塞尔伯温博物志》(Natural History of Selborne),1825年,第1卷,246页。
(359) 《笼鸟志》,1840年,252页。
(360) 博尔德(Bold),《动物学者》(Zoologist),1843—1844年,659页。
(361) D.巴林顿,《自然科学学报》,1773年,262页;贝奇斯坦(Bechstein),《笼鸟》(Stubenv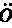 gel),1840年,4页。
gel),1840年,4页。
(362) 河鸟(water-ouzel)的情况也是如此,参阅赫伯恩(Hepburn)先生著述,见《动物学者》,1845—1846年,1068页。
(363) L.劳埃德《瑞典的猎鸟》,1867年,25页。
(364) 巴林顿,同上著作,264页;贝奇斯坦,同上著作,5页。
(365) 马尔(Malle)举出一个奇妙的事例,表明在他的花园内有些野生鸟鸫从一只笼鸟那里自然学会了一曲共和国的歌调。(《自然科学年刊》,第3辑,动物学部分,第10卷,118页)
(366) 毕肖普,《托德的解剖学和生理学全书》,第4卷,1496页。
(367) 如巴林顿在《自然科学学报》(1773年,262页)上所记述的。
(368) corroboree系澳洲土著庆祝胜利舞蹈晚会。——译者注
(369) 古尔德,《澳大利亚鸟类手册》,第1卷,1865年,308—310页;再参阅伍德先生在《学生》杂志(Student,1870年4月,125页)上的记述。
(370) 参阅古尔德的《蜂鸟科导论》,1861年,22页,有关这一效果的记述。
(371) 《加拿大的猎人和博物学者》(The Sportsman and Naturalist in Canada),罗斯·金(W.Ross King)少校著,1866年,144—146页。伍德先生在《学生》杂志(1870年4月,第116页)上对这种鸟在求偶季节的姿态和习性做过最好的记载。他说道,其耳簇毛或颈羽都竖起来了,因而在头冠上面相遇。参阅他的绘图,图39。
(372) 理查森,《美国边境地区动物志:鸟类》,1831年,359页。奥杜邦,同前著作,第4卷,507页。
(373) 关于这个问题最近发表的文章有:A.牛顿教授,见《彩鹳》(Ibis)1862年,107页;卡伦(Cullen)博士,同前著作,1865年,145页;弗劳尔先生,见《动物学会会报》,1865年,747页;以及默里博士,见《动物学会会报》,1868年,471页。后面这篇论文中有一幅绘图示明,一只雄澳洲鸨充分夸示其膨胀起来的喉囊,并非同一物种的所有雄者都有这种发达的喉囊,却是一个奇特的事实。
(374) 贝茨,《亚马孙河流域的博物学家》,1863年,第2卷,284页;华莱士,《动物学会会报》,1850年,206页。最近发现一个新的伞鸟物种(C.penduliger),它的颈部附器还要大些,见《彩鹳》,第1卷,457页。
(375) 毕肖普,见《托德的解剖学和生理学全书》,第4卷,1499页。
(376) 牛顿教授,《动物学会会报》,1871年,651页。
(377) 琵鹭(Platalea)的气管盘旋成8字形,虽然这种鸟(杰尔登,《印度鸟类》,第3卷,763页)是不会叫的;但布莱思先生告诉我说,气管的这样盘旋并非经常出现,所以它们正趋向发育不全。
(378) R.瓦格纳(Wagner)著,《比较解剖学原理》(Elements of Comp.Anat.),英译本,1845年,111页。上述有关鹅的情况,见雅列尔的《英国鸟类志》,第3卷,第2版,1845年,193页。
(379) 波那帕特(C.L.Bonaparte),在《博物学者丛书:鸟类》(Naturalist library:Birds),第6卷,126页。
(380) 劳埃德,《瑞典猎鸟》,1867年,22,81页。
(381) 詹纳,《自然科学学报》,1824年,20页。
(382) 上述事实有关极乐鸟类的,参阅布雷姆的《动物生活图解》,第3卷,325页。有关松鸡类的参阅理查森的《美国边境地区动物志:鸟类》,343,359页;罗斯·金少校的《加拿大的猎人》,1866年,156页;海蒙德先生,见柯克斯(Cox)教授的《印第安纳的地质调查》(Geol.Survey of Indiana),227页;奥杜邦,《美国鸟类志》,第1卷,216页。关于黑鹇,参阅杰尔登的《印度鸟类》,第3卷,533页。关于织布鸟(weavers),参阅利文斯通的《赞比西探险记》(Expedition to the Zambesi),1865年,425页。关于啄木鸟类,参阅麦克吉利夫雷的《英国鸟类志》,第3卷,1840年,84,88,89,95页。关于戴胜,参阅斯温赫先生的记述,见《动物学会会报》,1863年6月23日和1871年,348页。关于夜鹰,参阅奥杜邦的上述著作,第2卷,255页,以及《美国博物学者》,1873年,672页。英国夜鹰在春天快速飞行时也同样发出一种奇妙的响声。
(383) 参阅梅费斯的有趣论文,见《动物学会会报》,1858年,199页。关于丘鹬的习性,参阅麦克吉利夫雷的《英国鸟类志》,第4卷,371页。关于美国的丘鹬,参阅布莱基斯顿(Blakiston)的记述,见《彩鹳》,第5卷,1863年,131页。
(384) 沙尔文先生,《动物学会会报》,1867年,160页。我非常感谢这位著名的鸟类学家所提供的镰翅冠雉(Chamaepetes)的羽毛草图以及其他资料。
(385) 杰尔登,《印度鸟类》,第3卷,618,621页。
(386) 古尔德,《蜂鸟科导论》,1861年,49页;沙尔文,《动物学会会报》,1867年,160页。
(387) 斯克莱特,《动物学会会报》,1860年,90页;《彩鹳》,第4卷,1862年,175页;还有沙尔文的著述,见《彩鹳》,1860年,37页。
(388) 《埃塞俄比亚的尼罗河支流》(The Nile Tributaries of Abyssinia),1867年,203页。
(389) fairy-ring,蕈类在草地上形成的环状斑纹,从前迷信地认为这是由仙女跳舞而成的。——译者注
(390) 关于尖尾松鸡(Tetrao phasianellus)见理查森的《美国边境地区动物志》,361页,有关进一步的细节,参阅布莱基斯顿船长的记述,见《彩鹳》,1863年,125页。关于兀鹫属(Cathartes)和鹭属(Ardea),参阅奥杜邦的《鸟类志》,第2卷,51页;第3卷,89页。关于白喉雀,参阅麦克吉利夫雷的《英国鸟类志》,第2卷,354页;关于印度鸨,参阅杰尔登的《印度鸟类》,第3卷,618页。
(391) 古尔德,《澳洲鸟类手册》,第1卷,444,449,455页。萨丁造亭鸟的亭子可以在摄政公园(Regent's Park)内的动物学会花园里看到。
(392) 对于这一效果的意见,参阅J.肖(Shaw)先生的《动物的美感》(Feeling of Beauty among animals),见《科学协会会刊》(Athenaeum),1866年11月24日,681页。
(393) 参阅默里博士的文章,见《动物学会会报》,1872年,730页,附彩色图。
(394) 蒙蒂罗(Monteiro)先生,《彩鹳》,第4卷,1862年,339页。
(395) 《陆和水》,1868年,217页。
(396) 《鸟类头骨的骨质突起》,见《荷兰的动物学文献》,第1卷,第2期,1872年。
(397) 马歇尔博士,《论鸟尾》(Uber den Vogelschwanz),见前杂志,第l卷,第2期,1872年。
(398) 贾丁(Jardine)的《博物学家丛书:鸟类》,第14卷,166页。
(399) 斯克莱特,《彩鹳》,第6卷,1864年,114页。利文斯通,《赞比西探险记》,1865年,66页。
(400) 杰尔登,《印度鸟类》,第3卷,620页。
(401) 《动物学会会报》,1873年,429页。
(402) 华莱士,《博物学年刊杂志》,第20卷,1857年,416页,另见他的《马来群岛》,第2卷,1869年,390页。
(403) 参阅我的《动物和植物在家养下的变异》,第1卷,289,293页。
