上篇
正如必须预料的那样,试验进行缓慢。最初需要有耐心,但以后当几项试验同时进行时,情况就改善了。从春到秋,每天都日益焕发起我的兴趣,这样,为照料试验所必须付出的苦心操劳就大大地得到了补偿。此外,如果通过我的试验,我能在加快解决这些问题方面取得成功,我将感到加倍的愉快。
——孟德尔

孟德尔
给卡尔·耐格里的信 【1】
格雷戈·孟德尔(G. Mendel) 【2】
(1867年) 【3】
最尊敬的先生:
最诚挚地感谢您如此友善地寄给我这些出版物。《植物界中种的形成》、《关于推论的植物杂种》、《杂种形成的理论》、《植物种间的中间类型》及《根据种的中间型及种的区域对Hieracien的分类处理》诸文特别引人注目。根据当代科学对杂交种理论作出这一彻底的修正是最受欢迎的,再一次向您致以谢意。
关于阁下好意收下的那篇拙文,我想补充下述资料:所述试验是从1856年到1863年进行的。我知道,我取得的结果很难同我们当代的科学知识相容,而且在这种情况下发表一项如此孤立的试验有着双重危险性:对试验者以及主张进行这项试验的动机都是危险的。为此,我作了最大的努力用其他植物来验证在豌豆方面所得到的结果。1863年和1864年所做的一些杂交,使我相信难以找到适合于开展大量试验的植物,而且在不利的情况下时间消逝了,却没有得到我所需要的资料。我曾试图启发人们做一些对照试验,为此在自然科学家地区性学会会议上谈到了豌豆试验。如预期的那样,我遇到了分歧意见;但就我所知,却无人去重复此试验。去年,当要我把我的演讲在学会会议录上发表时,经过再次检查我在历年试验的记录而未发现有什么错误后,我同意予以公开发表。呈送给您的文章是上述讲话草稿未作修改的翻印本;对一个公开讲话作这样一个扼要的说明是必要的。
据闻阁下对我的试验持怀疑态度,这一点并不奇怪;在同样的情况下我也会有这样的想法。看来在您尊敬的信件中有两点非常重要,但没有答复。第一点涉及这样一个问题,当杂种Aa产生A植株,而这个A植株又继续只产生A植株时,是否可以作出结论:已经得到了类型的稳定性。
请允许我说明,作为一名试验工作者,我必须把类型的稳定性定义为在观察期间一个性状的保持力。这就是说我的提法,即一些杂种后代产生相同的类型只包括进行观察的那些世代;没有把它扩大到这些世代以外的情况。有两个世代全部试验都是用数目相当大的植株进行的。从第三代开始,由于试验地不足,必须限制植株数,因此,7项试验中的每一项试验,只能取样第二代的一部分植株(它们或产生相同类型的后代,或产生不同类型的后代),作进一步观察。观察扩大到四至六代。在产生相同后代的变种中,取一些植株观察四代。我必须进一步提及,有一个变种六代都产生相同类型后代的例子,尽管其亲本类型有4个性状是不同的。1859年,我从杂种第一代得到了一个可育性很好的后代,种子大、味道好。翌年,由于其后代保持了这些优良性状,并且整齐一致,该变种被栽种在我们的菜园里,直到1865年,每年都栽种许多植株。它的亲本植株为bcDg和BCdG:B代表胚乳黄色;C代表种皮灰棕色;D代表豆荚鼓起;G代表轴长;b代表胚乳绿色;c代表种皮白色;d代表豆荚皱缩;g代表轴短。上述杂交种为BcDG。
只有留种的植株方能确定胚乳的颜色,因为其他的豆荚在尚未成熟时就已经收获了。在这些植株中从来没有看到绿色胚乳,紫红色花朵(棕色种皮的一项标志)、皱缩豆荚或短轴。
这是我的一点经验。我不能判断这些发现是否能确定类型的稳定性;然而,我倾向于认为豌豆亲本性状在杂种后代中的分离是完全的,因而是永恒的。杂种后代带有这一个亲本性状或另一个亲本性状,或者带有两个亲本性状的杂种形式;我从未看到亲本性状之间的逐渐过渡或者从一个亲本性状渐渐接近于另一个亲本性状。发展过程简单说来是每一世代所出现的两个亲本性状都是分开的和没有改变的,没有看到一个亲本性状从另一个亲本性状遗传或取得些什么。例如,让我提到寄给您的1035~1088号口袋。所有的种子都来自棕色和白色种皮相结合的杂种第一代。在该杂种的棕色种子中,长出了几株种皮为纯白色、不夹带任何棕色的植株。我预期这些植株能保持像亲本植株那样的相同的性状稳定性。
我想扼要推敲的第二点包含下面这种说法:“你应当注意数字公式仅仅是试验性的,因为不能证明它们是合理的”。
我用单一性状所作的试验都得到相同的结果:杂种种子长出的植株,有一半还带有杂种性状(Aa),而另一半接受亲本性状A和a,其数量相等。这样,按平均计算,4株植株中有两株具有杂种性状Aa,一株具有亲本性状A,而另一株为亲本性状a。所以,对两个彼此不同的性状来说,2Aa+A+a或A+2Aa+a是经验的简单的展开级数。同样地,从经验的含义上看出,如果杂种结合了2个或3个不同的性状,其展开级数为2个或3个简单级的结合。在这一点上,我相信不能非难我离开了试验的范围。如若我把这种简单级数的结合扩大到两个亲本植株间任何一个差数上,我倒的确进入到合理的领域了。但看来这是可以允许的,因为我已经由早先的试验证实,不管哪两个不同性状的发育都是毫无差别地独立进行的。最后,关于我提到杂种胚珠和花粉细胞间的差异,也是以试验为依据的。看来这些试验和生殖细胞的类似试验是重要的,因为我相信试验结果将为豌豆方面所观察到的杂种发育提供解释。这些试验必须重复试验和验证。
非常抱歉,未能寄给阁下所需要的品种。如上所述,试验进行到1863年,那时候要结束试验,以便腾出土地和时间种植别的试验植物。所以不再能提供这些试验的种子。仅继续了一项开花期不同的试验,可提供该试验在1864年收获的种子,这是我最后一次搜集的种子。由于在下一年因豌豆象(Bruchus pisi)为害猖獗,而不得不放弃试验。在试验的早期年份,这种昆虫很少在植物中发现,1864年它造成了严重的损失,所损失的数字使翌年夏季第四批和第五批种子都所剩无几了。到最后几年,不得不在布隆(Brünn) 【4】 周围地区停止栽种豌豆。留下的种子可能仍然有用,其中有几个变种我估计保持着稳定性,它们来自由2个、3个和4个不同性状结合在一起的杂交种。全部种子都是从第一代成员,也就是从原始的杂种种子直接长成的植株上取得的。