(404) 引自德拉弗雷内(M.de Lafresnaye)的著述,见《博物学年刊杂志》,第13卷,1854年,157页;再参阅华莱士先生写的内容更为丰富的文章,见该刊第20卷,1857年,412页,以及见他的《马来群岛》。
(405) 华莱士,《马来群岛》,第2卷,1869年,405页。
(406) 斯克莱特先生,《知识界观察家》(Intellectual Observer),1867年1月。《沃特顿游记》(Waterton's Wanderings),118页。再参阅沙尔文先生的有趣论文,附图,见《彩鹳》,1865年,90页。
(407) 《陆和水》,1867年,394页。
(408) 埃利奥特先生,《动物学会会报》,1869年,589页。
(409) 尼奇(Nitzsch)的《羽区学》(Pterylography),斯克莱特校订,雷伊协会(Ray Soc.),1867年,14页。
(410) 雷鸟的有斑驳的褐色夏羽,作为一种保护色,对它来说其重要性不亚于白色冬羽;因为在斯堪的纳维亚的春季期间,当积雪融化时,这种鸟据了解在获得其夏羽之前受猛禽之害极甚。参阅威廉·冯·符里特(Wilhelm von Wright)的记述,见劳埃德的《瑞典的猎鸟》,1867年,125页。
(411) 上述换羽情况,有关鹬类的,参阅麦克吉利夫雷的《英国鸟类志》,第4卷,371页;有关燕鸻类、麻鹬类和鸨类的,参阅杰尔登的《印度鸟类》,第3卷,615,630,683页;有关鹬属(Totanus)的,参阅前书,700页;有关苍鹭羽饰的,同前书,738页,以及麦克吉利夫雷的前书,第4卷,435,444页,还有斯塔福德艾伦先生的著述,见《彩鹤》,第5卷,1863年,33页。
(412) 关于雷鸟的换羽,参阅古尔德的《英国的鸟类》。关于花蜜鸟,参阅杰尔登的《印度鸟类》,第1卷,359,365,369页。关于鹨类的换羽,参阅布赖茨著述,《彩鹳》,1867年,32页。
(413) 上述有关部分换羽和成年雄鸟保持其婚羽的情况,参阅杰尔登论述鸨类和鹬类的文章,见《印度鸟类》,第3卷,617,637,709,711页。还有布赖茨的文章,见《陆和水》,1867年,84页。关于极乐鸟的换羽,参阅马歇尔博士的有趣论文,见《荷兰文献》(Archives Neerlandaises),第6卷,1871年。关于黑羽长尾鸟,参阅《彩鹳》,第3卷,1861年,133页。关于庄哥-伯劳鸟,参阅杰尔登的前书,第1卷,435页。关于苍鹭(Herodias bubulcus)的春季换羽,参阅艾伦先生的著述,见《彩鹳》,1863年,33页。关于原鸡,参阅布赖茨的著述,见《博物学年刊杂志》,第1卷,1848年,455页;关于这个问题还可参阅我的《动物和植物在家养下的变异》,第1卷,236页。
(414) 麦克吉利夫雷,《英国鸟类志》,第5卷,34,70,223页。关于鸭科的换羽材料,引自沃特顿和蒙塔古。另参阅雅列尔著述,《英国鸟类志》,第3卷,243页。
(415) 关于鹈鹕,参阅斯克莱特文章,见《动物学会会报》,1868年,265页。关于美国的燕雀类,见奥杜邦的《鸟类志》,第1卷,174,221页,以及杰尔登的《印度鸟类》,第2卷,383页。关于马德拉的燕雀(Fringilla cannabina),参阅弗农·哈考特(Vernon Harcourt)先生的著述,见《彩鹳》,第5卷,1863年,230页。
(416) 再参阅《有装饰的家禽》(Ornamental Poultry),狄克逊(E.S.Dixon)牧师著,1848年,8页。
(417) 《印度鸟类》,第1卷,绪论,第24页;关于孔雀,第3卷,507页。参阅古尔德的《蜂鸟科导论》,1861年,15,111页。
(418) 《皇家地理学会会报》,第10卷,1840年,236页。
(419) 《博物学年刊杂志》,第8卷,1854年,157页;再参阅华莱士的前书,第20卷,1857年,412页,以及《马来群岛》,第2卷,1869年,252页,还有贝内特(Bennett)博士的著述,布雷姆在《动物生活图解》中予以引用,第3卷,326页。
(420) 伍德先生就这种夸示方法做过充分记载(《学生》,1870年4月,115页),一是关于金雉的,一是关于日本雉(Ph.versicolor)的,他称此为侧面的或单面的夸示。
(421) 《法则的支配》(The Reign of Law),1867年,203页。
(422) 有关这些鸟类的描述,参阅古尔德的《澳大利亚鸟类手册》,第1卷,1865年,417页。
(423) 《印度鸟类》,第2卷,96页。
(424) 关于非洲夜鹰(Cosmetornis)参阅利文斯通的《赞比西探险记》,1865年,66页。关于锦雉,参阅贾丁的《博物学丛书:鸟类》,第14卷,167页。关于极乐鸟类,参阅莱生的著述,布雷姆引用,见《动物生活图解》,第3卷,325页。关于黑羽长尾鸟,见巴罗(Barrow)的《非洲游记》(Travels in Africa),第1卷,243页,以及《彩鹳》,第3卷,1861年,133页。古尔德先生关于雄鸟胆怯的论述,见《澳大利亚鸟类手册》,第1卷,1865年,210,457页。
(425) 特格梅尔,《家禽之书》,1866年,139页。
(426) 诺曼(Nordman)描述[《莫斯科自然科学皇家学会公报》(Bull.Soc.Imp.des Nat.Moscou),1861,tom xxxiv.p.264]阿穆尔细嘴松鸡(Tetrao urogalloides)的巴尔兹舞会。他估计集合在舞场中的鸟约在一百只以上,不包括卧藏于周围灌木中的雌鸟。它们发出的喧嚣声和松鸡(urogallus)的有所不同。
(427) 关于上述松鸡的集会,见布雷姆的《动物生活》(Thierleben),第4卷,350页;再参阅劳埃德的《瑞典猎鸟》,1867年,19,78页。理查森,《美国边境地区动物志》,鸟类,362页。有关其他鸟类集会的参考材料已列举过了。关于极乐鸟类,参阅华莱士的著述,见《博物学年刊杂志》,第20卷,1857年,412页。关于鹬,参阅劳埃德的上述著作,221页。
(428) 伍德先生引用,见《学生》杂志,1870年4月,125页。
(429) 古尔德,《澳大利亚鸟类手册》,第1卷,300,308,448,451页。关于以上提到的雷鸟,参阅劳埃德的上述著作,129页。
(430) 关于喜鹊,参阅詹纳的论述,见《自然科学学报》,1824年,21页。麦克吉利夫雷,《英国鸟类志》,第1卷,570页。汤普孙,《博物学年刊杂志》,第8卷,1842年,494页。
(431) 关于游隼,参阅汤普孙的《爱尔兰博物学,鸟类》,第1卷,1849年,39页。关于枭、麻雀和鹧鸪,参阅怀特的《塞尔伯温博物志》,第1卷,1825年,第1版,139页。关于红尾鸲,参阅劳登主编的《博物学杂志》,第7卷,1834年,245页。布雷姆(《动物生活》,第4卷,991页)也提到了鸟类在同一天交配三次的例子。
(432) 参阅怀特关于在该季节的早期有小群雄鹧鸪存在的著述(《塞尔伯温博物志》,第1卷,1825年,140页),我还听到过有关这一事实的其他例子。参阅詹纳关于某些鸟类生殖器官延缓发育的著述,《自然科学学报》,1824年。关于三鸟同居,詹纳·韦尔先生为我提供了有关椋鸟和鹦鹉的事例,福克斯先生提供了有关鹧鸪的事例,谨此致谢。有关小嘴乌鸦,参阅《田野新闻》,1864年,415页。关于各种雄鸟在特定时期过后的歌唱,见詹尼斯牧师的《博物学观察》,1846年,87页。
(433) 下述事例是由莫里斯(F.O.Morris)牧师根据尊敬的福雷斯特牧师(Rev.O.W.Forester)的权威材料提出的,(《泰晤士报》,1868年8月6日),他说:“猎场看守人今年在此发现了一个鹰巢,内有五只小鹰。他捕杀了其中4只,留下一只剪短了翅膀的小鹰作为媒鸟,用以诱杀老鹰。次日,有两只老鹰给小鹰喂食,都被打死了,看守人以为事情就会至此完结。但第二天他来到那里,发现又有两只慈悲的老鹰怀着收养和救助孤雏的心情到了那里。他又把它们打死了,然后离开鹰巢而去。后来他回去时又发现两只更加慈悲的老鹰来办理同样的慈善事。他用枪打死了其中一只,另一只也被打中,但未能找到。此后就再没有老鹰来干这种无效的事了”。
(434) 下面一段是牛顿教授从亚当先生的《一个博物学家的游记》(Travels of a Naturalist,1870年,278页)中摘录的。他在谈到笼养的鳾(nut-hatches)时说道,日本五十雀通常的食物为浆果紫杉比较容易破裂的果实,有一次我用榛果代替了这种食物,这种鸟由于不能把榛果弄破,就把榛果一粒一粒地放到水盂里去,显然以为早晚能把它泡软。——这是有关这种鸟的智力的一个有趣证据。
(435) 《萨瑟兰郡游记》(A Tour in Sutherlandshire),第1卷,1849年,185页。布勒(Buller)博士说(《新西兰的鸟类》,1872年,56页),一只大型长尾雄鹦鹉被打死了;于是雌鸟“表现焦急和郁郁不乐,拒绝进食,因过度悲伤而死”。
(436) 《新南威尔士流浪记》(Wanderings in New South Wales),第2卷,1834年,62页。
(437) 巴克斯顿议员(C.Buxton,M.R.)著,《鹦鹉的驯化》(Acclimatization of Parrots),见《博物学年刊杂志》,1868年11月,381页。
(438) 《动物学者》,1847—1848年,1602页。
(439) 休伊特关于野鸭的著述,见《园艺杂志》,1863年1月13日,39页。奥杜邦关于野火鸡的著述,见《鸟类志》,第1卷,14页。关于“模拟画眉”,参阅上述著作,第1卷,110页。
(440) 参阅《彩鹳》,第2卷,1860年,344页。
(441) 关于蜂鸟的有装饰的鸟巢,参见古尔德的《蜂鸟科导论》,1861年,19页。关于造亭鸟参阅古尔德的《澳大利亚鸟类手册》,第1卷,1865年,444—461页。拉姆齐,《彩鹳》,1867年,456页。
(442) 《英国鸟类志》,第2卷,92页。
(443) 《动物学者》,1853—1854年,3946页。
(444) 沃特顿,《博物学论文集》(Essays on Nat.Hist.),第2辑,42,117页。在以下的叙述中,关于赤颈凫,参阅劳登主编的《博物学杂志》,第9卷,616页;劳埃德,《斯堪的纳维亚探险记》,第1卷,1854年,452页。狄克逊,《有装饰的家禽》(Ornamental and Domestic Poultry),137页;休伊特,《园艺杂志》,1863年1月13日,40页,贝希斯坦,《笼鸟志》(Stubenv gel),1840年,230页。詹纳·韦尔先生最近就鸭的两个物种向我提供了一个相似的事例。
gel),1840年,230页。詹纳·韦尔先生最近就鸭的两个物种向我提供了一个相似的事例。
(445) 奥杜邦,《鸟类志》,第1卷,191,349页;第2卷,42,275页;第3卷,2页。
(446) 《稀有和获奖的家禽》(Rare and Prize Poultry),1854年,27页。
(447) 《动物和植物在家养下的变异》,第2卷,103页。
(448) 包依塔和考尔比,《鸽类》,1824年,12页。吕卡(Lucas),《自然遗传的特点》(Traité de l'Héréd.Nat.),第2卷,1850年,296页,亲自观察到有关鸽类的几乎同样的事实。
(449) 《鸽的培育》(Die Taubenzucht),1824年,26页。
(450) 《鸟类志》,第1卷,13页。关于同样的效果,参阅布赖恩特(Bryant)博士的意见,见艾伦编著的《佛罗里达的哺乳类和鸟类》(Mammals and Birds of Florida),344页。
(451) 《动物学会会报》,1835年,54页。斯克莱特先生认为黑翼孔雀是一个独特的物种,并命名为Pavo nigripennis;但在我看来这些证据不过表明它只是一个变种而已。
(452) 鲁道菲,《人类学研究》(Beytr ge zur Anthropologie),1812年,184页。
ge zur Anthropologie),1812年,184页。
(453) 《达尔文学说及其在道德和宗教上的位置》(Die Darwin'sche Theorie,und ihre stellung zu Moral und Religion),1869年,59页。
(454) 这段叙述到利思·亚当斯(A.Leith Adams)先生所提供,参阅他的《田野和森林散记》(Field and Forest Rambles),1873年,76页,这同他自己的经验是一致的。
(455) 关于孔雀,参阅赫伦爵士的著述,见《动物学会学报》,1835年,54页,以狄克逊牧师的《有装饰的家禽》,1848年,8页。关于火鸡,见奥杜邦的上述著作,4页;关于雷鸟,参阅劳埃德的《瑞典的猎鸟》,1867年,23页。
(456) 休伊特先生的论述,在《特格梅尔的家禽之书》(1866年,165页)中引用。
(457) 在劳埃德的《瑞典的猎鸟》(345页)中引用。
(458) 按照布拉西乌斯(Blasius)博士的材料(《彩鹳》,第2卷,1860年,297页),在欧洲繁殖的有425个真实的物种,此外还有60个类型常常被看成为独特的物种。关于后者,布拉西乌斯认为只有10个类型确有疑问,而其余50个类型则应归入亲缘关系最近的物种;不过这表明了我们欧洲的某些鸟类必定有相当大的变异量。博物学家们对于一点还肯定不下来,即某些北美鸟类是否同其相应的欧洲物种有区别而列为不同的物种。还有许多北美类型不久以前被视为不同的物种,现在则认为不过是地方族而已。
(459) 《佛罗里达东部的哺乳类和鸟类》,以及《堪萨斯鸟类考察》等。尽管气候对鸟类的色彩有影响,但仍难说明栖息于某些地方的几乎所有物种何以是暗色的或黑色的,例如赤道下的加拉帕戈斯群岛,巴塔戈尼亚(Patagonia)温暖的广阔平原,以及像埃及那样的地方(参阅哈茨霍恩先生的论述,见《美国博物学家》,1873年,747页)。这些地方是开阔的,为鸟类提供的庇荫处很少;但具有明亮色彩的物种的缺如是否能用保护原理加以解释,似乎尚属可疑。因为彭巴斯草原虽为绿草所覆盖,却是同等开阔的,而鸟类也面临着同等的危险,然而许多具有鲜艳和显著色彩的物种在彼此却是常见的。我有时猜想,上述地方的景色普通都是暗淡的,这是不是会影响那里的鸟类欣赏鲜明色彩。
(460) 《物种起源》,第五版,1869年,104页。我一向见到构造上罕有的和极显著的偏差,值得称为畸形的,很少能够通过自然选择被保存下来,甚至高度有利的变异的保存在某种程度上也决定于机会。我还充分理解单纯的个体差异的重要性,这引导我如此强烈地主张人类无意识选择的重要性,其结果是各个品种的最有价值的个体得到保存,而不需要人类事先有任何改变这个品种性状的意图。但一直到我读了《北英评论》上所刊载的一篇有水平的文章以前(1867年3月,289页及以后各页),我没有看出单独个体所发生的变异,无论是轻微的或强烈显著的,被保存下来的机会是何等之多;这篇文章对我极为有用。
(461) 《蜂鸟科导论》,102页。
(462) 古尔德,《澳大利亚鸟类手册》,第2卷,32,68页。
(463) Tanagra来自南美图皮印第安语,意为颜色鲜明的鸟,这种鸟是中南美产的一个灰雀类。——译者注
(464) 奥杜邦,《鸟类志》,第4卷,1838年,389页。
(465) 杰尔登,《印度鸟类》,第1卷,108页;以及布莱思先生的论述,见《陆和水》,1868年,381页。
(466) 格拉伯,《法鲁旅游日记》(Tagebuch Reise nach F ro),1830年,51—54页。麦克吉利夫雷,《英国鸟类志》,第3卷,745页。《彩鹳》,第5卷,1863年,469页。
ro),1830年,51—54页。麦克吉利夫雷,《英国鸟类志》,第3卷,745页。《彩鹳》,第5卷,1863年,469页。
(467) 格拉伯,同上著作,54页。麦克吉利夫雷,同上著作,第5卷,327页。
(468) 《动物和植物在家养下的变异》,第2卷,92页。
(469) 关于这几点,再参阅《动物和植物在家养下的变异》,第1卷,253页;第2卷,73,75页。
(470) 譬如说,关于管足鸟(Podica)和堇鸡(Gallicrex)的虹彩,参阅《彩鹳》,第2卷,1860年,206页;第5卷,1863年,426页。
(471) 再参阅杰尔登的《印度鸟类》,第1卷,243—245页。
(472) 《贝格尔号舰航海中的动物学》,1841年,6页。
(473) 法国霍丹地方育成的品种。——译者注
(474) 法国品种,黑色,具羽冠。——译者注