对于遵照阁下的要求寄送这些种子给您做试验用,我感到迟疑不决,因为这同我本人的意愿并不完全一致。我担心这些种子中已有一部分丧失了生活力。此外,这些种子是在豌豆象已经为害猖獗的时候收获的,这就难以排除这种昆虫传递花粉的可能性;还有,我必须再一次说明,这些植物是预定作开花期差异的研究。其他的差异在收获时也进行过研究,但不像主要试验那样小心注意。我在一张单独的纸片上对口袋号码所添加的符号,是收获时用铅笔在种子口袋上记载每个植株情况的抄写。显性性状用A、B、C、D、E、F及G表示,有其双重含义。隐性性状以a、b、c、d、e、f及g表示;它们应当在第二代保持稳定,也就是说,从带有隐性性状植株的茎上所收获的种子,预期它会产生相同的植株(有关所研究的性状)。
请把种子袋号码同我的记录本上的号码比较一下,以便找出符号上是否有差错,每一口袋只装有一个单株的种子。
有些变种适合于作生殖细胞的试验,当年夏季就能取得试验结果。715、730、736、741、742、745、756及757各号口袋里的圆形黄色种子,以及另一方面,712、719、734、737、749和750号口袋里的角形绿色种子可用做这方面的试验。通过重复试验证明,如果绿色种子的植株被黄色种子的植株授精,那么所得种子的胚乳便失去绿色,取得黄色。种子的外形也同样如此。如果角形种子的植株被圆形或球形种子的植株授精,就产生圆形或球形种子。这样,由于通过外来花粉的授精而使种子的颜色和形状发生变化,就可能以此来识别授精花粉的组分。
试以B表示黄色胚乳、b表示绿色胚乳。
如果自花受精产生绿色和角形的种子,这类植株的花朵被外来花粉授精,而产生的种子仍然为绿色和角形,那么授体植株的花粉在这
两个性状方面是 ab
如果种子的形状改变了,花粉来自 Ab
如果种子的颜色改变了,花粉来自 aB
如果种子的形状和颜色都改变了,花粉来自 AB
以上列举的口袋包括从杂种ab+AB所产生的圆形和黄色、圆形和绿色、角形和黄色以及角形和绿色的种子。圆形和黄色的种子最适合于做试验。它们中间或许会产生AB、ABb、Aab和AaBb变种,因此,当绿色和角形种子长成的植株被上述圆形和黄色种子长成的植株的花粉授精,可能有4种情况:①ab+AB;②ab+ABb;③ab+AaB;④ab+AaBb。
假设杂种形成的许多种花粉细胞是可能不变的组合类型,如果这一假设是正确的话,植株的组成为:AB产生的花粉为AB;ABb产生的花粉为AB和Ab;AaB产生的花粉为AB和aB;AaBb产生的花粉为AB、Ab、aB和ab。
胚珠的受精情况为①胚珠ab同花粉AB;②胚珠ab同花粉AB和Ab;③胚珠ab同花粉AB和aB;④胚珠ab同花粉AB、Ab、aB和ab。
从上述受精可能得到下列变种:
①AaBb;②AaBb和Aab;③AaBb和aBb;④AaBb、Aab、aBb和ab。
如果各种花粉产生的数目相等,必然是:①全部种子为圆形和黄色;②一半是圆形和黄色、一半是圆形和绿色;③一半是圆形和黄色、一半是角形和黄色;④四分之一是圆形和黄色、四分之一是圆形和绿色、四分之一是角形和黄色、四分之一是角形和绿色。
此外,既然AB、ABb、AaB、AaBb之间的数字关系为1∶2∶2∶4,那么从圆形、黄色种子长成的每9株应当发现其平均数为AaBb4次;ABb和AaB各2次;以及AB1次。这样,情况④发生的机会常常是情况①的4倍;为②和③情况的两倍。
如果在另一方面,从上述圆形、黄色种子长成的植株,被绿色、角形植株的花粉所授精,而倘若胚珠的种类相同,比例相同,则结果必然与花粉方面的报道完全相同。
我本人没有进行这项试验,但我相信,根据相似的试验,人们能信赖所指出的结果。
在同样的方式下,可以对两个种子的性状中的任何一个性状分别进行单独试验,合适的做法是在同一株植株上分析全部圆形与角形种子,以及黄色与绿色种子。例如,如果一株绿色种子的植株被一株黄色种子的植株授精,由于黄色种子长成的植株是B和Bb变种,所得到的种子应该或是①全部为黄色;或②一半为黄色、一半为绿色。此外,由于B和Bb发生的比率为1∶2,第二种受精发生的机会常常是第一种受精的2倍。
有关其他性状,试验可以相同方式进行;但需待下一年才能得到结果……
正如必须预料的那样,试验进行缓慢。最初需要有耐心,但以后当几项试验同时进行时,情况就改善了。从春到秋,每天都日益焕发起我的兴趣,这样,为照料试验所必须付出的苦心操劳就大大地得到了补偿。此外,如果通过我的试验,我能在加快解决这些问题方面取得成功,我将倍加愉快。
最尊敬的先生,请接受我最诚挚的问候。
您忠实的 格雷戈·孟德尔
1867年4月18日,布隆
植物杂交的试验 【5】
格雷戈·孟德尔(G. Mendel)
(1866年)
引言
为了获得观赏植物新的颜色变异而进行人工授精的经验,引起了这里将要讨论的试验。每当相同物种之间发生受精,总是反复出现同样的杂种类型,这种惊人的规律性,促使我们进行更多的试验,其目的是为了探究杂种在其后代中的发育情况。
为了实现这个目的,许多细心的观察家,如克尔路特(Kölreuter)、盖尔特纳(Gärtner)、斯宾塞(Herbert Spencer)、Lecoq、Wichura等人,以其不知疲倦的、坚韧不拔的精神献出了他们部分生命。特别是盖特纳尔,在他的《植物界中杂种的产生》著作中记载了非常有价值的观察;而最近Wichura发表了关于柳杂种的一些深刻研究结果。到目前为止,还没有卓有成效地提出一个能普遍应用的控制杂种形成和发育的规律,这对任何一个熟悉这项工作的规模,并能懂得进行这类试验所必须面临困难的人来说,那是不足为奇的。只有当我们拥有多种植物的详尽试验结果,才能最终地解决这个问题。
谁全面研究一下这方面的工作,都会产生一种信念,即在所有这许许多多的试验中,没有一个试验就其规模和方法来说,能确定杂种后代出现的不同类型的数目,或者按照不同世代把这些类型进行可靠性归类,或者明确地查明它们在统计学上的关系。
要从事一项如此规模巨大的工作,的确需要一些勇气;但看来这是我们能最终解决这个问题的唯一正确的途径,这个问题的重要性对有机类型的进化历史是难以过分估计的。
现在提出的这篇论文记载了这样一个详细试验的结果。这个试验实际上局限于一个小的植物类群,而现在,经过8年研究之后,其全部主要工作都已结束。至于按照计划所进行的各项试验是否最好地达到了预期的目标,则留待读者作出公正的判断。
试验植物的选择
任何一项试验的价值和效用,决定于其材料是否适宜于它所使用的目的,因而摆在我们面前的情况,用哪一种植物进行试验和怎样进行这种试验,不能说是不重要的。
如果从一开始就希望避免一切可疑结果的危险,则必须尽可能仔细地挑选用于这种试验的植物类群。
试验植物必须是:①具有稳定的可以区分的性状;②这种植物的杂种在开花期必须能防止所有外来花粉的影响,或能容易地防止。
杂种及其子代在以后各代中其可育性应无明显的干扰。
如果在试验中发生外来花粉的偶然授精,而又没有识别出来,就会得出完全谬误的结论。如在许多杂种子代中碰到的某些类型可育性的降低或完全不育,会使试验十分困难或完全失效。为了发现杂种类型彼此间和对它们先代的关系,看来有必要做到这一点,即必须把每一个连续世代所产生的各批后代的全部成员都毫无例外地进行观察。
刚开始时,鉴于它们特殊的花器结构而把注意力特别集中到豆科植物。用这一科的几种植物进行了试验,结果发现豌豆属具备必要的合格条件。
豌豆属中有几个完全不同的类型,它们具有稳定、且肯定能很容易识别的性状,当它们的杂种彼此间杂交时,它们产生完全可育的后代。此外,不易发生外来花粉的干扰,因为受精器官紧密包在龙骨瓣中,且花药在花蕾里面开裂,因而在开花前柱头已布满花粉。这一情况特别重要。值得提到的另一些优点是这些植物在大田栽培和盆栽都很方便,还有它们的生长期比较短。人工授精当然是一项比较细致的手工程序,但几乎总是成功的。为进行人工授精,花蕾在完全发育前,就把它打开,去掉龙骨瓣,用镊子小心夹出每一个雄蕊,此后立即用外来花粉撒在柱头上。
总共从几个种子商人得到了34个或多或少有区别的豌豆品种,并进行了两年试种。有一个品种发现在许多全都相像的植株中,有几株明显不同。然而,下一年它们就没有变化,且与同一种子商那里得到的另一个品种完全相同;因而这些种子毫无疑问只是偶然的混杂。所有其他品种都产生完全稳定和相似的后代;无论如何,在两年试种期间没有看到什么重要的差异。在整个试验期间选用和栽培了其中的22个品种作受精用。它们都毫无例外地保持了稳定性。
难以可靠地对它们进行系统分类。如果我们采用物种的最严格的定义,根据这个定义,只有那些在恰好相同的情况下表现出性状精确相似的个体才算是同一个种,那么这些品种中没有两个可算是一个种。但根据专家们的意见,大多数属于豌豆(Pisum sativum)这一个种;而剩下的一些品种,有的认为应分类为豌豆的亚种,有的认为是独立的种,诸如P. quadratum、P. saccharatum和P. umbellatum,然而确定它们在分类系统中的位置,对于本试验目的来说是相当无关紧要的。一直发现,要在种和品种的杂种之间划分严格的界限,如同要在种和品种本身之间划分严格的界限一样,是不可能的。
试验的分组和布置
如果把两种在一个或几个性状上具有稳定差别的植物作杂交,许多试验证明,共同的性状不加改变地传给杂种及其后代;但另一方面,每对有区别的性状则在杂种中结合成一个新的性状,它在杂种后代中通常是有变异的。试验目的是对每对有区别的性状观察它们的变异,并推导出它们在连续世代中出现的规律。因此,把这个试验分成许多个别的试验,其数目相当于试验植物所出现的稳定差别的性状数字。
选作杂交的各种豌豆类型在以下方面有差异:茎长和茎色;叶片的大小和形状;花的位置、颜色和大小;花柄的长短;豆荚的颜色、形状和大小;种子的形状和大小;以及种皮和胚乳(子叶)的颜色。注意到有些性状难以作明确和肯定的划分,因为这种差别只是表现在程度上的不同,没有明确的划分标准。这一类性状不能用来作个别试验;试验只能针对在植物中表现清楚和明确的性状,最后,试验结果必须说明,就其总体而言,能否观察到它们在其杂种结合中表现为一种有规律的现象,和根据这些事实能否对那些在类型中不太重要的性状作出结论。
选作试验的性状同下述诸点有关:
1.成熟种子形状的差异
种子形状或圆形或略圆,如有凹陷,发生在表面,且总是很浅;或者种子形状不规则地带角和呈现皱缩(P. quadratum)。
2.种子胚乳颜色的差异 【6】
成熟种子的胚乳颜色或为淡黄、鲜黄和橙色,或多少带深绿色。种子颜色的差异是很容易看出的,因为它们的种皮是透明的。
3.种皮颜色的差异
种皮或为白色,而白花总是与这个性状相关;或灰色、灰褐色、皮革褐色、有或无堇紫色斑点,在这种情况下,旗瓣的颜色为堇紫色,翼瓣为紫色,而叶腋中的叶梗带红色。灰色的种皮在开水中变成深褐色。
4.成熟豆荚形状的差异
这些豆荚或是简单地膨胀鼓起,找不到缢缩,或者在种子之间深深缢缩,或多少有点皱缩(P. saccharatum)。
5.不成熟豆荚颜色的差异
它们或是从淡绿到深绿、或是嫩黄色,叶柄、叶脉、花萼都参与 【7】 这些颜色的差异。
6.花朵位置的差异
它们或长在轴上,这就是说花朵沿着主茎分布;它们或是顶生的,即在茎的顶端长成一簇花,并排列成几乎像一个假的伞形花序;在这种情况下,茎的上部切面多少要粗些(P. umbellatum)。
7.茎长度的差异
有几个类型茎的长度大不相同;但对一种类型来说,它是一个稳定的性状,以在相同土壤中生长的健康植株而言,这个性状只发生不重要的差异。
用这个性状进行试验时,为了能准确地辨认它们,总是用6~7英尺 【8】 的长轴类型同0.75~1.5英尺的短轴类型作杂交。
上述每两个可区分的性状用异花受精把它们结合起来。所作试验如下:
第一次试验,15株上作60次受精;
第二次试验,10株上作58次受精;
第三次试验,10株上作35次受精;
第四次试验,10株上作40次受精;
第五次试验,5株上作23次受精;
第六次试验,10株上作34次受精;
第七次试验,10株上作37次受精。
从同一品种的大量植株中,只挑选生活力最强的植株作受精用。瘦弱的植株总是造成不可靠的结果,因为即使在杂种第一代,甚至以后的世代,许多子代或完全不开花或只结少数低劣的种子。
此外,在所有试验中正反交是这样进行的,两个品种的每一个品种在这一组作为产生种子作受精,在另一组就用作为花粉植株。