(475) 贝希斯坦,《德国博物志》(Naturgeschichte Deutschlands),第4卷,1795年,31页,关于“僧侣鸽”的一个亚变种之论述。
(476) 这幅木刻系由特里门先生根据一幅美丽的绘图为我制成的,谨此致谢;再参阅他对这种蝴蝶翅膀的色彩和形状变异的非常变异量的描述,见其所著《非洲和澳洲的蝶类》(Rhopalocera Africae Australis),186页。
(477) 杰尔登,《印度鸟类》,第3卷,517页。
(478) 《动物和植物在家养下的变异》,第1卷,254页。
(479) 《田野新闻》,1870年5月28日。
(480) 《科学问题通俗讲演集》(Popular lectures on Scientific Subjects),英译本,1873年,219,227,269,390页。
(481) 《法则的支配》,1867年,247页。
(482) 《博物学者在尼加拉瓜》,1874年,112页。
(483) 《蜂鸟科导论》,1861年,110页。
(484) 《物种起源》,第4版,1866年,241页。
(485) 《威斯敏斯特评论》,1867年7月。《旅游杂志》(Journal of Travel),第1卷,1868年,73页。
(486) 特米克(Temminck)说,雌铜色雉(phasianus soemmerringii)的尾羽只有六英寸长,参阅《彩色版画》(Planches coloriées),第5卷,1838年,487和488页;以上面列举的数据是斯克莱特先生为我测得的。关于普通雉,参阅麦克吉利夫雷的《英国鸟类志》,第1卷,118—121页。
(487) 沙普伊(Chapius)博士,《比利时信鸽》,1865年,87页。
(488) 《田野新闻》,1872年9月。
(489) 贝希斯坦,《德国博物志》,1793年,第3卷,339页。
(490) 然而,戴恩斯·巴林顿认为会鸣唱的雌鸟只有少数,可能是因为这种本事在其孵卵期间会招来危险(《自然科学学报》,1773年,164页)。他进一步说,用类似的观点可能说明雌鸟的羽饰何以劣于雄鸟的羽饰。
(491) 拉姆齐先生,《动物学会会报》,1868年,50页。
(492) 默里编,《旅游杂志》,第1卷,1868年,78页。
(493) 同上书,281页。
(494) 奥杜邦,《鸟类志》,第1卷,233页。
(495) 杰尔登,《印度鸟类》,第2卷,108页。古尔德的《澳大利亚鸟类手册》,第1卷,463页。
(496) 例如:燕尾蜂鸟(Eupetomena macroura)的雌者,其头部和尾部呈暗蓝色,腰部为红色;火炬蜂鸟(Lampornis porphyrurus)雌者的上表面为黑绿色,其眼和喉部两侧呈艳红色;Eulampis jugularis雌者的头顶和背部呈绿色,但腰部和尾部为深红色。关于具有高度显著色彩的雌蜂鸟,还可以举出许多其他例子。请参阅古尔德先生有关这一科的巨著。
(497) 沙尔文先生在危地马拉注意到(《彩鹳》,1864年,375页),当太阳强烈照射时,蜂鸟类在很炎热天气里比在凉爽、阴天或下雨的时候更不愿意离开其鸟巢;它们的卵在炎热天气里好像更容易受到损害。
(498) 关于色彩暗淡的鸟类营造荫蔽的鸟巢,我可以列举八个澳洲属的物种作为例子,在古尔德的《澳大利亚鸟类手册》,第1卷,340,362,365,383,387,389,391,414等页均有对它们的描述。
(499) 霍恩先生,《动物学会会报》,1869年,243页。
(500) Superb Warblers系鸟名。——译者注
(501) 有关后面这些物种的造巢及其色彩,参阅古尔德的《澳大利亚鸟类手册》,第1卷,504,527页。
(502) 关于这个问题,我曾查阅过麦克吉利夫雷的《英国的鸟类》,虽然在某些场合中有关鸟巢荫蔽的程度,和雌者色彩的显著程度等问题还存在着一些疑问,可是把卵产于穴中或圆顶鸟巢内的下述鸟类,根据上述标准几乎不能认为它们是色彩显著者:麻雀,有两个物种;椋鸟(Sturnus)其雌者不及雄者鲜艳远甚;河鸟属(Cinclus),脊鸽(Motallica boarula(?));鸲属(Erithacus(?));灌木鹬属(Fruticola)有两个物种,石雕属(Saxicola);红尾鹟属(Ruticilla),有两个物种;莺属(Sylvia),有三个物种,山雀属(Parus),有三个物种,长尾鸟属(Mecistura),Anorthura,旋木雀属(Certhia),鳾属(Sitta),蚁鴷属(Yunx);鹟属(Muscicapa)有两个物种,燕属(Hirundo)有三个物种,以及雨燕(Cypselus)。下面12种鸟类之雌者按同一个标准衡量可视为色彩显著者,即,粉红椋鸟(Pastor),白鹡鸰(Motacilla alba),大山雀(Parus major)和青山雀(P.caeruleus),戴胜属(Upupa),啄木鸟属(Picus)四个物种,佛法僧属(Coracias),翠鸟属(Alcedo)和蜂虎属(Merops)。
(503) 默里编,《旅游杂志》,第1卷,78页。
(504) 参阅《鸟类志》中的许多论述,再参阅尤哥蒙·比托尼(Eugenio Bettoni)对意大利的鸟巢所作的一些奇妙考察,见《意大利科学协会会报》(Atti della Società Italiana),第6卷,1869年,487页。
(505) 见其《咬鹃科(Trogonidae)专论》一书,第1版。
(506) 即深蓝翠鸟(Cyanalcyon)。见古尔德的《澳大利亚鸟类手册》,第1卷,133,130,136页
(507) 雌雄之间各个级进差异可以在澳洲鹦鹉中追查出来。见古尔德的《澳大利亚鸟类手册》,第2卷,14—102页。
(508) 麦克吉利夫雷的《英国的鸟类》,第2卷,433页。杰尔登,《印度鸟类》,第2卷,282页。
(509) 所有以下事例全引自马勒布的巨著《啄木鸟类专论》(Monographie des Picidées)1861年。
(510) 奥杜邦的《鸟类志》,第2卷,75页;再参阅《彩鹳》,第1卷,268页。
(511) 古尔德的《澳大利亚鸟类手册》,第2卷,109—149页。
(512) 关于这种效果,参阅我的《动物和植物在家养下的变异》,第2卷,第十二章。
(513) 《彩鹳》,第6卷,1864年,122页。
(514) 雄鸟在追求雌鸟时,这些装饰都在颤动,并展翅以“卖弄其巨大优越性”:见利思·亚当斯的论述,《田野和森林散记》,1873年,153页。
(515) 关于鳽属(Ardetta),见居维叶的《动物界》(159页脚注),布莱思译。关于游隼,参阅布莱思先生的著述,见查尔斯沃思编的《博物学杂志》,第1卷,1837年,304页。关于卷尾属,见《禾鹳》,1863年,44页。关于琵鹭,见《禾鹳》,第6卷,1864年,366页,关于太平鸟属(Bombycilla),见奥杜邦的《鸟类志》,第1卷,229页。关于长尾鹦哥属(Palaeornis),再参阅杰尔登的《印度鸟类》,第1卷,263页。关于野火鸡,见奥杜邦的《鸟类志》,第1卷,15页;但我听卡顿(J.Catoń)说,在伊利诺伊,雌野火鸡获得一簇硬毛者很少。关于岩栖鸣禽类(Petrocossyphus)的雌者,夏普先生在《动物学会会报》(1872年,496页)举出过类似事例。
(516) 有关后面这些情况,布莱思先生曾就伯劳属(Lanius)、鹟属(Ruticilla)、朱顶雀属(Linaria)和鸭属(Anas)记载下许多例子,居维叶的《动物界》,英译本,158页。关于Pyranga aestiva,奥杜邦也记载过一个相似的事例(《鸟类志》,第5卷,519页)。
(517) 古尔德,《英国鸟类志》。
(518) 关于鸫、伯劳和啄木鸟,参阅布莱思的论述,查尔斯沃思主编的《博物学杂志》,第1卷,1837年,304页;以及他译的居维叶的《动物界》一书,159页脚注。我举出的交喙鸟属的例子系根据布莱思先生的材料。关于鸫,再参阅奥杜邦的《鸟类志》,第2卷,195页。关于金色杜鹃和印度鸽,参阅布莱思的论述,在杰尔登《印度鸟类》,第2卷,485页引用。关于瘤鸭(Sarkidiornis),参阅布莱思的论述,《彩鹳》,1867年,175页。