植株栽种于花圃内,少数进行盆栽,借助于使用木棒、树枝及拴于其间的绳索,使它们保持自然直立状态。每项试验在开花期把一些盆栽植株挪到温室,作为露天栽培主要试验的对照以防昆虫的可能干扰。在光顾豌豆的昆虫中,豌豆象(Bruchus pisi)如大量出现,则可能对试验有害。已知这种豌豆象的雌性昆虫在花里面产卵,这样就把龙骨瓣打开了,在一朵花里面捉到的一个跗节标本,在透镜下清楚地看到有几粒花粉。还必须提到的一种情况可能造成外来花粉的进入。例如在某些罕见的情况下,一朵其他方面发育都很正常的花朵,在某些部分凋萎了,结果使受精器官部分地暴露在外,也曾观察到龙骨瓣发育残缺不全,致使柱头和花药一直有一部分暴露在外面。有时也发生花粉发育不完全。在这种情况下,雌蕊在开花期间就逐渐伸长,直至柱头尖顶出龙骨瓣。在菜豆属和山黧豆属的杂种中也看到过这种异常的形态。
然而,对豌豆属来说,外来花粉造成错误受孕的危险是很小的,且决不能打乱总的结果。对10000个以上的植株作了仔细的检查,发现其中只有极少数不容置疑的例子是错误受孕。由于在温室中从未看到过这类情况,据推想豌豆象和上述花器结构的变态很可能是造成错误受孕的原因。
F1 杂种的类型
过去用观赏植物所作的试验已证明,杂种照例并不恰恰是亲本种间的中间类型。一些更为明显的性状,例如有关叶的大小和形状,有几个部分覆盖短柔毛等等,的确几乎常常看到中间型;但是在另一些情况下,两亲性状之一占压倒优势,以致很难或完全不可能在杂种中探查出另一个亲本的性状。
豌豆杂种的情况确实如此。7个杂交的每一个杂交,杂种性状同一个亲本的性状如此相像,以致另一个亲本的性状或完全看不到,或不能肯定地探查出来。这种情况对于确定和分类杂种后代所出现的类型是十分重要的。本文以后谈到的性状,凡是在杂交时整个或几乎不变地传给后代,从而它本身就构成杂种的性状则称为显性,而在传递过程中潜伏起来的性状则称为隐性。之所以选用隐性一词来表达,是因为这个性状在杂种中隐退或完全消失,但尽管如此,这个性状却又在它们的后代中毫无改变地重新出现,这一点将在后面予以说明。
此外,全部试验证明,显性性状究竟是属于产生种子一方还是花粉亲本一方,这一点完全是不重要的;在两种情况下,杂种的类型都是一样的。盖尔特纳也强调指出这一有趣的事实,他说即使最有实践经验的专家也难以在一个杂种中断定两个亲本种的性状是来自于母本植株还是父本植株。
本试验所用的可以区分的性状,其显性性状如下:
(1)种子形状圆或略圆,带有或不带有浅的凹陷;
(2)黄色种子子叶;
(3)种皮灰色、灰褐色或皮革褐色,及与此相联系的堇紫色的花和叶腋的红点;
(4)豆荚形状的简单膨胀;
(5)豆荚在未成熟时呈绿色,及与此相联系的茎、叶脉和萼也呈同样的颜色;
(6)花朵沿着茎分布;
(7)高茎。
必须说明,这最后一个性状,杂种通常超过两个亲本茎秆中更高的那一个,这种情况可能只是因为茎高极不相同者杂交时,植株各部分表现生长茂盛的结果。例如,经多次试验,1英尺和6英尺茎高杂交者都毫无例外地产生茎高在6~7.5英尺范围之内的杂种。
种皮试验的杂种种子常常斑点较多,而有时斑点连接成蓝紫色小块。即使当一个亲本的性状不存在时 【9】 ,这种斑点也经常出现。
杂种的种子形状和子叶颜色在人工授精后不久就显示出纯粹是外来花粉的影响。因此,在试验的当年就可看到这些性状,而所有其他性状当然只能出现在下一年从杂交种子长出的植株中。
F2 由杂种育成的第一代
这一代,同显性性状一起也出现其特点,得以充分显示的隐性性状,它们的出现明确地表现为3∶1的平均比例,因而这一代每4个植株中有3个植株为显性性状、一株为隐性性状。对于试验研究的全部性状,毫无例外地都是这种情况。种子带角的皱缩形状、子叶的绿色、种皮和花朵的白色、豆荚的缩缢、未成熟豆荚、叶柄、花萼和叶脉的黄色、伞状似花序和侏儒型茎秆,都以上述比例数字出现,而没有什么重大的改变。在任何试验中都没有观察到过渡类型。
由于正反交产生的杂种是一样的,且在它们以后的发育中没有可以觉察的差异,因此,每一次试验可以把正反交的结果合起来计算。每一对可区分性状所得到的相对数字如下:
试验1 种子形状——在第二个试验年度从253个杂种得到了7324粒种子。其中5474粒为圆形或略圆形、1850粒为带角皱缩形。据此推算出2.96∶1的比例。
试验2 子叶色——从258株得到8023粒种子,其中6022粒黄色、2001粒为绿色;因而其比例为3.01∶l。
在这两个试验中,每个豆荚通常都产生两种种子。在平均含有6到9粒种子的发育良好的豆荚里面,常常所有的种子都是圆的(试验1)或都是黄的(试验2);另一方面,从来没有看到在一个豆荚里面有超过5个以上的皱缩的或绿色的种子。杂种的豆荚发育得早或晚,或从主轴到侧枝长出,这都没有带来什么差异。一些少数植株,在最初形成的豆荚里面只发育了几粒种子,且两个性状只有一个性状,但以后发育的豆荚仍然保持了正常的比例。
像各个豆荚一样,各个植株中的性状分配也有不同。试以两组试验中头10个个体来说明。
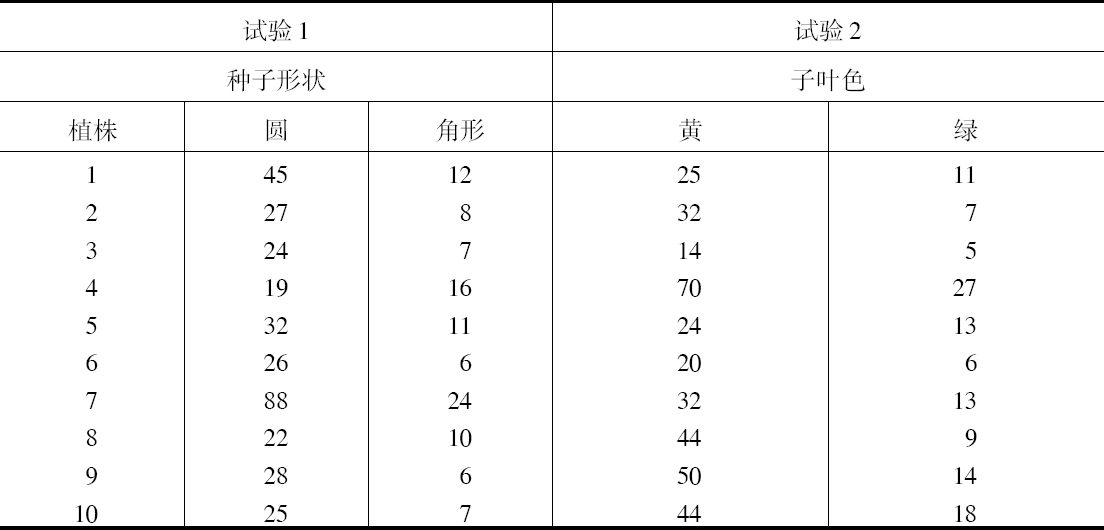
作为一个植株中两个种子性状截然不同的分配情况,试验1看到一个例子为43粒圆的、只有2粒带角的;而另一个例子为14粒圆的、15粒带角的。试验2有一例为32粒黄的和只有1粒绿的;但也有另一例为20粒黄的和19粒绿的。
这两个试验对确定平均比例是重要的,因为它说明,当试验植株数目较少时,可发生相当大的变动。还有,在计算种子时,特别是试验2,需要格外仔细,因为许多植株有些种子的子叶,其绿色不是那么明显,而初看起来容易被忽略掉。这种绿色的部分消失,其原因同植株的杂种性状无关,因为亲本品种也是如此。这种(漂白)特性只限于个体,而并不传给后代。在生长旺盛的植株中常常会看到这种情况。种子在发育期间由于昆虫的危害,常导致种子形状和颜色发生改变,但稍有挑选的实践经验后就很容易避免这种误差。豆荚必须保留在植株上直到它们熟透并发干,因为只有在那时候才充分显示出种子的形状和颜色,提及这一点几乎是多余的。
试验3 种皮色——929株中有705株产生紫红色花和灰褐色种皮,224株为白花和白色种皮,其比例为3.15∶1。
试验4 豆荚形状——在1181株中有882株豆荚为简单膨大、299株带缩缢。所得比例为2.95∶1。
试验5 豆荚未成熟时的颜色——供试植株580株,其中428株得绿色豆荚、152株为黄色。因此,其比例为2.82∶1。