(519) 请参阅例如古尔德对Cyanalcyon(蓝翠鸟的一种)的说明(《澳大利亚鸟类手册》,第1卷,133页),其幼年雄鸟虽与成年雌鸟相似,但色彩鲜艳较差。在鴗属的一些物种中,其雄鸟具有蓝色尾羽,而雌鸟的尾羽却呈褐色;夏普先生告诉我说,有一种鴗(D.gaudichaudi),其幼年雄鸟的尾羽最初呈褐色。古尔德先生曾描述过某些黑色美冠鹦鹉和大型长尾鹦鹉的雌雄二者及其幼鸟,它们也受同一规律所支配。还有杰尔登关于红色鹦哥(Palaeornis rosay)的论述(《印度鸟类》,第1卷,260页),它的幼鸟同雌鸟的相似胜于同雄鸟的相似。参阅奥杜邦关于雀形鸽(Columba passerina)雌雄二者及其幼鸟的论述(《鸟类志》,第2卷,475页)。
(520) 关于这个材料,我感谢古尔德先生,他给我看过这些标本,再参阅他的《蜂鸟科导论》,1861年,120页。
(521) 麦克吉利夫雷,《英国鸟类志》,第5卷,207—214页。
(522) 参阅他的可钦佩的论文,见《孟加拉亚细亚协会杂志》,第19卷,1850年,223页;再参阅杰尔登的《印度鸟类》,第1卷,导言,第29页。关于翠鸟类,施勒格尔(Schlegel)教授告诉布莱思先生说,他根据对成年雄鸟的比较,就能区别若干不同的族。
(523) Kalij pheasant,为Kallege的一个变种,同银雉的亲缘关系相近。——译者注
(524) 再参阅斯温赫先生的论述,《彩鹳》,1863年7月,131页,以前还有一篇论文,附载布赖茨先生的笔记摘要,见《彩鹳》,1861年1月,25页。
(525) 华莱士,《马来群岛》,第2卷,1869年,394页。
(526) 波伦(M.F.Pollen)曾描述过这些物种,见《彩鹳》,1866年,275页,附彩色插图。
(527) 参阅《动物和植物在家养下的变异》,第1卷,251页。
(528) 麦克吉利夫雷,《英国鸟类志》,第1卷,172—174页。
(529) 关于这个问题,参阅《动物和植物在家养下的变异》,第23章。
(530) 奥杜邦,《鸟类志》,第1卷,193页。麦克吉利夫雷,《英国鸟类志》,第3卷,85页。再参阅前此所举的有关印度啄木鸟的事例。
(531) 《威斯敏特评论》,1867年7月。默里《游记》,1868年,83页。
(532) 关于澳大利亚的物种,参阅古尔德的《澳大利亚鸟类手册》,第2卷,178,186,188页。在不列颠博物馆可以看到澳大利亚的漂鸟(Pedionomus tonquatus)标本也表现了相似的性差异。
(533) 杰尔登,《印度鸟类》,第3卷,596页。斯温赫先生的论述,见《彩鹳》,1865年,542页;1866年,131,405页。
(534) 杰尔登,《印度鸟类》,第3卷,677页。
(535) 古尔德的《澳大利亚鸟类手册》,第2卷,275页。
(536) 《印度的田野》(The Indian Field),1858年9月,3页。
(537) 《彩鹳》,1866年,298页。
(538) 关于这若干记述,参阅古尔德先生《大不列颠鸟类》,牛顿教授告诉我说,他根据自己和其他人的观察,长期一直相信上述物种的雄鸟全部地或者大部地担负起孵卵的义务,而且“在危险中它们对幼鸟的献身精神远比雌鸟大得多”。正如他告诉我的,斑尾塍鹬(Limosa lapponica)和少数一些涉禽类的雌鸟都比雄鸟大,而且具有更强烈对比的色彩。
(539) 塞兰岛土人断言(华莱士,《马来群岛》,第2卷,150页),其雄鸟和雌鸟轮流孵卵;但巴特利特先生认为这大概是雌鸟来巢下卵之误。
(540) 《学生》,1870年4月,124页。
(541) 关于这种鸟在圈养条件下的习性,参阅贝内特先生的杰出文章,见《陆和水》,1868年5月,233页。
(542) 斯克莱特先生关于鸵鸟孵卵问题的论述,见《动物学会学报》,1863年6月9日游鸵(Rhea darwinii)的情况也是如此;马斯特斯(Musters)船长说(《和巴塔哥尼亚的印第安人相处的日子》,1871年,128页),其雄鸟比雌鸟更大、更强壮而且动作更快,其色彩稍暗淡;但它担负孵卵和照看幼鸟的全部责任,正如游鸵属普通物种的雄鸟那样。
(543) 关于食腐肉的鸢,见《贝格尔舰航海中的动物学:鸟类》,1841年,16页。关于旋木鸟和欧夜鹰(Eurostopodus),见古尔德的《澳大利亚鸟类手册》,第1卷,602,97页。新西兰的麻鸭(Tadorna variegata)提供了一个完全相似的情况;其雌鸟的头部呈纯白色,而背部则比雄鸟的颜色更红,雄鸟的头部为一种鲜艳的暗青铜色,其背部覆以美丽的细纹鼠色羽衣,因此这两方面加起来它就会被认为是两性中更漂亮者。它比雌鸟大,更好斗,而且不孵卵,因而从所有这些方面看,这个物种应归入我们第一类;但斯克莱特先生(《动物学会学报》,1866年,150页)非常惊奇地看到幼年雌雄二者在孵出后三个月左右的时候,它们暗色的头部和颈部同成年雄鸟相似,而不同成年雌鸟相似;因此在这种场合中似乎是雌鸟发生了改变,而雄鸟和幼鸟则保持其羽衣的以往状态。
(544) 杰尔登,《印度鸟类》,第3卷,598页。
(545) 杰尔登,《印度鸟类》,第1卷,222,228页。古尔德,《澳大利亚鸟类手册》,第1卷,124,130页。
(546) 古尔德,同上著作,第2卷,37,46,56页。
(547) 奥杜邦,《鸟类志》,第2卷,55页。
(548) 《动物和植物在家养下的变异》,第2卷,79页。
(549) 查里沃思主编的《博物学杂志》,第1卷,1837年,305,306页。
(550) 《科普协会简报》(Bulletin de La Soc.Vaudoise des Sc.Nat.),第10卷,1869年,132页,波兰天鹅,即雅列尔命名的变天鹅(Cygnus immutabilis),其幼鸟永呈白色;但是,正如斯克莱特先生告诉我的,据认为,这个物种不过是普通天鹅(Cygnus olor)的一个变种而已。
(551) 我感谢布莱思先生提供有关这个属的材料。巴勒斯坦麻雀乃是一个石雀(Petronia)的亚属。
(552) 例如,夏鷽(Tanagra aestiva)和蓝燕雀(Fringilla cyanea)的雄鸟的漂亮羽衣完全长好,需时3年,而燕雀(Fringilla crris)的雄鸟需时4年(见奥杜邦的《鸟类志》,第1卷,233,280,378页),斑凫(Harlequin duck)需时三年(同上著作,第3卷,614页)。我听詹纳·韦尔先生说,雄金雉在孵出三个月左右就可同雌金雉区别开来,但其华丽的完善羽衣要到来年九月底才会出现。
(553) 这样,坦塔罗斯彩鹳(Ibis tantalus)和美洲鹤(Grus americanus)需时四年,红鹳(Flamingo)需时数年,而游鹭(Andea ludovicana)需时两年,才获得其完善的羽衣。参阅奥杜邦的上述著作,第1卷,221页;第3卷,133,139,211页。
(554) 布莱思先生的论述,见查理沃思主编的《博物学杂志》,第1卷,1837年,300页。有关金雉的材料是巴特利特先生告诉我的。
(555) 我曾注意到奥托邦的《鸟类志》中的下述事例。美国的红尾鹟(Muscapica ruticilla,第1卷,203页),坦塔罗斯彩鹳需时四年才达到完全成熟,但有时在孵出后第二年就繁育(第3卷,133页)。美洲鹤需要同样长的时间,但在获得其完善羽衣前就繁育(第3卷,211页)。青鹭(Ardea caerulea)的成鸟呈蓝色,幼鸟则呈白色;可以看到白色的、杂色的以及成熟的蓝色鸟全都在一块繁育(第4卷,58页):但布莱思先生告诉我说,某些苍鹭显然是二态的,因为可以看到同龄的白色和有色的个体。斑凫(Anas histrionica Linn.)需时三年才能获得其完善羽衣,虽然许多凫类在孵出后第二年就繁育(第3卷,614页)。白头隼(Falco leucocephalus,第3卷,210页)据知同样在未成熟时就繁育。黄鹂属的一些物种(根据布莱思先生和斯温赫先生的材料,见《彩鹤》,1863年7月,68页)同样在获得其完善羽衣前就繁育。
(556) 见上注。
(557) 属于完全不同纲的其他动物,无论是惯常地还是偶然地都能在完全获得其成年性状之前就可以繁育。鲑鱼的幼年雄者就是如此。有几种两栖动物,据知尚保持其幼体构造时就进行繁育。弗里茨·米勒指出(《支持达尔文的事实和论证》,英译本,1869年,79页)有几种异脚类甲壳动物的雄者在幼小时即达到性成熟了;我推论这是成熟前就进行繁育的一个例子,因为那时它们还没有获得其完全发育的抱握器。所有这类事实都是非常有趣的好像凭一种手段就可使物种实现性状的巨大改变。
(558) 杰尔登,《印度鸟类》,第3卷,507页,关于孔雀。马歇尔博士认为极乐鸟的较老和较鲜艳的雄鸟比较年幼的雄鸟有更大优越性;见《荷兰文献》,第6卷,1871年。——关于鹭属,见奥杜邦的上述著作,第3卷,139页。
(559) 关于图例见麦克吉利夫雷的《英国鸟类志》一书,关于鹬属见第229,271页;关于流苏鹬,见第172页;关于剑鸻(Charadrius hiaticula),见118页;关于雨鸻(Charadrius pluvialis),见第94页。
(560) 有关北美洲金翅雀、暗色燕雀见奥杜邦的《鸟类志》第1卷,172页。关于莫鲁里鸟,见古尔德的《澳大利亚鸟类手册》,第1卷,318页。
(561) 我感谢布莱思先生提供的有关印度白鹭的材料;再参阅杰尔登的《印度鸟类》,第3卷,749页。关于懒钳嘴鸭,见布莱思的论述,《彩鹳》,1867年,173页。
(562) 关于海雀,见麦克吉利夫雷的《英国鸟类志》,第5卷,347页。关于白冠燕雀见奥杜邦的上述著作,第2卷,89页。我以后还会提到某些苍鹭和白鹭的幼鸟呈白色的问题。
(563) 参阅《英国鸟类志》,第1卷,1839年,159页。
(564) 奥杜邦,《鸟类志》,第1卷,113页。
(565) 赖特(C.A.Wright)先生的著述,见《彩鹳》,第6卷,1864年,65页。杰尔登,《印度鸟类》,第1卷,515页。再参阅布莱思关于乌鸫的论述,见查理沃思主编的《博物学杂志》,第6卷,1837年,113页。
(566) 尚有数例补充如下:红鷽(Tanagra rubra)的年幼雄鸟同年幼雌鸟有区别(奥杜邦,《鸟类志》,第4卷,392页),一种印度的蓝鳾(Dendrophila frontalis),其雏鸟也同样地雌雄有别(杰尔登,《印度鸟类》,第1卷,389页)。布莱思先生也告诉我说,黑喉石雕(Saxicola rubicola)在很早的年龄其雌雄二者就可彼此区别。沙尔文先生举出(《动物学会会报》,1870年,206页)一个例子表明一种蜂鸟同下面所说的Eustephanus相似。
(567) 见《威斯敏特评论》,1867年7月,第5页。
(568) 《彩鹳》,1859年,第1卷,429页,及以下诸页。然而,罗尔夫斯(Rohlfs)博士在一封信中对我说,根据他在撒哈拉沙漠的经验,这个说法未免过火。
(569) 关于巨嘴鸟的巨嘴迄今尚无满意的解释,至于其鲜艳的色彩,有关解释还要少。贝茨先生说(《亚马孙河流域的博物学家》,第2卷,1863年,341页),它们是用嘴来啄取树枝末端的果实;同样地,像其他作者说过的,它们还用巨嘴去攫取其他鸟类巢里的卵和幼鸟。但是,正如贝茨先生所承认的,这种鸟嘴“对于用它所要达到的目的来说简直不能认为是形状很完善的工具”。如果说这种鸟嘴仅仅用做把握器官,则其宽度、深度、以及长度所示明的那样巨大体积乃是不可理解的。贝尔特先生相信这等鸟喙的基本用途在于抵御敌害,特别是在树洞里孵卵的雌鸟尤其需用它(《博物学者在尼加拉瓜》,197页)。
(570) 即龙首鵎鵼(Ramphastos Carinatus),见古尔德的《鵎鵼科专论》(Monograph Ramphastidae)。
(571) 关于鸥属、潜鸟以及燕鸥,见麦克吉利夫雷的《英国鸟类志》,第5卷,515,584,626页。关于雪雁,见奥杜邦的《鸟类志》,第4卷,562页。关于懒钳嘴鸭,见布莱思先生的论述,《彩鹳》,1867年,173页。
(572) 关于兀鹫,可以注意到它们在高空翱翔,既远又广,就像海洋上的水鸟那样,它们有三或四个物种几乎是完全白色或大部白色,许多其他物种则呈黑色。因此这里再一次表明了显著的色彩在生殖季节也许能帮助雌雄二者互相找到对方。
(573) 参阅杰尔登关于长尾鹦鹉属的论述,见《印度鸟类》,第1卷,258—260页。
(574) 美国的青色鹭和红色鹭的幼鸟也是白色的,成鸟的颜色犹如各自特有的名称。奥杜邦(《鸟类志》第3卷,416页;第4卷,58页)在想到羽衣的这种显著变化将大大使“分类学家们为难”时,似乎颇为高兴。
(575) 我非常感激斯克莱特先生为我审阅了有关鸟类的第四章以及后面有关哺乳动物的两章。这样在一些物种的名称上,以及在叙述任何一件为这位著名博物学家所清楚了解的事实上,我得以避免发生谬误。不过关于我从不同作者引用来的叙述的精确性如何,他当然完全没有责任。
(576) 参阅沃特顿关于两只野兔相斗的记载,《动物学者》(Zoologist),第1卷,1843年,211页。关于鼹鼠,贝尔,《英国兽类志》,第1版,100页。关于松鼠,奥杜邦和贝克曼,《北美的胎生四足兽》(Viviparous Quadrupeds of N.America),1846年,269页。关于河狸,格林先生,《林奈动物学会会刊》(Journal of Lin.Soc.Zoolog.),第10卷,1869年,362页。
(577) 关于海豹的战斗,艾博特(C.Abbott),《动物学会会报》(Proc.Zool.Soc.),1868年,191页;布朗先生,同上“会报”,1868年,436页;劳埃德,《瑞典的猎鸟》,1867年,412页;还可参阅彭南特(Pennant)的文章。关于抹香鲸,J.H.汤普孙,《动物学会会报》,1867年,246页。
(578) 参阅斯克罗普(《鹿的狩猎技术》,Art of Deer-stalking,17页)关于两只马鹿(Cervus elaphus)的角纠结在一起的记载。理查森,《美国边境地区动物志》,1829年,252页,他说,马鹿、驼鹿以及驯鹿的角都会这样地纠结在一起。史密斯在好望角发现过两只角马(gnus)的骨骼,它们的角也是如此。
(579) 拉蒙特(Lamont)先生说(《海象的交配季节》(Seasons with the Sea-Horses,1861年,143页),雄海象的良好獠牙重达4磅,比雌海象的獠牙为长,后者重约3磅。据描述雄海象相斗凶猛异常。关于雌海象偶尔缺少獠牙,参阅布朗的文章,《动物学会会报》,1868年,429页。
(580) 欧文,《脊椎动物解剖学》,第3卷,283页。
(581) 布朗先生,《动物学会会报》,1869年,553页。关于这个獠牙的同源性质,参阅特纳教授的论著,《解剖学及生理学杂志》,1872年,76页。关于雄者两只獠牙都发达的情况,参阅克拉克的论著,《动物学会会报》,1871年,42页。
(582) 关于抹香鲸和鸭嘴兽,欧文,同前杂志,第3卷,638,641页。祖特文(Zouteveen)在该书的荷兰文译本(第2卷,292页)中,曾引用哈廷的说法。
(583) 关于驯鹿角的构造及其脱落,霍夫勃格(Hoffberg),《(Amoenitates Acad)》,第4卷,1788年,149页。关于美国的变种或物种,理查森,《美国边境地区动物志》,241页;再参阅罗斯·金(Ross King)少校的《加拿大的狩猎爱好者》(Sportsman in Canada),1866年,80页。
(584) 小圣·伊莱尔,《动物学通论》(Essais de Zoolog.Générale),1841年,513页。除去角之外,其他雄性性状有时也可照样地传给雌者;例如,邦纳(Boner)先生在谈到雌性小羚羊时说道(《巴伐利亚山区中小羚羊的狩猎》,Chamois Hunting in the Mountains of Bavaria,1860年,第2版,363页),老龄雌性小羚羊“不仅其头部同雄者的极相似,而且沉其背部有一行长毛,通常这只在雄者身上才有”。
(585) 关于羌鹿,格雷博士,《大英博物馆哺乳动物目录》(Catalogue of Mammalia in the British Museum),第三部分,220页。关于美洲赤鹿,卡顿,《渥太华自然科学研究院院报》,1868年5月,9页。
(586) 此项材料蒙坎菲尔德(Canfield)博士提供,这篇论文见《动物学会会报》,1866年,105页。
(587) 例如,雌南非羚羊(Ant.euchore)的角同一个不同物种南美羚羊(Ant.dorcas.var.Corine)的角相似,参阅德马雷(Desmarest)的《哺乳动物学》(Mammalogie),455页。