试验6 花的位置——858例中,651株为轴生花序、207株为顶生。比例为3.14∶1。
试验7 茎的高度——1064株中,787例为高茎、277例矮茎。因此其互相比例为2.84∶1。这个试验要小心地把矮小植株挖出移栽到一个专用的试验圃中。这一预防措施是必要的,否则这些矮株会被其相邻的高株覆盖而死亡。即使在其幼龄状态,根据它们生长紧凑和密集的深绿色叶簇 【10】 ,很容易把它们挑选出来。
如现在把整个试验结果合在一起,就发现其显性和隐性性状数字之间的平均比例为2.98∶1或3∶1。
显性性状在此有一双重含义,即它既是一个亲本的性状,也是一个杂种性状。要知道在每一个个别情况下,它是这两种含义中的哪一种,只有在以后的世代中才能确定。作为一个亲本的性状,它必须毫无变化地传给所有的后代;另一方面,作为一个杂种性状,它必须保持如在第一代(F2 )中那样相同的行为。
F3 由杂种育成的第二代
凡是在第一代(F2 )表现隐性性状的类型,在第二代(F3 )这些性状就不再有变异;它们在后代中保持稳定。
(由杂种育成的)第一代中具有显性性状的类型则是另一种情景。它们中间有三分之二产生的后代,其显性和隐性性状之比为3∶1,从而表现出同杂种类型完全相同的比例,而只有1/3保持稳定的显性性状。
各试验的结果如下:
试验1 由第一代圆粒种子长出的565株中,193株只结圆粒种子,因而保持了这个性状的稳定性;然而,有372株产生圆粒和皱缩两种种子,比例为3∶1。所以杂种的数目与稳定的相比,为1.93∶1。
试验2 由第一代具黄色子叶种子长成的519植株中,166株只产生黄色子叶的种子、而353株产生黄色子叶和绿色子叶的种子,其比例为3∶1。因此,其结果是杂种和稳定类型划分的比例为2.13∶1。
下述试验的每一项个别试验选用了在第一代表现显性性状的100个植株,并且为了确定其含义,每株栽种10粒种子。
试验3 36株后代只产生灰褐色种皮;而64株后代中,有的灰褐色种皮,有的白色种皮。
试验4 29株后代只有简单膨大的豆荚;另一方面,71株后代中,有的膨大、有的缩缢。
试验5 40株后代只有绿色豆荚;60株后代中有的绿色、有的黄色。
试验6 33株后代只有轴生花朵;另一方面,67株后代中有的轴生、有的顶生。
试验7 28株后代遗传了长茎;72株后代中有的高茎、有的矮茎。
这些实验的每次试验中,都有一定数目的植株,其显性性状是稳定的。对于确定分离成具有持久稳定性状类型的比例,头两项试验尤为重要,因为这些试验可以比较大量植株。把1.93∶1和2.13∶1的比例合起来几乎正好是平均2∶1的比例。第六个试验结果十分相符;其余的试验比例多少有些变动,鉴于100株试验植株为数较少,这一点是可以预料到的。差距最大的试验5重复了一次,于是替代60和40的比例,结果是65和35的比例。因此,看来可以肯定地确定,平均比例为2∶1。从而得以证实,在第一代具有显性性状的那些类型,有2/3具有杂种性状,而1/3显性性状是稳定的。
因此,在第一代中,显性和隐性性状分配的3∶1比例,如果按照显性性状的意义可以把它分成杂种性状或亲本性状的话,这个比例在所有试验中都可以分解为2∶1∶1。由于第一代(F2 )的成员是从杂种(F1 )的种子直接产生的。现已清楚,杂种产生的种子具有两个可以区分性状中的这一个性状或另一个性状,这些种子有一半重新产生杂种,而另一半则产生稳定的植株,并分别获得显性或隐性性状,数目相等。
杂种育成的以后各代
杂种后代在第一和第二代中发育和分离的比例,大概对所有以后的后代都是适用的。试验1和2已进行了6代;试验3和4进行了5代;试验4、5、6进行了4代,这些试验从第三代起用少量植株继续试验,而没有看到与这一规律相偏离的情形,杂种后代在每一代都以2∶1∶1的比例分离成杂种和稳定的类型。
试以A代表两个稳定性状之一的显性性状,a代表隐性性状,并以Aa代表两者相结合的杂种类型,其公式为:
A+2Aa+a
此公式表示两个可区分性状在杂种后代系列中的各项。
盖尔特纳、克尔路特等人观察到杂种有回复到亲本类型的倾向,上述试验也证实了这一点。看来,从一次受精所产生的杂种数目,同变成稳定类型的数目相比较,在它们逐代相传的后代中总是不断地减少,但它们却不能完全消失。如假设所有世代中的全部植株能育性平均相等,此外,如果每个杂种所产生的种子有一半再次产生杂种,而另一半则以相等的比例产生两个性状稳定的类型,从以下综合结果可看出每一代子代的数目比例,这里A和a再一次指两个亲本的性状而Aa指杂种形式。为简便起见,假设每一代每1个植株只提供4粒种子。
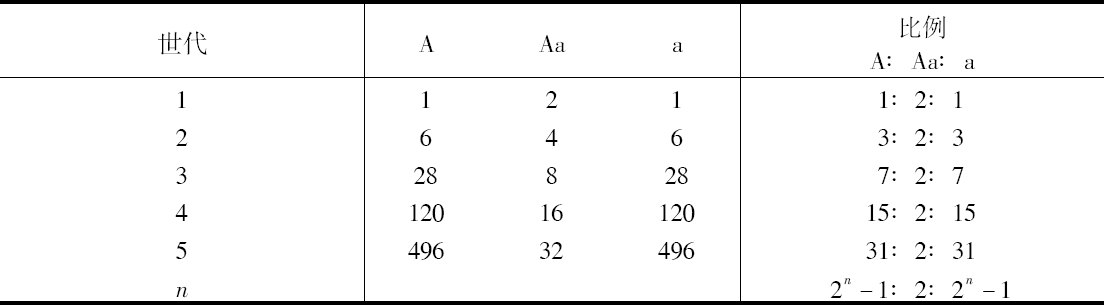
例如,在第十代,2n -1=1023。因此,这一代每产生2048个植株,有1023株具有稳定的显性性状、1023株具有隐性性状、只有两株是杂种。
同时具有几个可区分性状的杂种后代
上述试验中所用植物只有一个主要性状是不同的。第二项工作在于确定当通过杂交把几个不同的性状结合在杂种的时候,是否也能把所发现的性状的发育规律,应用于每对可区分的性状。关于在这种情况下的杂种形式,试验自始至终证明,这种杂种总是更接近于两个亲本植株中具有更多显性性状的那个亲本。例如,当母本植株的性状为短茎、顶生白花、简单膨大的豆荚;而父本植株为高茎,沿着茎着生的紫红色花和缩缢的豆荚时,杂种只有豆荚的形状像母本,其余的性状都同父本相像。倘若两个亲本类型中只有一个亲本具有显性性状,则杂种简直或根本不能与它区分。
用相当数目的植株作了两次试验。第一次试验,亲本植株在种子形状和子叶颜色方面有区别;第二项试验,则在种子形状、子叶颜色和种皮颜色上有区别。用种子性状进行的试验可以最简单和最肯定的方式得到试验结果。
为有助于研究这些试验资料,用A、B、C代表母本植株的不同性状,父本植株的性状以a、b、c表示,性状的杂种形式则以Aa、Bb和Cc表示。
试验1
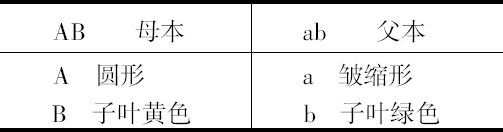
受精种子像母本,看起来是圆和黄色。从它们长成的植株产生4种种子,这些不同的种子常常出现在一个豆荚里面。总计,15株结了556粒种子,其中有:
315粒圆形和黄色;101粒皱缩和黄色;108粒圆形和绿色;32粒皱缩和绿色。
下一年把它们全部种下。圆形黄色种子中有11粒没有长成植株、有3株没有结种子。其余为:
38株圆形黄色的种子 AB
65株圆形黄色和绿色的种子 ABb
60株圆形黄色和皱缩黄色的种子 AaB
138株圆形黄色和绿色,皱缩黄色和绿色种子 AaBb
从皱缩黄色种子长成的96株植株,所结种子的情况如下:
28株只有皱缩黄色种子 aB
68株皱缩黄色和绿色种子 aBb
从108粒圆形绿色种子长出的102植株中,所结种子情况为:
35株只有圆形绿色种子 Ab
67株圆形和皱缩绿色种子 Aab
皱缩绿色种子长出30个植株,它们所结的种子性状都一样,保持了稳定的ab。
因而,看来杂种后代有9种不同的形式,其中有些数目很不相等。当我们把它们合在一起并加以整理后发现:
| 38株为AB符号; | 65株为ABb符号; |
| 35株为Ab符号; | 68株为aBb符号: |
| 28株为aB符号; | 60株为AaB符号; |
| 30株为ab符号; | 67株为Aab符号; |
| 138株为AaBb符号。 |
全部类型可以分为3个主要的不同组。第一组包括符号AB、Ab、aB和ab的植株,它们只具有稳定的性状并在下一代中不再有变异。这些类型每一个都代表平均出现33次。第二组包括符号ABb、aBb、AaB、Aab的植株,这些植株一个性状是稳定的,另一个性状是杂种的,并在下一代中只有杂种性状有变化。每一种植株平均各出现65次。AaBb类型发生138次,它的两个性状都是杂种形式,其行为同产生它们的杂种完全相同。
如把这三组类型出现的数字作一比较,1、2、4的比例就非常清楚。33、65、138的数字非常接近于33、66、132的比例数字。
因此,发育系列包括9组,其中有4组仅出现一次,且两个性状都是稳定的;AB、ab类型像它们的亲本类型;其他两种为A、a、B、b结合性状之间的组合,这些组合也可能是稳定的。有4组经常出现两次,一个性状是稳定的,另一个是杂种的。有一组出现4次,且两个性状都是杂种的。因此,杂种后代如果结合了两种可区分的性状,可用以下公式表示:
AB+Ab+aB+ab+2ABb+2aBb+2AaB+2Aab+4AaBb
这个公式无可争辩地是一个组合系列,其中把A和a的性状、B和b的性状这两个公式结合在一起。把这两个公式结合起来我们就得到系列中所有各组的全部数字:
A+2Aa+a
B+2Bb+b
试验2

本试验严格按照上述试验的相同方式进行。在全部试验中要求付出最多的时间并与困难周旋。从24个杂种总共得到了687粒种子,这些种子或为带斑点的灰褐色或灰绿色、圆或皱缩。下年有639株结实,进一步研究说明了它们中间有:
| 8株ABC | 22株ABCc | 45株ABbCc |
| 14株ABc | 17株AbCc | 36株aBbCc |
| 9株AbC | 25株aBCc | 38株AaBCc |
| 11株Abc | 20株abCc | 40株AabCc |
| 8株aBC | 15株ABbC | 49株AaBbC |
| 10株aBc | 18株ABbc | 48株AaBbc |
| 10株abC | 19株aBbC | |
| 7株abc | 24株aBbc | |
| 14株AaBC | 78株AaBbCc | |
| 18株AaBc | ||
| 20株AabC | ||
| 16株Aabc |
整个式子包括27项。其中8项所有性状都是稳定的,每项平均出现10次;12项有两个性状是稳定的,第三个性状是杂种的,每项平均出现19次;6项有一个性状是稳定的,另两个性状是杂种的,每项平均出现43次;有一个类型出现78次且所有性状都是杂种的。10、19、43、78的比例是如此接近于10、20、40、80或1、2、4、8的比例,致使后者比例可毫无疑问地代表它真正的数值。
这样,当原始亲本有3个性状不同时,杂种的发展可按下述式子进行:
ABC+ABc+AbC+Abc+aBC+aBc+abC+abc+2ABCc
+2AbCc+2aBCc+2abCc+2ABbC+2ABbc+2aBbC
+2aBbc+2AaBC+2AaBc+2AabC+2Aabc+4ABbCc
+4aBbCc+4AaBCc+4AabCc+4AaBbC+4AaBbc+8AaBbCc
这也是一个组合系列,在这个系列中把A和a、B和b、C和c的式子结合在一起,其公式为:
A+2Aa+a
B+2Bb+b
C+2Cc+c
这个式子可得出系列的全部类别。其中产生的稳定组合同性状A、B、C、a、b、c之间各种可能的组合都是符合的;这中间有两类ABC和abc同两个原有的亲本原种相似。
此外,用少数试验植株作了进一步的试验,把剩下的其余性状两个一起,3个一起地结合成杂种,全都得到了大致相同的结果。因此,毫无疑问,对试验所包括的全部性状来说,可应用的原理就是由几个主要不同的性状结合成的杂种后代,表现在组合系列的各项,这里面把每对可区分的性状都合并到一起。同时证明,每对不同性状在杂种结合中的关系,同两个最初的亲本原种在其他方面的差异是独立无关的。
设以n代表两个原种的可区分性状的数目,3n 就得出组合系列的项数,4n 为属于这个系列的个体数,而2n 则为保持稳定的组合数。因而,假如原种有4个性状不同,这系列就具有34 =81个类别,44 =256个个体和24 =16个稳定的类型;或者,换一种说法,每256个杂种后代,有81个不同组合,其中16个是稳定的。
在豌豆中,用重复杂交的方法实际上已经得到了上述7个可区分性状可能结合的全部稳定的组合。其数字是27 =128。由此得到证明,按照(数学的)组合定律,可能推算的所有组合,通过反复人工授精,可以得到在一群植物的几个品种中出现的稳定性状。
有关杂种的开花期,试验尚未结束。但已经可以说,杂种开花时间几乎正好在母本和父本之间,而且,杂种在这个性状方面的结构,大体上是遵循在其他性状方面所确定的规律。选用这类试验的类型,彼此间在开花中期方面的差别,至少需要20天,而且,播种时所有播种深度必须一致,以使它们能同时发芽。还有,在整个开花期,必须考虑到更重要的温度变化和由此引起花期的部分提早或推迟。显然,这个试验面临着许多需要克服的困难,并需要巨大的注意力。