(588) 格雷,《大英博物馆哺乳动物目录》,第三部分,1852年,160页。
(589) 理查森,《美国边境地区动物志》,278页。
(590) 产于爪哇及东印度群岛的野牛,行动迅速,常为小群,马来人有饲养这种牛的。——译者注
(591) 《陆和水》,1867年,346页。
(592) 安德鲁·史密斯,《南非洲动物学》(Zoology of S.Africa),图版第十九。欧文,《脊椎动物解剖学》,第3卷,624页。
(593) 这是赛德利茨(Seidlitz)的结论,《达尔文学说》(Die Darwinsche Theorie),1871年,47页。
(594) 我非常感激维克托·卡勒斯教授,他为我在萨克森做过有关这一问题的调查。冯·纳修西亚斯(H.von Nathusius)说,如对绵羊在其幼年时进行阉割,它们的角或者完全消失,或者仅留一点残迹(《牲畜饲养》Viehzucht,1872年,64页);但我不知道他所谈的是美利奴羊,还是普通品种。
(595) 我曾举出各种试验以及其他证据,证明情况确系如此,见我的著作《动物和植物在家养下的变异》,1868年,39—47页。
(596) 埃默森·坦南特(J.Emerson Tennent)爵士,《锡兰》(Ceylon),第2卷,1859年,274页。关于马六甲,见《印度群岛杂志》(Journal of Indian Archipelago),第4卷,357页。
(597) 《加尔各答博物学杂志》(Calcutta Journal of Nar.Hist.),第2卷,1843年,526页。
(598) 布莱思先生,《陆和水》,1867年3月,134页,系根据赫顿上尉以及其他人士的材料。关于彭布罗克希尔郡山羊,参阅《田野新闻》,1869年,150页。
(599) 贝利,《关于兽角的使用》(Surl'usage des Cornes),见《自然科学年刊》,第2卷,1824年,369页。
(600) 关于马鹿的角,欧文,《英国化石哺乳类动物》(Brifish Fossil Mammals),1949年,478页;关于驯鹿的角,理查森,《美国边境地区动物志》,1829年,24页。关于莫里茨堡的材料,系维克托·卡勒斯教授提供,特此致谢。
(601) 卡顿说,“当在鹿群中的优势问题一旦解决并为全群所接受之后”,美洲鹿就用其前足相斗。贝利,《兽角的使用》,见《自然科学年刊》,第2卷,1824年,371页。
(602) 参阅上述卡顿的论文附录,其中有非常有趣的记载。
(603) 《美国博物学者》,1869年12月,552页。
(604) musk-deer,一名香獐,形似鹿而小。雌雄皆无角,雄之上颚甚发达,有细长犬齿突出口外,长约3寸。——译者注
(605) 帕拉斯,《动物学专论》(Spicilegia Zoologica),第13分册,1779年,18页。
(606) 拉蒙特,《海象的交配季节》,1861年,141页。
(607) 关于短獠牙的莫克那(Mooknah)变种攻击其他象的方法,再参阅科斯(Corse)的文章,见《自然科学学报》,1799年,212页。
(608) 体重约30磅,毛角呈赤黄褐色,角小而不分枝,上颚犬齿发达,叫声如狐。——译者注
(609) 欧文,《脊椎动物解剖学》,第3卷,349页。
(610) 吕佩尔(Rüppell)论鹿和羚羊的犬齿,见《动物学会会报》,1836年1月12日,3页,其中并有马丁先生关于雌美洲鹿的附注。再参阅法克纳的关于一只成年雌鹿的犬齿的报道,见《古生物学的专题研究及记录》(Palaeont、Memoirs and Notes),第1卷,1868年,576页。老龄雄麝的犬齿有时长达三英寸(《动物学专论》,第13分册,1779年,18页),而老龄雌麝的犬齿仅系残迹,高出牙床半英寸。
(611) 埃默森·坦南特,《锡兰》,第2卷,1859年,275页;欧文,《英国化石哺乳类动物》,1846年,245页。
(612) 理查森,《美国边境地区动物志》,关于驼鹿,236,237页;关于驼鹿角的扩张,《陆和水》,1869年,143页。再参阅欧文的《英国化石哺乳类动物》,关于爱尔兰驼鹿,447,455页。
(613) 《森林动物》(Forest Creatures),博纳,1861年,60页。
(614) 或称海熊、腽肭兽。——译者注
(615) 参阅艾伦先生的很有趣的论文,见《剑桥大学有袋类比较动物学学报》(Bull.Mus.Comp.Zoolog.of Cambridge),美国版,第2卷,第1号,82页。其重量系由谨慎的观察家勃兰特上尉所确定。吉尔博士,《美国博物学者》,1871年1月;关于雌雄鲸的相对大小,《美国博物学者》,1873年1月。
(616) 《动物的身体机构》(Animal Economy),45页。
(617) blood-hound,善嗅血腥,可训练其追蹑负伤的猎物,或在战场寻觅伤兵,或破获凶杀案件。——译者注
(618) 再参阅理查森的《关于犬的手册》(Manual on the Dog),50页。麦克尼尔先生曾就苏格兰猎鹿狗提出过非常有价值的报告,他首先提醒人们注意到这种狗雌雄二者大小不相等,见斯克罗普的《鹿的狩猎技术》。卡波勒斯有意发表有关这一著名品种的全部材料及其历史,我翘首以待。
(619) 布雷姆,《动物生活》,第2卷,729—732页。
(620) 参阅华莱士先生关于这种动物的有趣报道,见《马来群岛》,第1卷,1869年,435页。
(621) 《意大利自然科学协会会刊》(Atti della Soc.Italiana di Sc.Nat.),第15卷,第4分册,1873年。
(622) 《时代》(The Times),1857年11月10 日。关于加拿大山猫,参阅奥杜邦和贝克曼的《北美四足兽》(Quadrupeds of North America),1846年,139页。
(623) 鬃海狗亦有译作“海狮”,为海驴(Otaria stelleri Less.,又名steller’s sea lion)的一种,常群集于福克兰岛。斑海豹(seal,phoca vitulina L.),多产于北太平洋。二者均属鳍脚类。
(624) 默里博士,关于海狗,《动物学会会报》,1869年,109页。艾伦先生在上述论文中(75页)曾提到,雄者的颈毛固然长于雌者的颈毛,但是否值得叫做鬃,尚可杯疑。
(625) 博纳先生曾对德国赤鹿做过最好的描述(《森林动物》,1861年,81页),他说,“当雄鹿保卫其权利,反击一只入侵者的时候,另一只入侵者趁机侵入其妻妾禁区,一一驱走为其战利品。完全一样的情况也发生于海豹,参阅艾伦先生的论文,同前杂志,100页。
(626) 艾伦先生,《剑桥大学有袋类比较动物学学报》,美国版,第2卷,第1号,99页。
(627) 梅休著,《狗:它们的管理》(Dogs:their Management),第2版,1864年,187—192页。
(628) 艾力克斯·沃克(Alex Walker),《论血族通婚姻》(On Intermarriage),1838年,276页,244页。
(629) 形小,常用于助猎,以捕获鼬、獾、兔或水獭。——译者注
(630) 《自然遗传专论》(Traitéde I'Héréd.Nat.),第2卷,1850年,296页。
(631) 《动物研究院院报》(Amoenitates Acad.),第4卷,1788年,160页。
(632) 欧文,《脊椎动物解剖学》,第3卷,585页。
(633) 同上书,595页。
(634) 关于驼鹿和野生驯鹿的习性,参阅罗斯·金(Ross King)的著作,见《加拿大的猎人》,1866年,53,131页。
(635) 欧文,《脊椎动物解剖学》,第3卷,600页。
(636) 格林先生,《林奈学会会刊》,第10卷,动物学部分,1869年,362页。
(637) 马丁,《哺乳动物志大纲》(General Introduction to the Nat.Hist.of Mamm.Animals),1841年,431页。
(638) 《巴拉圭哺乳动物志》,1830年,15,31页。
(639) 关于象海豹,参阅莱生的文章,见《博物分类学辞典》,第13卷,418页。关于冠海豹,参阅德凯(Dekay)的文章,见《纽约博物学会年报》(Annals of Lyceum of Nat.Hist.New York),第1卷,1824年,94页。彭南特也曾从海豹猎人那里搜集过有关这种动物的材料。最充分的材料是由布朗先生提出的,见《动物学会会报》,1868年,435页。
(640) 关于河狸,参阅摩尔根先生的很有趣的著作《美国河狸》(AmericanBeaver),1868年,300页。帕拉斯很好地讨论了哺乳动物散发气味的腺体,见《动物学专论》,第8卷,1779年,23页。欧文也曾记载过这种腺体,其中包括象和鼩鼱的腺体,见《脊椎动物解剖学》,第3卷,634,763页。关于蝙蝠,参阅多布森先生的文章,见《动物学会会报》,1873年,241页。
(641) 伦格尔,《巴拉圭哺乳动物志》,1830年,355页。这位观察家还举出一些有关这种气味的奇妙特性。
(642) 欧文,《脊椎动物解剖学》,第3卷,632页。再参阅默里博士对这等腺体的观察材料,见《动物学会会报》,1870年,340页。德马雷关于红斑羚羊(Antilope subgutturosa),见《哺乳动物学》(Mammalogie),1820年,455页。
(643) 帕拉斯,《动物学专论》,第13卷,1799年,24页。德穆兰(Desmoulins),《博物分类学辞典》,第3卷,586页。
(644) 格雷博士,《诺斯雷动物园采访记》(Gleanings frorn the Menagerie at KnowSley),28页。
(645) 卡顿论北美马鹿,《渥太华自然科学院院报》(Transact.Ottawa Acad.Nat.Sciences),1868年,36,40页;布莱思,关于喜马拉雅野山羊,《陆和水》,1867年,37页。
(646) 欧文编,《亨特的论文及其观察材料》(Hunter'sEssays and Observations),第1卷,1861年,236页。
(647) 参阅格雷博士的《大英博物馆哺乳动物目录》,第三部分,1852年,144页。
(648) 伦格尔,《哺乳动物志》,14页;德马雷,《哺乳动物学》,86页。
(649) 参阅我的《动物和植物在家养下的变异》第1卷中所载的这几种动物;还有第2卷,73页;以及第二十章中所载的有关半开化人实行选择的情况。关于伯布拉山羊,参阅格雷的《目录》,157页。
(650) 关于岩大袋鼠(Osphranter rufus),古尔德,《澳大利亚哺乳动物》(Mammals of Australia),1863年,第2卷。关于负鼠,德马雷,《哺乳动物学》,256页。
(651) 为圭亚那的一处地方。——译者注
(652) 《博物学年刊杂志》,1867年11月,325页。关于巢鼠,德马雷,《哺乳动物学》,304页。
(653) 艾伦,《剑桥大学有袋类比较动物学学报》,美国版,1869年,207页。多布森博士关于翼手目性征的文章,见《动物学会会报》,1873年,241页。格雷博士关于树懒的文章,同上杂志,1871年,436页。
(654) 德马雷,《哺乳动物学》,1820年,220页。关于线斑猫(Felis mitis),伦格尔,同前书,194页。
(655) 默里博士关于海狗的文章,见《动物学会会报》,1869年,108页。布朗先生关于格陵兰海豹的文章,同上杂志,1868年,417页。关于海豹的颜色,再参阅德斯玛列司特的文章,同上杂志,243,249页。
(656) 卡顿,《渥太华自然科学院院报》,1868年,4页。
(657) 格雷博士,《大英博物馆哺乳动物目录》,第三部分,1852年,134—142页;再参阅格雷博士的《诺斯雷动物园访问记》,该书载有一张漂亮的德比大角斑羚的图:参阅该书有关羚属的章节。关于好望角大角斑羚(Orea canna),参阅安德鲁·史密斯的《南非动物学》,41,42页。在伦敦动物园中也有许多这等羚羊。
(658) 关于黑印度羚(Ant.niger),参阅《动物学会会报》,1850年,133页。有一个亲缘近似的物种,在体色方面有同等的性差异,参阅贝克爵士的《阿伯特·尼安萨》(Albert Nyanza),第2卷,1866年,627页。关于水草地印度羚,格雷,《大英博物馆哺乳动物目录》,100页。德马雷,《哺乳动物学》,468页,关于卡玛印度羚。安德鲁·史密斯,《南非动物学》,关于角马。
(659) 《渥太华自然科学院院报》,1868年5月21日,3,5页。
(660) S.米勒,关于爪哇牛,《马来群岛动物志》(Zoog.Indischen Archipel.),1839—1844年,35彩图;再参阅拉弗尔斯(Raffles)的文章,布赖茨先生引用,见《陆和水》,1867年,476页。关于山羊,格雷博士,《大英博物馆目录》,146页;德马雷,《哺乳动物学》,482页。关于南美沼地鹿,伦格尔,同前书,345页。
(661) 斯克莱特,《动物学会会报》,1866年,1页。M.M.波伦和范达姆(van Dam)也充分肯定了同一事实。再参阅格雷博士的文章,《博物学年刊杂志》,1871年5月,340页。
(662) 关于吼猴,伦格尔,同前书,14页;布雷姆,《动物生活图解》,第1卷,96,107页。关于悬猴,德马雷,《哺乳动物学》,75页。关于长臂猿,布莱思,《陆和水》,1867年,135页。关于瘦猴(Semnopithecus),S.米勒,《马来群岛动物志》,10彩图。
(663) 热尔韦(Gervais),《哺乳动物志》,1854年,103页,载有雄西非山魈的头骨图。再参阅德马雷的《哺乳动物学》,70页。小圣伊莱尔和居维叶,《哺乳动物志》,第1卷,1824年。
(664) 即Phalangista vulpina,属食果有袋类,形似松鼠,体长约一尺半,尾长一尺许,每产一二仔,幼仔渐长,离去育儿囊,尚负于母背上,产于澳大利亚。——译者注
(665) 即Phascolomys wombat,属啮齿有袋类。体肥大,长二三尺,四肢粗短而强壮,皆有五趾。后肢除第二趾皆有长曲之爪,以爪掘穴而居。食木根及草等,故其齿列颇似啮齿类。——译者注
(666) 《动物和植物在家养下的变异》,第2卷,1868年,102,103页。
(667) 欧文编,《亨特的论文及其观察材料》,第1卷,1861年,194页。
(668) 贝克爵士,《阿比尼西亚的尼罗河支流》,1867年。
(669) 关于麝鼠,奥杜邦和贝奇曼,《北美的四足兽》(The Quadrupeds of N.America),1846年,109页。
(670) 《关于四足兽的新种》,1778年,7页。我所谓的狍(roe)就是帕拉斯命名的Capreolus sibiricus subecaudatus。
(671) 产于西藏、青海以及蒙古等处,或谓即驴之原种,体高达四尺,毛色概灰带赤、或栗色,背部中脊有黑纹一条,体下白色。幼驴毛色黄中带赤,与栖处之砂质色相合。——译者注
(672) 《博物学者在尼加拉瓜》,249页。
(673) 参阅史密斯《南非动物学》及格雷博士的《诺斯雷动物园访问记》二书中的精美图版。
(674) 《威斯敏特评论》,1867年7月1日,5页。
(675) 《南非游记》,第2卷,1824年,315页。
(676) 格雷博士,《诺斯雷动物园访问记》,64页。布赖茨在谈到锡兰豚鹿(hog-deer)时说道,在换角季节它的白色斑点比普通猪鹿的白色斑点更为鲜明。
(677) 福尔克纳(Falconer)和考特雷(Cautley),《地质学会会报》(Proc.Geolog.Soc.),1843年;以及法克纳的《古生物学论文集》(Pal.Memoirs),第1卷,196页。
(678) 《动物和植物在家养下的变异》,第1卷,1868年,61—64页。
(679) 《动物学会会报》,1862年,164页。再参阅哈特曼(Hartmann)的文章,见《农业年鉴》(Ann.d.Landw.),第43册,222页。
(680) 我在伦敦动物园曾见过这种情况,在小圣伊莱尔和居维叶的《哺乳动物志》的彩色图版中也可看到许多这种情形。
(681) 贝茨,《亚马孙河流域的博物学家》,第2卷,1863年,310页。
(682) 我在动物学会的动物园中曾见到大部分上述猴类。关于眉线瘦猴的描述系引自马丁的《哺乳动物志》,1841年,460页;再参阅475,523页。