假如我们打算用一种简单的方式来整理所得到的结果,我们发现那些在试验材料中易于肯定地识别的可区分性状,在它们的杂种相互关系中,全部行为都精确相似。每一对可区分性状的杂种后代,有一半仍然是杂种,而另一半则是稳定的,其中母本和父本性状的比例相等。假如通过异花受精把几个可区分的性状结合在一个杂种里面,所产生的后代就得到一个组合系列的各项,这个组合系列是把每一对可区分性状的组合系列合到一起。
用作试验的全部性状,其表现行为整齐一致,这使我们得以充分证实接受以下的原则是正确的,即有些其他性状在植物中不是表现得那么界限分明,因而不能在各个试验中把它们包括进去,但也存在着类似的关系。用长度不同的花梗进行的一次试验,总的来说,结果是相当令人满意的,虽然在类型的区分和系列的排列方面,它不可能做到像准确试验所必不可缺的那样肯定。
杂种的生殖细胞
上述试验结果导致进一步试验,试验结果看来适合于对杂种的卵和花粉细胞的成分作出一些结论。豌豆方面所提供的一条重要线索是在杂种后代中间出现的稳定类型。同样地,在相连性状的全部组合中也发生这种情况。单凭经验来说,我们发现每种情况下皆可证实,只有当卵细胞和受精花粉具有相同性状时,才能产生稳定的后代,因而卵细胞和授精花粉这二者都具有创造十分相似的个体的材料,其情况犹如纯种的正常受精一样。因此我们必须肯定地认为,当杂种植株产生稳定类型时,必然有恰恰相似的因素在起作用。既然一个植株,甚或一个植株的一朵花产生出各种稳定的类型,那么看来得出下面的结论是合乎逻辑的,即杂种的子房里形成许多种卵细胞,而花药中形成许多种花粉细胞,其情况如同有可能的稳定组合类型一般,并且这些卵和花粉细胞的内部组成同各个类型的卵和花粉细胞是一致的。
事实上,有可能从理论上证明,这个假设完全足以说明各代杂种的发育情况,只要我们同时假定杂种所形成的各种卵和花粉细胞,其平均数目相等。
为了用试验证明这些假设,设计了以下试验。通过受精把两种在种子形状和子叶颜色方面不同而稳定的类型结合起来。
试以A、B、a、b再次代表可区分的性状,我们就有:

把这种人工授精的种子同两个亲本原种各取几粒种子一起种下,并取样生长最健壮的植株作正反交。受精情况如下:①用AB花粉给杂种受精;②用ab花粉给杂种受精;③用杂种的花粉给AB受精;④用杂种的花粉给ab受精。
这四项试验,每项试验3个植株上的全部花朵都受了精。如上述理论是正确的话,杂种必定产生AB、Ab、aB、ab类型的卵和花粉细胞,其结合情况如下:①卵细胞AB、Ab、aB、ab和花粉细胞AB;②卵细胞AB、Ab、aB、ab和花粉细胞ab;③卵细胞AB和花粉细胞AB、Ab、aB、ab;④卵细胞ab和花粉细胞AB、Ab、aB、ab。
以上各项试验只能产生以下类型:①AB、ABb、AaB、AaBb;②AaBb、Aab、aBb、ab;③AB、ABb、AaB、AaBb;④AaBb、Aab、aBb、ab.
此外,如果所产生的杂种的卵和花粉细胞的几种类型,其数目平均相等,那么在每项试验的上述4种组合应当彼此间比例相同。但不能指望这种数字关系完全相符,这是因为每一次受精,即使在正常情况下总有一些卵细胞不能发育,或以后死亡,而且有许多即使发育良好的种子播种时却不能发芽。上述假设还受到以下方面的限制,即尽管它要求产生同等数目的各种卵和花粉细胞,但它并不要求每一个杂种在数学上准确无误。
第一和第二项试验的主要目的为证明杂种卵细胞的组成,而第三和第四项试验则是确定花粉细胞的组成,如上述证明所示,第一和第三项试验同第二和第四项试验应当刚好产生同样的组合,而且即使在下一年,在人工授精种子的形状和颜色方面,应当部分地看出这个结果。在第一和第三项试验,种子形状和颜色的显性性状A和B,在每个组合中都出现,且部分是稳定的,部分与隐性性状a和b成杂种结合,为此,显性性状必定把它们的特点印刻在全部种子上。因此,如果这个理论证明是正确的话,全部种子应当是圆和黄的。另一方面,在第二和第四项试验,一种组合在种子形状和颜色方面都是杂种,因而种子是圆和黄的;另一种组合种子形状是杂种,但颜色是稳定的隐性性状,所以种子是圆和绿的;第三种组合种子形状的隐性性状是稳定的,但颜色是杂种的,其结果种子是皱缩和黄的;第四种组合种子的两个隐性性状都是稳定的,因而种子是皱缩和绿的。这样,在这两项试验中都可以指望有4种种子,即圆和黄、圆和绿、皱和黄、皱和绿。
收获结果同预期情况完全相符。所得结果如下:
第一项试验,98粒都是圆、黄种子;
第二项试验,94粒都是圆、黄种子;
第三项试验,31粒圆和黄、26粒圆和绿、27粒皱和黄、26粒皱和绿的种子;
第四项试验,24粒圆和黄、25粒圆和绿、22粒皱和黄、26粒皱和黄的种子。
现不再对试验取得成功有任何怀疑;下一代必能提供最终的证据。从播下的种子中,第一项试验得90株,第三项试验得87株。所结种子情况如下:

第二和第四项试验中圆和黄的种子产生具有圆和皱、黄和绿色种子AaBb的植株;从圆绿种子中,产生具有圆和皱绿种子Aab的植株;皱黄种子产生皱黄和绿色种子aBb的植株;从皱绿种子长成的植株,只产生皱绿种子ab。
虽然这两项试验都有一些种子不能发芽,但并不影响上年取得的结果,因为每一种种子所产生的植株,就它们的种子来说,彼此相似,而与其他植株不同。因此,所得结果如下:

因此,在全部试验中,出现了所提出理论要求的所有类型,且数目几乎相等。
在另一项试验中,进一步试验了花色和茎长的性状,假若以上理论是正确的话,选择进行到试验第三年,每一个性状应该出现在全部植株的一半。试以A、B、a、b再次代表各种性状。
A 紫红色; B 长轴; a 白花; b 短轴。
类型Ab为ab所受精,产生杂种Aab。此外,aB也为ab受精,而产生杂种aBb。翌年,为进一步受精,Aab杂种用作母本,而杂种aBb用作父本:
母本 Aab, 可能的卵细胞 Ab、ab;
父本 aBb, 花粉细胞 aB、ab。
卵和花粉细胞间受精可能产生4种组合,即AaBb+aBb+Aab+ab
由此看出,按上述理论,在试验的第三年,全部植株情况如下:
半数应为紫红色花(Aa),第1、3组;
半数应为白花(a),第2、4组;
半数应为长轴(Bb),第1、2组;
半数应为短轴(b),第3、4组。
从第二年的45次受精产生了187粒种子,其中只有166粒种子在第三年长到开花期。其中各组出现的数目如下:

由此出现了:
85株紫红色花(Aa);81株白花(a);87株长茎(Bb);79株短茎(b)。
因此,所提出的理论在这个试验里同样令人满意地得到了证实。
在豆荚形状、豆荚颜色和花的位置等性状方面,也作了小规模的试验,所得结果完全相符。由可区分性状联合成全部可能的组合,都充分地表现出来,并且数目基本相等。
因而,通过试验证实了这样一种理论,即豌豆杂种所形成的卵和花粉细胞,在它们的组成方面,数目相等地代表着受精中性状联合而组成的全部稳定的类型。
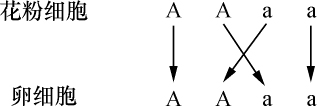
杂种后代中各种类型的差异以及观察到它们在数目上的相应比例,可以在以上提出的原理中找到充分的解释。每一对可区分性状的展开系列可提供这方面最简单的例子。这个系列用A+2Aa+a式子表示。其中A和a指具有稳定的可区分性状的类型,而Aa为这两者的杂种类型。它把4个个体分成3个不同的组别。产生这种结果时,A和a类型的花粉和卵细胞均等参与受精;因此每一类型发生两次,而形成4个个体。这样参与受精的是:
花粉细胞 A+A+a+a
卵细胞 A+A+a+a
因此,两种花粉中哪一种花粉同每一个个别的卵细胞相结合,纯粹是一种机会而已。然而,根据概率定律,按许多情况的平均来看,经常会发生的情况是每一种花粉类型A和a同每一个卵细胞类型A和a的结合往往机会等同,因此,两个花粉细胞A之一在受精中将同卵细胞A相遇,而另一个则同一卵细胞a相遇;同样地,一个花粉细胞a将同一个卵细胞A结合,而另一个与卵细胞a结合。
设以相结合的卵和花粉细胞的符号以分数的形式表示,把花粉细胞放在线上,卵细胞在线下,就可以弄清楚受精的结果。这样我们可写成:
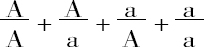
第一和第四项,卵和花粉细胞为相同种类,因而它们相结合的产物必然是稳定的,即A和a;另一方面,第二和第三项又产生原种的两个可区分性状的结合,因此,由这些受精所产生的类型与原种所产生的杂种类型相同。因而这就发生一种重复杂交。这说明了一个惊人的事实,即杂种除了产生两个亲本类型外,还能够产生同它本身相像的后代:
两者都得到同样的结合Aa,这在前面已经提到,花粉或卵细胞属于这两个性状中的哪一个性状,对受精的结果不会带来差异。因而我们可以写成:
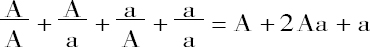
这个式子代表当两个可区分性状在杂种中结合时,其自花受精的平均结果。但是,个别的花朵和个别的个体,这一系列类型所产生的比例可能有明显的变动。除了在事实上只能把种子器官中发生的两种卵细胞的数目,看做是平均相等外,究竟两种花粉中哪一种花粉同每一个个别的卵细胞受精,却仍然是一种纯粹的机会。为此,个别的数值必然有所起伏,甚至有可能出现极端的例子,这在以上有关种子形状和子叶颜色的试验方面已经谈到过。数目的真正比例,只能通过从尽可能多的单个数值的总和中平均推算出来;数目越多,越能消除单纯的机会效应。
由两种可区分性状结合的杂种,其展开系列在16个个体中有9个不同的类型,即:
AB+Ab+aB+ab+2ABb+2aBb+2AaB+2Aah+4AaBb
在原种的可区分性状Aa和Bb之间,可能有4种稳定的组合,因而杂种就产生相应的4种卵和花粉细胞AB、Ab、aB、ab。每一种在受精中平均出现4次,因为此系列包括16个个体。而受精的参与者是:
花粉细胞
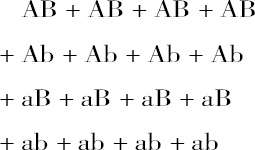
卵细胞
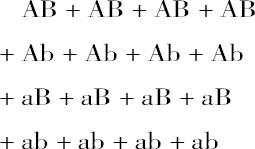
在受精过程中每一种花粉总是以平均相等的机会同每一种卵细胞结合,因此4个花粉细胞AB的每一个,都有一次机会同卵细胞的各种类型AB、Ab、aB、ab中间的一个相结合。其余的几种花粉细胞Ab、aB、ab也以完全相同的方式同所有其他的卵细胞相结合。因而,我们得到:
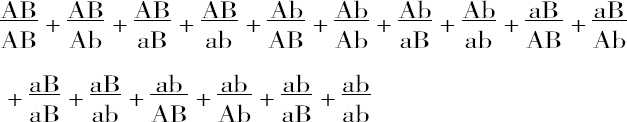
或
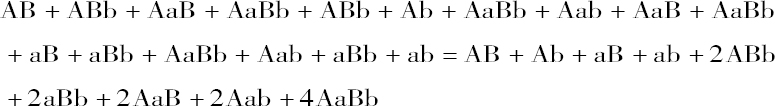
当3种可区分性状结合在杂种中时,其杂种的发展系列也按此完全相同的方式表示之。杂种形成8种不同的卵和花粉细胞ABC、ABc、AbC、Abc、aBC、aBc、abC、abc,而且每种花粉再次平均地使自己同每一种卵细胞结合一次。
因此,控制杂种发育的不同性状的组合规律,可以在所阐明的原理中找到根据和解释,即杂种所产生的卵细胞和花粉细胞,以相等数目代表着受精中性状结合所产生的全部稳定的类型。


谈家桢(1909—2008),我国现代遗传学奠基人,杰出的科学家和教育家。图为谈家桢在科研与教学时的照片。
注释
【1】 梁宏译自G. Mendel. Letters to Carl Nägeli. The birth of genetics, part Ⅱ. Genetics , 1950, 35(5): (In supplement, pp 3-5).
【2】 格雷戈·孟德尔(Johann Gregor Mendel,1822—1884),奥地利植物遗传学家,被誉为“现代遗传学之父”。他通过豌豆杂交实验,发现了遗传学的两个基本规律——分离规律和自由组合规律,开始了现代遗传学研究。
【3】 表示该文章首次发表时的年份。全书同解。
【4】 现为捷克的布尔诺。
【5】 梁宏译自G. Mendel. Experiments in plant hybridization. Classic Papers in Genetics , 1962: 1-20.
【6】 孟德尔以“胚乳”一词不太确切地指种子里面含有养分的子叶。
【7】 有一个种具有颜色极美的褐红色豆荚,它在成熟时变成堇紫色和蓝色。去年才开始用这个性状做试验。
【8】 1英尺=0.3048米。
【9】 这里指F1 植株产生的种子的种皮。
【10】 侏儒型或“Cupid”甜豌豆也是同样如此。
